
Carbon Assimilation, Biomass Partitioning and Productivity in Grasses


{kind=link}
Abstract
:1. Introduction

2. Carbon Fixation
3. Plant Morphology as a Partial Consequence of Photosynthetic N Requirement
4. Nutrient and Water Availability
5. Competition
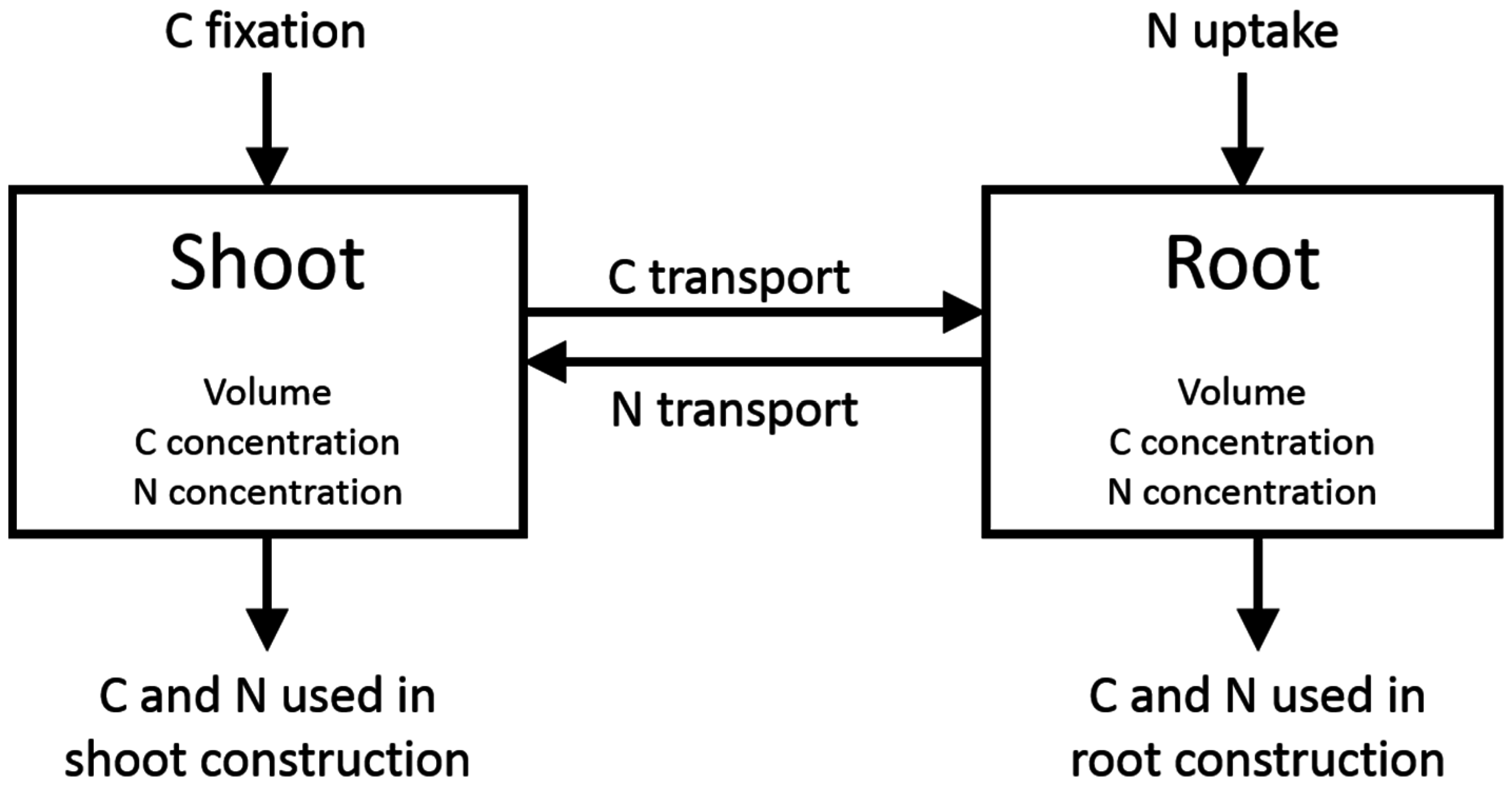
6. Carbon Allocation
7. Conclusions
Acknowledgements
Conflicts of Interest
References
- Kruger, E.L.; Volin, J.C. Reexamining the empirical relationship between plant growth and leaf photosynthesis. Funct. Plant Biol. 2006, 33, 421–429. [Google Scholar] [CrossRef]
- Wright, I.J.; Reich, P.B.; Westoby, M.; Ackerly, D.D.; Baruch, Z.; Bongers, F.; Cavender-Bares, J.; Chapin, T.; Cornelissen, J.H.C.; Diemer, M.; et al. The worldwide leaf economics spectrum. Nature 2004, 428, 821–827. [Google Scholar] [CrossRef] [PubMed]
- Poorter, H.; Niklas, K.J.; Reich, P.B.; Oleksyn, J.; Poot, P.; Mommer, L. Biomass allocation to leaves, stems and roots: Meta-analysis of interspecific variation and environmental control. New Phytol. 2012, 193, 30–50. [Google Scholar] [CrossRef] [PubMed]
- Raven, J.A.; Thomas, H. Grasses. Curr. Biol. 2010, 20, R837–R839. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Rehuel, D.; de Cauwer, B.; Cougnon, M. The role of forage crops in multifunctional agriculture. In Fodder Crops and Amenity Grasses; Boller, B., Posselt, U.K., Veronesi, F., Eds.; Springer-Verlag: New York, NY, USA, 2010; pp. 1–12. [Google Scholar]
- Fulkerson, W.J.; Donaghy, D.J. Plant-soluble carbohydrate reserves and senescence-key criteria for developing an effective grazing management system for ryegrass-based pastures: A review. Aust. J. Exp. Agric. 2001, 41, 261–275. [Google Scholar] [CrossRef]
- Robinson, D. Resource capture by localized root proliferation: Why do plants bother? Ann. Bot. 1996, 77, 179–185. [Google Scholar] [CrossRef]
- Thornley, J.H.M. A balanced quantitative model for root:shoot ratios in vegetative plants. Ann. Bot. 1972, 36, 431–441. [Google Scholar]
- Robinson, D. Compensatory changes in the partitioning of dry matter in relation to nitrogen uptake and optimal variations in growth. Ann. Bot. 1986, 58, 841–848. [Google Scholar]
- Hirose, T. A vegetative plant growth model: Adaptive significance of phenotypic plasticity in matter partitioning. Funct. Ecol. 1987, 1, 195–202. [Google Scholar] [CrossRef]
- Durand, J.-L.; Varlet-Grancher, C.; Lemaire, G.; Gastal, F.; Moulia, B. Carbon partitioning in forage crops. Acta Biotheor. 1991, 39, 213–224. [Google Scholar] [CrossRef]
- Sheehy, J.E.; Gastal, F.; Mitchell, P.L.; Durand, J.-L.; Lemaire, G.; Woodward, F.I. A nitrogen-led model of grass growth. Ann. Bot. 1996, 77, 165–177. [Google Scholar] [CrossRef]
- Lemaire, G.; Millard, P. An ecophysiological approach to modelling resource fluxes in competing plants. J. Exp. Bot. 1999, 50, 15–28. [Google Scholar] [CrossRef]
- Gastal, F.; Lemaire, G.; Durand, J.-L.; Louarn, G. Quantifying crop responses to nitrogen and avenues to improve nitrogen-use efficiency. In Crop Physiology, Applications for Genetic Improvement and Agronomy, 2nd ed.; Sadras, V.O., Calderini, D., Eds.; Academic Press/Elsevier: Oxford, UK, 2015; pp. 159–206. [Google Scholar]
- Amthor, J.S. The role of maintenance respiration in plant growth. Plant Cell Environ. 1984, 7, 561–569. [Google Scholar]
- Yamori, W.; Irving, L.J.; Adachi, S.; Busch, F.A. Strategies for optimizing photosynthesis with biotechnology to improve crop yield. Handb. Photosynth. 2016, in press. [Google Scholar]
- Galmés, J.; Kapralov, M.V.; Andralojc, P.J.; Conesa, M.À.; Keys, A.J.; Parry, M.A.J.; Flexas, J. Expanding knowledge of the Rubisco kinetics variability in plant species: Environmental and evolutionary trends. Plant Cell Environ. 2014, 37, 1989–2001. [Google Scholar] [CrossRef] [PubMed]
- Evans, J.R. Photosynthesis and nitrogen relationships in leaves of C3 plants. Oecologia 1989, 78, 9–19. [Google Scholar] [CrossRef]
- Makino, A.; Sato, T.; Nakano, H.; Mae, T. Leaf photosynthesis, plant growth and nitrogen allocation in rice under different irradiances. Planta 1997, 203, 390–398. [Google Scholar] [CrossRef]
- Parkinson, K.J.; Penman, H.L.; Tregunna, E.B. Growth of plants in different oxygen concentrations. J. Exp. Bot. 1974, 25, 132–145. [Google Scholar] [CrossRef]
- Yoshida, S. Effects of CO2 enrichment at different stages of panicle development on yield components and yield of rice (Oryza sativa L.). Soil Sci. Plant Nutr. 1973, 19, 311–316. [Google Scholar] [CrossRef]
- Myers, S.S.; Zanobetti, A.; Kloog, I.; Huybers, P.; Leakey, A.D.B.; Bloom, A.J.; Carlisle, E.; Dietterich, L.H.; Fitzgerald, G.; Hasegawa, T.; et al. Increasing CO2 threatens human nutrition. Nature 2014, 510, 139–142. [Google Scholar] [CrossRef] [PubMed]
- Bloom, A.J. The increasing importance of distinguishing among plant nitrogen sources. Curr. Opin. Plant Biol. 2015, 25, 10–16. [Google Scholar] [CrossRef] [PubMed]
- Bloom, A.J.; Asensio, J.S.R.; Randall, L.; Rachmilevitch, S.; Cousins, A.B.; Carlisle, E.A. CO2 enrichment inhibits shoot nitrate assimilation in C3 but not C4 plants and slows growth under nitrate in C3 plants. Ecology 2012, 93, 355–367. [Google Scholar] [CrossRef] [PubMed]
- Sicher, R.C.; Bunce, J.A. Relationship of photosynthetic acclimation to changes of Rubisco activity in field grown winter wheat and barley during growth in elevated carbon dioxide. Photosynth. Res. 1997, 52, 27–38. [Google Scholar] [CrossRef]
- Garcia, R.L.; Long, S.P.; Wall, G.W.; Osborne, C.P.; Kimball, B.A.; Nie, G.Y.; Pinter, P.J., Jr.; LaMorte, R.L.; Wechsung, F. Photosynthesis and conductance of spring-wheat leaves: Field response to continuous free-air atmospheric CO2 enrichment. Plant Cell Environ. 1998, 21, 659–669. [Google Scholar] [CrossRef]
- Ghannoum, O.; Evans, J.R.; Chow, W.S.; Andrews, T.J.; Conroy, J.P.; von Caemmerer, S. Faster Rubisco is the key to superior nitrogen use efficiency in NADP-Malic enzyme relative to NAD-Malic enzyme C4 grasses. Plant Physiol. 2005, 137, 638–650. [Google Scholar] [CrossRef] [PubMed]
- Sage, R.F.; Li, M.; Monson, R.K. The taxonomic distribution of C4 photosynthesis. In C4 Plant Biology; Sage, R.F., Monson, R.K., Eds.; Academic Press: San Diego, USA, 1999; pp. 551–584. [Google Scholar]
- Studer, R.A.; Christin, P.A.; Williams, M.A.; Orengo, C.A. Stability-activity tradeoffs constrain the adaptive evolution of RubisCO. Proc. Natl. Acad. Sci. USA 2014, 111, 2223–2228. [Google Scholar] [CrossRef] [PubMed]
- Von Caemmerer, S.; Quick, W.P.; Furbank, R.T. The development of C4 rice: Current progress and future challenges. Science 2012, 336, 1671–1672. [Google Scholar] [CrossRef] [PubMed]
- Lin, M.T.; Occhialini, A.; Andralojc, P.J.; Parry, M.A.J.; Hanson, M.R. A faster Rubisco with potential to increase photosynthesis in crops. Nature 2014, 513, 547–550. [Google Scholar] [CrossRef] [PubMed]
- Conley, M.M.; Kimball, B.A.; Brooks, T.J.; Pinter, P.J., Jr.; Hunsaker, D.J.; Wall, G.W.; Adam, N.R.; LaMorte, R.L.; Matthias, A.D.; Thompson, T.L.; et al. CO2 enrichment increases water-use efficiency in sorghum. New Phytol. 2001, 151, 407–412. [Google Scholar] [CrossRef]
- Sudo, E.; Suzuki, Y.; Makino, A. Whole plant growth and N utilization in transgenic rice plants with increased or decreased Rubisco content under different CO2 partial pressures. Plant Cell Physiol. 2014, 55, 1905–1911. [Google Scholar] [CrossRef] [PubMed]
- Cheng, S.H.; Moore, B.D.; Seemann, J.R. Effects of short and long-term elevated CO2 on the expression of ribulose-1,5-bisphosphate carboxylase/oxygenase genes and carbohydrate accumulation in leaves and Arabidopsis thaliana (L.) Heynh. Plant Physiol. 1998, 116, 715–723. [Google Scholar] [CrossRef] [PubMed]
- Moore, B.D.; Cheng, S.H.; Sims, D.; Seemann, J.R. The biochemical and molecular basis for photosynthetic acclimation to elevated atmospheric CO2. Plant Cell Environ. 1999, 22, 567–582. [Google Scholar] [CrossRef]
- Seneweera, S.; Makino, A.; Hirotsu, N.; Norton, R.; Suzuki, Y. New insight into the photosynthetic acclimation to elevated CO2: The role of leaf nitrogen and ribulose-1,5-bisphosphate carboxylase/oxygenase content in rice leaves. Environ. Exp. Bot. 2011, 71, 128–136. [Google Scholar] [CrossRef]
- Moore, B.D.; Cheng, S.H.; Rice, J.; Seemann, J.R. Sucrose cycling, Rubisco expression, and prediction of photosynthetic acclimation to elevated atmospheric CO2. Plant Cell Environ. 1998, 21, 905–915. [Google Scholar] [CrossRef]
- Hocking, P.J.; Meyer, C.P. Effects of CO2 enrichment and nitrogen stress on growth, and partitioning of dry matter and nitrogen in wheat and maize. Aust. J. Plant Physiol. 1991, 18, 339–356. [Google Scholar] [CrossRef]
- Gastal, F.; Lemaire, G. N uptake and distribution in crops: An agronomical and ecophysiological perspective. J. Exp. Bot. 2002, 53, 789–799. [Google Scholar] [CrossRef] [PubMed]
- Hikosaka, K.; Terashima, I. A model of the acclimation of photosynthesis in the leaves of C3 plants to sun and shade with respect to nitrogen use. Plant Cell Environ. 1995, 18, 605–618. [Google Scholar] [CrossRef]
- Anten, N.P.R.; Schieving, F.; Werger, M.J.A. Patterns of light and nitrogen distribution in relation to whole canopy carbon gain in C3 and C4 mono- and dicotyledonous species. Oecologia 1995, 101, 504–513. [Google Scholar] [CrossRef]
- Makino, A. Rubisco and nitrogen relations in rice: Leaf photosynthesis and plant growth. Soil Sci. Plant Nutr. 2003, 49, 319–327. [Google Scholar] [CrossRef]
- Mae, T.; Makino, A.; Ohira, A. Changes in the amounts of ribulose bisphosphate carboxylase synthesized and degraded during the life-span of rice life (Oryza sativa L.). Plant Physiol. 1983, 24, 1079–1086. [Google Scholar]
- Irving, L.J.; Robinson, D. A dynamic model of Rubisco turnover in cereal leaves. New Phytol. 2006, 169, 493–504. [Google Scholar] [CrossRef] [PubMed]
- Mae, T.; Ohira, K. The relationship of nitrogen related to leaf growth and senescence in rice plants (Oryza sativa L.). Plant Cell Physiol. 1981, 22, 1067–1074. [Google Scholar]
- Santos, P.M.; Thornton, B.; Corsi, M. Nitrogen dynamics in the intact grasses Poa trivialis and Panicum maximum receiving contrasting supplies of nitrogen. J. Exp. Bot. 2002, 53, 2167–2176. [Google Scholar] [CrossRef] [PubMed]
- Wada, S.; Hayashida, Y.; Izumi, M.; Kurusu, T.; Hanamata, S.; Kanno, K.; Kojima, S.; Yamaya, T.; Kuchitsu, K.; Makino, A.; et al. Autophagy Supports Biomass Production and Nitrogen Use Efficiency at the Vegetative Stage in Rice. Plant Physiol. 2015, 168, 60–73. [Google Scholar] [CrossRef] [PubMed]
- Khaembah, E.N.; Irving, L.J.; Thom, E.R.; Faville, M.J.; Easton, H.S.; Matthew, C. Leaf Rubisco turnover in a perennial ryegrass (Lolium perenne L.) mapping population: Genetic variation, identification of associated QTL, and correlation with plant morphology and yield. J. Exp. Bot. 2013, 64, 1305–1316. [Google Scholar] [CrossRef] [PubMed]
- Khaembah, E.N.; Gastal, F.; Carre, S.; Irving, L.J.; Barre, P.; Matthew, C. Morphology and Rubisco turnover characteristics of perennial ryegrass breeding populations after two and four cycles of divergent selection for long or short leaf length. Crop Pasture Sci. 2013, 64, 687–695. [Google Scholar] [CrossRef]
- Bélanger, G.; Gastal, F.; Lemaire, G. Growth analysis of a tall fescue sward fertilised with different rates of nitrogen. Crop Sci. 1992, 6, 1371–1376. [Google Scholar] [CrossRef]
- MacAdam, J.W.; Volenec, J.J.; Nelson, C.J. Effects of nitrogen on mesophyll cell division and epidermal cell elongation in tall fescue leaf blades. Plant Physiol. 1989, 89, 549–556. [Google Scholar] [CrossRef] [PubMed]
- Anten, N.P.R. Optimal photosynthetic characteristics of individual plants in vegetation stands and implications for species coexistence. Ann. Bot. 2005, 95, 495–506. [Google Scholar] [CrossRef] [PubMed]
- Anten, N.P.R. Evolutionarily stable leaf area production in plant populations. J. Theor. Biol. 2002, 217, 15–32. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Gastal, F.; Bélanger, G. The effects of nitrogen fertilization and the growing season on photosynthesis of field-grown tall fescue (Festuca arundinacea Shreb.) canopies. Ann. Bot. 1993, 72, 401–408. [Google Scholar] [CrossRef]
- Bélanger, G.; Gastal, F.; Warembourg, F.R. Carbon balance of tall fescue (Festuca arundinacea Shreb.): Effects of nitrogen fertilization and the growing season. Ann. Bot. 1994, 74, 653–659. [Google Scholar] [CrossRef]
- Gastal, F.; Bélanger, G.; Lemaire, G. A model of the leaf expansion rate of tall fescue in response to nitrogen and temperature. Ann. Bot. 1992, 70, 437–442. [Google Scholar]
- Anten, N.P.R.; Hirose, T. Biomass allocation and light partitioning among dominant and subordinate individuals in Xanthium canadense stands. Ann. Bot. 1998, 82, 665–673. [Google Scholar] [CrossRef] [Green Version]
- Poorter, H.; Evans, J.R. Photosynthetic nitrogen-use efficiency of species that differ inherently in specific leaf area. Oecologia 1998, 116, 26–37. [Google Scholar] [CrossRef]
- Andrews, M.; Raven, J.A.; Lea, P.J. Do plants need nitrate? The mechanisms by which nitrogen form affects plants. Ann. Appl. Biol. 2013, 163, 174–199. [Google Scholar] [CrossRef]
- Lemaire, G.; van Oosterom, E.; Sheehy, J.; Jeuffroy, M.H.; Massignam, A.; Rossato, L. Is crop N demand more closely related to dry matter accumulation or leaf area expansion during vegetative growth? Field Crops Res. 2007, 100, 91–106. [Google Scholar] [CrossRef]
- Hikosaka, K. Leaf canopy as a dynamic system: Ecophysiology and optimality in leaf turnover. Ann. Bot. 2005, 95, 521–533. [Google Scholar] [CrossRef] [PubMed]
- Thomas, H.; Davies, A. Effect of shading on the regrowth of Lolium perenne swards in the field. Ann. Bot. 1977, 42, 705–715. [Google Scholar] [CrossRef]
- Thomas, H.; Sadras, V.O. The capture and gratuitous disposal of resources by plants. Funct. Ecol. 2001, 15, 3–12. [Google Scholar]
- Lambers, H.; Simpson, R.J.; Beilharz, V.C.; Dalling, M.J. Translocation and utilization of carbon in wheat (Triticum aestivum). Physiol. Plant. 1982, 56, 18–22. [Google Scholar] [CrossRef]
- Scheible, W.R.; Lauerer, M.; Schulze, E.D.; Caboche, M.; Stitt, M. Accumulation of nitrate in the shoot acts as a signal to regulate shoot–root allocation in tobacco. Plant J. 1997, 11, 671–691. [Google Scholar] [CrossRef]
- Miller, A.J.; Fan, X.; Orsel, M.; Smith, S.J.; Wells, D.M. Nitrate transport and signalling. J. Exp. Bot. 2007, 58, 2297–2306. [Google Scholar] [CrossRef] [PubMed]
- Andrews, M.; Morton, J.D.; Lieffering, M.; Bisset, L. The partitioning of nitrate assimilation between root and shoot of a range of temperate cereals and pasture grasses. Ann. Bot. 1992, 70, 271–276. [Google Scholar]
- Laperche, A.; Devienne-Barret, F.; Maury, O.; le Gouis, J.; Ney, B. A simplified conceptual model of carbon/nitrogen functioning for QTL analysis of winter wheat adaptation to nitrogen deficiency. Theor. Appl. Genet. 2006, 113, 1131–1146. [Google Scholar] [CrossRef] [PubMed]
- Drew, M.C. Comparison of the effects of a localized supply of phosphate, nitrate, ammonium and potassium on the growth of the seminal root system, and the shoot, in barley. New Phytol. 1975, 75, 479–490. [Google Scholar] [CrossRef]
- Zhang, H.; Jennings, A.; Barlow, P.W.; Forde, B.G. Dual pathways for regulation of root branching by nitrate. Proc. Natl. Acad. Sci. USA 1999, 96, 6529–6534. [Google Scholar] [CrossRef] [PubMed]
- Lima, J.E.; Kojima, S.; Takahashi, H.; von Wirén, N. Ammonium triggers lateral root branching in Arabidopsis in AMMONIUM TRANSPORTER1;3-dependent manner. Plant Cell 2010, 22, 3621–3633. [Google Scholar] [CrossRef] [PubMed]
- Raven, J.A.; Wollenweber, B.; Handley, L.L. A comparison of ammonium and nitrate as nitrogen sources for photolithotrophs. New Phytol. 1992, 121, 19–32. [Google Scholar] [CrossRef]
- Zerihun, A.; McKenzie, B.A.; Morton, J.D. Photosynthate costs associated with the utilization of different nitrogen-forms: Influence on the carbon balance of plants and shoot-root biomass partitioning. New Phytol. 1998, 138, 1–11. [Google Scholar] [CrossRef]
- Scheurwater, I.; Cornelissen, C.; Dictus, F.; Welschien, R.; Lambers, H. Why do fast- and slow-growing grass species differ so little in their rate of root respiration, considering the large differences in the rate of growth and ion uptake? Plant Cell Environ. 1998, 21, 995–1005. [Google Scholar] [CrossRef]
- Andrews, M.; Raven, J.A.; Lea, P.J.; Sprent, J.I. A role for shoot protein in shoot—Root dry matter allocation in higher plants. Ann. Bot. 2006, 97, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Irving, L.J.; Cameron, D.D. You are what you eat: Interactions between root parasitic plants and their hosts. Adv. Bot. Res. 2009, 50, 87–138. [Google Scholar]
- Huang, B.; Fu, J. Photosynthesis, respiration, and carbon allocation of two cool-season perennial grasses in response to surface soil drying. Plant Soil 2000, 227, 17–26. [Google Scholar] [CrossRef]
- Hare, P.D.; Cress, W.A.; van Staden, J. Dissecting the roles of osmolyte accumulation during stress. Plant Cell Environ. 1998, 21, 535–553. [Google Scholar] [CrossRef]
- Araya, T.; Noguchi, K.; Terashima, I. Effect of carbohydrate accumulation on photosynthesis differ between sink and source leaves of Phaseolus vulgaris L. Plant Cell Physiol. 2006, 47, 644–652. [Google Scholar] [CrossRef] [PubMed]
- Hamblin, A.; Tennant, D. Root length density and water uptake in cereals and grain legumes: How well are they correlated? Aust. J. Agric. Res. 1987, 38, 513–527. [Google Scholar] [CrossRef]
- Saarinen, T.; Haansuu, P. Shoot density of Carex rostrate Stokes in relation to internal carbon:nitrogen balance. Oecologia 2000, 122, 29–35. [Google Scholar] [CrossRef]
- Davies, A. Changes in growth rate and morphology of perennial ryegrass swards at high and low nitrogen levels. J. Agric. Sci. 1971, 77, 123–134. [Google Scholar] [CrossRef]
- Gautier, H.; Varlet-Grancher, C.; Hazard, L. Tillering responses to the light environment and to defoliation in populations of perennial ryegrass (Lolium perenne L.) selected for contrasting leaf length. Ann. Bot. 1999, 83, 423–429. [Google Scholar] [CrossRef]
- Smith, H. Phytochromes and light signal perception by plants—An emerging synthesis. Nature 2000, 407, 585–591. [Google Scholar] [CrossRef] [PubMed]
- Toyota, M.; Tatewaki, M.; Morokuma, M.; Kusutani, A. Tillering responses to high red/far red ratio of four Japanese wheat cultivars. Plant Prod. Sci. 2014, 17, 124–130. [Google Scholar] [CrossRef]
- Evers, J.B.; Vos, J.; Andrieu, B.; Struik, P.C. Cessation of tillering in spring wheat in relation to light interception and red: Far-red ratio. Ann. Bot. 2006, 97, 649–658. [Google Scholar] [CrossRef] [PubMed]
- Sparkes, D.L.; Holme, S.J.; Gaju, O. Does light quality initiate tiller death in wheat? Eur. J. Agron. 2006, 24, 212–217. [Google Scholar] [CrossRef]
- Simons, R.G.; Davies, A.; Troughton, A. The effect of spacing on the growth of two genotypes of perennial ryegrass. J. Agric. Res. 1973, 80, 495–502. [Google Scholar] [CrossRef]
- Lonsdale, W.M.; Watkinson, A.R. Light and self-thinning. New Phytol. 1982, 90, 431–445. [Google Scholar] [CrossRef]
- Lonsdale, W.M. The self-thinning rule: Dead or alive? Ecology 1990, 71, 1373–1388. [Google Scholar] [CrossRef]
- Enquist, B.J.; Brown, J.H.; West, G.B. Allometric scaling of plant energetics and population density. Nature 1998, 395, 163–165. [Google Scholar] [CrossRef]
- Matthew, C.; Lemaire, G.; Sackville-Hamilton, N.R.; Hernandez-Garay, A. A modified self-thinning equation to describe size/density relationships for defoliated swards. Ann. Bot. 1995, 76, 579–587. [Google Scholar] [CrossRef]
- Davies, A.; Thomas, H. Rates of leaf and tiller production in young spaced perennial ryegrass plants in relation to soil temperature and solar radiation. Ann. Bot. 1983, 57, 591–597. [Google Scholar]
- Minchin, P.E.H.; Lacointe, A. New understanding on phloem physiology and possible consequences for modelling long-distance carbon transport. New Phytol. 2005, 166, 771–779. [Google Scholar] [CrossRef] [PubMed]
- Matthew, C.; Kemball, W.D. Allocation of carbon-14 to roots of different ages in perennial ryegrass (Lolium perenne L.). In Proceedings of the XVIII International Grassland Congress, Calgary: Association Management Center, Section 7, Calgary, Canada, 8–19 June, 1997; pp. 1–2.
- Nikinmaa, E.; Hölttä, T.; Hari, P.; Kolari, P.; Mäkelä, A.; Sevanto, S.; Vesala, T. Assimilate transport in phloem sets conditions for leaf gas exchange. Plant Cell Environ. 2013, 36, 655–669. [Google Scholar] [CrossRef] [PubMed]
- Lattanzi, F.A.; Schnyder, H.; Thornton, B. The sources of carbon and nitrogen supplying leaf growth. Assessment of the role of stores with compartmental models. Plant Physiol. 2005, 137, 383–395. [Google Scholar] [CrossRef] [PubMed]
- Matsuhashi, S.; Fujimaki, S.; Kawachi, N.; Sakamoto, K.; Ishioka, N.S.; Kume, T. Quantitative modeling of photoassimilate flow in an intact plant using the positron emitting tracer imaging system (PETIS). Soil Sci. Plant Nutr. 2005, 51, 417–423. [Google Scholar] [CrossRef]
- Thornton, B.; Paterson, E.; Midwood, A.J.; Sim, A.; Pratt, S.M. Contribution of current carbon assimilation in supplying root exudates of Lolium perenne measured using steady-state 13C labelling. Physiol. Plant. 2004, 120, 434–441. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, D.D.; Irving, L.J.; Carnevalli, R.A.; Hodgson, J.; Matthew, C. Distribution of current photosynthate in two Guinea grass (Panicum maximum Jacq.) cultivars. J. Exp. Bot. 2006, 57, 2015–2024. [Google Scholar] [CrossRef] [PubMed]
- Osaki, M.; Shinano, T.; Yamada, M.; Yamada, S. Function of node unit in photosynthate distribution to root in higher plants. Photosynthetica 2004, 42, 123–131. [Google Scholar] [CrossRef]
- Osaki, M.; Morikawa, K.; Matsumoto, M.; Shinano, T.; Iyoda, M.; Tadano, T. Productivity of high yielding crops. III. Accumulation of ribulose-1,5-bisphosphate carboxylase/oxygenase and chlorophyll in relation to productivity of high yielding crops. Soil Sci. Plant Nutr. 1993, 39, 399–408. [Google Scholar] [CrossRef]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Irving, L.J. Carbon Assimilation, Biomass Partitioning and Productivity in Grasses. Agriculture 2015, 5, 1116-1134. https://doi.org/10.3390/agriculture5041116
Irving LJ. Carbon Assimilation, Biomass Partitioning and Productivity in Grasses. Agriculture. 2015; 5(4):1116-1134. https://doi.org/10.3390/agriculture5041116
Chicago/Turabian StyleIrving, Louis J. 2015. "Carbon Assimilation, Biomass Partitioning and Productivity in Grasses" Agriculture 5, no. 4: 1116-1134. https://doi.org/10.3390/agriculture5041116
APA StyleIrving, L. J. (2015). Carbon Assimilation, Biomass Partitioning and Productivity in Grasses. Agriculture, 5(4), 1116-1134. https://doi.org/10.3390/agriculture5041116