Abstract
Two of the most troublesome weeds in soybean, cotton, and corn in cropping systems of mid-south United States (US) are Senna obtusifolia and Xanthium strumarium. Understanding their population dynamics, particularly weed seedling emergence patterns, is important for the timely implementation and the success of weed management strategies. Identifying the sources of variation of emergence patterns could greatly improve our ability to predict emergence timing. A three-years field study was conducted to determine the effect of environmental conditions on S. obtusifolia and X. strumarium seedling emergence and seedbank potential. The experiment was conducted with two seed sources; X. strumarium burs and S. obtusifolia seeds from a single maternal plant source, and X. strumarium burs and S. obtusifolia seeds from multiple maternal plant sources, both being exposed either to 5 cm burial depth (buried) or left on the soil surface (soil surface) in the fallow or planted in spring after their storage under chilled (chill) or room temperature (no chill) conditions. X. strumarium and S. obtusifolia seedling emergence was lower from burs and seeds that were planted in the soil in September as compared with the chill and/or no chill seeds/burs stored for six months. X. strumarium seedling emergence was reduced from 37 to 1% when burs were left on the soil surface when compared to buried burs. S. obtusifolia seedling emergence was reduced from 47 to 13% when seeds were left on the soil surface as compared to buried. At the end of the experimental period, the soil seedbank of X. strumarium had been significantly depleted, whereas the remaining seeds of S. obtusifolia were viable.
1. Introduction
Senna obtusifolia (Sicklepod) and Xanthium strumarium (common cocklebur) are two of the most important weeds species in row cropping systems of the mid- and southeastern United States (US) [1,2] as they have the potential to become major weeds of many crops, including soybean, cotton, and maize in short time periods [3]. In addition, these species can invade and dominate pastures, roadsides, fence lines, creek banks, and other disturbed areas [4].
Leon et al. [1] reported that integrated approaches which include multiple tactics and the incorporation of pre- with sequential post-application of herbicides are necessary to ensure S. obtusifolia seedbank reductions as plants that survived a single herbicide application are capable of producing abundant viable seeds. Consequently, weed management should aim at preventing weed reproduction and reducing weed emergence after crop planting [5]. Effective weed management systems will consist of multiple control tactics, which are integrating weed biology and population dynamics [5] as a vehicle for the development of cropping systems that minimize the opportunity for weed growth, reproduction, seedling emergence, and recruitment [6,7,8]. Therefore, the determination of the soil seedbank is of great importance for understanding weed population dynamics and implementing effective weed management tactics since potential weed problems exist as long as weed seeds remain viable in the soil [9]. Understanding the ecophysiological processes that control weed seed dormancy and germination with the intention of explaining and eventually predicting weed seedling emergence patterns is of great interest [10,11].
In addition, research has been conducted to characterize how tillage systems affect seedling emergence patterns [12,13,14,15]. The magnitude of X. strumarium emergence is influenced by tillage, crop presence and various interspecific competition regimes [16,17]. Tillage increases X. strumarium and S. obtusifolia seedling emergence through soil aeration and increased temperature [18]. Under tilled conditions Bararpour and Oliver [16] reported 11% seedling emergence for both species, in relation to the initial seedbank, the first year of seed deposition. On the contrary, seedling emergence under no-tilled conditions was reduced to 0.7% and 1.6% for X. strumarium and S. obtusifolia, respectively. These authors also recorded high seed deposition for both species, even when they were grown under intra- and interspecific competitive conditions for two consecutive years. However, since weed seeds persist in the soil, understanding seedbank dynamics over multiple years is important for devising multiyear strategies to manage weeds. The purpose of this paper is to investigate how simulated tillage strategies (buried vs. unburied) in combination with chill or no chill conditions can affect the soil seedbank of both X. strumarium and S. obtusifolia. In addition, to examine the seed survival mechanisms when seeds for both X. strumarium and S. obtusifolia originated from single or multiple parental plants.
2. Materials and Methods
2.1. Seed Material and Experimental Set up
Hundred S. obtusifolia seeds and X. strumarium burs from a single maternal source (one plant), three days after their collection in September, were planted at a 5-cm depth within a circle of 7-cm in diameter. Another hundred S. obtusifolia seeds and X. strumarium burs were left on the soil surface of an experimental plot, which was enclosed by a 0.5- by 0.5-m wood frame 15 cm in height to prevent seeds/burs from being washed or blown out of the plot area. The planting area had no previous history of the weed species under investigation.
Some of the harvested S. obtusifolia seeds and X. strumarium burs were stored at room temperature (no chill) 18 to 24 °C, and some of the seeds and burs were exposed under chilling conditions at 0 to 2 °C and relative humidity between 25–30%. In May, one-hundred no chill S. obtusifolia seeds and X. strumarium burs and one-hundred chill seeds from both of the species were planted at a 5-cm depth in the middle of 0.5 × 0.5-m plots. Experimental plots, the middle of which was marked with a flag, were arranged in a completely randomized design with four replications. The soil was a Taloka silt loam (fine, mixed, thermic Mollic albaqualfs), with 25% sand, 62% silt, 13% clay, 1% organic matter, and pH of 6.7. The second year of the study, another experiment was conducted using the same procedure as above, except that the seed source was originating from a multiple maternal seed sources of the initial cocklebur and S. obtusifolia populations, had already established in previous year, and left to grow under intense intraspecific interference. The first germination experiment was run over three years, while the second was an independent treatment that was run over two years. Seedling emergence was recorded every two weeks until the end of the growing season.
2.2. Monitoring Soil Seed Reserves
After three years, soil samples were taken from the middle of each plot (buried treatment) with an 11-cm-diameter soil core at a 15-cm depth to retrieve the remaining S. obtusifolia seeds and X. strumarium burs. Each soil sample was passed through a series of four sieves containing screens of the following sizes: 4.75 mm to collect X. strumarium, 2.00 mm to collect S. obtusifolia, 1.00 mm security sieves for escaped S. obtusifolia seeds, and 0.25 mm to collect soil so that most of the soil would not go down the sink drain. Prior to screening, soil cores were soaked until a slurry was obtained. Water was run through the sieves with the sample slurry to enhance the movement through the screens. Seeds and burs were separated and were counted according to species. The seeds that were left on the soil surface of the 0.5 m × 0.5 m framed plots were collected by hand. To secure that the entire seed sample was collected, the soil surface of the sampling area was swapped and the debris was moved to the lab for further processing.
2.3. Germination Tests
Visual examination of the X. strumarium burs by bur dissection indicated whether enclosed seeds were desiccated, therefore not suitable for germination test. Germination tests were conducted to assess the viability of collected seeds that were retrieved from the sampling areas. S. obtusifolia seeds before expose to germination test were scarified with H2S04 (98%) for 10 min. For the germination evaluations, the remaining S. obtusifolia seeds were placed in a 9-cm diameter plastic petri dish (Fisher Scientific, Suwanee, GA, USA) lined with two layers of Whatman filter paper (Whatman’s No. 1, Fisher Scientific, Suwanee, GA, USA) and moistened with 10 mL of distilled water containing 100 ppm of captan [N-(trichloromethyl-thio)-r-cyclohexene-1,2-dicarboximide] (Drexel Chemical Company, Memphis, TN, USA) to control pathogens. These were incubated for seven days at 25 °C. Distilled water was added as needed to remoisten Whatman filter papers. Germinated seeds were counted and removed at seven days after seeds were placed in the incubator. Then, non-germinated S. obtusifolia seeds were scarified for a second time for 12 min and were placed back to the petri dishes and incubator. After five days, the remaining non-germinated seeds were scarified for a third time for 15 min.
2.4. Data Analysis
The expected number of remaining seeds after seedling emergence was estimated based on Equation (1).
The number of seeds loss was estimated based on Equation (2).
E = Expected No. of seeds; I = Initial seed deposition; SE = Seedling emergence; SL = Seed loss; R = Retrieved seeds.
Data were subjected to analysis of variance and means were separated with least significant difference (LSD) at the 5% level of probability using SAS 9.3 software (SAS Institute Inc., Cary, NC, USA).
3. Results
3.1. Single Maternal Seed Source
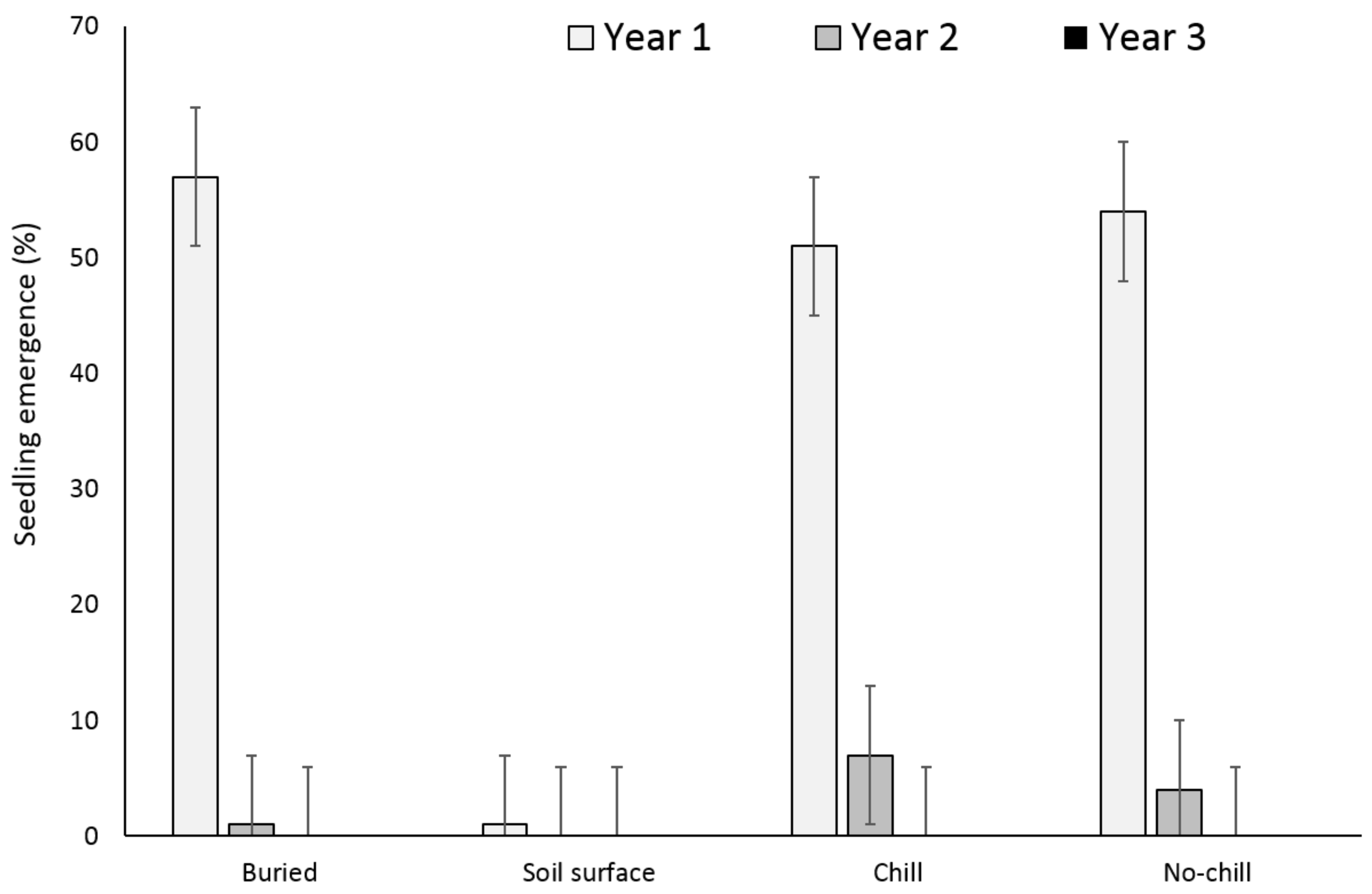
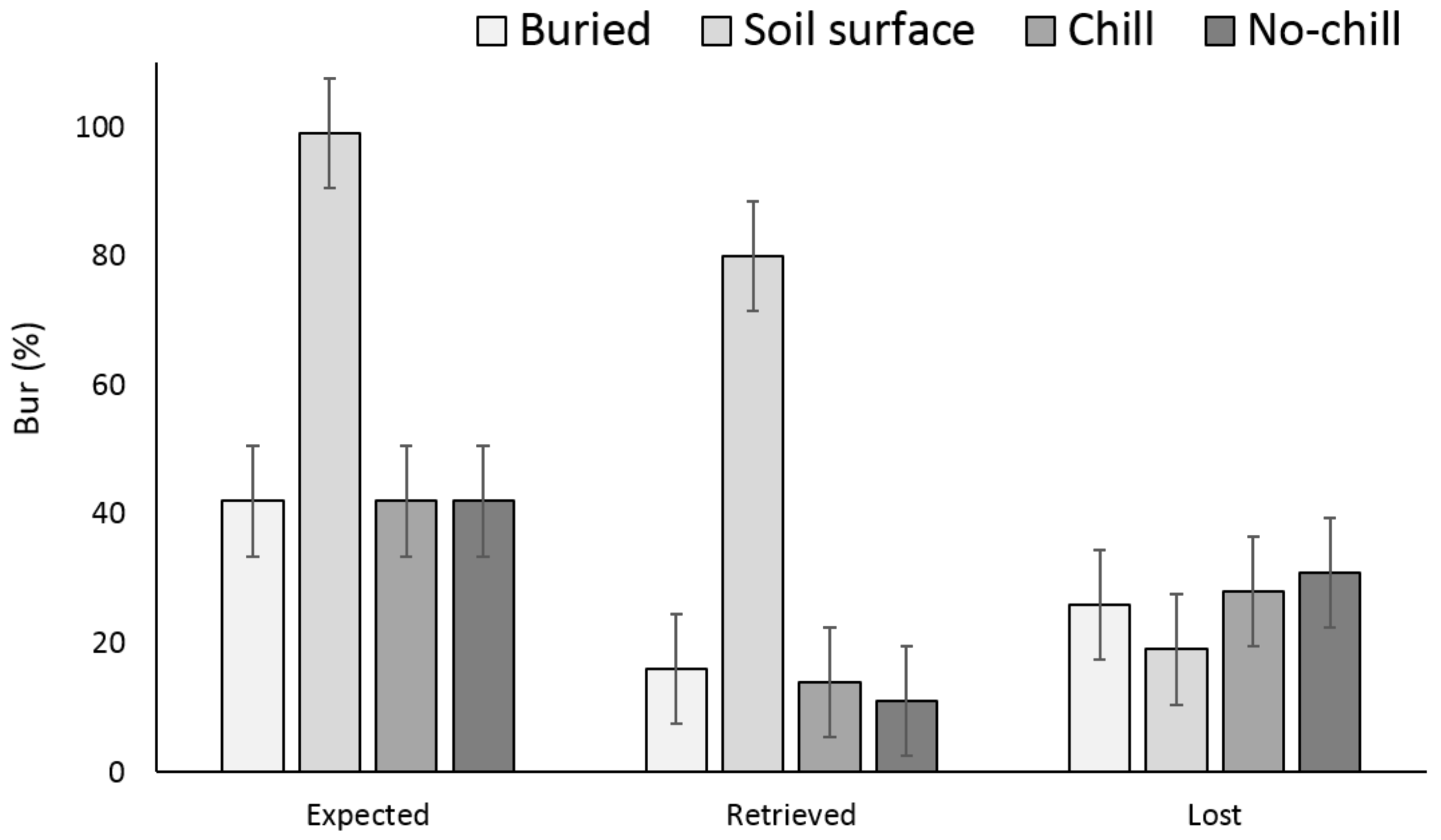
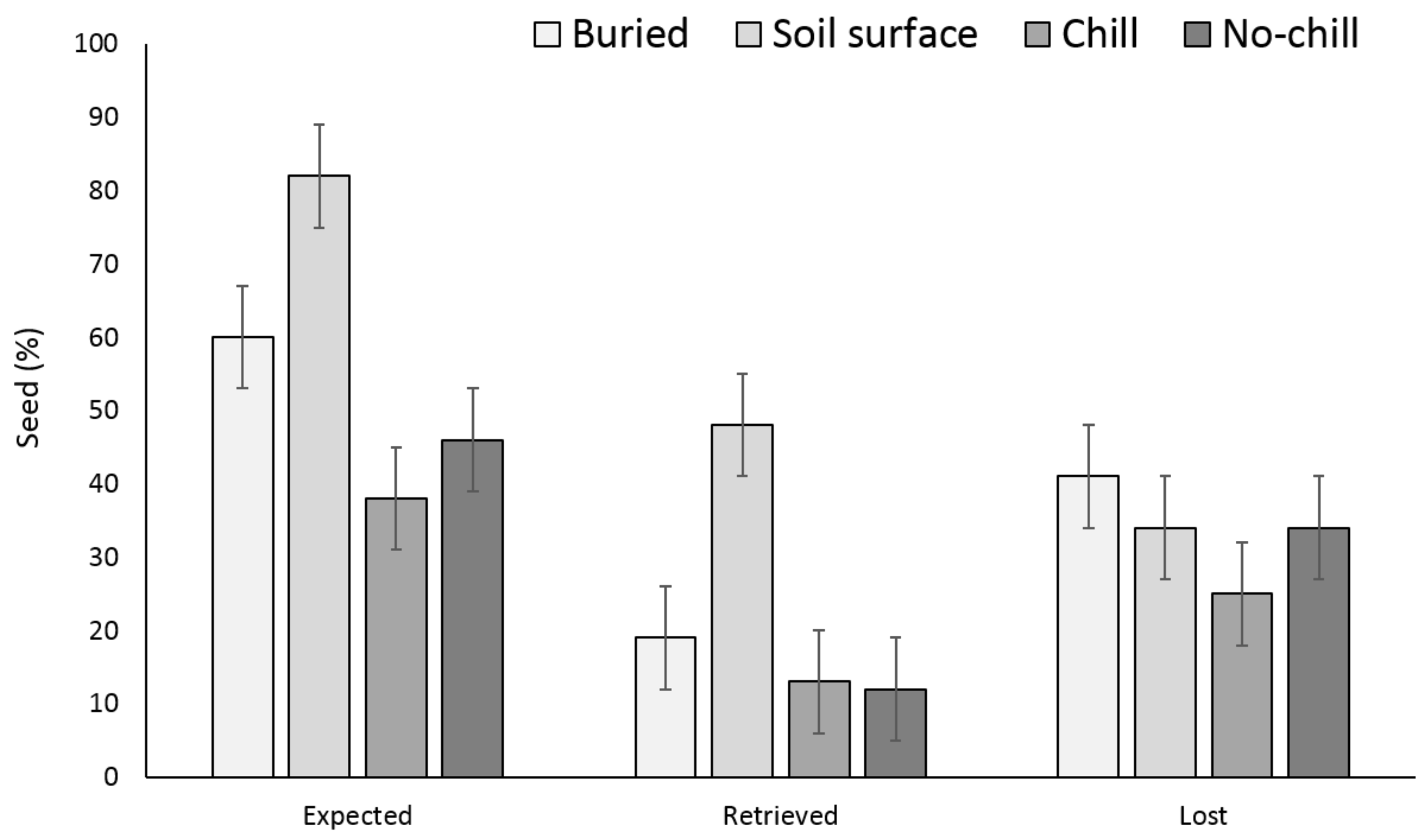
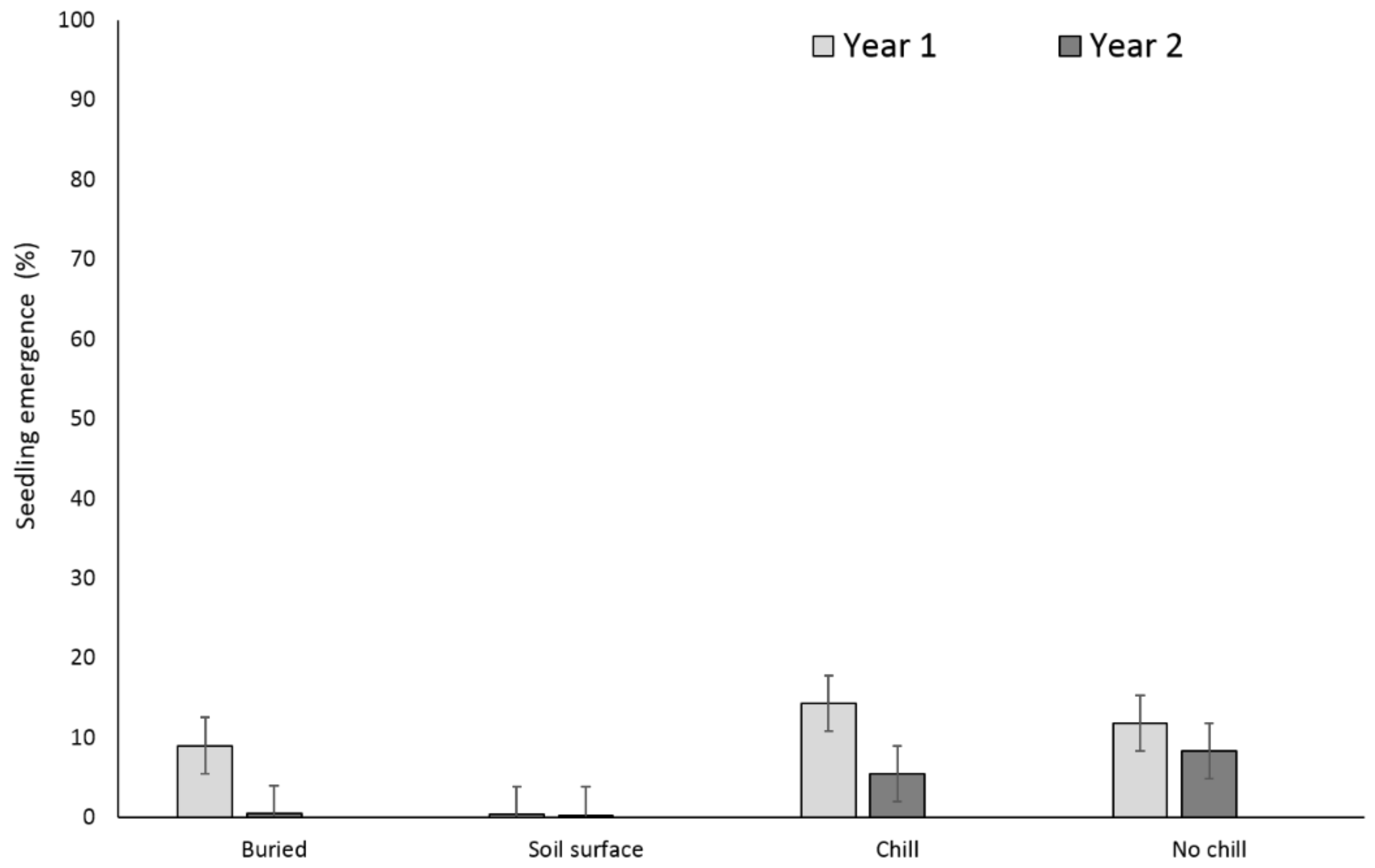
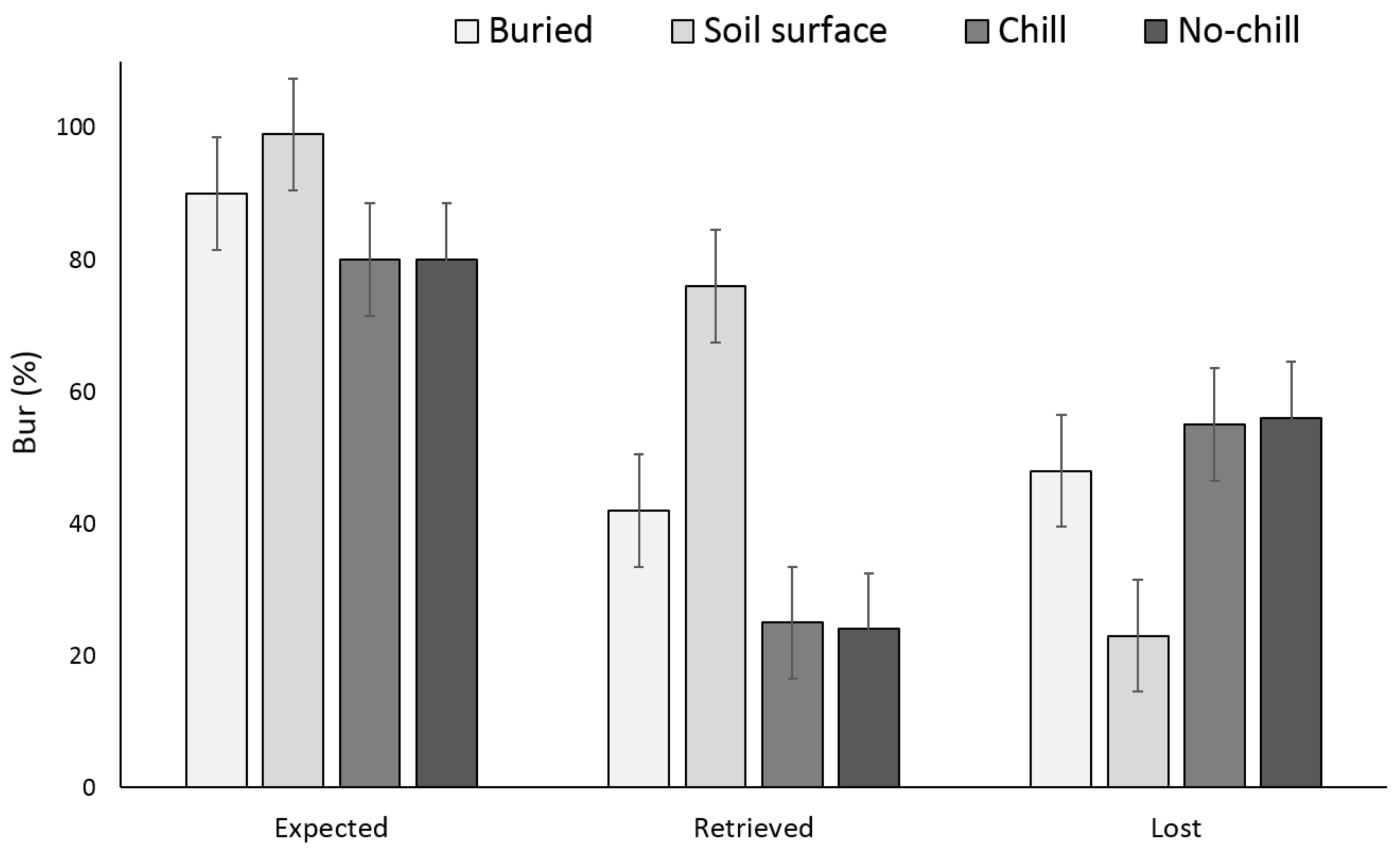
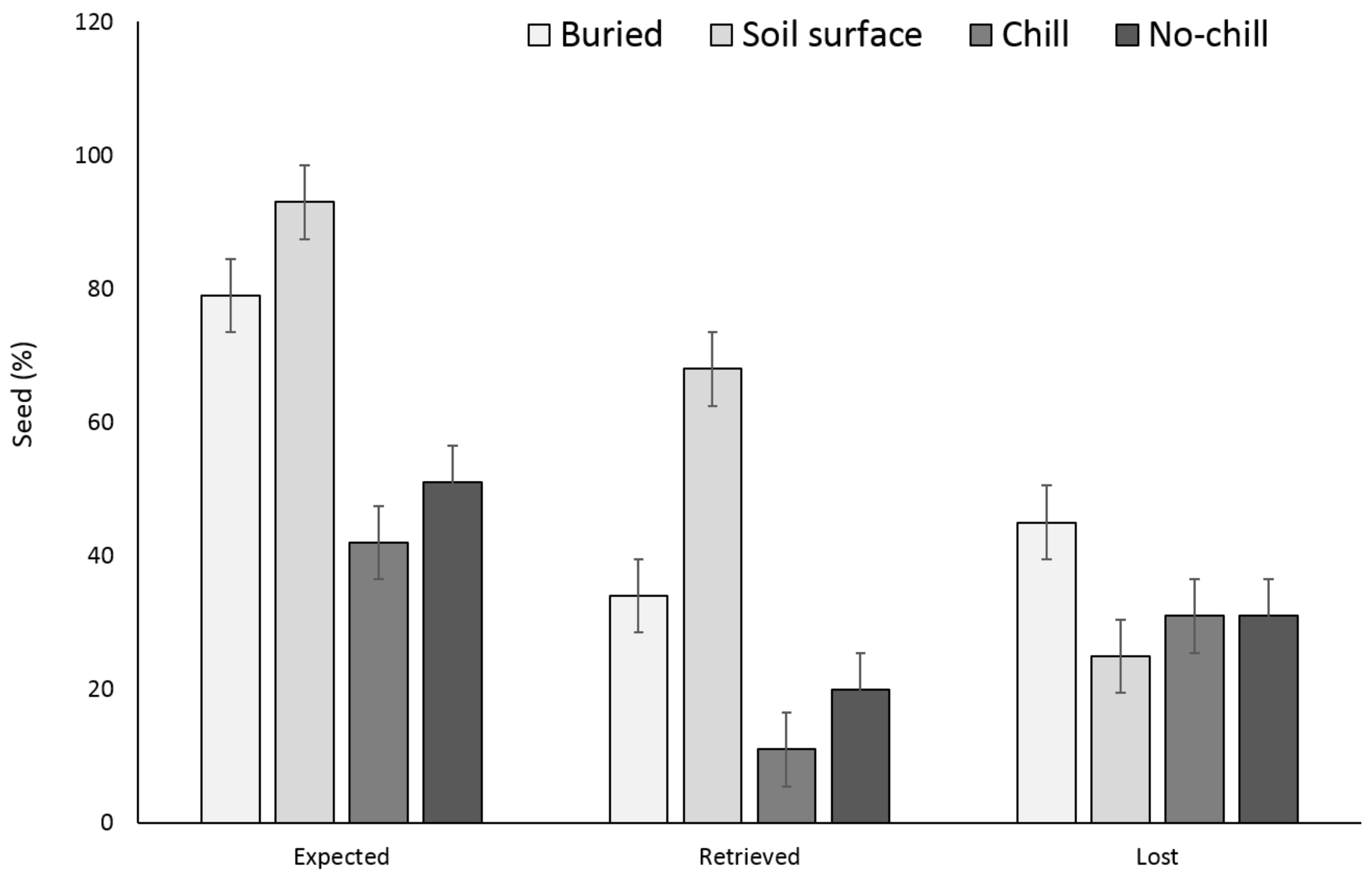
There were no significant differences between the buried seed treatments (i.e., buried in September or May after having exposed to chill or no chill conditions) for X. strumarium or S. obtusifolia seedling emergence (Figure 1 and Figure 2). Seedling emergence of X. strumarium and S. obtusifolia was 54% and 46% (an average of September and May planting dates), respectively, when seeds and burs were buried. When the seeds and burs remained on the soil surface, only 1% and 5% of X. strumarium burs and S. obtusifolia seeds emerged, respectively. The second year the X. strumarium seedbank was depleted (Figure 1). After three years, an average of 58% of X. strumarium burs had emerged regardless of whether burs were planted in September or exposed to chill or no-chill conditions. Soil sampling indicated that an average of 28 and 19% of the burs were lost when they were buried or left on the soil surface, respectively (Figure 3). None of the X. strumarium burs that were collected by soil sampling contained viable achenes. Three years after, the S. obtusifolia seedbank was not completely depleted (Figure 2). The total percent seedling emergence by that time was recorded at 62% and 54%, respectively, for seeds under chill conditions or seeds that were exposed under room temperature (no chill). Buried seeds or seeds that were left on the soil surface in September resulted in 40% and 18% seedling emergence, respectively. Soil sampling indicated that an average of 36% of the seeds were lost when seeds were planted (Figure 4) versus 34% when seeds were left on the soil surface. Germination test indicated that 100% of remaining S. obtusifolia seeds were viable and would germinate (Table 1).
Figure 1.
Effect of environmental conditions on X. strumarium emergence originating from a single maternal source. Vertical bars represent least significant difference (LSD) at 5% significance level.
Figure 2.
Effect of environmental conditions on S. obtusifolia emergence originating from a single maternal source. Vertical bars represent LSD at 5% significance level.
Figure 3.
Effect of environmental conditions on X. strumarium bur loss originating from a single maternal source after three years. Vertical bars represent LSD at 5% significance level. Expected is the sum of non-germinated seeds.
Figure 4.
Effect of environmental conditions on S. obtusifolia seed loss originating from a single maternal source after three years. Vertical bars represent LSD at 5% significance level. Expected is the sum of non-germinated seeds.

Table 1.
Percentage germination of S. obtusifolia seeds from single maternal source under various environmental conditions.
3.2. Multiple Maternal Seed Source
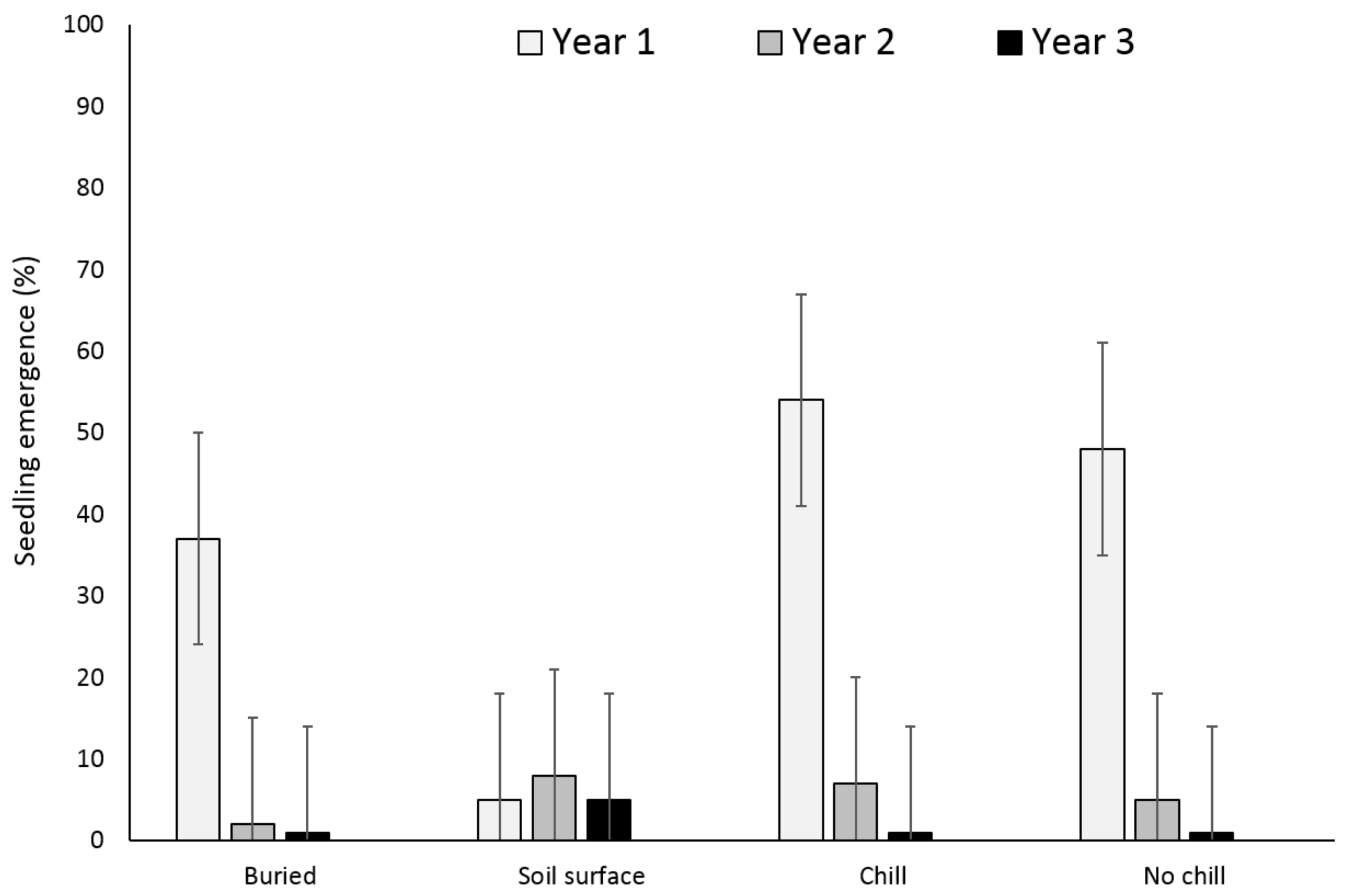
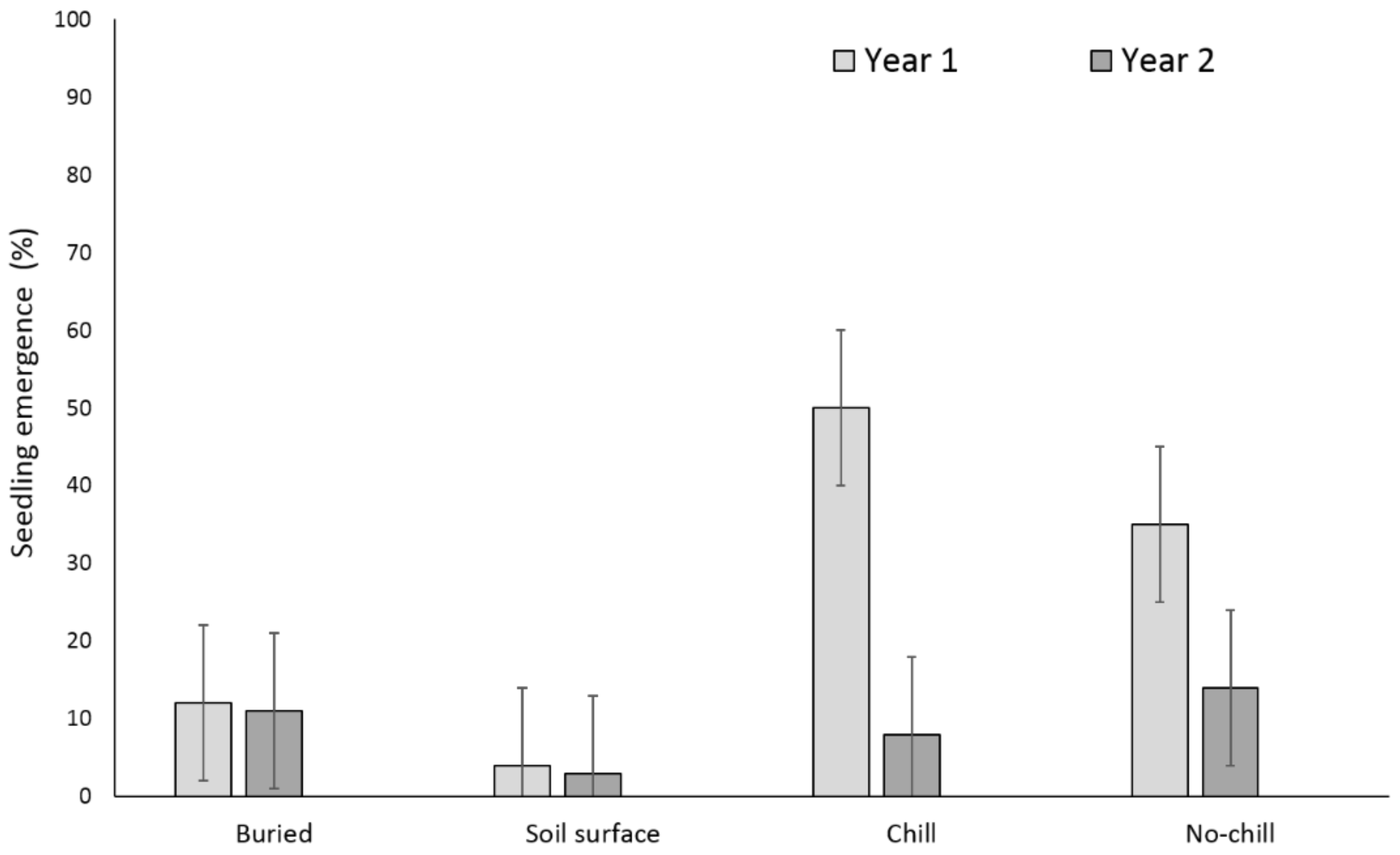
Two years after, X. strumarium and S. obtusifolia seedling emergence was highest when the seeds or burs were exposed to chill conditions for six months and were planted in the field in May (Figure 5 and Figure 6). X. strumarium and S. obtusifolia seedling emergence was the lowest when seeds and burs were left on the soil surface; only 0.4% and 4% emerged, respectively. An average of 12% and 34% of X. strumarium burs and S. obtusifolia seeds emerged, respectively, when seeds/burs were planted in the soil. Upon the completion of the experiment (two years after the establishment of the study), there were differences in X. strumarium seedling emergence at different planting dates or when burs were left on the soil surface (Figure 5). S. obtusifolia seedling emergence was the lowest when seeds were left on the soil surface when compared to seeds planted in soil (Figure 6). Total seedling counts indicate that when X. strumarium burs and S. obtusifolia seeds were exposed under chill or no-chill conditions (for 6 months), seedling emergence was the highest (Figure 5 and Figure 6). Soil sampling indicated that an average of 53 and 36% of X. strumarium burs and S. obtusifolia seeds were lost when burs and seeds were buried (Figure 7 and Figure 8). X. strumarium burs and S. obtusifolia seed lost were 23% and 25% when burs and seeds were left on the soil surface (Figure 7 and Figure 8). Seed or bur loss probably was due to decay, disease, and predation. Germination of remaining X. strumarium burs was 0% due to decomposition. The germination test indicated that 100% of the remaining S. obtusifolia seeds were viable and would germinate.
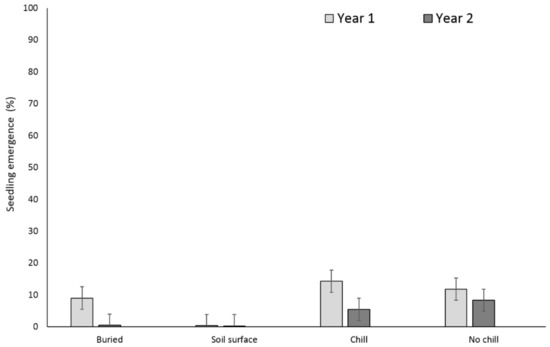
Figure 5.
Effect of environmental conditions on X. strumarium emergence originating from a multiple maternal source. Vertical bars represent LSD at 5% significance level.
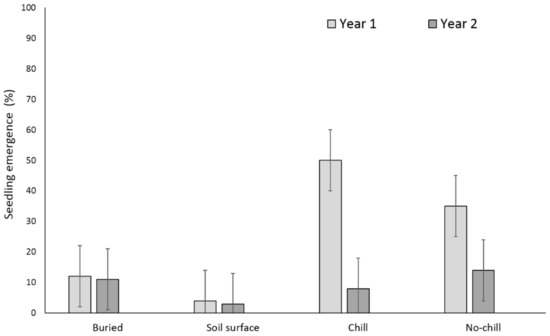
Figure 6.
Effect of environmental conditions on S. obtusifolia emergence originating from a multiple maternal source. Vertical bars represent LSD at 5% significance level.
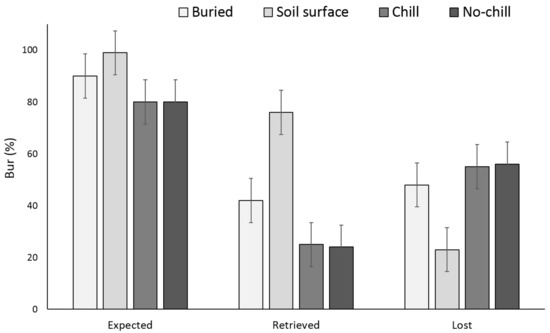
Figure 7.
Effect of environmental conditions on X. strumarium bur loss originating from a multiple maternal source after two years. Vertical bars represent LSD at 5% significance level. Expected” is the sum of non-germinated seeds.
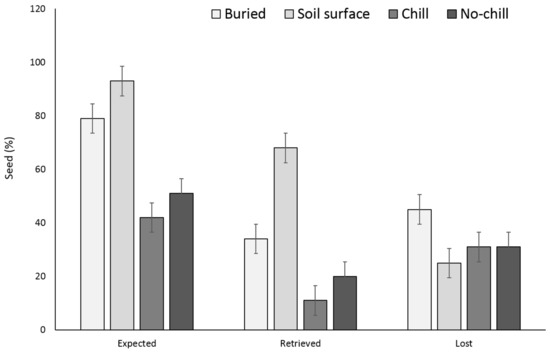
Figure 8.
Effect of environmental conditions on S. obtusifolia seed loss originating from a multiple maternal source after two years. Vertical bars represent LSD at 5% significance level. Expected” is the sum of non-germinated seeds.
Single maternal seed source buried in the soil had approximately 75% germination after the first scarification (Table 1); however, seeds on the surface had more resistant seed coats with only 30% germination (Table 1). A second scarification doubled germination of the seeds remained on the soil surface.
On the contrary, the multiple maternal seed source exhibited a greater variation in seed germination after one scarification (Table 2). Results from both experiments indicated that seedling emergence was lower when S. obtusifolia seeds and X. strumarium burs were buried at 5-cm depth than when seeds/burs were stored at 18 to 24 °C (no-chill) or at 0 to 2 °C (chill) for six months (Figure 1, Figure 2, Figure 5, and Figure 6), which was possibly due to differing planting days of September and May for both of the treatments. The lower emergence was due to loss of seed viability and seed decay when seed material was buried hence exposed to seed predators and microorganism attack. X. strumarium seedling emergence was reduced from 58 to 1% from single maternal seed source (Figure 1) and from 16 to 0.7% from multiple maternal seed source (Figure 5) when burs were left on the soil surface as compared to burs that were buried in the soil. S. obtusifolia seedling emergence was reduced from 52 to 18% from single maternal seed source (Figure 2) and from 43 to 7% from multiple maternal seed source when seeds were left on the soil surface compared with seeds buried in the soil.
Table 2.
Percentage germination of S. obtusifolia seeds from multiple maternal sources under various environmental conditions.
4. Discussion
These results indicate that a no-till production system that does not favor seed burial may effectively reduce X. strumarium seedling emergence [16,19], and, to a lesser extent, S. obtusifolia emergence. The germination test indicated that 100% of the remaining X. strumarium burs were decayed, but 100% of S. obtusifolia seeds were viable. These results also indicate that the X. strumarium seedbank was depleted after three years, but the S. obtusifolia seedbank was not depleted and the remaining seeds were viable. Thus, S. obtusifolia seed remain viable in or on the soil longer than those of X. strumarium. An average of 36 and 33% of X. strumarium burs and S. obtusifolia seeds were lost after three years. The large number of seeds or burs that were lost during the experiment was probably due to decay and/or predation [20,21]. Scarification of S. obtusifolia seed indicates that there are different levels of seed coat hardness (Bararpour and Oliver 1997) [22]; the hardness may be due to a thin impermeable waxy gel covering the S. obtusifolia seed. The hard seed coat of S. obtusifolia seed (i.e., survival mechanism) [22] accounts for the long-term survival of S. obtusifolia seed in the soil seedbank.
The soil seedbank of X. strumarium, as shown in Figure 3 and Figure 7, can be declined dramatically within three years after the deposition of seed on the soil. This indicates the relatively short life span of X. strumarium burs, hence the seedbank persistence of X. strumarium, therefore timing for management implementation tactics against this species, especially these that have developed herbicide resistance.
X. strumarium achenes, as revealed by germination and viability tests, are very sensitive to decay, particularly these that are left on the soil surface (a simulated no-tillage cropping system) as their emergence was recorded at 1% in the first year and 0% for the second and third year (Figure 1). Therefore, no-tillage can be an effective management practice to disturb X. strumarium seedbank dynamics and reduce the population of this species. Similar results were observed by Korres et al. (2018) [23] when the seeds of Amaranthus palmeri and A. tuberculatus were placed at two depths (0 and 15 cm for three years.
On the contrary, the seedbank of S. obtusifolia, as shown in Figure 4, was not depleted. Germination and viability tests revealed the resistance of S. obtusifolia seed against decay, which was most probably due to the characteristic hard seed coat. Therefore, in the case of this species, no tillage is less effective management practice for the reduction of S. obtusifolia population. These results are indicative of changes cropping system might exert on weed composition. In support of these results, Sosnoskie et al. (2006) [24] stated the effects of no-tillage systems as opposed to conventional or minimum tillage on the composition and diversity of weed communities.
5. Conclusions-Practical Applications
No tillage cropping systems that are suitable for conservational agriculture practices can contribute significantly in X. strumarium control tactics through seedbank reductions. X. strumarium bur prickles prevented good soil contact under no-till and many burs remained on the soil surface therefore fail to germinate and progressively were exposed to decay. In contrast, the smooth waxy surface of the smaller S. obtusifolia seeds allowed for good soil contact, even in no-till systems. S. obtusifolia can be a more problematic species to control in comparison with X. strumarium, especially under no-tillage conditions.
Acknowledgments
We would like to thank Lawrence R. Oliver for his useful comments and guidance during the completion of this project.
Author Contributions
Taghi Bararpour conducted the experiments, collected and analyzed the data and wrote the manuscript. Nicholas E. Korres analyzed the data and wrote the paper.
Conflicts of Interest
No conflicts of interest are declared.
References
- Leon, R.G.; Ferrell, J.A.; Sellers, B.A. Seed production and control of sicklepod (Senna obtusifolia) and pitted morningglory (Ipomoea lacunosa) with 2,4-D, dicamba, and glyphosate combinations. Weed Technol. 2016, 30, 76–84. [Google Scholar] [CrossRef]
- Norsworthy, J.K.; Oliveira, M.J. Tillage and soybean canopy effects on common cocklebur (Xanthium strumarium) emergence. Weed Sci. 2007, 55, 474–480. [Google Scholar] [CrossRef]
- Anonymous. Sicklepods: Sicklepod (Senna obtusifolia), Foetid Senna (Senna tora) and Hairy Senna (Senna hirsuta). Restricted Invasive Plant. Department of Agriculture and Fisheries. Biosecurity Queensland. 2016. Available online: www.biosecurity.qld.gov.au (accessed on 27 December 2017).
- CABI. Invasive Species Compendium Online Data Sheet. Senna obtusifolia (sicklepod). CABI Publishing, 2011. Available online: www.cabi.org/isc (accessed on 27 December 2017).
- Buhler, D.D. Challenges and opportunities for integrated weed management. Weed Sci. 2002, 50, 273–280. [Google Scholar] [CrossRef]
- Cardina, J.; Webster, T.M.; Herms, C.P.; Regnier, E.E. Development of weed IPM: Levels of integration for weed management. In Expanding the Context of Weed Management; Buhler, D.D., Ed.; Haworth Press, Inc.: New York, NY, USA, 1999; pp. 239–267. [Google Scholar]
- Honda, Y. Ecological correlations between the persistence of the soil seed bank and several plant traits, including seed dormancy. Plant Ecol. 2008, 196, 301–309. [Google Scholar] [CrossRef]
- Ooi, M.K.J.; Auld, T.D.; Whelan, R.J. Distinguishing between persistence and dormancy in soil seed banks of three shrub species from fi re-prone southeastern Australia. J. Veg. Sci. 2007, 18, 405–412. [Google Scholar] [CrossRef]
- Egley, G.H.; Chandler, J.M. Germination and viability of weed seeds after 2.5 years in a 50-year buried seed study. Weed Sci. 1978, 26, 230–231. [Google Scholar]
- Forcella, F. Seedling emergence model for velvetleaf. Agron. J. 1993, 85, 929–933. [Google Scholar] [CrossRef]
- Vleeshouwers, L.M.; Kropff, M.J. Modelling field emergence patterns in arable weeds. New Phytol. 2000, 148, 445–457. [Google Scholar] [CrossRef]
- Dyer, W. Exploiting weed seed dormancy and germination requirements through agronomic practices. Weed Sci. 1995, 43, 498–503. [Google Scholar]
- Mulugeta, D.; Stoltenberg, D.E. Increased weed emergence and seedbank depletion by soil disturbance in a no-tillage system. Weed Sci. 1997, 45, 234–241. [Google Scholar]
- Webster, T.M.; Cardina, J.; Norquay, H.M. Tillage and seed depth effects on velvetleaf (Abutilon theophrasti) emergence. Weed Sci. 1998, 46, 76–82. [Google Scholar]
- Yenish, J.P.; Doll, J.D.; Buhler, D.D. Effects of tillage on vertical distribution and viability of weed seed in soil. Weed Sci. 1992, 40, 429–433. [Google Scholar]
- Bararpour, M.T.; Oliver, L.R. Effect of tillage and interference on common cocklebur (Xanthium strumarium) and sicklepod (Senna obtusifolia) population, seed production, and seedbank. Weed Sci. 1998, 46, 424–431. [Google Scholar]
- Norsworthy, J.K. Soybean canopy formation effects on pitted morningglory (Ipomoea lacunosa), common cocklebur (Xanthium strumarium), and sicklepod (Senna obtusifolia) emergence. Weed Sci. 2004, 52, 961–967. [Google Scholar] [CrossRef]
- Litch, M.A.; Al-Kaisi, M. Strip-tillage on seedbed soil temperature and other soil physical properties. Soil Tillage Res. 2005, 80, 233–249. [Google Scholar]
- Bararpour, M.T. Effects of Tillage and Interference on Sicklepod (Senna obtusifolia L.) and Common Cocklebur (Xanthium strumarium L.) Growth Population, Seed Production, and Seedbank Potential. Ph.D. Dissertation, University of Arkansas, Fayetteville, AR, USA, 1995. [Google Scholar]
- Cavers, P.B. Seed demography. Can. J. Bot. 1983, 61, 3578–3590. [Google Scholar] [CrossRef]
- Davis, A.S.; Anderson, K.I.; Hallett, S.G.; Renner, K.A. Weed seed mortality in soils with contrasting agricultural management histories. Weed Sci. 2006, 54, 291–297. [Google Scholar] [CrossRef]
- Bararpour, M.T.; Oliver, L.R. Survival mechanisms of sicklepod (Senna obtusifolia L.) seed. In Proceedings of the Southern Weed Science Society, Houston, TX, USA, 20–22 January 1997; p. 151. [Google Scholar]
- Korres, N.E.; Norsworthy, J.K.; Young, B.G.; Reynolds, D.B.; Johnson, W.G.; Conley, S.P.; Smeda, R.J.; Mueller, T.C.; Bagavathiannan, M.V. Seedbank persistence of Palmer amaranth and waterhemp in the mid-south United States. In Proceedings of the Weed Science Society of America, Annual Meeting, Arlington, VA, USA, 29 January–1 February 2008. [Google Scholar]
- Sosnoskie, L.M.; Herms, C.P.; Cardina, J. Weed seedbank community composition in a 35-yr-old tillage and rotation experiment. Weed Sci. 2006, 54, 263–273. [Google Scholar] [CrossRef]
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).