Carbon and Nitrogen Content of Soil Organic Matter and Microbial Biomass under Long-Term Crop Rotation and Tillage in Illinois, USA



Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Sites
2.2. Soil Analyses
2.3. Statistical Analyses
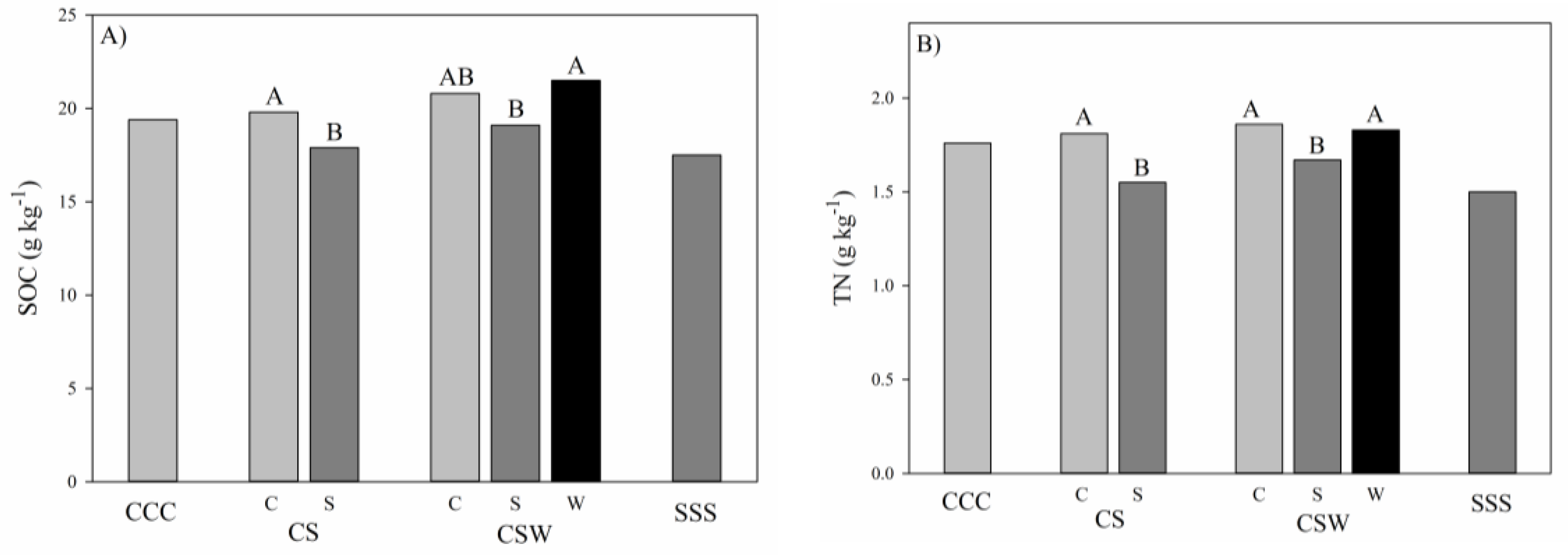
3. Results and Discussion
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| SOM | Soil organic matter |
| SOC | Soil organic carbon |
| MBC | Microbial biomass carbon |
| MBN | Microbial biomass nitrogen |
| CCC | Continuous corn |
| CS | Corn-soybean |
| CSW | Corn-soybean-wheat |
| SSS | Continuous soybean |
| NT | No-till |
| CT | Chisel tillage |
References
- Schimel, J.P.; Schaeffer, S.M. Microbial control over carbon cycling in soil. Front. Microbiol. 2012, 3, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Miltner, A.; Bombach, P.; Schmidt-Brücken, B.; Kästner, M. Som genesis: Microbial biomass as a significant source. Biogeochemistry 2012, 111, 41–55. [Google Scholar] [CrossRef]
- Blanco-Canqui, H.; Lal, R. Mechanisms of carbon sequestration in soil aggregates. Crit. Rev. Plant Sci. 2004, 23, 481–504. [Google Scholar] [CrossRef]
- Amézketa, E. Soil aggregate stability: A review. J Sustain. Agric. 1999, 14, 83–151. [Google Scholar] [CrossRef]
- Needelman, B.A.; Wander, M.M.; Bollero, G.A.; Boast, C.W.; Sims, G.K.; Bullock, D.G. Interaction of tillage and soil texture biologically active soil organic matter in Illinois. Soil Sci. Soc. Am. J. 1999, 63, 1326–1334. [Google Scholar] [CrossRef]
- Kumar, S.; Kadono, A.; Lal, R.; Dick, W. Long-term no-till impacts on organic carbon and properties of two contrasting soils and corn yields in Ohio. Soil Sci. Soc. Am. J. 2012, 76, 1798–1809. [Google Scholar] [CrossRef]
- McDaniel, M.D.; Tiemann, L.K.; Grandy, A.S. Does agricultural crop diversity enhance soil microbial biomass and organic matter dynamics? A meta-analysis. Ecol. Appl. 2014, 24, 560–570. [Google Scholar] [CrossRef] [PubMed]
- Stockmann, U.; Adams, M.A.; Crawford, J.W.; Field, D.J.; Henakaarchchi, N.; Jenkins, M.; Minasny, B.; McBratney, A.B.; De Remy de Courcelles, V.; Singh, K.; et al. The knowns, known unknowns and unknowns of sequestration of soil organic carbon. Agric. Ecosyst. Environ. 2013, 164, 80–99. [Google Scholar] [CrossRef]
- West, T.O.; Post, W.M. Soil organic carbon sequestration rates by tillage and crop rotation: A global data analysis. Soil Sci. Soc. Am. J. 2002, 66, 1930–1946. [Google Scholar] [CrossRef]
- Karlen, D.L.; Varvel, G.E.; Bullock, D.G.; Cruse, R.M. Crop rotations for the 21st century. Adv. Agron. 1994, 53, 1–45. [Google Scholar]
- Divito, G.A.; Rozas, H.R.S.; Echeverría, H.E.; Studdert, G.A.; Wyngaard, N. Long term nitrogen fertilization: Soil property changes in an argentinean pampas soil under no tillage. Soil Tillage Res. 2011, 114, 117–126. [Google Scholar] [CrossRef]
- Zuber, S.M.; Behnke, G.D.; Nafziger, E.D.; Villamil, M.B. Crop rotation and tillage effects on soil physical and chemical properties in Illinois. Agron. J. 2015, 107, 971–978. [Google Scholar] [CrossRef]
- Johnson, A.M.; Hoyt, G.D. Changes to the soil environment under conservation tillage. HortTechnology 1999, 9, 380–393. [Google Scholar]
- Martens, D.A. Nitrogen cycling under different soil management systems. Adv. Agron. 2001, 70, 143–192. [Google Scholar]
- Govaerts, B.; Mezzalama, M.; Unno, Y.; Sayre, K.D.; Luna-Guido, M.; Vanherck, K.; Dendooven, L.; Deckers, J. Influence of tillage, residue management, and crop rotation on soil microbial biomass and catabolic diversity. Appl. Soil Ecol. 2007, 37, 18–30. [Google Scholar] [CrossRef]
- Zuber, S.M.; Villamil, M.B. Meta-analysis approach to assess effect of tillage on microbial biomass and enzyme activities. Soil Biol. Biochem. 2016, 97, 176–187. [Google Scholar] [CrossRef]
- Balesdent, J.; Chenu, C.; Balabane, M. Relationship of soil organic matter dynamics to physical protection and tillage. Soil Tillage Res. 2000, 53, 215–230. [Google Scholar] [CrossRef]
- Nannipieri, P.; Ascher, J.; Ceccherini, M.T.; Landi, L.; Pietramellara, G.; Renella, G. Microbial diversity and soil functions. Eur. J. Soil Sci. 2003, 54, 655–670. [Google Scholar] [CrossRef]
- Soil Survey Staff Web Soil Survey. Available online: http://websoilsurvey.nrcs.usda.gov/ (accessed on 7 March 2018).
- McGeehan, S.L.; Naylor, D.V. Automated instrumental analysis of carbon and nitrogen in plant and soil samples. Commun. Soil Sci. Plant Anal. 1988, 19, 493–505. [Google Scholar] [CrossRef]
- Nelson, D.W.; Sommers, L.E. Total carbon, organic carbon, and organic matter. In Methods of Soil Analyses, Part 3, Chemical Methods; Sparks, D.L., Ed.; Soil Science Society of America: Madison, WI, USA, 1996; pp. 961–1010. [Google Scholar]
- Rice, C.W.; Moorman, T.B.; Beare, M. Role of microbial biomass carbon and nitrogen in soil quality. In Methods for Assessing Soil Quality; Doran, J.W., Jones, A.J., Eds.; Soil Science Society of America: Madison, WI, USA, 1996; pp. 203–215. [Google Scholar]
- Sparling, G.P.; West, A.W. Importance of soil water content when estimating soil microbial C, N and P by the fumigation-extraction methods. Soil Biol. Biochem. 1989, 21, 245–253. [Google Scholar] [CrossRef]
- Littell, R.C.; Milliken, G.A.; Stroup, W.W.; Wolfinger, R.D.; Schabenberger, O. Sas for Mixed Models, 2nd ed.; SAS Institute, Inc.: Cary, NC, USA, 2006; p. 745. [Google Scholar]
- Van Eerd, L.L.; Congreves, K.A.; Hayes, A.; Verhallen, A.; Hooker, D.C. Long-term tillage and crop rotation effects on soil quality, organic carbon, and total nitrogen. Can. J. Soil Sci. 2014, 94, 303–315. [Google Scholar] [CrossRef]
- Ajwa, H.; Tabatabai, M. Decomposition of different organic materials in soils. Biol. Fertil. Soils 1994, 18, 175–182. [Google Scholar] [CrossRef]
- Melillo, J.M.; Aber, J.D.; Linkins, A.E.; Ricca, A.; Fry, B.; Nadelhoffer, K.J. Carbon and nitrogen dynamics along the decay continuum: Plant litter to soil organic matter. In Ecology of Arable Land—Perspectives and Challenges, Proceedings of the International Symposium, Swedish University of Agricultural Sciences, Uppsala, Sweden, 9–12 June 1987; Clarholm, M., Bergström, L., Eds.; Springer: Dordrecht, The Netherlands, 1989; pp. 53–62. [Google Scholar]
- Shahbaz, M.; Kuzyakov, Y.; Heitkamp, F. Decrease of soil organic matter stabilization with increasing inputs: Mechanisms and controls. Geoderma 2017, 304, 76–82. [Google Scholar] [CrossRef]
- Xiao, C.; Guenet, B.; Zhou, Y.; Su, J.; Janssens, I.A. Priming of soil organic matter decomposition scales linearly with microbial biomass response to litter input in steppe vegetation. Oikos 2015, 124, 649–657. [Google Scholar] [CrossRef]
- Moreno-Cornejo, J.; Zornoza, R.; Doane, T.A.; Faz, Á.; Horwath, W.R. Influence of cropping system management and crop residue addition on soil carbon turnover through the microbial biomass. Biol. Fertil. Soils 2015, 51, 839–845. [Google Scholar] [CrossRef]
- Havlin, J.L.; Kissel, D.E.; Maddux, L.D.; Claassen, M.M.; Long, J.H. Crop rotation and tillage effects on soil organic carbon and nitrogen. Soil Sci. Soc. Am. J. 1990, 54, 448–452. [Google Scholar] [CrossRef]
- Varvel, G.E. Rotation and nitrogen fertilization effects on changes in soil carbon and nitrogen. Agron. J. 1994, 86, 319–325. [Google Scholar] [CrossRef]
- Ciampitti, I.A.; Vyn, T.J. Physiological perspectives of changes over time in maize yield dependency on nitrogen uptake and associated nitrogen efficiencies: A review. Field Crops Res. 2012, 133, 48–67. [Google Scholar] [CrossRef]
- Delogu, G.; Cattivelli, L.; Pecchioni, N.; De Falcis, D.; Maggiore, T.; Stanca, A.M. Uptake and agronomic efficiency of nitrogen in winter barley and winter wheat. Eur. J. Agron. 1998, 9, 11–20. [Google Scholar] [CrossRef]
- Gentry, L.E.; Below, F.E.; David, M.B.; Bergerou, J.A. Source of the soybean N credit in maize production. Plant Soil 2001, 236, 175–184. [Google Scholar] [CrossRef]
- Salvagiotti, F.; Cassman, K.G.; Specht, J.E.; Walters, D.T.; Weiss, A.; Dobermann, A. Nitrogen uptake, fixation and response to fertilizer n in soybeans: A review. Field Crops Res. 2008, 108, 1–13. [Google Scholar] [CrossRef]
- Varvel, G.E.; Wilhelm, W.W. No-tillage increases soil profile carbon and nitrogen under long-term rainfed cropping systems. Soil Tillage Res. 2011, 114, 28–36. [Google Scholar] [CrossRef]
- Franzluebbers, A.J.; Hons, F.M. Soil-profile distribution of primary and secondary plant-available nutrients under conventional and no tillage. Soil Tillage Res. 1996, 39, 229–239. [Google Scholar] [CrossRef]
- Chabbi, A.; Rumpel, C. Organic matter dynamics in agro-ecosystems—The knowledge gaps. Eur. J. Soil Sci. 2009, 60, 153–157. [Google Scholar] [CrossRef]
- Fontaine, S.; Barot, S. Size and functional diversity of microbe populations control plant persistence and long-term soil carbon accumulation. Ecol. Lett. 2005, 8, 1075–1087. [Google Scholar] [CrossRef]
- Russell, A.E.; Laird, D.A.; Mallarino, A.P. Nitrogen fertilization and cropping system impacts on soil quality in midwestern mollisols. Soil Sci. Soc. Am. J. 2006, 70, 249–255. [Google Scholar] [CrossRef]
- Ekenler, M.; Tabatabai, M. Β-glucosaminidase activity of soils: Effect of cropping systems and its relationship to nitrogen mineralization. Biol. Fertil. Soils 2002, 36, 367–376. [Google Scholar] [CrossRef]
- Bonkowski, M. Protozoa and plant growth: The microbial loop in soil revisited. New Phytol. 2004, 162, 617–631. [Google Scholar] [CrossRef]
- Nannipieri, P.; Eldor, P. The chemical and functional characterization of soil N and its biotic components. Soil Biol. Biochem. 2009, 41, 2357–2369. [Google Scholar] [CrossRef]
- Schimel, J.P.; Bennett, J. Nitrogen mineralization: Challenges of a changing paradigm. Ecology 2004, 85, 591–602. [Google Scholar] [CrossRef]
- Carter, M.R.; Gregorich, E.G.; Angers, D.A.; Beare, M.H.; Sparling, G.P.; Wardle, D.A.; Voroney, R.P. Interpretation of microbial biomass measurements for soil quality assessment in humid temperate regions. Can. J. Soil Sci. 1999, 79, 507–520. [Google Scholar] [CrossRef]


{kind=link}
| Source | SOC | TN | MBC | MBN |
|---|---|---|---|---|
| Rotation (R) | 0.057 | 0.001 | 0.557 | 0.507 |
| Tillage (T) | 0.072 | 0.021 | 0.249 | 0.112 |
| R × T | 0.550 | 0.319 | 0.819 | 0.358 |
| Depth (D) | 0.001 | 0.352 | <0.0001 | <0.0001 |
| R × D | 0.590 | 0.117 | 0.600 | 0.363 |
| T × D | 0.074 | 0.001 | 0.098 | 0.086 |
| R × T × D | 0.901 | 0.669 | 0.267 | 0.339 |
| Depth (cm) | Depth (cm) | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 0–10 | 10–20 | 0–20 | 0–10 | 10–20 | 0–20 | ||||||||
| SOC (g kg−1) | TN (g kg−1) | ||||||||||||
| Rotation | Tillage | Mean | SE | Mean | SE | Mean | SE | Mean | SE | Mean | SE | Mean | SE |
| CCC * | CT | 19.8 | 6.4 | 17.3 | 6.3 | 18.5 | 6.3 | 1.83 | 0.31 | 1.57 | 0.13 | 1.70 | 0.17 |
| NT | 22.2 | 18.3 | 20.2 | 2.03 | 1.62 | 1.82 | |||||||
| CS | CT | 19.2 | 17.3 | 18.3 | 1.81 | 1.50 | 1.65 | ||||||
| NT | 21.7 | 17.1 | 19.4 | 2.37 | 1.50 | 1.94 | |||||||
| CSW | CT | 21.4 | 19.2 | 20.3 | 2.04 | 1.65 | 1.84 | ||||||
| NT | 23.2 | 18.2 | 20.7 | 2.05 | 1.55 | 1.80 | |||||||
| SSS | CT | 16.8 | 16.2 | 16.5 | 1.39 | 1.38 | 1.38 | ||||||
| NT | 20.1 | 17.7 | 18.6 | 1.78 | 1.46 | 1.62 | |||||||
| CCC | 21.0 | 6.4 | 17.8 | 6.4 | 19.4 AB § | 6.3 | 1.93 | 0.23 | 1.60 | 0.13 | 1.76 A | 0.14 | |
| CS | 20.5 | 17.2 | 18.8 AB | 2.09 | 1.50 | 1.80 A | |||||||
| CSW | 22.3 | 18.7 | 20.5 A | 2.04 | 1.60 | 1.82 A | |||||||
| SSS | 18.4 | 16.6 | 17.5 B | 1.58 | 1.42 | 1.50 B | |||||||
| CT | 19.3 b †,‡ | 6.3 | 17.5 b | 6.3 | 18.4 B | 6.3 | 1.76 b | 0.17 | 1.52 ab | 0.12 | 1.64 B | 0.11 | |
| NT | 21.8 a | 17.7 b | 19.7 A | 2.06 a | 1.53 ab | 1.79 A | |||||||
| Depth (cm) | |||||||
|---|---|---|---|---|---|---|---|
| 0–10 | 10–20 | 0–20 | |||||
| Rotation | Tillage | Mean | SE | Mean | SE | Mean | SE |
| CCC * | CT | 3.75 (42.7) | 0.31 | 3.32 (27.6) | 0.31 | 3.53 (34.3) | 0.28 |
| NT | 4.12 (61.5) | 0.31 | 3.43 (30.8) | 0.31 | 3.77 (43.5) | 0.28 | |
| CS | CT | 4.02 (55.5) | 0.28 | 3.57 (35.4) | 0.28 | 3.79 (44.3) | 0.26 |
| NT | 4.37 (79.3) | 0.28 | 3.61 (36.8) | 0.28 | 3.99 (54.0) | 0.26 | |
| CSW | CT | 4.12 (61.6) | 0.27 | 3.29 (26.9) | 0.27 | 3.71 (40.7) | 0.26 |
| NT | 4.23(68.6) | 0.27 | 3.52 (33.7) | 0.27 | 3.87 (48.1) | 0.26 | |
| SSS | CT | 4.13 (61.9) | 0.31 | 3.66 (39.0) | 0.31 | 3.89 (49.1) | 0.28 |
| NT | 4.31 (74.7) | 0.31 | 3.32 (27.8) | 0.31 | 3.82 (45.6) | 0.28 | |
| CCC | 3.94 (51.2) | 0.27 | 3.37 (29.1) | 0.27 | 3.65 (38.6) | 0.25 | |
| CS | 4.19 (66.3) | 0.25 | 3.59 (36.1) | 0.25 | 3.89 (48.9) | 0.24 | |
| CSW | 4.17 (65.0) | 0.24 | 3.40 (30.1) | 0.24 | 3.79 (44.3) | 0.24 | |
| SSS | 4.22 (68.0) | 0.27 | 3.49 (32.9) | 0.27 | 3.86 (47.3) | 0.25 | |
| CT | 4.00 (54.8) a †,‡ | 0.24 | 3.46 (31.8) b | 0.24 | 3.73 (41.8) | 0.23 | |
| NT | 4.26 (70.7) a | 0.24 | 3.47 (32.1) b | 0.24 | 3.86 (47.6) | 0.23 | |
| Depth (cm) | |||||||
|---|---|---|---|---|---|---|---|
| 0–10 | 10–20 | 0–20 | |||||
| Rotation | Tillage | Mean | SE | Mean | SE | Mean | SE |
| CCC * | CT | 1.80 (6.04) | 0.36 | 0.61 (1.85) | 0.29 | 1.21 (3.34) | 0.28 |
| NT | 1.86 (6.43) | 0.36 | 1.68 (5.37) | 0.28 | 1.77 (5.87) | 0.27 | |
| CS | CT | 2.19 (8.94) | 0.27 | 1.54 (4.67) | 0.22 | 1.87 (6.46) | 0.22 |
| NT | 2.06 (7.86) | 0.27 | 1.51 (4.52) | 0.21 | 1.79 (5.96) | 0.21 | |
| CSW | CT | 2.10 (8.14) | 0.23 | 1.37 (3.94) | 0.19 | 1.73 (5.66) | 0.19 |
| NT | 2.04 (7.66) | 0.23 | 1.61 (4.98) | 0.19 | 1.82 (6.18) | 0.19 | |
| SSS | CT | 2.17 (8.73) | 0.36 | 1.13 (3.11) | 0.28 | 1.65 (5.21) | 0.27 |
| NT | 2.66 (14.28) | 0.36 | 1.60 (4.97) | 0.28 | 2.13 (8.43) | 0.27 | |
| CCC | 1.83 (6.23) | 0.27 | 1.15 (3.15) | 0.22 | 1.49 (4.43) | 0.21 | |
| CS | 2.13 (8.38) | 0.21 | 1.52 (4.59) | 0.18 | 1.83 (6.21) | 0.17 | |
| CSW | 2.07 (7.90) | 0.19 | 1.49 (4.43) | 0.16 | 1.78 (5.92) | 0.16 | |
| SSS | 2.41 (11.17) | 0.27 | 1.37 (3.93) | 0.21 | 1.89 (6.62) | 0.21 | |
| CT | 2.06 (7.87) a †,‡ | 0.17 | 1.16 (3.20) c | 0.14 | 1.61 (5.02) | 0.14 | |
| NT | 2.15 (8.62) a | 0.17 | 1.60 (4.95) b | 0.14 | 1.88 (6.53) | 0.14 | |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zuber, S.M.; Behnke, G.D.; Nafziger, E.D.; Villamil, M.B. Carbon and Nitrogen Content of Soil Organic Matter and Microbial Biomass under Long-Term Crop Rotation and Tillage in Illinois, USA. Agriculture 2018, 8, 37. https://doi.org/10.3390/agriculture8030037
Zuber SM, Behnke GD, Nafziger ED, Villamil MB. Carbon and Nitrogen Content of Soil Organic Matter and Microbial Biomass under Long-Term Crop Rotation and Tillage in Illinois, USA. Agriculture. 2018; 8(3):37. https://doi.org/10.3390/agriculture8030037
Chicago/Turabian StyleZuber, Stacy M., Gevan D. Behnke, Emerson D. Nafziger, and Maria B. Villamil. 2018. "Carbon and Nitrogen Content of Soil Organic Matter and Microbial Biomass under Long-Term Crop Rotation and Tillage in Illinois, USA" Agriculture 8, no. 3: 37. https://doi.org/10.3390/agriculture8030037
APA StyleZuber, S. M., Behnke, G. D., Nafziger, E. D., & Villamil, M. B. (2018). Carbon and Nitrogen Content of Soil Organic Matter and Microbial Biomass under Long-Term Crop Rotation and Tillage in Illinois, USA. Agriculture, 8(3), 37. https://doi.org/10.3390/agriculture8030037