Abstract
Most studies about the effects of N addition on soil microbial biomass evaluate soil microbial and physicochemical characteristics using single-test methods, and these studies have not been integrated and analyzed to comprehensively assess the impact of N fertilization on soil microbial biomass. Here, we conduct a meta-analysis to analyze the results of 86 studies characterizing how soil microbial biomass C (MBC), N (MBN), and P (MBP) pools respond to exogenous N addition across multiple land use types. We found that low N addition (5–50 kg/hm2) rates significantly affect soil microbial biomass, mainly by increasing MBC but also by decreasing MBP and significantly increasing MBC/MBP. N addition affects soil physicochemical properties, significantly reducing pH and significantly increasing the soil dissolved organic N and inorganic N content. Our analysis also revealed that the effects of N application vary across ecosystems. N addition significantly decreases MBP and total P in planted forests but does not significantly affect soil microbial biomass in grasslands. In farmland soil, N addition significantly increases total P, NH4+, NO3−, MBN, and MBP but significantly decreases pH. Although N addition can strongly influence soil microbial biomass, its effects are modulated by ecosystem type. The addition of N can negatively affect MBC, MBN, and MBP in natural forest ecosystems, thereby altering global ecosystem balance.
1. Introduction
N is an important limiting nutrient that constrains productivity and often regulates net C uptake rates in terrestrial ecosystems. Atmospheric deposition of reactive N has increased dramatically due to fossil fuel combustion and the widespread use of N fertilizers, with important impacts across ecosystems at the global scale [1,2,3]. N modulates biological and geochemical processes, and reactive N deposition affects the accumulation, decomposition, and persistence of organic matter in terrestrial ecosystems. These changes interfere with the biogeochemical cycling of C, N, and P in ways that influence ecosystem services [4,5]. Therefore, soil C, N, and P form important links in the global C and N cycles, and the decomposition, transformation, and accumulation of soil organic matter impacts plant growth and global C and N balances [6]. Global nutrient cycles are also impacted by microbial biomass C, N, and P dynamics, and the proportion of soil organic matter formed by each of these sub-pools can be altered by N addition.
The microbial biomass C (MBC) pool is composed of dead microbial biomass and accounts for 1–3% of total soil organic C (SOC). In addition to its utility as an overall indicator of the intensity of soil microbial activity, it regulates the decomposition and formation of organic C in soils [7,8]. MBC is also a primary driver of soil N mineralization and as such plays an important role in nutrient cycling and organic matter decomposition [9,10,11]. Because MBC forms a reservoir of unstable, plant-available nutrients [12], understanding the dynamics of this pool is essential for understanding soil C cycling, C balance, and biochemical properties [13,14]. Importantly, the size of the MBC pool and its fluxes are often used in evaluations of soil fertility [15,16,17], soil effective nutrient status, and biological activity. Moreover, MBC is a source of C contributing to soil humus, and the new humus formation that it promotes supports soil fertility and ecology [18]. Previous studies have demonstrated that the size of the MBC pool tends to increase proportionally to the amount of exogenous N applied, although these increases are not permanent; this relationship may result from the lower soil C/N ratio promoted by N application [19]. Other work has shown that N addition increases microbial biomass C and soluble organic C stocks and is correlated with total SOC content [20,21]. Some research also suggests that N deposition alters the MBC pool more in grasslands than in forests or croplands, which is generally consistent with patterns observed in larger SOC pools [22].
Microbial biomass N (MBN) is the main form of N sequestered in soil organic matter and is involved in nutrient cycling and energy conversion in soils [23,24]. Characteristics of the MBN pool influence exchanges of N between organic and inorganic forms, and variation in its lability and concentration is widely acknowledged as an essential regulator of internal soil N cycling [25,26,27]. Prior work has demonstrated that soil microbial biomass is influenced by many factors, such as forest type, seasonal temperature variation, rainfall, plant development, and organic matter accumulation driven by apoplankton. These variables drive the strong temporal and spatial variation observed in soil microbial biomass via their influence on its quantity and function [28]. Many indicators used to assess soil N pools are modulated by the quantity and activity of soil microorganisms, which are key players in soil N transformations [29]. More generally, MBN has been used as an early and sensitive indicator of the extent to which a soil has been disturbed, and the size of this pool in agroecosystems is influenced by many management practices [30]. Some work has shown that N addition can reduce MBN content, which may result from the inhibition of bacterial growth driven by deviations from neutral pH [31], thereby altering the soil’s bacterial diversity. Moreover, N addition has been shown to promote plant growth and plant N uptake, altering the dynamics of plant–microbe competition for nutrients and water. Under fertilization, microbial growth and reproduction are inhibited, decreasing MBN content [32].
Microbial biomass P (MBP) is an irreplaceable source of plant P and a key regulator of the soil P cycle [33], and exogenous N inputs modulate P retention by soil microbes [34]. The application of ammonium N reduces soil pH, thereby enhancing the adsorption of inorganic P by soil cations and reducing the amount of P available for microbial fixation [35]. Guo et al. showed that N addition caused soil acidification and inhibited microbial growth, weakening the ability of soil microorganisms to retain inorganic phosphates [36].
We hypothesized that N addition may affect environmental variables relevant to soil microbial communities, resulting in significant changes in soil MBC, MBN, and MBP. However, most studies assess soil microbial biomass and physicochemical properties using single-test methods; there is also a lack of integration and analysis of previous studies related to the impact of N addition on soil microbial biomass. Meta-analysis is an important statistical method that combines the results of multiple independent trials from similar studies to draw comprehensive conclusions. Here, we conducted a meta-analysis to elucidate the effects of N addition on soil microbial C, N, and P pools in different ecosystems. This work provides a scientific basis for future research on soil biogeochemical cycling across ecosystems and may be important for predicting how alterations to the global N cycle may induce ecological changes.
2. Materials and Methods
2.1. Literature Search and Screening
To investigate the overall effect of anthropogenic exogenous N addition on MBC, MBN, and MBP, we conducted a search using the databases China Knowledge (https://www.cnki.net, accessed on 20 May 2023) and Web of Science (https://webofscience.clarivate.cn/wos/woscc/basic-search) on 20 May 2023. We employed the following search terms: (“microbial biomass C” OR “MBC” OR “microbial biomass N” OR “MBN” OR “microbial biomass P” OR “MBP”) AND (“fertilization” OR “N fertilizer” OR “inorganic N” OR “urea” OR “N addition” OR “N deposition” OR “organic N” OR “N” OR “ammonium N” OR “nitrate N”). We selected articles published between 2000 and 2023 using the following criteria: (1) experimental treatments should compare the results of exogenous N application to a control group without N application; (2) each treatment group should have at least three replicates; (3) sample site data (e.g., geographic coordinates, soil type, laboratory vs. field experiment) should be available; (4) microbial C, N, and P content data should be available.
2.2. Compilation
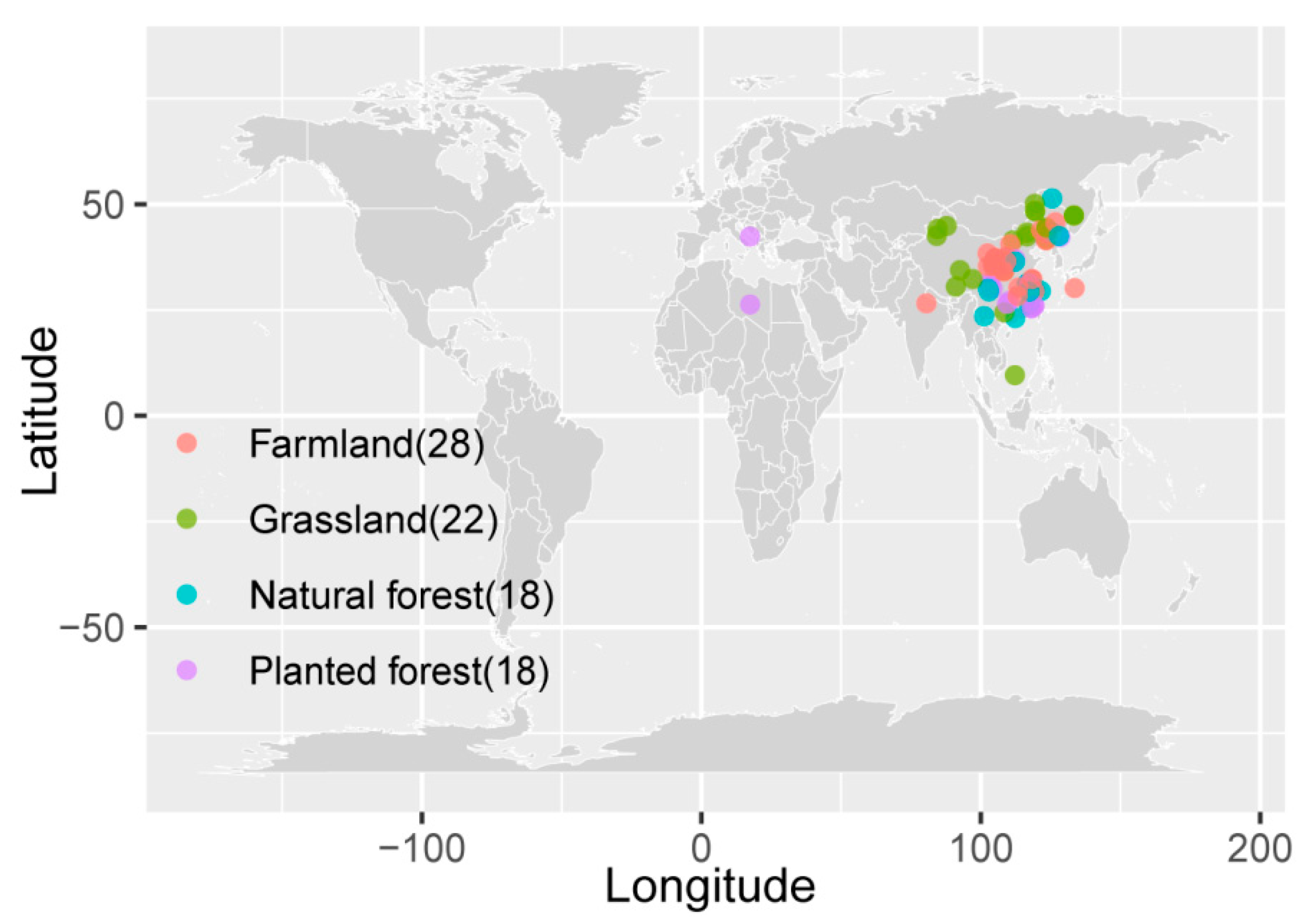
Variables compiled from the literature search included the following: ecosystem type; altitude; mean annual temperature (MAT) and precipitation (MAP); MBC, MBN, and MBP; and MBC/MBN, MBC/MBP, and MBN/MBP. Graphical information was digitized using Getdata software (v2.25, Russia). Eighty-six studies published between 1 January 2000 and 1 January 2023 were selected for a total of 658 datasets. Both indoor cultures and field trials are represented in this collection. The geographic and frequency distribution of the selected studies is presented in Figure 1 and Supplementary Materials.

Figure 1.
Geographic distribution of selected studies.
2.3. Data Analysis
Effect values (Ei) were calculated for each group using standardized units for each indicator. Groups not receiving N addition were classified as part of the control. The response ratio (RR) and its logarithm (lnRR) were used as the Ei to assess how different indicators of MBC, MBN, and MBP responded to N addition. The following equation was used: Ei = lnRR = ln (Xe/Xc), where Xe is the mean value of the response variable in the test group and Xc is the mean value of the response variable in the control group without added N.
Variance (v) was calculated as v = , where ne and nc are the sample sizes of the variable under N addition and CK treatments and Se and Sc are the standard deviations of the variable under N addition and CK treatments. The mean response ratio (RR++) was calculated from the individual pairwise comparison between N and CK treatments: , , where m is the group number and k is the number of comparisons in the group. We used the R package “metafor” to calculate the 95% confidence intervals. If the 95% confidence interval overlapped with 0, the difference between the test and control group was considered non-significant; if the 95% confidence interval did not overlap with 0, the difference between the two groups was considered significant. Data analysis was conducted using R (v4.2.3), and the package plotrix was used to generate graphs. Bootstrap statistical analysis was conducted using SPSS 22.0 (version 25.0, IBM, Armonk, NY, USA).
3. Results
3.1. Effect of N Addition on Soil Microbial Biomass
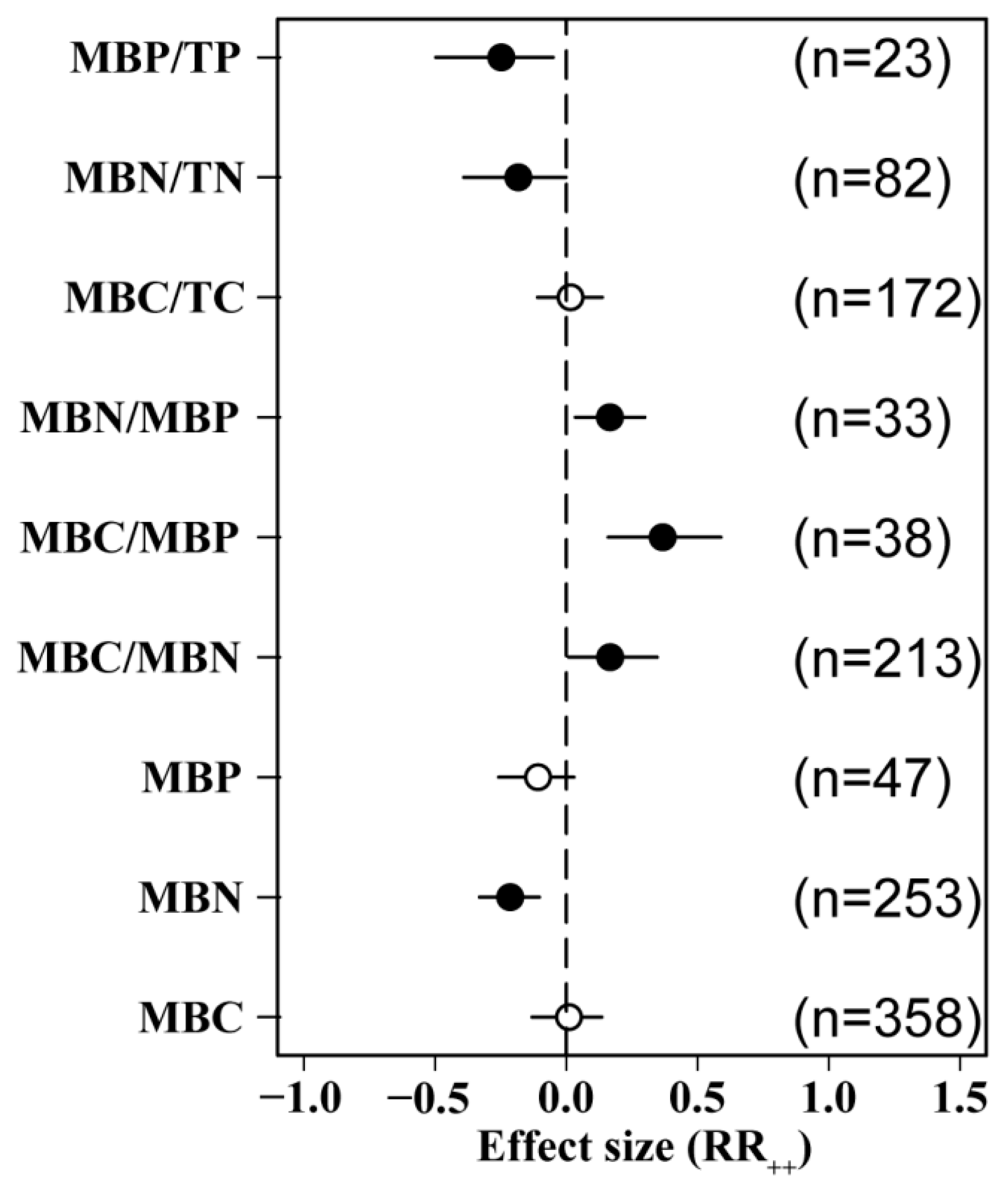
N addition significantly reduced MBN did not have a significant effect on MBC or MBP. N addition significantly increased MBC/MBN, MBC/MBP, and MBN/MBP ratios by 36.7%, 16.7%, and 16.6%, respectively, but the difference between the indicators was not significant. Our analysis also revealed that N positively influenced MBC/TC and negatively influenced MBN/TN and MBP/TP (Figure 2).
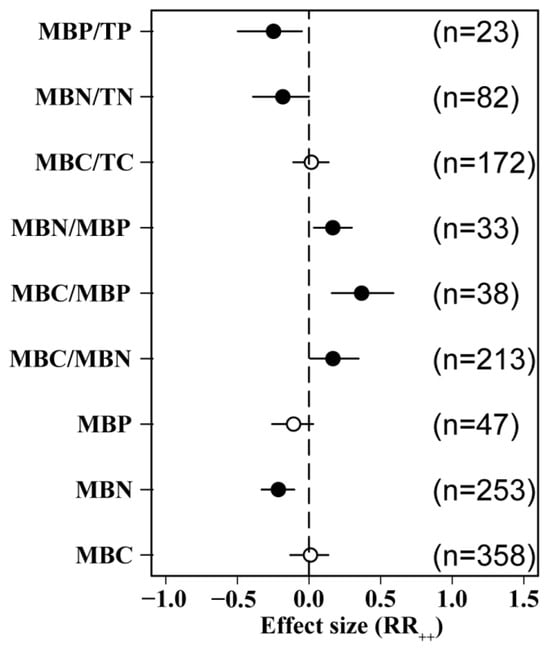
Figure 2.
Effects of N addition on soil microbial biomass. MBC: soil microbial biomass C, MBN: soil microbial biomass N, MBP: soil microbial biomass P, TC: soil total organic C, TN: soil total N, TP: soil total P. Number of samples is represented by n. Closed and open symbols indicate significant and non-significant effects, respectively.
3.2. Effects of N Addition Rates on Microbial Diversity and Plant Growth
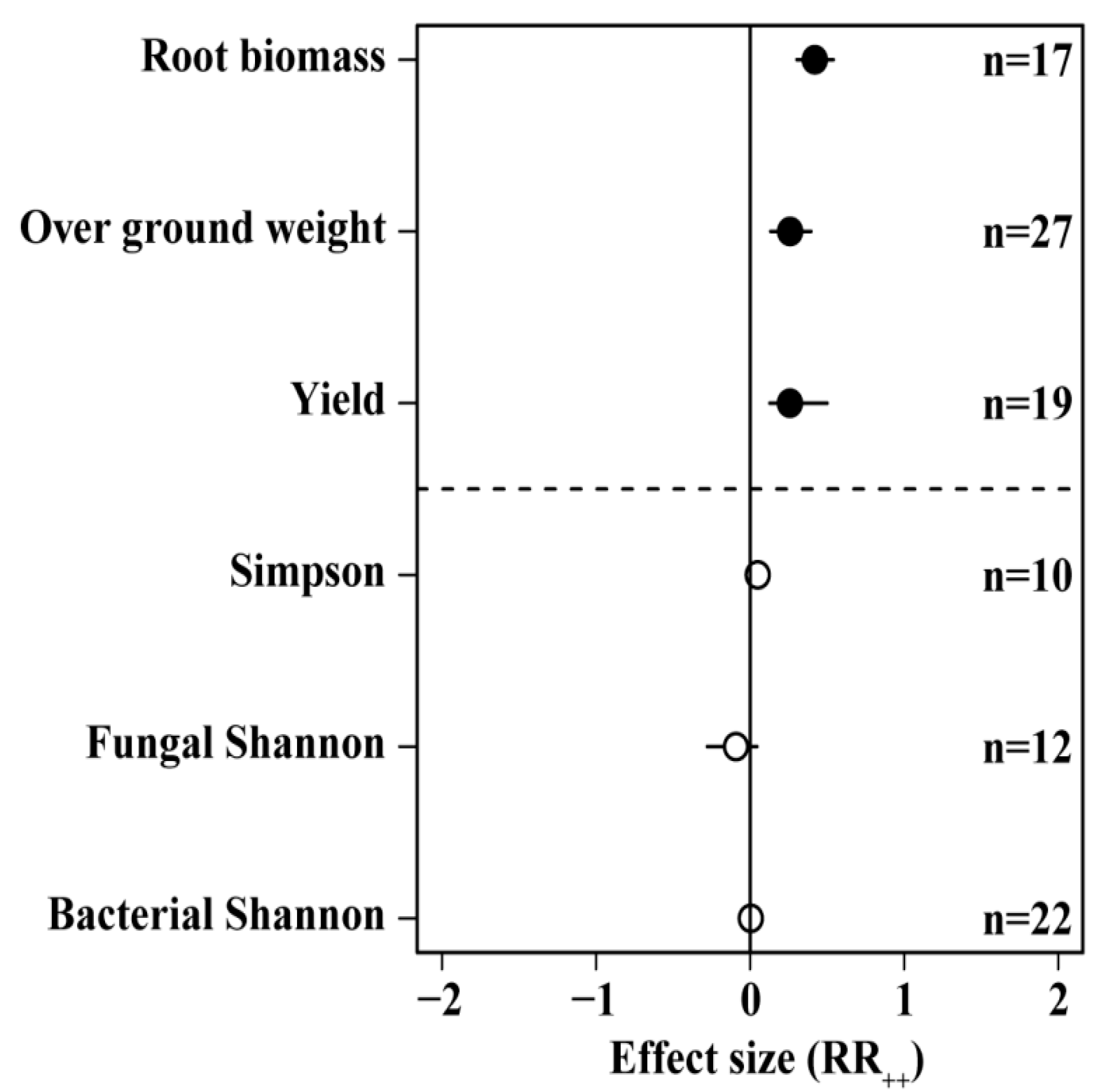
We found that N addition did not have a significant positive effect on soil microbial diversity, but it did have significant positive effects on plant yield (25.8%), above-ground biomass (25.9%), and root biomass (42%) (Figure 3).
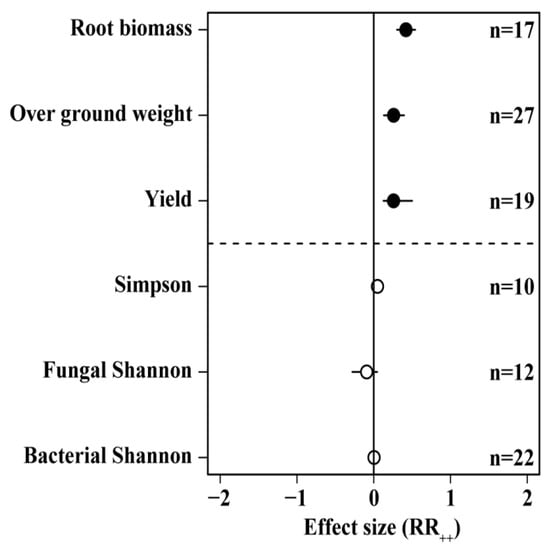
Figure 3.
Effects of different N application rates on microbial diversity and plant growth. Number of samples is represented by n. Closed and open symbols indicate significant and non-significant effects, respectively.
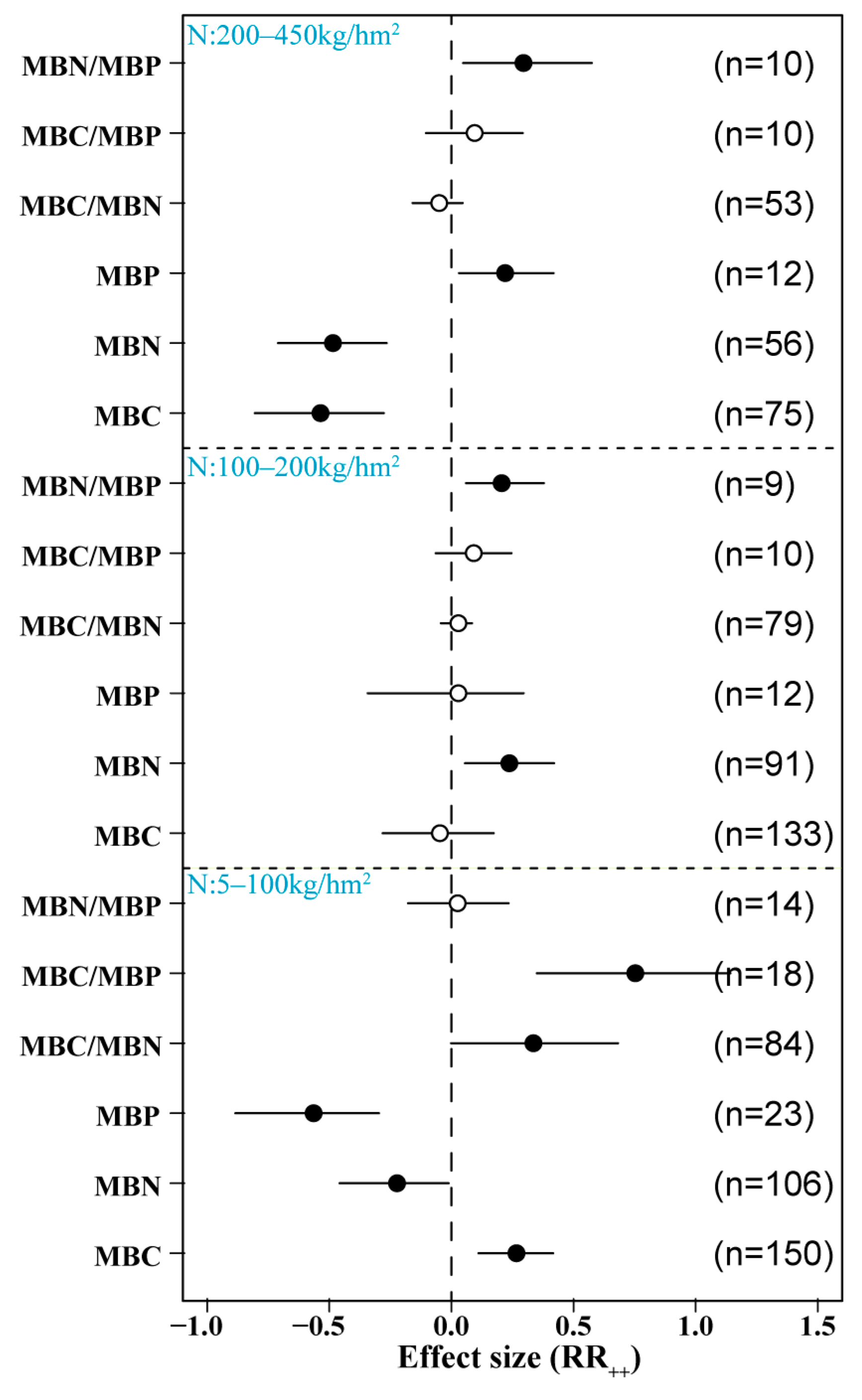
3.3. Effects of Different N Application Rates on Soil Microbial Biomass
We found that N addition had a significant effect on soil microbial C, N, and P. Low N addition (5–50 kg/hm2) significantly increased soil MBC content and MBC/MBP but significantly reduced MBP. It had no significant effect on MBN, MBC/MBN, or MBN/MBP. However, medium levels of N addition (50–100 km/hm2) significantly increased soil MBC/MBP content, significantly reduced MBP, and had no significant effect on other indices. High N addition (100–450 km/hm2) significantly reduced MBC and MBN and significantly increased MBN/MBP content but had no significant effects on MBP, MBC/MBN, or MBC/MBP (Figure 4).
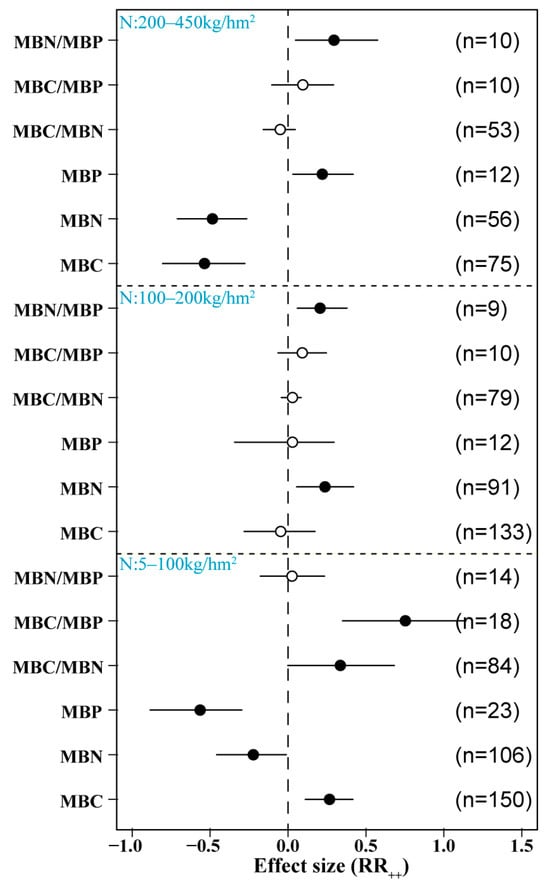
Figure 4.
Effects of different N doses on microbial biomass. Low N: 5–50 kg/hm2; medium N: 50–100 kg/hm2; high N: 100–450 kg/hm2. MBC: soil microbial biomass C, MBN: soil microbial biomass N, MBP: soil microbial biomass P. Number of samples is represented by n. Closed and open symbols indicate significant and non-significant effects, respectively.
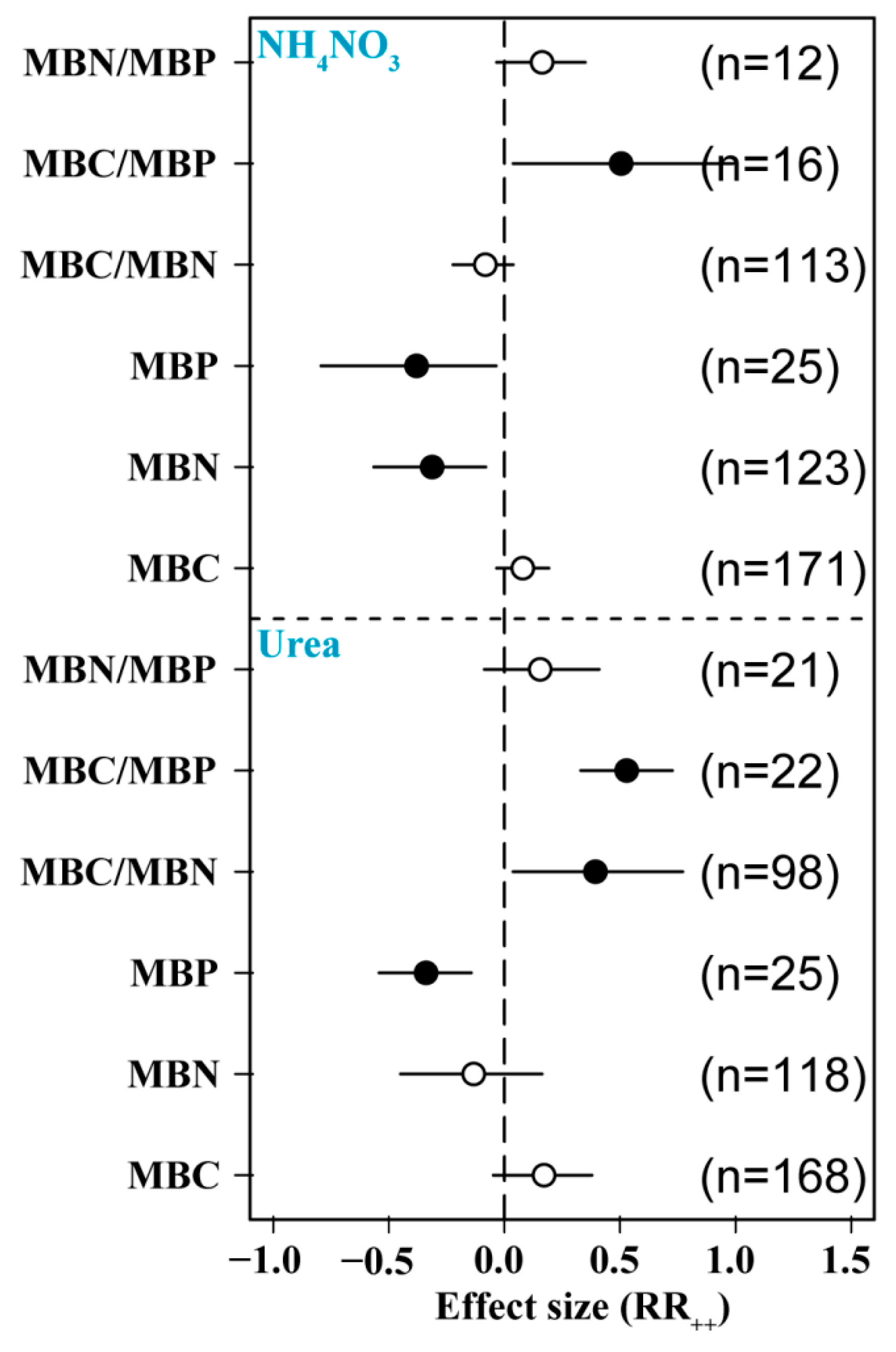
3.4. Effects of N Form on Soil Microbial Biomass
We found differences in the effects of the two major N forms on soil MBC, MBN, and MBP. The application of urea and ammonium nitrate significantly reduced MBP content and significantly increased MBC/MBP. The addition of ammonium nitrate significantly reduced MBN, and the addition of urea significantly increased MBC/MBN, but the addition of different forms of N had no significant effect on other indices (Figure 5).
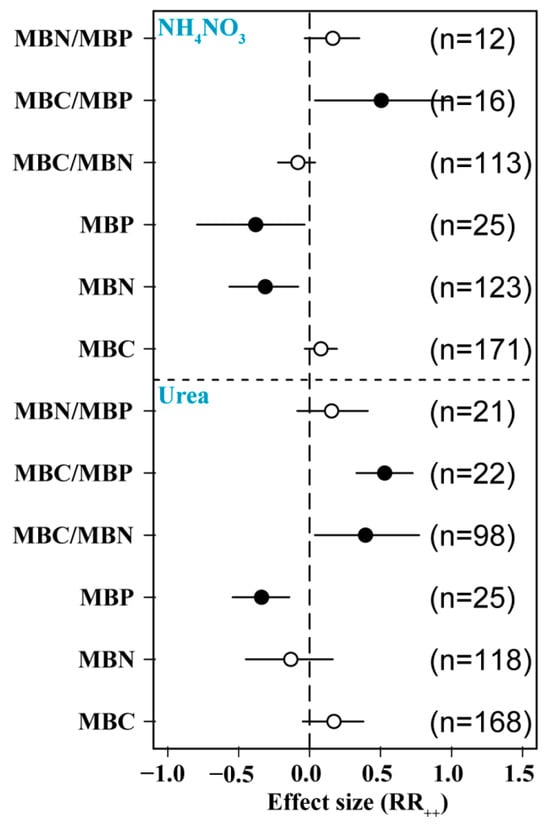
Figure 5.
Effects of N form on soil microbial biomass. MBC: soil microbial biomass C, MBN: soil microbial biomass N, MBP: soil microbial biomass P. Number of samples is represented by n. Closed and open symbols indicate significant and non-significant effects, respectively.
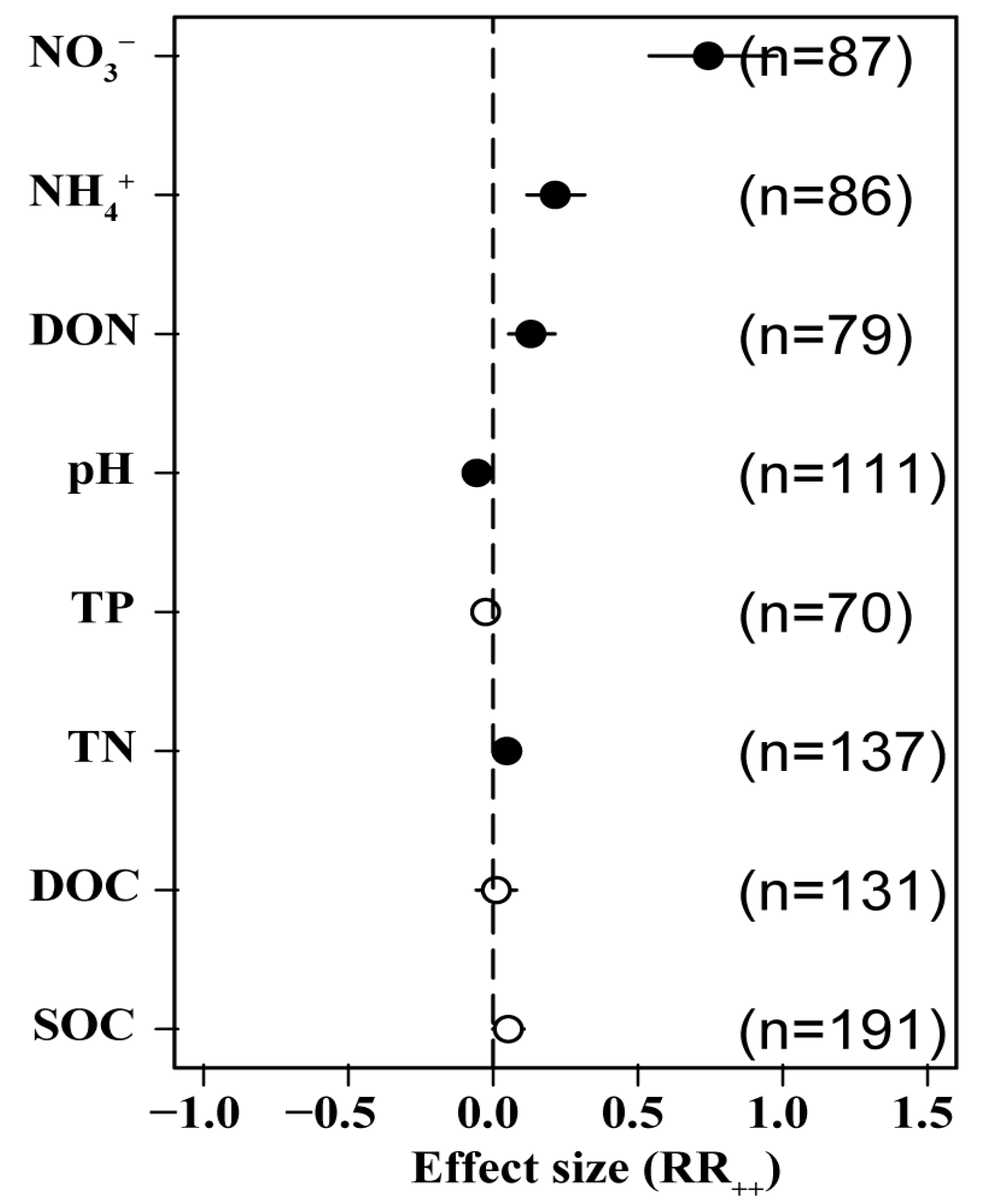
3.5. Effects of N Addition on Soil Chemical Properties
Our integrated analysis revealed that N addition had a positive effect on SOC (5.2%) and dissolved organic C (DOC) (1.1%) and had a significant positive effect on TN (4.7%). It had a negative effect on TP (−2.6%); drove a significant decline in soil pH (−5.6%); and was significantly and positively correlated with dissolved organic N (DON) (13.0%), NH4+ (21.4%), and NO3− (74.3%). This suggests that N application lowers soil pH significantly, causing soil acidification in addition to driving significant increases in soil soluble N (Figure 6).
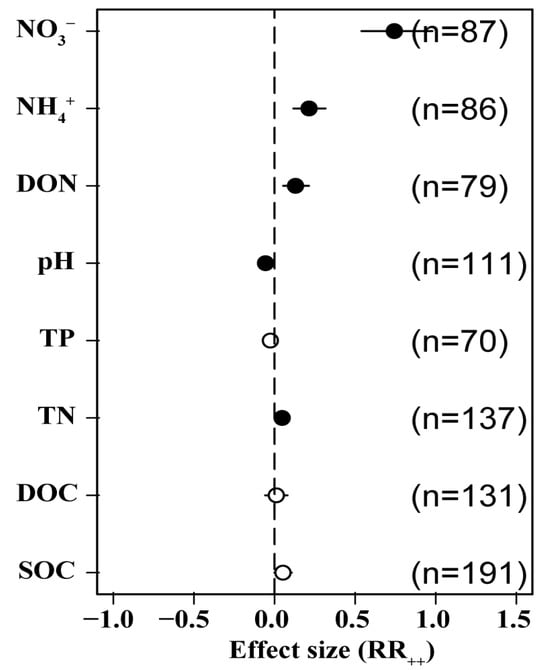
Figure 6.
Effects of N addition on soil physical and chemical properties. SOC: soil organic C, DOC: soluble organic C, TN: soil total N, TP: soil total P, DON: dissolved organic N, NH4+: ammonium N, NO3−: nitrate N. Number of samples is represented by n. Closed and open symbols indicate significant and non-significant effects, respectively.
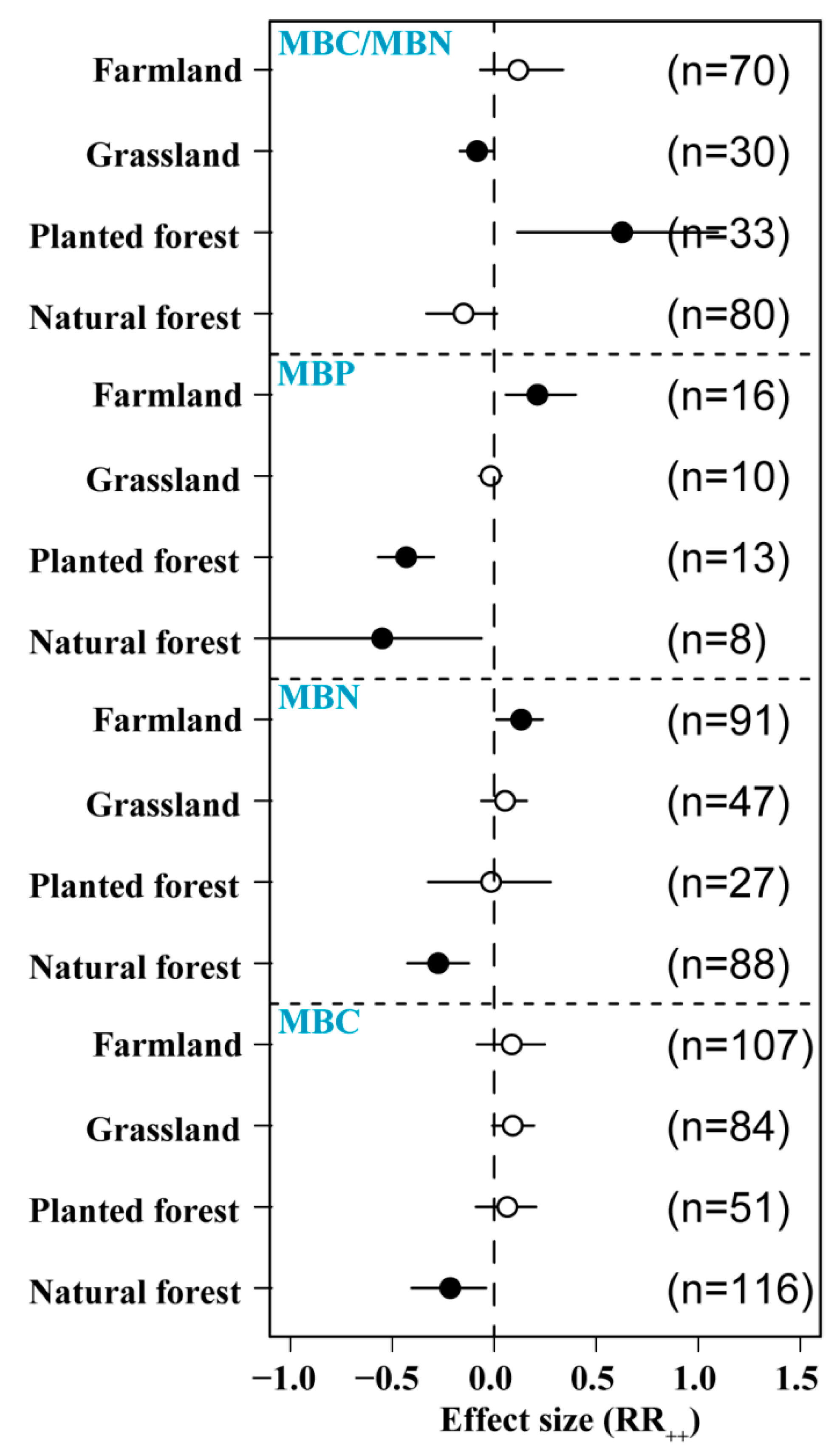
3.6. Effects of N Addition on Microbial Biomass in Different Ecosystems
N addition affected MBC differently across ecosystems, with a significant negative effect in forest soils (−6.5%) and a positive effect in planted forests (9.1%), grasslands (10.3%), and farmlands (12.6%). N addition significantly reduced MBN in natural forest soils (−27.4%) but increased MBN in farmland (13.2%). The increase in MBN was significantly higher in grassland than in forest soils. The application of exogenous N had a significant negative effect on natural forest (−54.9%) and planted forest MBP (−43.1%) but a significant positive effect on farmland MBP (21.1%). The increase in MBP was significantly higher in farmland than in other soil types. We found that N addition significantly increased MBC/MBN in planted forest but significantly reduced MBC/MBN in grassland and had no significant effect on other ecological types (Figure 7).
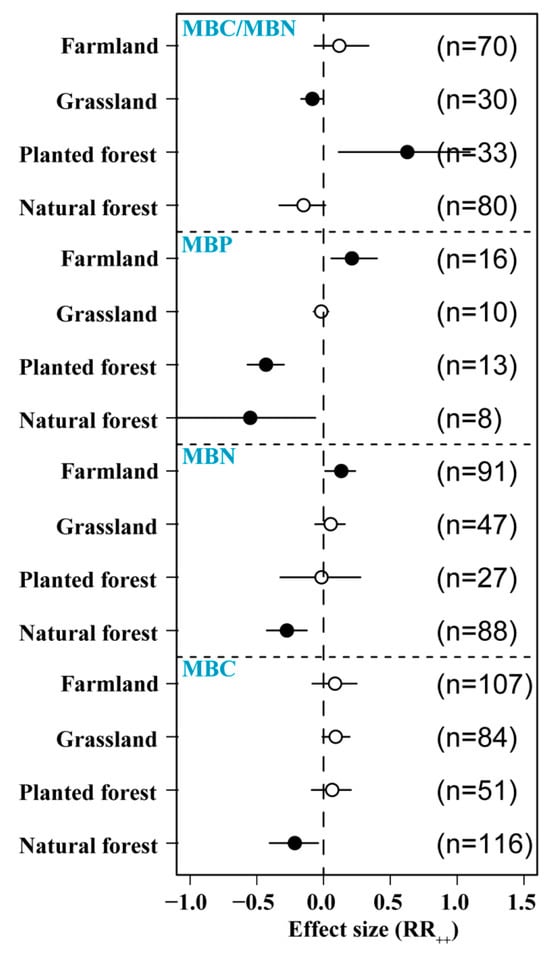
Figure 7.
Effects of N addition on soil microbial biomass in different ecosystems (natural forest, planted forest, grassland, and farmland). Sample number is represented by n. Closed and open symbols indicate significant and non-significant effects, respectively.
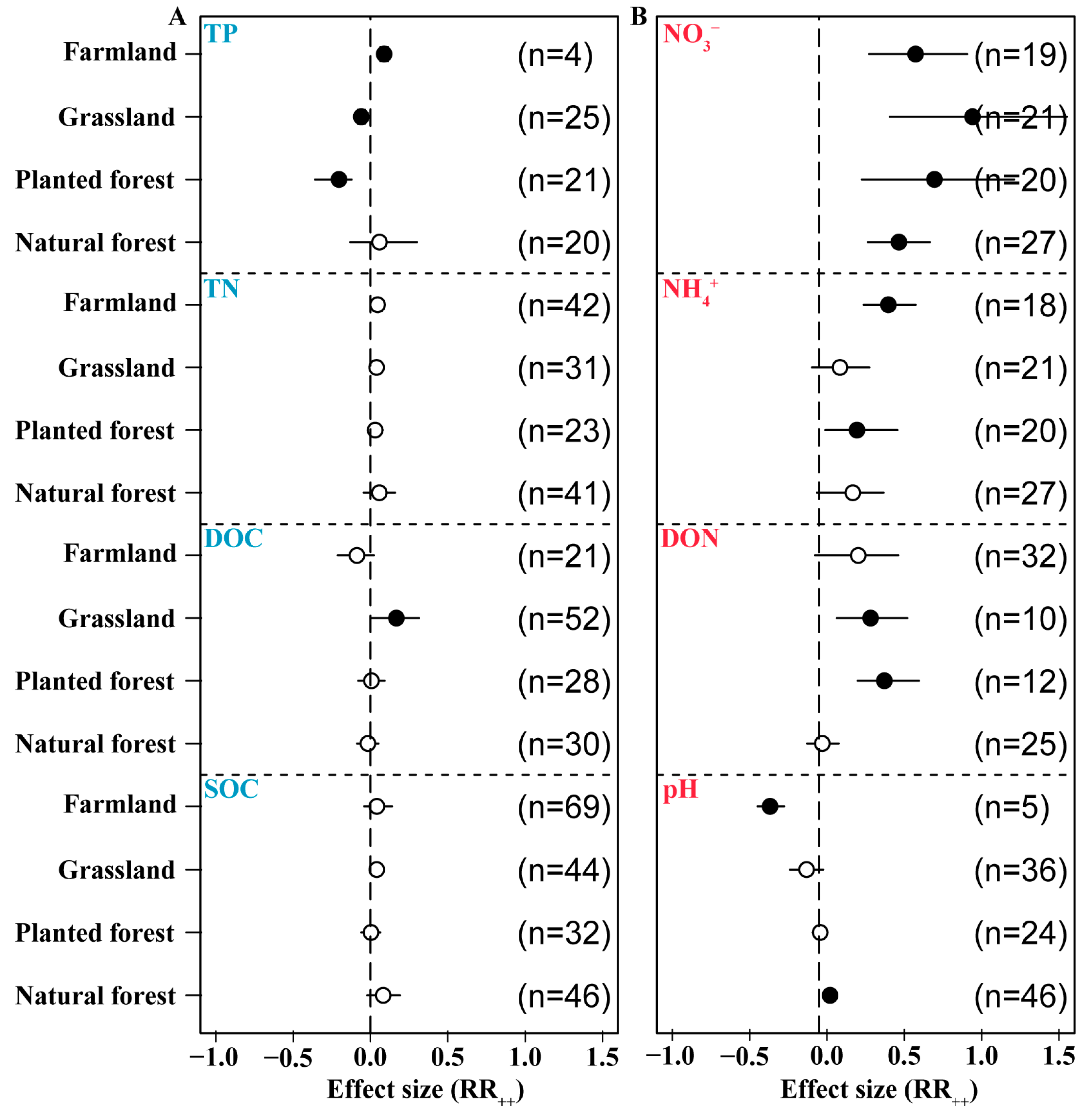
3.7. Effects of N Addition on Soil Chemical Properties in Different Ecosystems
N addition had a positive effect on SOC in different ecosystems, increasing the size of the pool by 8.2%, 0.3%, 4%, and 4.1% in natural forest, planted forest, grassland, and farmland, respectively. Integration analysis revealed that exogenous N application had a negative effect on DOC in forest (-2%) and farmland (−9%) soil but a significant positive effect in grassland (16.7%) soil. N addition was associated with increases in TN in all ecosystem types (5.7%, 3%, 3.9%, 4.6%), but TN only increased significantly in farmland. N application had a positive effect on TP in forest (5.8%) and farmland soils (8.8%) and significant negative effects in planted forest (−20.4%) and grassland soils (−6%) (Figure 8A). N addition had a positive effect on pH in forest soil (2%) but a significant negative effect on soil pH in planted forest (−4.4%), grassland (−13.2%), and farmland (−36.8%). Exogenous N application had a significant positive effect on DON in planted forest (37.1%) and grassland (28.2%). It also had a positive effect on forest, farmland, and grassland soil NH4+ (16.7%, 19.4%, 8.4%) and significantly increased farmland NH4+ (39.7%) as well as NO3− in all ecosystem types (46.5%, 69.5%, 94.1%, 57.3%) (Figure 8B).
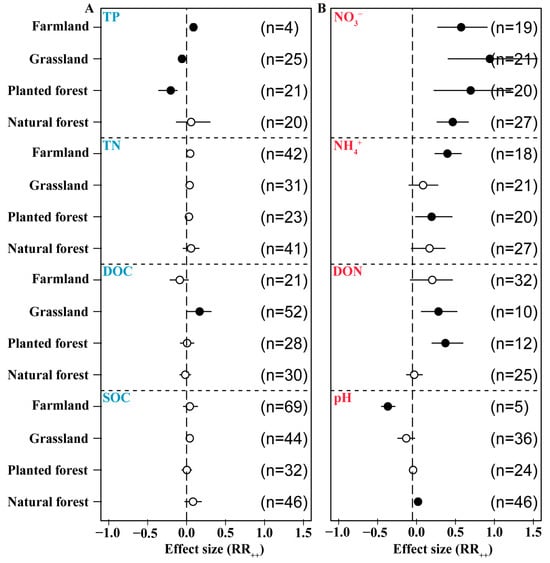
Figure 8.
Effects of N addition on soil physical and chemical factors across ecosystem types. Soil organic C (SOC) and dissolved organic C (DOC), total N (TN), and total P (TP) (A). Soil dissolved organic N (DON), pH, NH4+, and NO3− (B). The number of samples is represented by n. Closed and open symbols indicate significant and non-significant effects, respectively.
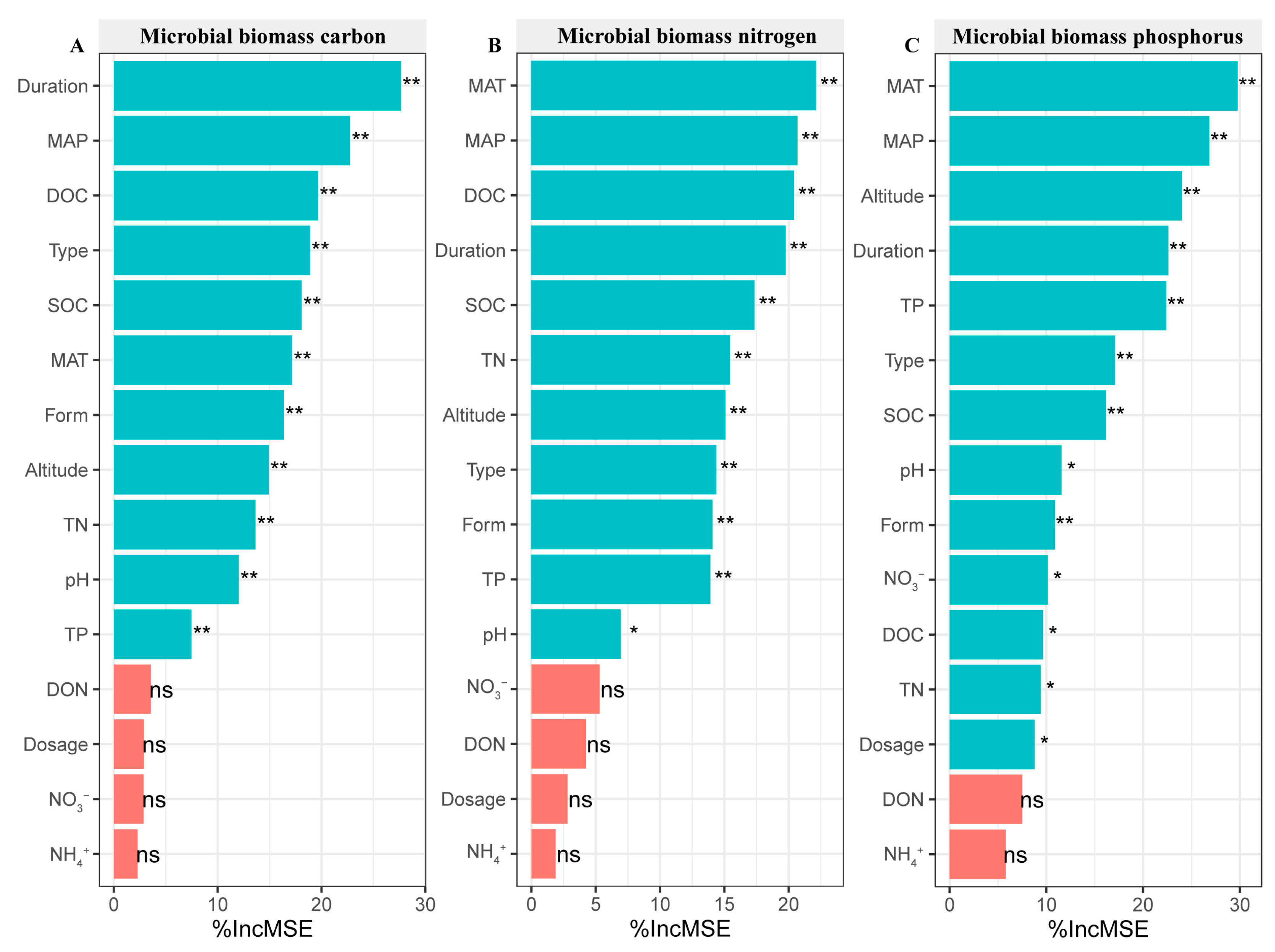
3.8. N Addition Affecting Soil Microbial Biomass Based on Random Forest Analysis
The duration was the most important factor influencing the response of MBC to N addition (Figure 9A). The climate was an important factor affecting the response of MBN and MBP to N addition, and inorganic N, DON, and N dosage use were non-important factors affecting the response of MBC and MBN to N addition (Figure 9B,C).
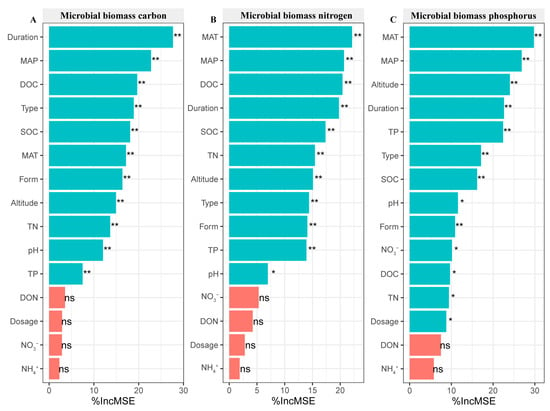
Figure 9.
The effect of N addition on soil microbial biomass C (A), microbial biomass N (B), and microbial biomass P (C) based on random forest analysis. * indicates significance at 0.05 level. ** indicates significance at 0.01 level. ns indicates no significance. SOC: soil organic C, DOC: soluble organic C, TN: soil total N, TP: soil total P, DON: dissolved organic N, NH4+: ammonium N, NO3−: nitrate N, Dosage: N dosage, MAT: mean annual temperature, MAP: mean annual precipitation, Type: soil use type.
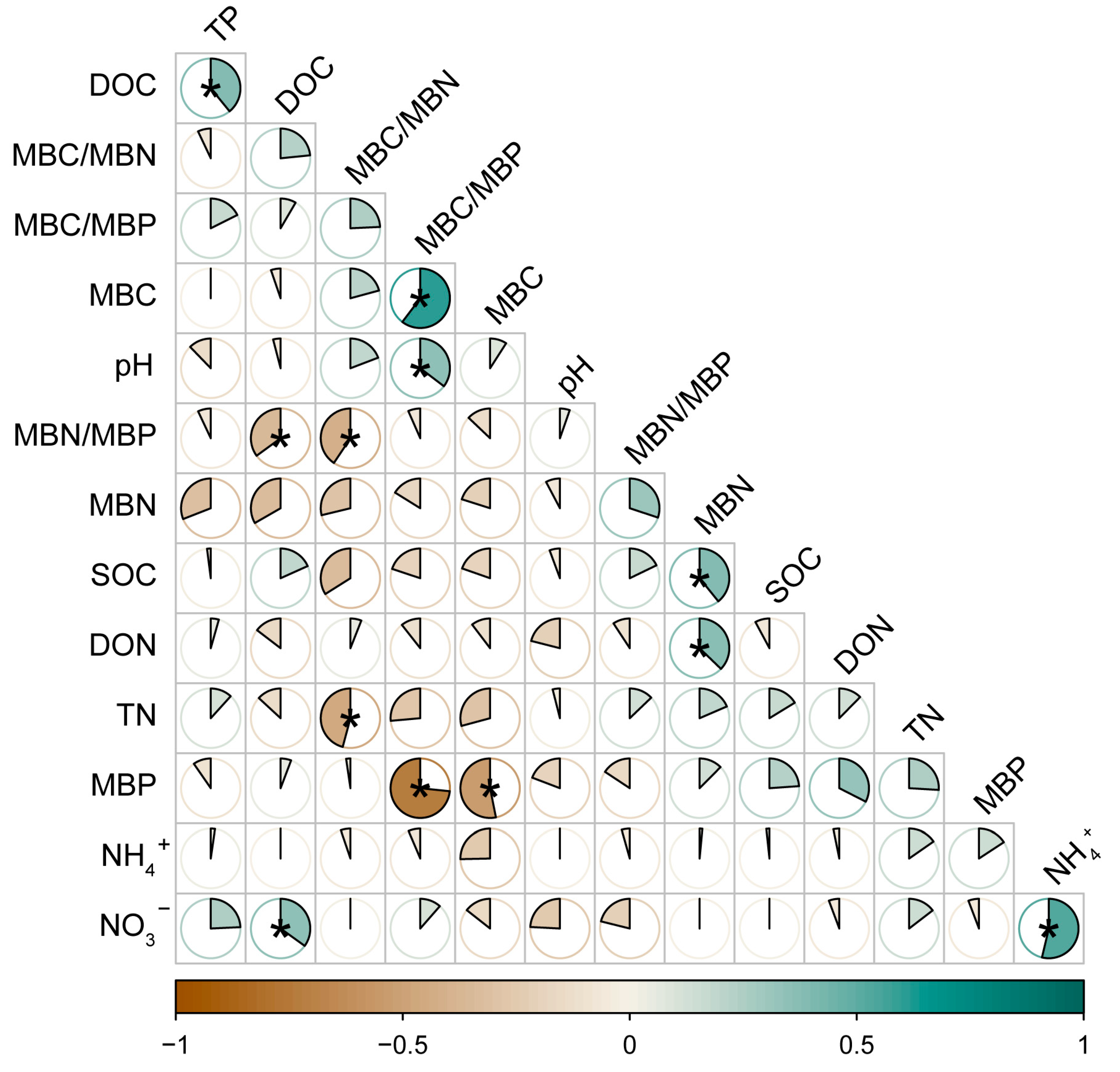
3.9. Correlation Analysis of Soil Physical and Chemical Factors
Our analysis of soil indicators revealed that DOC and TP were significantly positively correlated, MBC/MBP was significantly positively correlated with MBC and pH, and MBN was significantly positively correlated with SOC and DON. MBN/MBP was significantly negatively correlated with DOC, TN was significantly negatively correlated with MBC/MBN, and MBP was significantly negatively correlated with MBC (Figure 10).
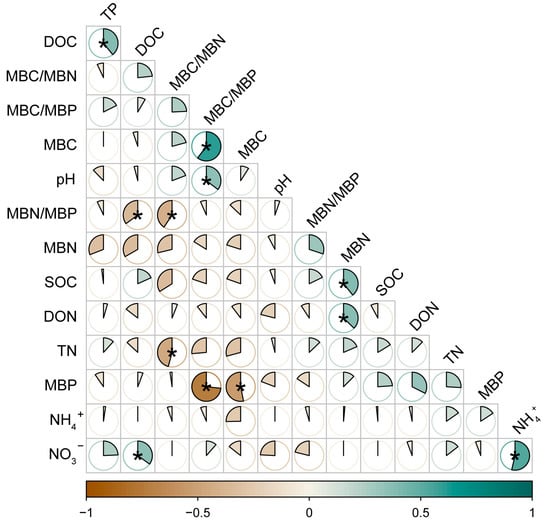
Figure 10.
Correlation analysis between soil chemical properties and soil microbial biomass under N addition. * indicates significance at the 0.05 level. SOC: soil organic C, DOC: soluble organic C, TN: soil total N, TP: soil total P, DON: dissolved organic N, NH4+: ammonium N, NO3−: nitrate N. MBC: soil microbial biomass C, MBN: soil microbial biomass N, MBP: soil microbial biomass P.
3.10. Linear Relationship between Environmental Factors and Soil Microbial Biomass
Integration analysis did not reveal significant linear relationships between any of the environmental factors (altitude, MAT, MAP) and MBC, TN, or TP. However, we identified a positive correlation between altitude and MBP (Figure S1C) and a negative correlation between ln (RR) of MBP and MAT (Figure S1F) and MAP (Figure S1I). Other indicators did not differ significantly (Figure S1A,B,D,E,G,H). Ln (RR) of MBC and MBN were significantly negatively correlated with ln (RR) of MBP and MBC/MBN, respectively. This indicates that an increase in MBC content reduced MBP content, which may have been driven by reductions in MBP resulting from N addition, despite the accumulation of MBC that also results from N fertilization (Figure S1J,N). Ln (RR) of MBN was positively correlated with ln(RR) of MBP, but this relationship was not significant (Figure S1L). Correlations between other indicators were not significant (Figure S1K,M,O).
4. Discussion
4.1. Effect of N Addition on Soil Physical and Chemical Properties
Soil microorganisms control feedback between SOC pools and the atmosphere and thus play a key role in terrestrial C cycling [37,38,39]. In terrestrial ecosystems, soil MBC is an important regulator of the C cycle [40,41,42]. Different land use practices and fertilization treatments alter microbial C use indirectly, likely by changing plant community structure, root systems and their secretions, soil organic matter concentration, and soil pH. Changes induced by human activity also alter microbial activity directly via their influence on soil pH and water content [43,44]. Moreover, N addition can promote microbial nitrification [45,46,47,48] and cause soil acidification, thus reducing populations of soil microbes [49,50,51]. It is widely acknowledged that changes in subsurface soil properties may be caused by differences in soil microbial community structure, as microorganisms are key drivers of biogeochemical processes [52,53]. In this study, we found that N addition drove significantly lower soil pH while causing significant increases in DON and inorganic N. These changes to soil physical and chemical properties likely affected soil microbial community composition to some degree.
4.2. Effects of N Addition on Positive and Negative Directions of Soil MBC and MBN
N addition can increase populations of soil bacteria (especially Actinomycetes) and fungi during the crop growth period [54,55,56], but high levels of fertilization can reduce their size [57,58,59]. Some research suggests that MBC and MBN content is strongly influenced by N addition, with the application of moderate amounts optimizing MBC and MBN content [60,61] but the application of excessive amounts reducing MBN content [62,63]. In this study, N addition (low and high N) also reduced MBN content and significantly impacted MBN in forest systems. Previous research has demonstrated that different fertilization regimes (C addition, N addition, and C + N addition) increase MBC and MBN to varying degrees [64,65,66]. Some work suggests that MBC decreases associated with N fertilization may result from the wide distribution of microorganisms and the high microbial taxonomic variability across different forest sites [22,67,68,69,70]. A ten-year-long experiment conducted by Pregitzer et al. in Michigan, USA, found that long-term N deposition increased tree biomass and SOC stocks [71]. Similarly, a five-year-long tropical experiment conducted by Cusack et al. revealed that improved soil nutrient status resulting from N addition may increase MBC by enhancing nutrient availability, thereby supporting microbial growth and reproduction [72]. Consistent with the variability documented in previous work, we found that low N addition increases MBC content but decreases MBP content. This variability is likely due to the fact ecosystem type strongly influences MBC and MBN and how they respond to N application. In contrast to our findings, other experiments have found that long-term fertilization reduces soil MBC. This inconsistency may be due to differences in temporal scale: short-term fertilization experiments may not provide sufficient time for microorganisms to respond to changing environmental conditions. Forest soil represents a complex system in which many factors, including soil moisture, pH, and apoplankton content [73], influence soil microbial biomass content. Some studies have shown that N addition reduces MBC and MBN in grassland and forest soils [74]. Here, we found that N addition did not significantly affect microbial biomass in grassland soil but significantly reduced grassland MBC/MBN. Other work found that temperature differences did not influence MBC or MBN. This may be due to the fact that soil microorganisms have become well adapted to temperature fluctuations over time [75]. Recent work found that MBC and MBN decreased under both high soil moisture conditions and fertilization regimes in which large quantities of N were applied [11]. Other studies have shown that the toxicity resulting from the accumulation of NH4+ and NO3− under fertilization can directly affect soil microorganisms. A synthesis of 65 N addition trials found that N addition reduced MBC content by 9.5% [76,77]. Moderate N addition can alleviate N limitation and provide a substrate and energy for microbial growth and basal metabolism, thereby stimulating soil nutrient effectiveness and improving microbial functional activity [78]. Reductions in soil MBC and MBN content may also be driven by excessive N addition, which reduces salt ion concentrations while increasing the concentration of Al(III). The resulting cell lysis drives overall lower microbial activity [79]. In this study, different ecosystems respond differently to N addition. Diminished MBC, MBN, and MBP in forest ecosystems, which are characterized by low N content, may have been caused by the inability of soil microbes to cope with the addition of large quantities of N. In contrast, microbial community composition in agroecosystems tends to be more homogeneous and resistant to the influence of exogenous N inputs. Therefore, these microorganisms can absorb large amounts of N to support growth and reproduction, thereby increasing microbial biomass.
4.3. Effects of N Addition on MBP and MBC/MBN
Recent work found that N addition did not significantly influence MBP [80], consistent with the results of a meta-analysis of 192 N addition studies. This work reported a slight, non-significant decline in MBP that increased with the duration and amount of N addition [81]. Sensitivity to N addition appears to vary across microbial taxa. Studies in subtropical forest soils found no significant change in soil fungal biomass after N application [82] but reported reduced bacterial biomass with continued N input [83]. Despite these findings, other work has observed increases in all soil P fractions following N application [84,85]. Microbial C/P and N/P reflect the potential for microbial P release and ecosystem nutrient limitation, respectively, and both ratios may vary as MBC/MBP changes. This further suggests that N deposition attenuates microbial P release and exacerbates soil P limitation [86,87]. Here, correlation analysis found a significant negative correlation between MBC and MBP, indicating that N addition may have reduced MBP indirectly via its promotion of MBC accumulation. In contrast, we also found that N addition increased MBP in farmland soils, but this may have been caused by tillage practices and altered soil microbial community composition. Soil MBC/MBN can be used as an indicator of altered feedback between microbial community composition and soil N status within a region [88,89]. Studies have shown that MBC/MBN values for soil bacterial and fungal communities are in the ranges of 3–5 and 4–15, respectively [90,91]. We found that changes to microbial stoichiometric ratios were larger than changes to overall soil stoichiometry, but that microbial and soil C/N/P ratios mirrored one another. This may be due to the relatively greater sensitivity of microbial C, N, and P to changes in soil nutrient status [92]. Moreover, soil microbial biomass C/N is influenced by the ratio of fungi/bacteria [93]. Previous findings that MBC, MBN, and MBP declined in response to fertilization may have been the result of reduced microbial biomass driven by the substrate acidification and lignin and cellulose degradation that results from N addition [94,95,96,97,98]. In this study, N addition significantly increased MBC/MBN. We found that the intensity of this effect varied across land use type (forest, planted forest, grassland, and farmland), but the relationship between MBC/MBN and land cover was not significant. We also found that MBN was significantly negatively correlated with MBC/MBN.
5. Conclusions
This integrated analysis of 86 studies conducted across the globe revealed that N addition significantly affected soil microbial biomass. This effect was most pronounced under low N addition (5–50 kg/hm2), which significantly affected soil microbial biomass, mainly by increasing soil MBC but also by decreasing MBP and driving higher MBC/MBP. N addition affected soil physicochemical properties, significantly reducing soil pH and significantly increasing soil DON and inorganic N. Our analysis also revealed that the effects of N application vary across ecosystems: N addition significantly decreased MBP and TP in planted forest but did not significantly affect soil microbial biomass in grassland. In farmland soil, N addition significantly increased total P, NH4+, NO3−, MBN, and MBP but significantly decreased pH. Moreover, although N addition increases soil microbial biomass and thus supports higher yields resulting from the accumulation of above-ground biomass, the effects of fertilization vary with the quantity applied. Excessive N application can negatively affect forest ecosystems, thus affecting the global N balance.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/agriculture14091616/s1, Figure S1. Linear relationship between environmental factors and soil microbial biomass C, N, and P under N addition. MBC: soil microbial biomass C, MBN: soil microbial biomass N, MBP: soil microbial biomass P, MAT: mean annual temperature, MAP: mean annual precipitation; Table S1. Literature resources.
Author Contributions
Writing—original draft preparation, C.H.; writing—review and editing, Z.J. and Y.R. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by the Project of Sanya Yazhou Bay Science and Technology City (SCKJ-JYRC-2022-94), Strategic Priority Research Program of Chinese Academy of Sciences (XDA28020203), Postgraduate Innovation Research Project of Hainan Province (Qhyb2022-67), PhD Scientific Research and Innovation Foundation of Sanya Yazhou Bay Science and Technology City (HSPHDSRF-2023-12-008), and the National Natural Science Foundation of China (91751204, 32160750).
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Acknowledgments
We would like to thank Daniel Petticord at the University of Cornell for his assistance with English language and grammatical editing of the manuscript.
Conflicts of Interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
References
- Canfield, D.E.; Glazer, A.N.; Falkowski, P.G. The Evolution and Future of Earth’s Nitrogen Cycle. Science 2010, 330, 192–196. [Google Scholar] [CrossRef] [PubMed]
- Ti, C.; Gao, B.; Luo, Y.; Wang, S.; Chang, S.X.; Yan, X. Dry deposition of N has a major impact on surface water quality in the Taihu Lake region in southeast China. Atmos. Environ. 2018, 190, 1–9. [Google Scholar] [CrossRef]
- Wang, W.; Liu, X.J. Research progress on nitrogen deposition and grassland ecosystem response in Qinghai-Tibet Plateau. J. China Agric. Univ. 2018, 23, 151–158. [Google Scholar]
- Deng, S.P.; Moore, J.M.; Tabatabai, M.A. Characterization of active nitrogen pools in soils under different cropping systems. Biol. Fertil. Soils 2000, 32, 302–309. [Google Scholar] [CrossRef]
- Belay-Tedla, A.; Zhou, X.; Su, B.; Wan, S.; Luo, Y. Labile, recalcitrant, and microbial carbon and nitrogen pools of a tall grass prairie soil in the US Great Plains subjected to experimental warming and clipping. Soil Biol. Biochem. 2009, 41, 110–116. [Google Scholar] [CrossRef]
- Zhang, G.S.; Huang, G.B. Soil organic carbon sequestration potential in cropland. Acta Ecol. Sin. 2005, 25, 351–357. [Google Scholar]
- Sun, T.; Wang, Y.; Hui, D.; Jing, X.; Feng, W. Soil properties rather than climate and ecosystem type control the vertical variations of soil organic carbon, microbial carbon, and microbial quotient. Soil Biol. Biochem. 2020, 148, 107905. [Google Scholar] [CrossRef]
- Qualls, R.; Haines, B.; Swank, W.; Tyler, S. Retention of soluble organic nutrients by a forested ecosystem. Biogeochemistry 2002, 61, 135–171. [Google Scholar] [CrossRef]
- Hartmann, M.; Frey, B.; Mayer, J.; Mäder, P.; Widmer, F. Distinct soil microbial diversity under long-term organic and conventional farming. ISME J. 2015, 9, 1177–1194. [Google Scholar] [CrossRef]
- Dou, X.L.; He, P.; Cheng, X.; Zhou, W. Long-term fertilization alters chemically-separated soil organic carbon pools: Based on stable C isotope analyses. Sci. Rep. 2016, 6, 19061. [Google Scholar] [CrossRef]
- Zhang, T.A.; Chen, H.Y.; Ruan, H. Global negative effects of nitrogen deposition on soil microbes. ISME J. 2018, 12, 1817–1825. [Google Scholar] [CrossRef] [PubMed]
- Yuan, X.C.; Lin, W.S.; Pu, X.T.; Yang, Z.R.; Zheng, W.; Chen, Y.; Yang, Y. Effects of forest regeneration patterns on the quantity and chemical structure of soil solution dissolved organic carbon matter in subtropical forest. Chin. J. Appl. Ecol. 2016, 27, 1845–1852. [Google Scholar]
- Bargali, K.; Manral, V.; Padalia, K.; Bargali, S.S.; Upadhyay, V.P. Effect of vegetation type and season on microbial biomass carbon in Central Himalayan forest soils, India. Catena 2018, 171, 125–135. [Google Scholar] [CrossRef]
- Liu, Z.J.; Zhou, W.; Shen, J.B.; Li, S.T.; Ai, C. Soil quality assessment of yellow clayey paddy soils with different productivity. Biol. Fertil. Soils 2014, 50, 537–548. [Google Scholar] [CrossRef]
- Zhou, Y.; Chen, Y.; Jiang, F.; Hu, Y.; Long, L.; Pei, L.; Li, J.; Xu, K. Responses of soil microbial biomass carbon, nitrogen and microbial entropy to different materials returned to corn fields. J. Soil Water Conserv. 2020, 34, 173–180. [Google Scholar]
- Kiboi, M.; Ngetich, K.; Mugendi, D.; Muriuki, A.; Adamtey, N.; Fliessbach, A. Microbial biomass and acid phosphomonoesterase activity in soils of the Central Highlands of Kenya. Geoderma Reg. 2018, 15, e00193. [Google Scholar] [CrossRef]
- Xu, Y.; Zhong, Z.; Zhang, W.; Han, X.; Yang, G.; Ren, C.; Feng, Y.; Ren, G.; Wang, X. Responses of soil nosZ–type denitrifying microbial communities to the various land–use types of the Loess Plateau, China. Soil Tillage Res. 2019, 195, 104378. [Google Scholar] [CrossRef]
- Zheng, J.S.; Hu, J.M.; Wei, X.H.; Wei, Y.; Su, S.; Li, T.; Xia, X.; Yu, Y.; Zhang, J. Effects of green manure returning on soil microbial biomass carbon and mineralization of organic carbon in smash ridging paddy field. Chin. J. Eco-Agric. 2021, 29, 691–703. [Google Scholar]
- Kuzyakov, Y.; Friedel, J.; Stahr, K. Review of mechanisms and quantification of priming effects. Soil Biol. Biochem. 2000, 32, 1485–1498. [Google Scholar] [CrossRef]
- Lin, M.; Deng, S.; Su, Y.; Liu, K.; Li, F. Effects of fertilization on soil active organic carbon and carbon sequestration of forage in Karst region. Plant Nutr. Fertil. Sci. 2012, 18, 1119–1126. [Google Scholar]
- Allison, S.D.; Lu, Y.; Weihe, C.; Goulden, M.L.; Martiny, A.C.; Treseder, K.K.; Martiny, J.B. Microbial abundanceand composition influence litter decomposition responseto environmental change. Ecology 2013, 94, 714–725. [Google Scholar] [CrossRef] [PubMed]
- Wallenstein, M.D.; McNulty, S.; Fernandez, I.J.; Boggs, J.; Schlesinger, W.H. Nitrogen fertilization decreases forest soil fungal and bacterial biomass in three long-term experiments. For. Ecol. Manag. 2006, 222, 459–468. [Google Scholar] [CrossRef]
- Zhang, T.T.; Chen, S.T.; Wang, J.; Wang, Z.H.; Hu, Z.H. Effects of warming and straw application on soil microbial biomass carbon and nitrogen and bacterial community structure. Environ. Sci. 2019, 40, 4718–4724. [Google Scholar]
- Yan, G.; Yao, Z.; Zheng, X.; Liu, C. Characteristics of annual nitrous and nitri coxide emissions from majorce-realcropsinthe North China Pain under alternative fertilizer management. Agric. Eosc Ystems Environ. 2015, 207, 67–78. [Google Scholar] [CrossRef]
- Liu, S.; Li, Y.; Wu, J.; Huang, D.; Su, Y.; Wei, W. Spatial variability of soil microbial biomass carbon, nitrogen and phosphorus in a hilly red soil landscape in subtropical China. Soil Sci. Plant Nutr. 2010, 56, 693–704. [Google Scholar] [CrossRef]
- Li, T.T.; Nie, F.; Yang, W.; Yang, K.; He, R.Y.; Zhang, L.; Li, Z.J.; Xu, Z.F. Seasonal dynamics of labile soil nitrogen from four plantations at the western edge of the Sichuan basin. Chin. J. Appl. Environ. Biol. 2018, 24, 744–750. [Google Scholar]
- Hobbie, E.A.; Ouimette, A.P. Controls of nitrogen isotope patterns in soil profiles. Biogeochemistry 2009, 95, 355–371. [Google Scholar] [CrossRef]
- Iqbal, J.; Hu, R.; Feng, M.; Lin, S.; Malghani, S.; Ali, I.M. Microbial biomass, and dissolved organic carbon and nitrogen strongly affect soil respiration in different land uses: A case study at Three Gorges Reservoir Area, South China. Agric. Ecosyst. Environ. 2010, 137, 294–307. [Google Scholar] [CrossRef]
- Zhang, J.; Wen, X.; Liao, Y.; Liu, Y. Effects of different amount of maize straw returning on soil fertility and yield of winter wheat. Plant Nutr. Fertil. Sci. 2010, 16, 612–619. [Google Scholar]
- Dempster, D.N.; Gleeson, D.; Solaiman, Z.; Jones, D.L.; Murphy, D. Decreased soil microbial biomass and nitrogen mineralisation with Eucalyptus biochar addition to a coarse textured soil. Plant Soil 2012, 354, 311–324. [Google Scholar] [CrossRef]
- Wang, R.; Filley, T.R.; Xu, Z.; Wang, X.; Li, M.-H.; Zhang, Y.; Luo, W.; Jiang, Y. Coupled response of soil carbon and nitrogen pools and enzyme activities to nitrogen and water addition in a semi-arid grassland of Inner Mongolia. Plant Soil 2014, 381, 323–336. [Google Scholar] [CrossRef]
- Wei, C.; Yu, Q.; Bai, E.; Lü, X.; Li, Q.; Xia, J.; Kardol, P.; Liang, W.; Wang, Z.; Han, X. Nitrogen deposition weakens plant–microbe interactions in grassland ecosystems. Glob. Chang. Biol. 2013, 19, 3688–3697. [Google Scholar] [CrossRef] [PubMed]
- Lin, Q.M. Interference of soil soluble inorganic phosphorus on the determination of microbial biomass phosphorus. Acta Ecol. Sin. 2001, 21, 993–996. [Google Scholar]
- Chen, G.C.; He, Z.L.; Wang, Y.J. Impact of pH on microbial biomass carbon and microbial biomass phosphorus in red soils. Pedosphere 2004, 14, 9–15. [Google Scholar]
- Parfitt, R.L. Phosphate adsorption on an oxisol. Soil Sci. Soc. Am. J. 1977, 41, 1064–1067. [Google Scholar] [CrossRef]
- Guo, Q. Soil acidification induced by nitrogen addition and its responses to water addition in Inner Mongolia Temperate Steppe, China. J. Appl. Ecol. 2019, 30, 3285–3291. [Google Scholar]
- Chen, D.D.; Zhang, S.H.; Dong, S.K.; Wang, X.T.; Du, G.Z. Effect of land-use on soil nutrients and microbial biomass of an alpine region on the northeastern Tibetan plateau, China. Land Degrad. Dev. 2010, 21, 446–452. [Google Scholar] [CrossRef]
- Li, Y.; Dong, S.; Wen, L.; Wang, X.; Wu, Y. The effects of fencing on carbon stocks in the degraded alpine grasslands of the Qinghai-Tibetan Plateau. J. Environ. Manag. 2013, 128, 393–399. [Google Scholar] [CrossRef]
- Nie, M.; Pendall, E.; Bell, C.; Gasch, C.K.; Raut, S.; Tamang, S.; Wallenstein, M.D. Positive climate feedbacks of soil microbial communities in a semi-arid grassland. Ecol. Lett. 2013, 16, 234–241. [Google Scholar] [CrossRef]
- Li, X.; Chen, Z. Soil microbial biomass C and N along a climatic transect in the Mongolian steppe. Biol. Fertil. Soils 2004, 39, 344–351. [Google Scholar] [CrossRef]
- Qiu, L.; Wei, X.; Zhang, X.; Cheng, J.; Gale, W.; Guo, C.; Long, T. Soil organic carbon losses due to land use change in a semiarid grassland. Plant Soil 2012, 355, 299–309. [Google Scholar] [CrossRef]
- Qiao, N.; Xu, X.; Cao, G.; Ouyang, H.; Kuzyakov, Y. Land use change decreases soil carbon stocks in Tibetan grasslands. Plant Soil 2015, 395, 231–241. [Google Scholar] [CrossRef]
- Liu, S.; An, N.; Yang, J.; Dong, S.; Wang, C.; Yin, Y. Prediction of soil organic matter variability associated with different land use types in mountainous landscape in southwestern Yunnan province, China. Catena 2015, 133, 137–144. [Google Scholar] [CrossRef]
- Lu, X.; Yan, Y.; Sun, J.; Zhang, X.; Chen, Y.; Wang, X.; Cheng, G. Carbon, nitrogen, and phosphorus storage in alpine grassland ecosystems of Tibet: Effects of grazing exclusion. Ecol. Evol. 2015, 5, 4492–4504. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Tian, D.; Wang, B.; Wang, J.; Wang, S.; Chen, H.Y.H.; Xu, X.; Wang, C.; He, N.; Niu, S. Microbes drive global soil nitrogen mineralization and availability. Glob. Chang. Biol. 2019, 25, 1078–1088. [Google Scholar] [CrossRef]
- Miller, K.S.; Geisseler, D. Temperature sensitivity of nitrogen mineralization in agricultural soils. Biol. Fertil. Soils 2018, 54, 853–860. [Google Scholar] [CrossRef]
- Taylor, A.E.; Myrold, D.D.; Bottomley, P.J. Temperature affects the kinetics of nitrite oxidation and nitrification coupling in four agricultural soils. Soil Biol. Biochem. 2019, 136, 107523. [Google Scholar] [CrossRef]
- Thangarajan, R.; Bolan, N.S.; Naidu, R.; Surapaneni, A. Effects of temperature and amendments on nitrogen mineralization in selected Australian soils. Environ. Sci. Pollut. Res. 2015, 22, 8843–8854. [Google Scholar] [CrossRef]
- Wang, C.; Liu, D.; Bai, E. Decreasing soil microbial diversity is associated with decreasing microbial biomass under nitrogen addition. Soil Biol. Biochem. 2018, 120, 126–133. [Google Scholar] [CrossRef]
- Högberg, M.N.; Högberg, P.; Myrold, D.D. Is microbial community composition in boreal forest soils determined by pH, C-to-N ratio, the trees, or all three? Oecologia 2007, 150, 590–601. [Google Scholar] [CrossRef]
- Lebauer, D.S.; Treseder, K.K. Nitrogen limitation of net primary productivity in terrestrial ecosystems is globally distributed. Ecology 2008, 89, 371–379. [Google Scholar] [CrossRef] [PubMed]
- Cheng, H.; Yuan, M.; Duan, Q.; Sun, R.; Shen, Y.; Yu, Q.; Li, S. Influence of phosphorus fertilization patterns on the bacterial community in upland farmland. Ind. Crop. Prod. 2020, 155, 112761. [Google Scholar] [CrossRef]
- Liu, C.; Gong, X.; Dang, K.; Li, J.; Yang, P.; Gao, X.; Deng, X.; Feng, B. Linkages between nutrient ratio and the microbial community in rhizosphere soil following fertilizer management. Environ. Res. 2020, 184, 109261. [Google Scholar] [CrossRef] [PubMed]
- Vepsäläinen, M.; Kukkonen, S.; Vestberg, M.; Sirviö, H.; Niemi, R.M. Application of soilenzyme activity test kit in a field experiment. Soil Biol. Biochem. 2001, 33, 1665–1672. [Google Scholar] [CrossRef]
- Fageria, N.K.; Baligar, V.C. Enhancing Nitrogen Use Efficiency in Crop Plants. Adv. Agron. 2005, 88, 97–185. [Google Scholar]
- Changhui, W.; Feng, Z.; Xiang, Z.; Kuanhu, D. The effects of N and P additions on microbial N transformations and biomass on saline-alkaline grassland of loess plateau of northern china. Geoderma 2014, 213, 419–425. [Google Scholar] [CrossRef]
- Guo, T.C.; Song, X.; Ma, D.Y.; Cha, F.N.; Yue, Y.J.; Zhang, Y.; Li, Y.Z. Effect of nitrogen fertilizer on soil enzymatic activity and rhizosphere microorganisms of wheat. J. Soil Water Conserv. 2006, 20, 129–132. [Google Scholar]
- Liu, W.; Jiang, L.; Hu, S.; Li, L.; Liu, L.; Wan, S. Decoupling of soil microbes and plants with increasing anthropogenic nitrogen inputs in a temperate steppe. Soil Biol. Biochem. 2014, 72, 116–122. [Google Scholar] [CrossRef]
- Zhang, Y.; Han, X.; He, N.; Long, M.; Huang, J.; Zhang, G.; Wang, Q.; Han, X. Increase in ammonia volatilization from soil in response to N deposition in Inner Mongolia grasslands. Atmos. Environ. 2014, 84, 156–162. [Google Scholar] [CrossRef]
- Mangalassery, S.; Kalaivanan, D.; Philip, P.S. Effect of inorganic fertilisers and organic amendments on soil aggregation and biochemical characteristics in a weathered tropical soil. Soil Tillage Res. 2019, 187, 144–151. [Google Scholar] [CrossRef]
- Ren, F.; Sun, N.; Xu, M.; Zhang, X.; Wu, L.; Xu, M. Changes in soil microbial biomass with manure application in cropping systems: A meta-analysis. Soil Tillage Res. 2019, 194, 104291. [Google Scholar] [CrossRef]
- Shen, S.M.; Hart, P.B.S.; Powlson, D.S.; Jenkinson, D.S. The nitrogen cycle in the Broadbalk wheat experiment: 15N-labeled fertilizer residues in soil and in the soil microbial biomass. Soil Biol. Biochem. 1989, 21, 529–533. [Google Scholar] [CrossRef]
- Qiu, S.; Peng, P.; Liu, Q.; Rong, X. Soil microbial biomass nitrogen and its role in nitrogen cycling. Chin. J. Ecol. 2006, 25, 443–448. [Google Scholar]
- Huang, G.; Li, Y.; Su, Y.G. Divergent responses of soil microbial communities to water and nitrogen addition in a temperate desert. Geoderma 2015, 251–252, 55–64. [Google Scholar] [CrossRef]
- Sinsabaugh, R.L.; Belnap, J.; Rudgers, J.; Kuske, C.R.; Martinez, N.; Sandquist, D. Soil microbial responses to nitrogen addition in arid ecosystems. Front. Microbiol. 2015, 6, 819. [Google Scholar] [CrossRef]
- Geisseler, D.; Scow, K.M. Long-term effects of mineral fertilizers on soil microorganisms-A review. Soil Biol. Biochem. 2014, 75, 54–63. [Google Scholar] [CrossRef]
- Cusack, D.F.; Silver, W.L.; Torn, M.S.; Burton, S.D.; Firestone, M.K. Changes in microbial community characteristics and soil organic matter with nitrogen additions in two tropical forests. Ecology 2011, 92, 621–632. [Google Scholar] [CrossRef]
- Heinze, S.; Raupp, J.; Joergensen, R.G. Effects of fertilizer and spatial heterogeneity in soil pH on microbial biomass indices in a long-term field trial of organic agriculture. Plant Soil 2010, 328, 203–215. [Google Scholar] [CrossRef]
- Van Diepen, L.T.A.; Lilleskov, E.A.; Pregitzer, K.S.; Miller, R.M. Simulated Nitrogen Deposition Causes a Decline of Intra- and Extraradical Abundance of Arbuscular Mycorrhizal Fungi and Changes in Microbial Community Structure in Northern Hardwood Forests. Ecosystems 2010, 13, 683–695. [Google Scholar] [CrossRef]
- Wang, Q.; Zhang, W.; Sun, T.; Chen, L.; Pang, X.; Wang, Y.; Xiao, F. N and P fertilization reduced soil autotrophic and heterotrophic respiration in a young Cunninghamia lanceolata forest. Agric. For. Meteorol. 2017, 232, 66–73. [Google Scholar] [CrossRef]
- Pregitzer, K.S.; Burton, A.J.; Zak, D.R.; Talhelm, A.F. Stimulated chronic nitrogen deposition increase carbon storage in Northern Temperate forests. Glob. Change Biol. 2008, 14, 142–153. [Google Scholar] [CrossRef]
- Hooker, T.D.; Stark, J.M. Soil C and N cycling in three semiarid vegetation types: Response to an in situ pulse of plant detritus. Soil Biol. Biochem. 2008, 40, 2678–2685. [Google Scholar] [CrossRef]
- Luo, X.B.; Liang, R.B.; Wang, Y.X. Response of soil microbial carbon and nitrogen pools to increasing atmospheric nitrogen deposition in forest surface soil. Ecol. Environ. Sci. 2014, 23, 365–370. [Google Scholar]
- Zhang, W.; Parker, K.M.; Luo, Y.; Wan, S.; Wallace, L.L.; Hu, S. Soil microbial responses to experimental warming and clipping in a tallgrass prairie. Glob. Chang. Biol. 2007, 11, 27–28. [Google Scholar] [CrossRef]
- Zhang, N.; Liu, W.; Yang, H.; Yu, X.; Gutknecht, J.L.M.; Zhang, Z.; Wan, S.; Ma, K. Soil microbial responses to warming and increased precipitation and their implications for ecosystem C cycling. Oecologia 2013, 173, 1125–1142. [Google Scholar] [CrossRef]
- Jian, S.; Li, J.; Chen, J.I.; Wang, G.; Mayes, M.A.; Dzantor, K.E.; Hui, D.; Luo, Y. Soil extracellular enzymc activities, soil carbon and nitrogen storage under nitrogen fertilization. A meta-analysis. Soil Biol. Biochem. 2016, 101, 32–43. [Google Scholar] [CrossRef]
- Chen, D.; Li, J.; Lan, Z.; Hu, S.; Bai, Y. Soil acidification exerts a greater control on soil respiration than soil nitrogen availability in grasslands subjected to long-term nitrogen enrichment. Funct. Ecol. 2016, 30, 658–669. [Google Scholar] [CrossRef]
- Li, H.; Yang, S.; Xu, Z.; Yan, Q.; Li, X.; van Nostrand, J.D.; He, Z.; Yao, F.; Han, X.; Zhou, J.; et al. Rsponses of soil microbial functional genes to global changes are indirectly influenced by aboveground plant biomass variation. Soil. Biol. Biochem. 2017, 104, 18–29. [Google Scholar] [CrossRef]
- Pietri JC, A.; Brookes, P.C. Relationships between soil pH and microbial properties in a UK arable soil. Soil Biol. Biochem. 2008, 40, 1856–1861. [Google Scholar] [CrossRef]
- Chen, Y.Q.; Chen, G.T.; Liang, Z.; Li, R.H.; Ma, H.Y.; Tu, L.H. Ten-year nitrogen addition did not significantly affect soil phosphorus fractions in a Pleioblastus amarus plantation. Ecol. Environ. Sci. 2018, 27, 677–684. [Google Scholar]
- Deng, Q.; Hui, D.; Dennis, S.; Reddy, K.C. Responses of terrestrial ecosystem phosphorus cycling to nitrogen addition: A meta-analysis. Glob. Ecol. Biogeogr. 2017, 26, 713–728. [Google Scholar] [CrossRef]
- Cheng, L.; Zhou, J.C.; Lin, K.M.; Zhang, Q.F.; Zhou, J.R.; Lin, Q.Y.; Zheng, W.; Wang, T.; Chen, Y.M. Effects of nitrogen addition on soil microbial community structure in a subtropical Phyllostachy pubescens. Chin. J. Ecol. 2020, 39, 1929–1937. [Google Scholar]
- Fan, Y.; Zhong, X.; Lin, F.; Liu, C.; Yang, L.; Wang, M.; Chen, G.; Chen, Y.; Yang, Y. Responses of soil phosphorus fractions after nitrogen addition in a subtropical forest ecosystem: Insights from decreased Fe and Al oxides and increased plant roots. Geoderma 2019, 337, 246–255. [Google Scholar] [CrossRef]
- Ahmed, W.; Liu, K.; Qaswar, M.; Huang, J.; Huang, Q.; Xu, Y.; Ali, S.; Mehmood, S.; Asghar, R.M.A.; Mahmood, M.; et al. Long-Term Mineral Fertilization Improved the Grain Yield and Phosphorus Use Efficiency by Changing Soil P Fractions in Ferralic Cambisol. Agronomy 2019, 9, 784. [Google Scholar] [CrossRef]
- Wan, W.; Li, X.; Han, S.; Wang, L.; Luo, X.; Chen, W.; Huang, Q. Soil aggregate fractionation and phosphorus fraction driven by long-term fertilization regimes affect the abundance and composition of P-cycling-related bacteria. Soil Tillage Res. 2020, 196, 104475. [Google Scholar] [CrossRef]
- Chen, C.; Condron, L.; Davis, M.; Sherlock, R. Seasonal changes in soil phosphorus and associated microbial properties under adjacent grassland and forest in New Zealand. For. Ecol. Manag. 2003, 177, 539–557. [Google Scholar] [CrossRef]
- Griffiths, B.S.; Spilles, A.; Bonkowski, M. C:N:P stoichiometry and nutrient limitation of the soil microbial biomass in a grazed grassland site under experimental P limitation or excess. Ecol. Process. 2012, 1, 6. [Google Scholar] [CrossRef]
- Foote, J.; Boutton, T.; Scott, D. Soil C and N storage and microbial biomass in US southern pine forests: Influence of forest management. For. Ecol. Manag. 2015, 355, 48–57. [Google Scholar] [CrossRef]
- Yang, J.; Yu, F.; Yu, Y.; Zhang, J.; Wang, R.; Srinivasulu, M.; Vasenev, V.I. Characterization, source apportionment, and risk assessment of polycyclic aromatic hydrocarbons in urban soil of Nanjing, China. J. Soils Sediments 2016, 17, 1116–1125. [Google Scholar] [CrossRef]
- Cao, C.; Jiang, S.; Ying, Z.; Zhang, F.; Han, X. Spatial variability of soil nutrients and microbiological properties after the establishment of leguminous shrub Caragana microphylla Lam. plantation on sand dune in the Horqin Sandy Land of Northeast China. Ecol. Eng. 2011, 37, 1467–1475. [Google Scholar] [CrossRef]
- Huang, G.; Li, Y.; Su, Y.G. Effects of increasing precipitation on soil microbial community composition and soil respiration in a temperate desert, Northwestern China. Soil Biol. Biochem. 2015, 83, 52–56. [Google Scholar] [CrossRef]
- Wu, J.P.; Han, X.H.; Xu, Y.D.; Ren, C.J.; Yang, G.H.; Ren, G.X. Ecological stoichiometry of soil end soil microbial biomass C, N, P under grain-to-green program in Loess Hilly region. Acta Agrestia Sin. 2016, 24, 783–792. [Google Scholar]
- Wei, S.Z.; Zhao, Q.; Liao, M.Q.; Zhou, S.X.; He, C.; Wang, L.; Huang, C. Effects of simulated nitrogen deposition on microbial biomass during litter decompositionin a natural evergreen broad-leaved forest in the rainy area of West China. Acta Ecol. Sin. 2018, 38, 8001–8007. [Google Scholar]
- Zhao, H.; Huang, G.; Li, Y.; Ma, J.; Sheng, J.; Jia, H.; Li, C. Effects of increased summer precipitation and nitrogen addition on root decomposition in a temperate desert. PLoS ONE 2015, 10, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Wang, S.; He, T.; Liu, L.; Wu, J. Response of organic carbon mineralization and microbial community to leaf litter and nutrient additions in subtropical forest soils. Soil Biol. Biochem. 2014, 71, 13–20. [Google Scholar] [CrossRef]
- Qiao, N.; Xu, X.; Hu, Y.; Blagodatskaya, E.; Liu, Y.; Schaefer, D.; Kuzyakov, Y. Carbon and nitrogen additions induce distinct priming effects along an organicmatter decay continuum. Sci. Rep. 2016, 6, 19865. [Google Scholar] [CrossRef]
- Micks, P.; Aber, J.D.; Boone, R.D.; Davidson, E.A. Short-term soil respiration and nitrogen immobilization response to nitrogen applications in control and nitrogen-enriched temperate forests. For. Ecol. Manag. 2004, 196, 57–70. [Google Scholar] [CrossRef]
- Zhu, Y.F.; Sun, Z.L.; Wang, Q.K. Effects of biochar and nitrogen additions on soil organic carbon decomposition and balance in a subtropical forest. Chin. J. Ecol. 2020, 39, 2851–2859. [Google Scholar]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).