Nitrogen Use Efficiency and the Genetic Variation of Maize Expired Plant Variety Protection Germplasm


Abstract
:1. Introduction
2. Materials and Methods
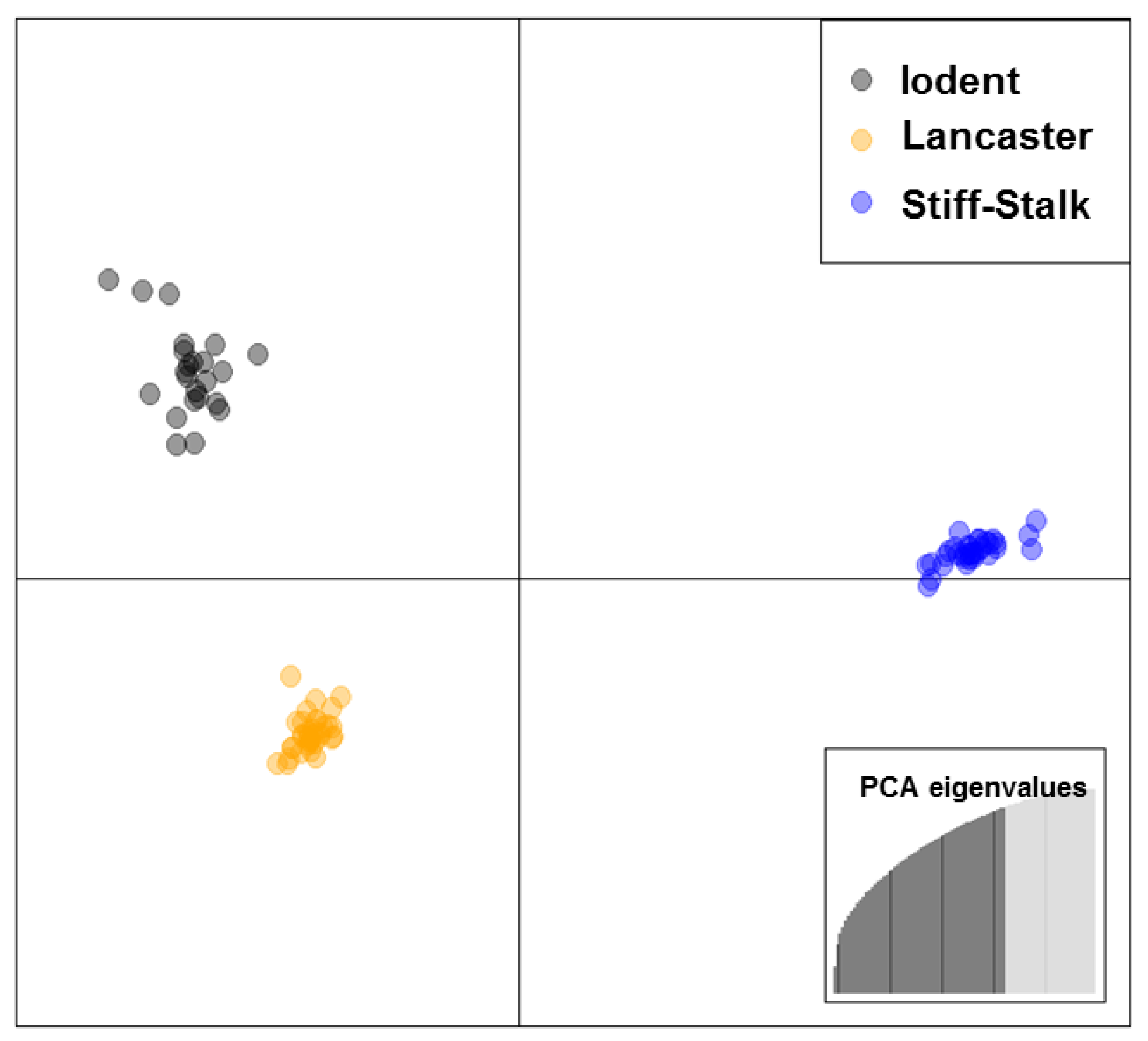
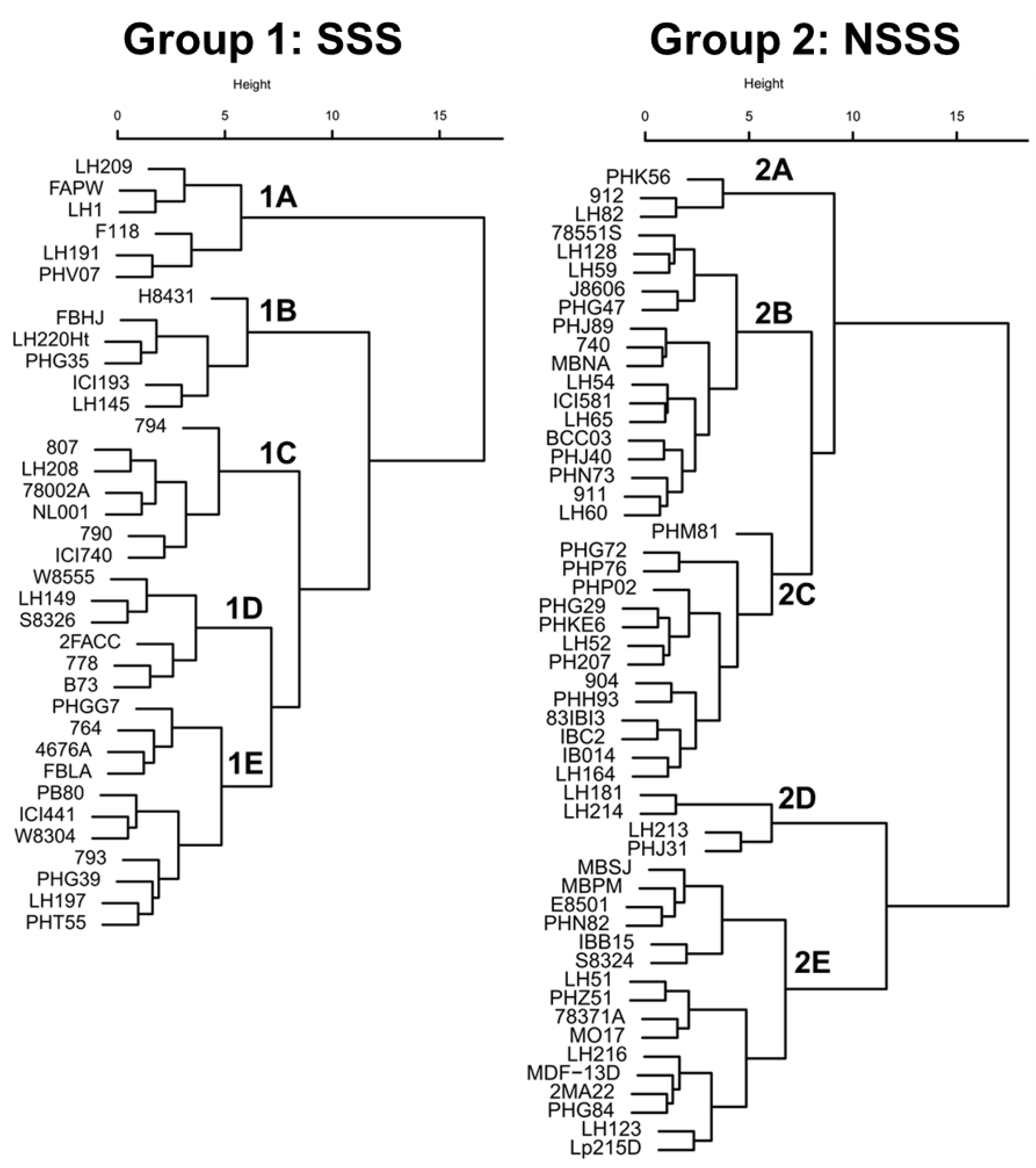
2.1. Germplasm and Genomic Data
2.2. Research Sites and Crop Management
2.3. Experimental Treatments and Design
2.4. Phenotype Measurements
2.5. Statistical Analysis
3. Results and Discussion
3.1. Phenotypic Variation of N-Use Traits
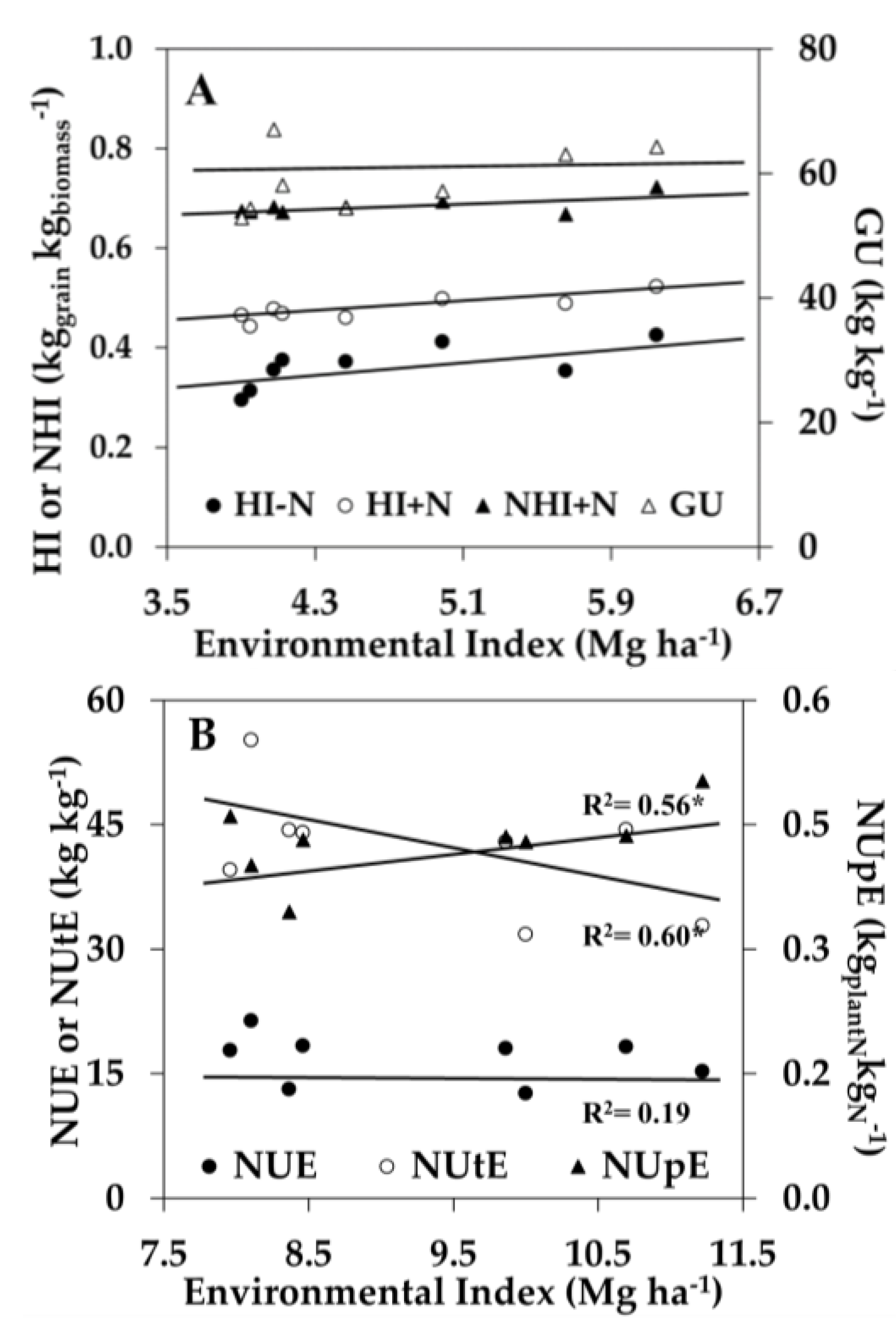
3.2. Genotype × Environment Interaction of N-Use Traits
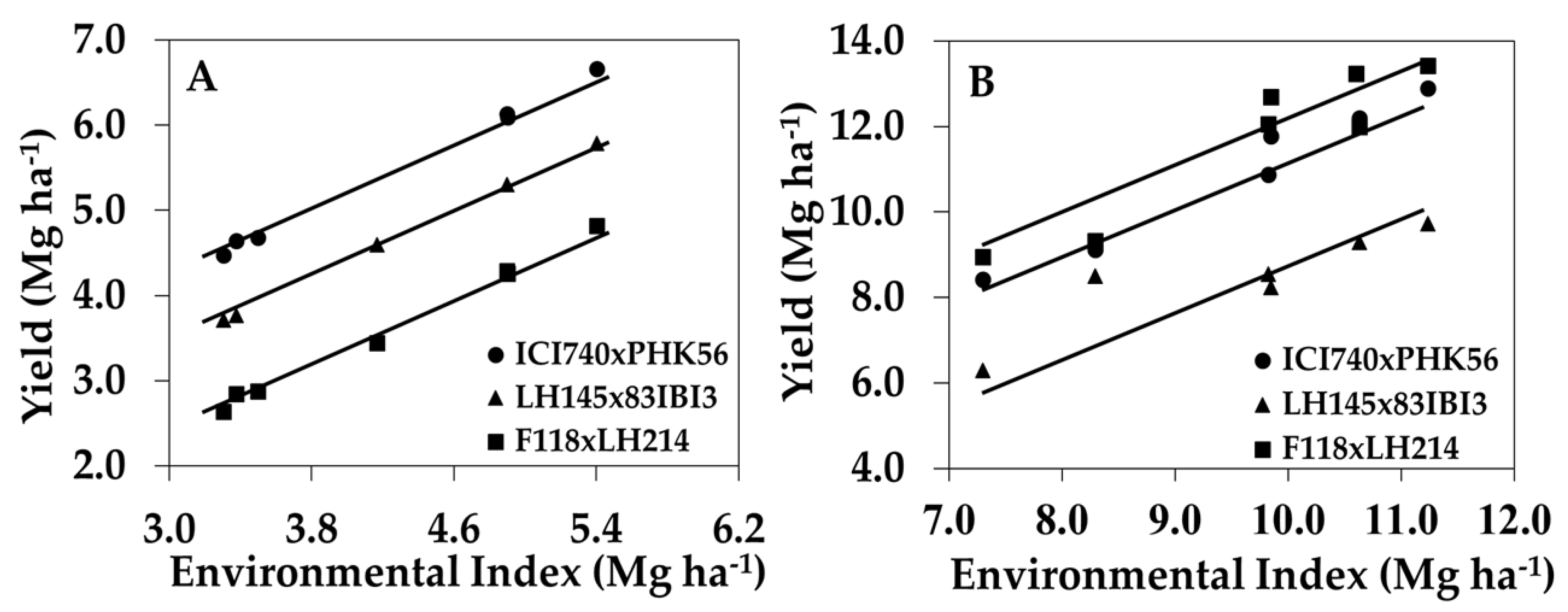
3.3. Identification of Maize Genotypes with Improved NUE
4. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- FAOSTAT. Production. Food and Agricultural Organization of the United Nations, Statistics Division. 2014. Available online: http://faostat3.fao.org/home/E (accessed on 13 October 2017).
- USDA-NASS (USDA National Agricultural Statistics Service). National Statistics for Corn: Corn Grain—Yield, Measured in Bu/acre. Statistics by Subject; USDA-NASS: Washington, DC, USA, 2014. Available online: http://www.nass.usda.gov/Statistics_by_Subject/index.php (accessed on 14 February 2016).
- Bender, R.R.; Haegele, J.W.; Ruffo, M.L.; Below, F.E. Nutrient uptake, partitioning, and remobilization in modern, transgenic insect-protected maize hybrids. Agron. J. 2013, 105, 161–170. [Google Scholar] [CrossRef]
- Duvick, D.N. The contribution of breeding to yield advances in maize (Zea mays L.). Adv. Agron. 2005, 86, 83–145. [Google Scholar]
- Kihara, J.; Nziguheba, G.; Zingore, S.; Coulibaly, A.; Esilaba, A.; Kabambe, V.; Njoroge, S.; Palm, C.; Huising, J. Understanding variability in crop response to fertilizer and amendments in sub-Saharan Africa. Agric. Ecosyst. Environ. 2016, 229, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Cassman, K.G.; Dobermann, A.; Walters, D.T. Agroecosystems, nitrogen-use efficiency, and nitrogen management. Ambio 2002, 31, 132–140. [Google Scholar] [CrossRef] [PubMed]
- Raun, W.R.; Johnson, G.V. Improving nitrogen use efficiency for cereal production. Agron. J. 1999, 91, 357–363. [Google Scholar] [CrossRef]
- Moll, R.H.; Kamprath, E.J.; Jackson, W.A. Analysis and interpretation of factors which contribute to efficiency of nitrogen utilization. Agron. J. 1982, 74, 562–564. [Google Scholar] [CrossRef]
- Moose, S.P.; Below, F.E. Biotechnology approaches to improving maize nitrogen use efficiency. In Molecular Genetic Approaches to Maize Improvement. Biotechnology in Agriculture and Forestry Series; Kriz, A.L., Larkins, B.A., Eds.; Springer: Berlin/Heidelberg, Germany, 2009; Volume 63, pp. 65–77. ISBN 978-3-540-68919-5. [Google Scholar]
- Uribelarrea, M.; Crafts-Brandner, S.J.; Below, F.E. Physiological N response of field-grown maize hybrids (Zea mays L.) with divergent yield potential and grain protein concentration. Plant Soil 2009, 316, 151–160. [Google Scholar] [CrossRef]
- Seebauer, J.R.; Moose, S.P.; Fabbri, B.J.; Crossland, L.D.; Below, F.E. Amino acid metabolism in maize earshoots. Implications for assimilate preconditioning and nitrogen signaling. Plant Physiol. 2004, 136, 4326–4334. [Google Scholar] [CrossRef] [PubMed]
- Hirel, B.; Bertin, P.; Quillere, I.; Bourdoncle, W.; Attagnant, C.; Dellay, C.; Gouy, A.; Cadiou, S.; Retaillau, C.; Falque, M.; et al. Towards a better understanding of the genetic and physiological basis for nitrogen use efficiency in maize. Plant Physiol. 2001, 125, 1258–1270. [Google Scholar] [CrossRef] [PubMed]
- Hammer, G.L.; Dong, Z.; McLean, G.; Doherty, A.; Messina, C.; Schussler, J.; Zinselmeier, C.; Paskiewicz, S.; Cooper, M. Can changes in canopy and/or root system architecture explain historical maize yield trends in the U.S. Corn Belt? Crop Sci. 2009, 49, 299–312. [Google Scholar] [CrossRef]
- Leach, K.A.; Tran, T.M.; Slewinski, T.L.; Meeley, R.B.; Braun, D.M. Sucrose transporter2 contributes to maize growth, development, and crop yield. J. Integr. Plant Biol. 2017, 59, 390–408. [Google Scholar] [CrossRef] [PubMed]
- Peng, Y.; Li, C.; Fritschi, F.B. Diurnal dynamics of maize leaf photosynthesis and carbohydrate concentrations in response to differential N availability. Environ. Exp. Bot. 2014, 99, 18–27. [Google Scholar] [CrossRef]
- Bihmidine, S.; Hunter, C.T., III; Johns, C.E.; Koch, K.E.; Braun, D.M. Regulation of assimilate import into sink organs: Update on molecular drivers of sink strength. Front. Plant Sci. 2013, 4, 177. [Google Scholar] [CrossRef] [PubMed]
- Weiland, R.T.; Ta, T.C. Allocation and retranslocation of N-15 by maize (Zea mays L.) hybrids under field conditions of low and high-N fertility. Aust. J. Plant Physiol. 1992, 19, 77–88. [Google Scholar] [CrossRef]
- Kamprath, E.J.; Moll, R.H.; Rodriguez, N. Effects of nitrogen fertilization and recurrent selection on performance of hybrid populations of corn. Agron. J. 1982, 74, 955–958. [Google Scholar] [CrossRef]
- Uribelarrea, M.; Moose, S.P.; Below, F.E. Divergent selection for grain protein affects nitrogen use in maize hybrids. Field Crop. Res. 2007, 100, 82–90. [Google Scholar] [CrossRef]
- Haegele, J.W.; Cook, K.A.; Nichols, D.M.; Below, F.E. Changes in nitrogen use traits associated with genetic improvement for grain yield of maize hybrids released in different decades. Crop Sci. 2013, 53, 1256–1268. [Google Scholar] [CrossRef]
- Tollenaar, M.; Nissanka, S.P.; Rajcan, I.; Bruulsema, T.W. Yield response of old and new corn hybrids to nitrogen. Better Crops. 1997, 81, 3–5. [Google Scholar]
- Presterl, T.; Groh, S.; Landbeck, M.; Seitz, G.; Schmidt, W.; Geiger, H.H. Nitrogen uptake and utilization efficiency of European maize hybrids developed under conditions of low and high nitrogen input. Plant Breed. 2002, 121, 480–486. [Google Scholar] [CrossRef]
- Gallais, A.; Coque, M. Genetic variation and selection for nitrogen use efficiency in maize: A synthesis. Maydica 2005, 50, 531–537. [Google Scholar]
- Laftte, H.R.; Edmeades, G.O. Stress tolerance in tropical maize is linked to constitutive changes in ear growth characteristics. Crop Sci. 1995, 35, 820–826. [Google Scholar] [CrossRef]
- Mikel, M.A.; Dudley, J.W. Evolution of North American dent corn from public to proprietary germplasm. Crop Sci. 2006, 46, 1193–1205. [Google Scholar] [CrossRef]
- Nelson, P.T.; Coles, N.D.; Holland, J.B.; Bubeck, D.M.; Smith, S.; Goodman, M.M. Molecular characterization of maize inbreds with expired US plant variety protection. Crop Sci. 2008, 48, 1673–1685. [Google Scholar] [CrossRef]
- Hauck, A.L.; Johnson, G.R.; Mikel, M.A.; Mahone, G.S.; Morales, A.J.; Rocheford, T.R.; Bohn, M.O. Generation means analysis of elite ex-plant variety protection commercial inbreds: A new public maize genetics resource. Crop Sci. 2014, 54, 174–189. [Google Scholar] [CrossRef]
- Bari, M.A.A.; Carena, M.J. Can expired proprietary maize (Zea mays L.) industry lines be useful for short-season breeding programs? II. Agronomic traits. Euphytica 2016, 207, 69–77. [Google Scholar] [CrossRef]
- Smith, J.S.C.; Duvick, D.N.; Smith, O.S.; Cooper, M.; Feng, L. Changes in pedigree backgrounds of Pioneer brand maize hybrids widely grown from 1930 to 1999. Crop Sci. 2004, 44, 1935–1946. [Google Scholar] [CrossRef]
- Doyle, J.J.; Doyle, J.L. A rapid DNA isolation procedure for small quantities of fresh leaf tissue. Phytochem. Bull. 1987, 19, 11–15. [Google Scholar]
- Elshire, R.J.; Glaubitz, J.C.; Sun, Q.; Poland, J.A.; Kawamoto, K.; Buckler, E.S.; Mitchell, S.E. A robust, simple genotyping-by-sequencing (GBS) approach for high diversity species. PLoS ONE 2011, 6. [Google Scholar] [CrossRef] [PubMed]
- Bradbury, P.J.; Zhang, Z.; Kroon, D.E.; Casstevens, T.M.; Ramdoss, Y.; Buckler, E.S. TASSEL: Software for association mapping of complex traits in diverse samples. Bioinformatics 2007, 23, 2633–2635. [Google Scholar] [CrossRef] [PubMed]
- Jombart, T.; Devillard, S.; Balloux, F. Discriminant analysis of principal components: A new method for the analysis of genetically structured populations. BMC Genet. 2010, 11, 94. [Google Scholar] [CrossRef] [PubMed]
- The R Project for Statistical Computing. Available online: http://www.r-project.org (accessed on 10 December 2016).
- Bernardo, R. Molecular markers and selection for complex traits in plants: Learning from the last 20 years. Crop Sci. 2008, 48, 1649–1664. [Google Scholar] [CrossRef]
- Ritchie, S.W.; Hanway, J.J.; Benson, G.O. How a Corn Plant Develops; Spec. Rep. No. 48; Iowa State University of Science and Technology Cooperative Extension Service: Ames, IA, USA, 1997. [Google Scholar]
- Betrán, F.J.; Beck, D.; Bänziger, M.; Edmeades, G.O. Genetic analysis of inbred and hybrid grain yield under stress and nonstress environments in tropical maize. Crop Sci. 2003, 43, 807–817. [Google Scholar] [CrossRef]
- Stuber, C.W.; Cockerham, C.C. Gene effects and variances in hybrid populations. Genetics 1966, 54, 1279–1286. [Google Scholar] [PubMed]
- SAS Institute. SAS 9.2 User’s Guide; SAS Institute: Cary, NC, USA, 2013. [Google Scholar]
- Holland, J.B.; Nyquist, W.E.; Cervantes-Martinez, C.T. Estimating and interpreting heritability for plant breeding: An update. Plant Breed. Rev. 2003, 22, 9–112. [Google Scholar]
- Falconer, D.S.; Mackay, T.F. Introduction to Quantitative Genetics, 4th ed.; Longman Technical: Essex, UK, 1996. [Google Scholar]
- Gilmour, A.; Cullis, B.; Welham, S.; Gogel, B.; Thompson, R. An efficient computing strategy for prediction in mixed linear models. Comput. Stat. Data Anal. 2004, 44, 571–586. [Google Scholar] [CrossRef]
- Brun, E.L.; Dudley, J.W. Nitrogen responses in the USA and Argentina of corn populations with different proportions of flint and dent germplasm. Crop Sci. 1989, 29, 565–569. [Google Scholar] [CrossRef]
- Trachsel, S.; Leyva, M.; Lopez, M.; Suarez, E.A.; Mendoza, A.; Montiel, N.G.; Macias, M.S.; Burgueno, J.; San Vicente, F. Identification of tropical maize germplasm with tolerance to drought, nitrogen deficiency, and combined heat and drought stresses. Crop Sci. 2016, 56, 3031–3045. [Google Scholar] [CrossRef]
- Bänziger, M.; Betrán, F.J.; Lafitte, H.R. Efficiency of high-nitrogen selection environments for improving maize for low-nitrogen target environments. Crop Sci. 1997, 37, 1110–1117. [Google Scholar] [CrossRef]
- Abe, A.; Menkir, A.; Moose, S.P.; Adetimirin, V.O.; Olaniyan, A.B. Genetic variation for nitrogen-use efficiency among selected tropical maize hybrids differing in grain yield potential. J. Crop Improv. 2012, 27, 31–52. [Google Scholar] [CrossRef]
- Duvick, D.N.; Cassman, K.G. Post-green revolution trends in yield potential of temperate maize in the northcentral United States. Crop Sci. 1999, 39, 1622–1630. [Google Scholar] [CrossRef]
- Van Eeuwijk, F.A.; Bustos-Korts, D.V.; Malosetti, M. What should students in plant breeding know about the statistical aspects of genotype × environment Interactions? Crop Sci. 2016, 56, 2119–2140. [Google Scholar] [CrossRef]
- Smiciklas, K.D.; Below, F.E. Influence of heterotic pattern on nitrogen use and yield of maize. Maydica 1990, 35, 209–213. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
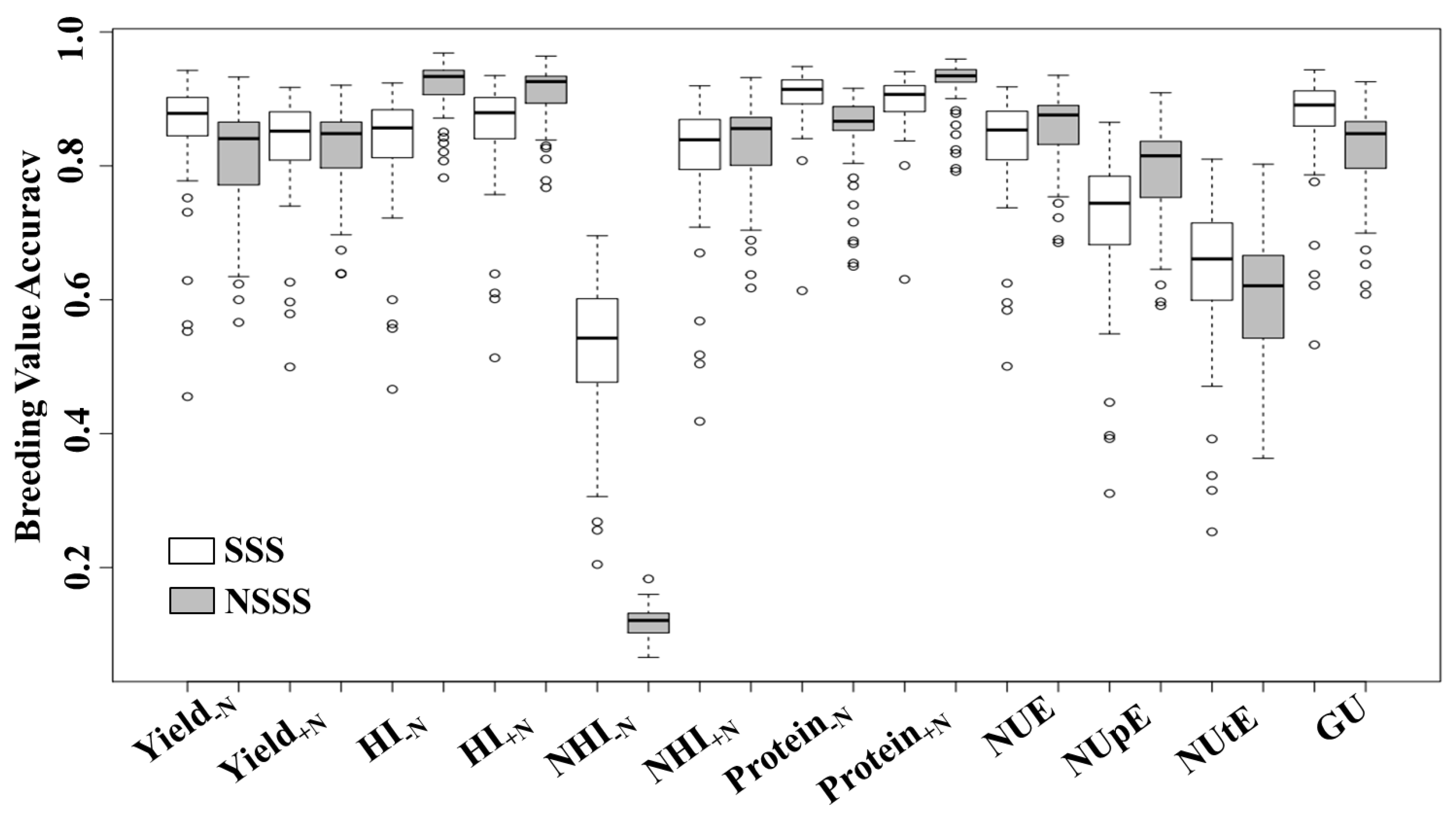{kind=link}
| GCASSS † | GCANSSS | SCA | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Trait | Mean ± SE ‡ | Range Min./Max. | σ2SSS | Range Min./Max. | σ2NSSS | Range Min./Max. | σ2SCA | σ2E | σ2SCA×E | σ2R | H2 |
| Yield−N | 4.9 ± 0.19 | −0.7/0.7 | 0.13 | −0.8/0.5 | 0.12 | - | 0.00 | 0.70 | 0.01 | 1.31 | 0.31 |
| Yield+N | 9.1 ± 0.28 | −0.9/+0.9 | 0.25 | −1.2/0.9 | 0.32 | −0.2/+0.3 | 0.06 | 2.12 | 0.36 | 1.07 | 0.61 |
| HI−N | 0.36 ± 0.01 | −0.05/+0.09 | 6 × 10−4 | −0.10/+0.06 | 1 × 10−3 | −0.01/+0.01 | 1 × 10−4 | 2 × 10−3 | 3 × 10−4 | 4 × 10−3 | 0.63 |
| HI+N | 0.47 ± 0.01 | −0.02/+0.03 | 2 × 10−4 | −0.06/+0.03 | 4 × 10−4 | −0.01/+0.01 | 5 × 10−5 | 6 × 10−4 | 1 × 10−5 | 1 × 10−3 | 0.73 |
| NHI−N | 0.56 ± 0.01 | −0.01/+0.01 | 2 × 10−4 | −0.01/+0.01 | 1 × 10−5 | −0.03/+0.02 | 4 × 10−4 | 9 × 10−3 | 2 × 10−3 | 8 × 10−3 | 0.11 |
| NHI+N | 0.68 ± 0.01 | −0.04/+0.02 | 3 × 10−4 | −0.05/+0.03 | 5 × 10−4 | −0.02/+0.01 | 1 × 10−4 | 3 × 10−4 | 1 × 10−4 | 3 × 10−3 | 0.44 |
| Protein−N | 62 ± 1.3 | −6.1/+5.9 | 0.8 | −5.2/+5.4 | 0.8 | −3.2/+2.3 | 0.4 | 1.7 | 1.3 | 2.1 | 0.74 |
| Protein+N | 85 ± 1.3 | −3.8/+4.9 | 0.7 | −6.0/+4.6 | 1.3 | −2.5/+2.5 | 0.2 | 5.5 | 0.5 | 3.0 | 0.77 |
| NUE | 16.7 ± 1.14 | −3.6/+3.9 | 3.81 | −5.7/+5.2 | 5.50 | −0.95/+1.20 | 0.80 | 10.13 | 4.97 | 18.56 | 0.60 |
| NUpE | 0.43 ± 0.03 | −0.03/+0.05 | 6 × 10−4 | −0.08/+0.09 | 1 × 10−3 | −0.01/+0.42 | 2 × 10−4 | 2 × 10−3 | 1 × 10−3 | 0.01 | 0.27 |
| NUtE | 41.8 ± 1.79 | −3.4/+2.9 | 5.5 | −2.7/+4.2 | 5.2 | −0.58/+0.60 | 1.3 | 59.3 | 7.8 | 201.7 | 0.11 |
| GU | 59.0 ± 2.2 | −7.8/+8.9 | 17.8 | −9.9/+7.4 | 16.0 | −3.2/+2.9 | 5.5 | 29.1 | 7.8 | 88.7 | 0.58 |
| Yield−N | Yield+N | HI−N | HI+N | NHI−N | NHI+N | Protein−N | Protein+N | NUE | NUpE | NUtE | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Yield+N | 0.31 ** | - | - | - | - | - | - | - | - | - | - |
| HI−N | 0.63 *** | −0.33 ** | - | - | - | - | - | - | - | - | - |
| HI+N | 0.49 *** | NS | 0.77 *** | - | - | - | - | - | - | - | - |
| NHI−N | NS | NS | NS | NS | - | - | - | - | - | - | - |
| NHI+N | 0.51 *** | NS | 0.65 *** | 0.78 *** | NS | - | - | - | - | - | - |
| Protein−N | −0.38 *** | −0.22 * | NS | NS | NS | NS | - | - | - | - | - |
| Protein+N | NS | −0.39 *** | NS | NS | NS | 0.37 *** | 0.73 *** | - | - | - | - |
| NUE | −0.33 ** | 0.74 *** | −0.73 *** | −0.42 *** | NS | −0.35 *** | NS | −0.26 * | - | - | - |
| NUpE | NS | 0.64 *** | −0.59 *** | −0.43 *** | NS | −0.27 * | 0.22 * | NS | 0.77 *** | - | - |
| NUtE | −0.29 * | 0.44 *** | −0.46 *** | NS | NS | −0.21 * | NS | −0.47 *** | 0.66 *** | NS | - |
| GU | 0.67 *** | NS | 0.82 *** | 0.59 *** | NS | 0.50 *** | −0.51 *** | NS | −0.59 *** | −0.48 *** | −0.32 ** |
| Group | N † | Yield | HI | NHI | Grain Protein | NUE | NUpE | NUtE | GU | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Low N | High N | Low N | High N | Low N | High N | Low N | High N | ||||||
| Mg ha−1 | kg kg−1 | kggrainN kgplantN−1 | g kg−1 | kg kgNfert.−1 | kgplantN kgNfert−1 | kg kgplantN−1 | kg kgplantN−1 | ||||||
| SSS | |||||||||||||
| 1A | 6 | 4.57 | 9.15 | 0.33 | 0.47 | 0.57 | 0.67 | 86.4 | 68.3 | 18.29 | 0.44 | 42.90 | 53.26 |
| 1B | 6 | 5.05 | 8.82 | 0.39 | 0.48 | 0.57 | 0.69 | 85.0 | 64.1 | 15.37 | 0.42 | 41.10 | 64.41 |
| 1C | 7 | 5.24 | 9.13 | 0.38 | 0.48 | 0.57 | 0.69 | 85.9 | 66.1 | 15.45 | 0.42 | 40.51 | 60.55 |
| 1D | 6 | 4.93 | 9.29 | 0.37 | 0.49 | 0.57 | 0.69 | 82.2 | 64.8 | 17.90 | 0.42 | 43.42 | 60.44 |
| 1E | 11 | 4.88 | 9.05 | 0.36 | 0.48 | 0.57 | 0.68 | 86.1 | 67.1 | 16.87 | 0.43 | 41.47 | 57.56 |
| LSD ‡ p ≤ 0.05 | 0.29 | 0.50 | 0.01 | 0.01 | 0.01 | 0.02 | 2.6 | 2.3 | 1.39 | 0.02 | 1.30 | 1.40 | |
| NSSS | |||||||||||||
| 2A | 3 | 5.31 | 9.32 | 0.42 | 0.50 | 0.57 | 0.70 | 83.5 | 64.0 | 16.21 | 0.40 | 42.70 | 64.92 |
| 2B | 16 | 5.02 | 9.17 | 0.37 | 0.48 | 0.57 | 0.69 | 83.3 | 65.0 | 16.71 | 0.42 | 42.22 | 60.18 |
| 2C | 14 | 5.08 | 8.74 | 0.40 | 0.49 | 0.57 | 0.70 | 88.1 | 67.2 | 14.85 | 0.42 | 40.22 | 61.26 |
| 2D | 4 | 4.31 | 9.52 | 0.29 | 0.45 | 0.57 | 0.66 | 86.3 | 69.1 | 21.11 | 0.48 | 43.75 | 51.91 |
| 2E | 16 | 4.78 | 9.18 | 0.34 | 0.47 | 0.57 | 0.67 | 84.5 | 66.1 | 17.47 | 0.44 | 42.06 | 56.65 |
| LSD p ≤ 0.05 | 0.21 | 0.52 | 0.02 | 0.02 | 0.00 | 0.02 | 3.2 | 2.3 | 1.38 | 0.03 | 1.05 | 1.47 | |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mastrodomenico, A.T.; Hendrix, C.C.; Below, F.E. Nitrogen Use Efficiency and the Genetic Variation of Maize Expired Plant Variety Protection Germplasm. Agriculture 2018, 8, 3. https://doi.org/10.3390/agriculture8010003
Mastrodomenico AT, Hendrix CC, Below FE. Nitrogen Use Efficiency and the Genetic Variation of Maize Expired Plant Variety Protection Germplasm. Agriculture. 2018; 8(1):3. https://doi.org/10.3390/agriculture8010003
Chicago/Turabian StyleMastrodomenico, Adriano T., C. Cole Hendrix, and Frederick E. Below. 2018. "Nitrogen Use Efficiency and the Genetic Variation of Maize Expired Plant Variety Protection Germplasm" Agriculture 8, no. 1: 3. https://doi.org/10.3390/agriculture8010003
APA StyleMastrodomenico, A. T., Hendrix, C. C., & Below, F. E. (2018). Nitrogen Use Efficiency and the Genetic Variation of Maize Expired Plant Variety Protection Germplasm. Agriculture, 8(1), 3. https://doi.org/10.3390/agriculture8010003