Evaluating the Effectiveness of Rhizobium Inoculants and Micronutrients as Technologies for Nepalese Common Bean Smallholder Farmers in the Real-World Context of Highly Variable Hillside Environments and Indigenous Farming Practices

Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site, Climate, and Soil
2.2. Experimental Design
2.3. Plant Materials, Rhizobium Strains, and Inoculum Preparation
2.4. Planting and Application of Micronutrients (B and Mo)
2.5. Sample Collection
2.6. Statistical Analysis
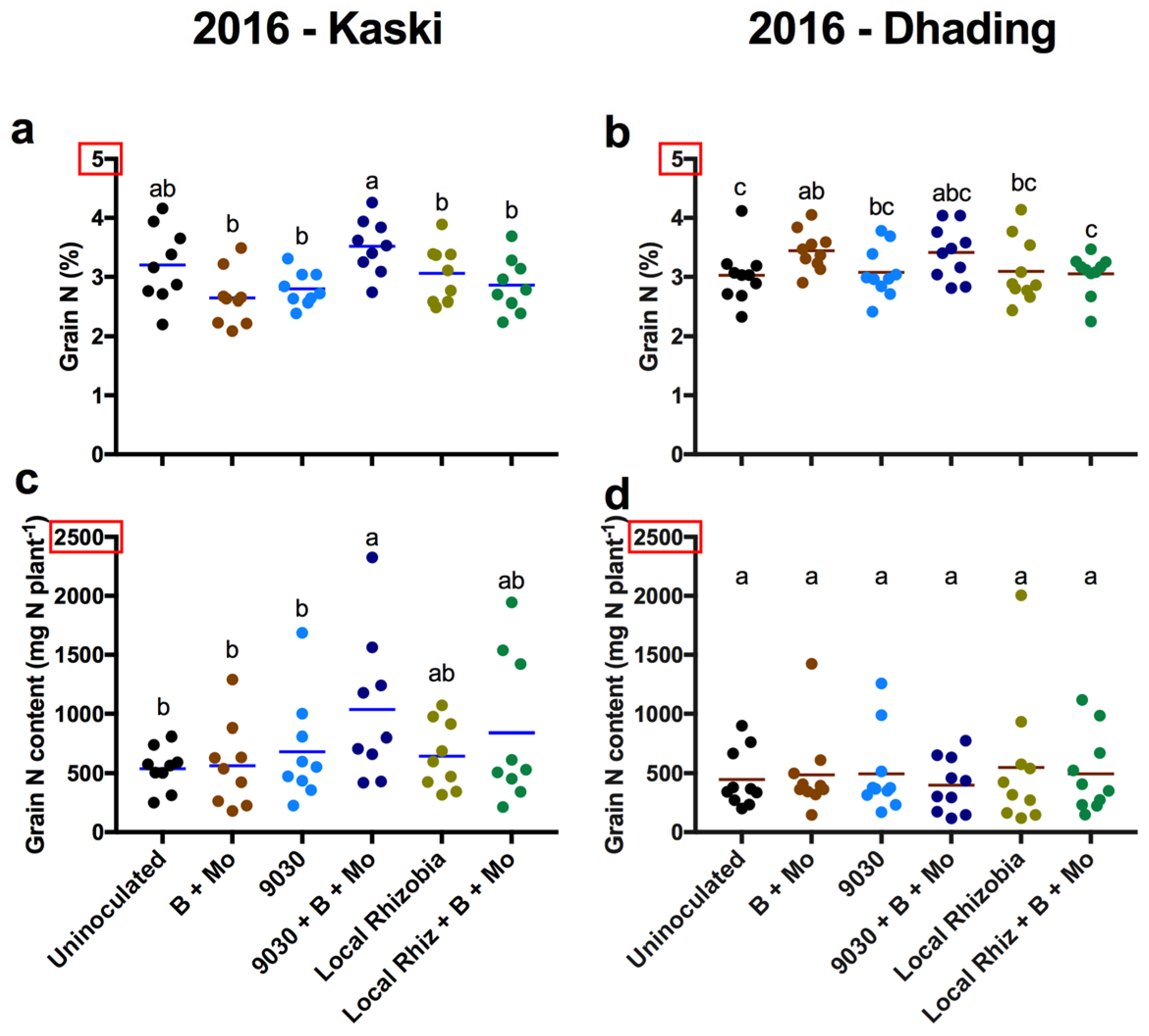
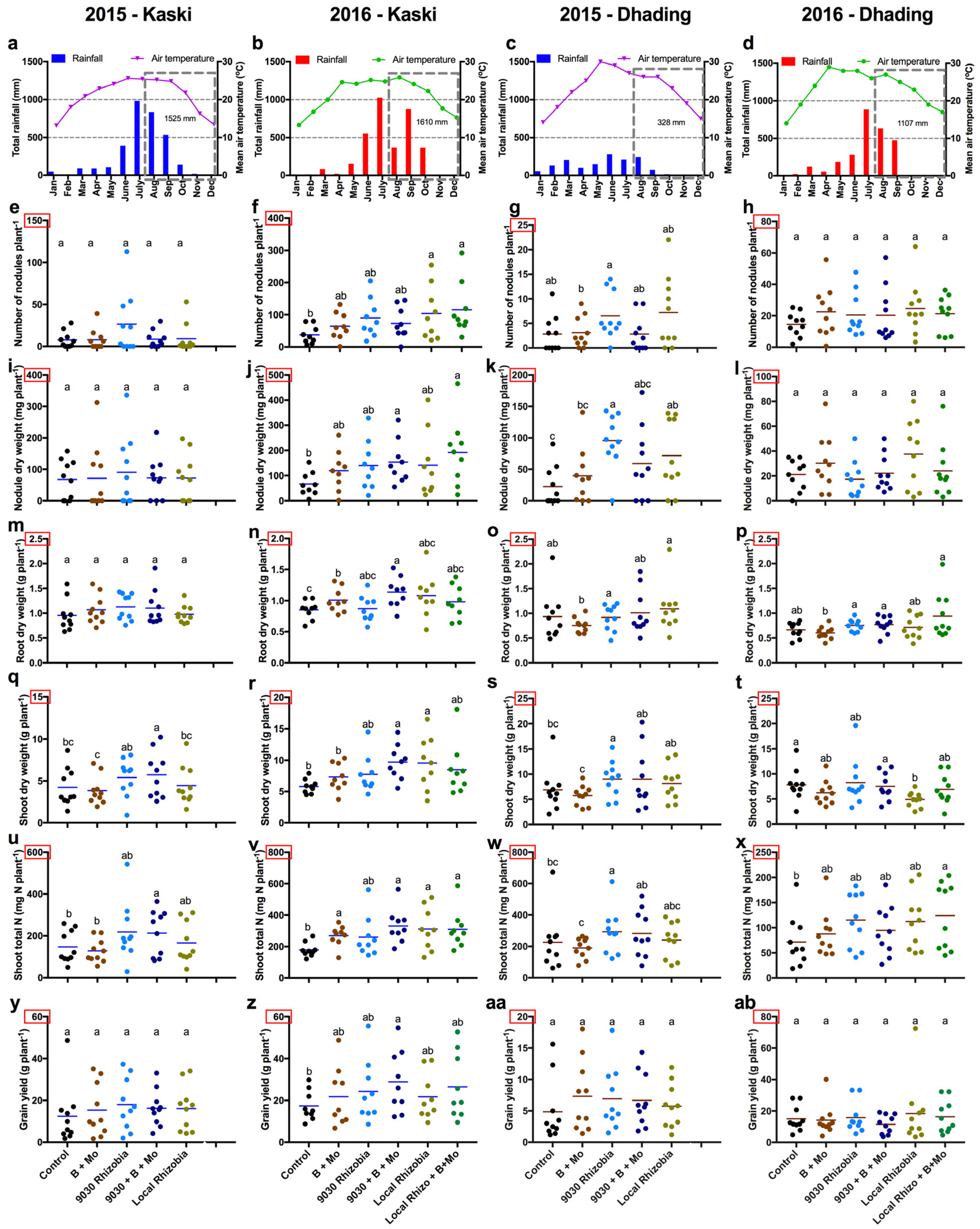
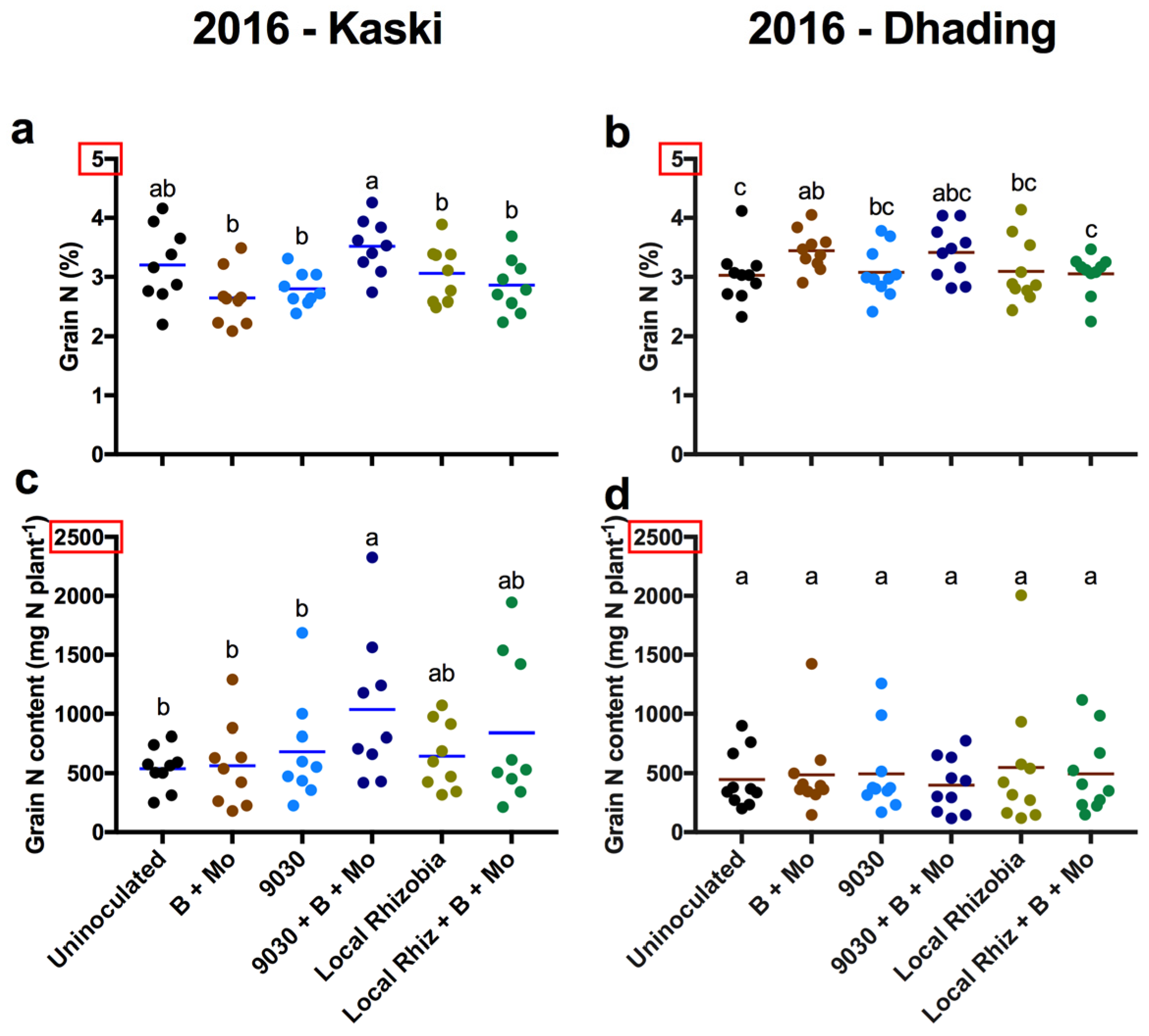
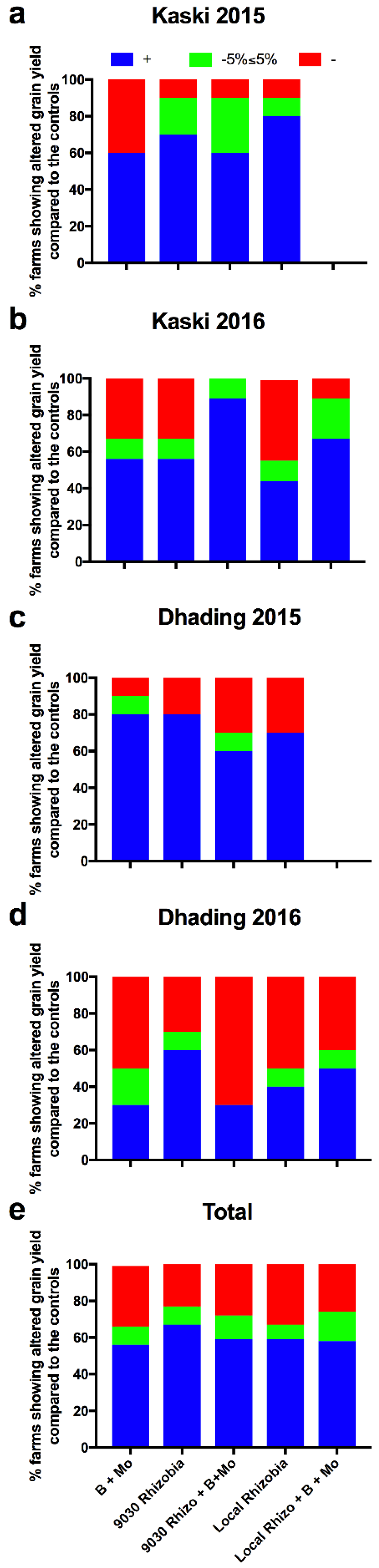
3. Results
3.1. Nodulation Data
3.2. General Effects of the Treatments on Agronomic Traits
3.3. Effects of the Treatments on Agronomic Traits at an Individual Farm Level
4. Discussion
4.1. Understanding Neutral or Negative Effects of Treatments
4.2. Understanding Variability between Trials: the Importance of Soil Moisture
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Leigh, G.J. The Worlds Greatest Fix: A History of Nitrogen and Agriculture; Oxford University Press: Oxford, UK, 2004. [Google Scholar]
- Foyer, C.H.; Lam, H.; Nguyen, H.T.; Siddique, K.H.M.; Varshney, R.K.; Colmer, T.D.; Cowling, W.; Bramley, H.; Mori, T.A.; Hodgson, J.M.; et al. Neglecting legumes has compromised human health and sustainable food production. Nat. Plants 2016, 2, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Mpepereki, S.; Javaheri, F.; Davis, P.; Giller, K.E. Soyabeans and sustainable agriculture Promiscuous soyabeans in southern Africa. Field Crops Res. 2000, 65, 137–149. [Google Scholar] [CrossRef]
- Graham, P.H.; Vance, C.P. Legumes: Importance and constraints to greater use. Plant Physiol. 2003, 131, 872–877. [Google Scholar] [CrossRef] [PubMed]
- Herridge, D.F.; Peoples, M.B.; Boddey, R.M. Global inputs of biological nitrogen fixation in agricultural systems. Plant Soil 2008, 311, 1–18. [Google Scholar] [CrossRef]
- Hauggaard-Nielsen, H.; Jørnsgaard, B.; Kinane, J.; Jensen, E.S. Grain legume-cereal intercropping: The practical application of diversity, competition and facilitation in arable and organic cropping systems. Renew. Agric. Food Syst. 2008, 23, 3–12. [Google Scholar] [CrossRef]
- Fustec, J.; Lesuffleur, F.; Mahieu, S.; Cliquet, J.B. Nitrogen rhizodeposition of legumes. A review. Agron. Sustain. Dev. 2010, 30, 57–66. [Google Scholar] [CrossRef]
- Thilakarathna, M.S.; Papadopoulos, Y.A.; Grimmett, M.; Fillmore, S.A.E.; Crouse, M.; Prithiviraj, B. Red clover varieties show nitrogen fixing advantage during the early stages of seedling development. Can. J. Plant Sci. 2018, 98, 1–10. [Google Scholar] [CrossRef]
- Thilakarathna, M.S.; Mcelroy, M.S.; Chapagain, T.; Papadopoulos, Y.A.; Raizada, M.N. Belowground nitrogen transfer from legumes to non-legumes under managed herbaceous cropping systems. A review. Agron. Sustain. Dev. 2016, 36, 58. [Google Scholar] [CrossRef]
- Chapagain, T.; Riseman, A. Barley—Pea intercropping: Effects on land productivity, carbon and nitrogen transformations. Field Crops Res. 2014, 166, 18–25. [Google Scholar] [CrossRef]
- Thilakarathna, M.S.; Papadopoulos, Y.A.; Rodd, A.V.; Grimmett, M.; Fillmore, S.A.E.; Crouse, M.; Prithiviraj, B. Nitrogen fixation and transfer of red clover genotypes under legume–grass forage based production systems. Nutr. Cycl. Agroecosyst. 2016, 106, 233–247. [Google Scholar] [CrossRef]
- Gan, Y.; Hamel, C.; O’Donovan, J.T.; Cutforth, H.; Zentner, R.P.; Campbell, C.A.; Niu, Y.; Poppy, L. Diversifying crop rotations with pulses enhances system productivity. Sci. Rep. 2015, 5, 14625. [Google Scholar] [CrossRef] [Green Version]
- Thilakarathna, M.S.; Raizada, M.N. A meta-analysis of the effectiveness of diverse rhizobia inoculants on soybean traits under field conditions. Soil Biol. Biochem. 2017, 105, 177–196. [Google Scholar] [CrossRef]
- Zahran, H.H. Rhizobium-legume symbiosis and nitrogen fixation under severe conditions and in an arid climate. Microbiol. Mol. Biol. Rev. 1999, 63, 968–989. [Google Scholar] [PubMed]
- Giller, K.E. Nitrogen Fixation in Tropical Cropping Systems; CABI: Wallingford, UK, 2001. [Google Scholar]
- Bolaños, L.; Esteban, E.; De Lorenzo, C.; Fernandez-Pascual, M.; De Felipe, M.R.; Garate, A.; Bonilla, I. Essentiality of boron for symbiotic dinitrogen fixation in pea (Pisum sativum) Rhizobium nodules. Plant Physiol. 1994, 104, 85–90. [Google Scholar] [CrossRef]
- Gepts, P.; Debouck, D. Origin, domestication, and evolution of the common bean (Phaseolus vulgaris L.). In Common Bean: Research for Crop Improvement; Schoonhoven, A., van Voysest, O., Eds.; CIAT: Cali, Colombia, 1991; pp. 7–53. [Google Scholar]
- Broughton, W.J.; Herna’ndez, G.; Blair, M.; Beebe, S.; Gepts, P.; Vanderleyden, J. Beans (Phaseolus spp.)—Model food legumes. Plant Soil 2003, 252, 55–128. [Google Scholar] [CrossRef]
- Graham, P.H.; Ranalli, P. Common bean (Phaseolus vulgaris L.). Field Crops Res. 1997, 53, 131–146. [Google Scholar] [CrossRef]
- Beebe, S.; Ramirez, J.; Jarvis, A.; Rao, I.M.; Mosquera, G.; Bueno, J.M.; Blair, M.W. Genetic improvement of common beans and the challenges of climate change. In Crop Adaptation to Climate Change; Yadav, S.S., Redden, R.J., Hatfield, J.L., Lotze-Campen, H., Hall, A.E., Eds.; John Wiley & Sons, Ltd.: Richmond, Australia, 2011; pp. 356–369. [Google Scholar]
- Bliss, F.A. Breeding common bean for improved biological nitrogen fixation. Plant Soil 1993, 152, 71–79. [Google Scholar] [CrossRef]
- Hardarson, G.; Atkins, C. Optimising biological N2 fixation by legumes in farming systems. Plant Soil 2003, 252, 41–54. [Google Scholar] [CrossRef]
- Rurangwa, E.; Vanlauwe, B.; Giller, K.E. Benefits of inoculation, P fertilizer and manure on yields of common bean and soybean also increase yield of subsequent maize. Agric. Ecosyst. Environ. 2018, 261, 219–229. [Google Scholar] [CrossRef] [PubMed]
- Chekanai, V.; Chikowo, R.; Vanlauwe, B. Response of common bean (Phaseolus vulgaris L.) to nitrogen, phosphorus and rhizobia inoculation across variable soils in Zimbabwe. Agric. Ecosyst. Environ. 2018, 266, 167–173. [Google Scholar] [CrossRef] [PubMed]
- Ndakidemi, P.A.; Dakora, F.D.; Nkonya, E.M.; Ringo, D.; Mansoor, H. Yield and economic benefits of common bean (Phaseolus vulgaris) and soybean (Glycine max) inoculation in northern Tanzania. Aust. J. Exp. Agric. 2006, 46, 571–577. [Google Scholar] [CrossRef]
- Da Silva, P.M.; Tsai, S.M.; Bonetti, R. Response to inoculation and N-fertilization for increased yield and biological nitrogen-fixation of common bean (Phaseolus vulgaris L.). Plant Soil 1993, 152, 123–130. [Google Scholar] [CrossRef]
- Chianu, J.; Nkonya, E.; Mairura, F.; Chianu, J.; Akinnifesi, F. Biological nitrogen fixation and socioeconomic factors for legume production in sub-Saharan Africa: Review. Agron. Sustain. Dev. 2011, 31, 139–154. [Google Scholar] [CrossRef]
- Musandu, A.A.O.; Joshua, O.O. Response of common beans to Rhizobium innoculation and fertilizers. J. Food Technol. Afr. 2001, 6, 121–125. [Google Scholar] [CrossRef]
- Giller, K.E. N2Africa Putting Nitrogen Fixation to Work for Smallholder Farmers in Africa, Podcaster No. 28, October, November, December 2014 (No. 28). N2Africa Project. 2014. Available online: https://n2africa.org/sites/default/files/N2Africa-Podcaster-28.pdf (accessed on 10 January 2019).
- Bambara, S.; Ndakidemi, P.A. Effects of Rhizobium inoculation, lime and molybdenum on photosynthesis and chlorophyll content of Phaseolus vulgaris L. Afr. J. Microbiol. Res. 2009, 3, 791–798. [Google Scholar]
- Silva, A.; Franzini, V.I.; Piccolla, C.D.; Muraoka, T. Molybdenum supply and biological fixation of nitrogen by two Brazilian common bean cultivars. Rev. Bras. Eng. Agrícola e Ambient. 2017, 21, 100–105. [Google Scholar] [CrossRef] [Green Version]
- Vieira, R.F.; Cardoso, E.J.B.N.; Vieira, C.; Cassini, S.T.A. Foliar application of molybdenum in common beans. I. Nitrogenase and reductase activities in a soil of high fertility. J. Plant Nutr. 1998, 21, 169–180. [Google Scholar] [CrossRef]
- Vieira, R.F.; Vieira, C.; Cardoso, E.J.B.N.; Mosquim, P.R. Foliar application of molybdenum in common bean. II. Nitrogenase and nitrate reductase activities in a soil of low fertility. J. Plant Nutr. 1998, 21, 2141–2151. [Google Scholar] [CrossRef]
- Ganie, M.A.; Akhter, F.; Bhat, M.A.; Najar, G.R. Growth, yield and quality of french bean (Phaseolus vulgaris L.) as influenced by sulphur and boron application on inceptisols of Kashmir. Bioscan 2014, 9, 513–518. [Google Scholar]
- Redondo-Nieto, M.; Wilmot, A.R.; El-Hamdaoui, A.; Bonilla, I.; Bolaños, L. Relationship between boron and calcium in the N2-fixing legume-rhizobia symbiosis. Plant Cell Environ. 2003, 26, 1905–1915. [Google Scholar] [CrossRef]
- Chapagain, T.; Raizada, M.N. Agronomic challenges and opportunities for smallholder terrace agriculture in developing countries. Front. Plant Sci. 2017, 8, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Small, F.A.A.; Raizada, M.N. Mitigating dry season food insecurity in the subtropics by prospecting drought-tolerant, nitrogen-fixing weeds. Agric. Food Secur. 2017, 6, 1–14. [Google Scholar] [CrossRef]
- Franke, A.C.; Baijukya, F.; Kantengwa, S.; Reckling, M.; Vanlauwe, B.; Giller, K.E. Poor farmers–poor yields: Socio-economic, soil fertility and crop management indicators affecting climbing bean productivity in northern Rwanda. Exp. Agric. 2016, 1–21. [Google Scholar] [CrossRef]
- Gharti, D.B.; Darai, R.; Subedi, S.; Sarker, A.; Kumar, S. Grain legumes in Nepal: Present scenario and future prospects. World J. Agric. Res. 2014, 2, 216–222. [Google Scholar] [CrossRef]
- Andersen, P. A review of micronutrient problems in the cultivated soil of Nepal. Mt. Res. Dev. 2007, 27, 331–335. [Google Scholar] [CrossRef]
- Anderson, J.M.; Ingram, J.S.I. Tropical Soil Biology and Fertility: A Handbook of Methods, 2nd ed.; CAB International: Wallingford, UK, 1993. [Google Scholar]
- Somasegaran, P.; Hoben, H.J. Handbook for Rhizobia. Methods in Legume–Rhizobium Technology; Springer: Heidelberg, Germany, 1994. [Google Scholar]
- van Berkum, P.; Beyene, D.; Eardly, B.D. Phylogenetic relationships among Rhizobium species nodulating the common bean (Phaseolus vulgaris L.). Int. J. Syst. Bacteriol. 1996, 46, 240–244. [Google Scholar] [CrossRef] [PubMed]
- Thilakarathna, M.S.; Raizada, M.N. A review of nutrient management studies involving finger millet in the Semi-Arid Tropics of Asia and Africa. Agronomy 2015, 5, 262–290. [Google Scholar] [CrossRef]
- Thilakarathna, M.S.; Raizada, M.N. Challenges in using precision agriculture to optimize symbiotic nitrogen fixation in legumes: Progress, limitations, and future improvements needed in diagnostic testing. Agronomy 2018, 8, 78. [Google Scholar] [CrossRef]
- Van Kessel, C.; Hartley, C. Agricultural management of grain legumes: Has it led to an increase in nitrogen fixation? Field Crops Res. 2000, 65, 165–181. [Google Scholar] [CrossRef]
- Sanz-Sáez, Á.; Heath, K.D.; Burke, P.V.; Ainsworth, E.A. Inoculation with an enhanced N2-fixing Bradyrhizobium japonicum strain (USDA110) does not alter soybean (Glycine max Merr.) response to elevated [CO2]. Plant Cell Environ. 2015, 38, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Hungria, M.; Vargas, M.A.T. Environmental factors affecting N2 fixation in grain legumes in the tropics, with an emphasis on Brazil. Field Crops Res. 2000, 65, 151–164. [Google Scholar] [CrossRef]
- Albareda, M.; Rodrı’guez-Navarro, D.N.; Temprano, F.J. Soybean inoculation: Dose, N fertilizer supplementation and rhizobia persistence in soil. Field Crops Res. 2009, 113, 352–356. [Google Scholar] [CrossRef]
- Liu, Y.; Wu, L.; Baddeley, J.A.; Watson, C.A. Models of biological nitrogen fixation of legumes. A review. Agron. Sustain. Dev. 2011, 31, 155–172. [Google Scholar] [CrossRef]
- van Heerwaarden, J.; Baijukya, F.; Kyei-Boahen, S.; Adjei-Nsiah, S.; Ebanyat, P.; Kamai, N.; Wolde-meskel, E.; Kanampiu, F.; Vanlauwe, B.; Giller, K. Soyabean response to rhizobium inoculation across sub-Saharan Africa: Patterns of variation and the role of promiscuity. Agric. Ecosyst. Environ. 2018, 261, 211–218. [Google Scholar] [CrossRef] [PubMed]
- Ronner, E.; Franke, A.C.; Vanlauwe, B.; Dianda, M.; Edeh, E.; Ukem, B.; Bala, A.; Van Heerwaarden, J.; Giller, K.E. Understanding variability in soybean yield and response to P-fertilizer and rhizobium inoculants on farmers’ fields in northern Nigeria. Field Crops Res. 2016, 186, 133–145. [Google Scholar] [CrossRef]
- Bogacki, P.; Peck, D.M.; Nair, R.M.; Howie, J.; Oldach, K.H. Genetic analysis of tolerance to Boron toxicity in the legume Medicago truncatula. BMC Plant Biol. 2013, 13, 54. [Google Scholar] [CrossRef] [PubMed]
- Bagheri, A.; Paull, J.G.; Rathjen, A.J.; Ali, S.M.; Moody, D.B. Genetic variation in the response of pea (Pisum sativum L.) to high soil concentrations of boron. Plant Soil 1992, 146, 261–269. [Google Scholar] [CrossRef]
- Ribet, J.; Drevon, J.J. The phosphorus requirement of N2-fixing and urea-fed Acacia mangium. New Phytol. 1996, 132, 383–390. [Google Scholar] [CrossRef]
- Puppo, A.; Groten, K.; Bastian, F.; Carzaniga, R.; Soussi, M.; Lucas, M.M.; De Felipe, M.R.; Harrison, J.; Vanacker, H.; Foyer, C.H. Legume nodule senescence: Roles for redox and hormone signalling in the orchestration of the natural aging process. New Phytol. 2005, 165, 683–701. [Google Scholar] [CrossRef]
- Chapagain, T.; Pudasaini, R.; Ghimire, B.; Gurung, K.; Choi, K.; Rai, L.; Magar, S.; BK, B.; Raizada, M.N. Intercropping of maize, millet, mustard, wheat and ginger increased land productivity and potential economic returns for smallholder terrace farmers in Nepal. Field Crops Res. 2018, 227, 91–101. [Google Scholar] [CrossRef]
- Adhikari, D.; Kaneto, M.; Itoh, K.; Suyama, K.; Pokharel, B.B.; Gaihre, Y.K. Genetic diversity of soybean-nodulating rhizobia in Nepal in relation to climate and soil properties. Plant Soil 2012, 357, 131–145. [Google Scholar] [CrossRef]
- Adhikari, D.; Itoh, K.; Suyama, K. Genetic diversity of common bean (Phaseolus vulgaris L.) nodulating rhizobia in Nepal. Plant Soil 2013, 368, 341–353. [Google Scholar] [CrossRef]
- Baral, B.; Gupta, V.N. Symbiotic properties of rhizobial isolates of Manang and Kathmandu districts. Nepal J. Sci. Technol. 2010, 11, 95–100. [Google Scholar] [CrossRef]
- Risal, C.P.; Djedidi, S.; Dhakal, D.; Ohkama-Ohtsu, N.; Sekimoto, H.; Yokoyama, T. Phylogenetic diversity and symbiotic functioning in mungbean (Vigna radiata L. Wilczek) bradyrhizobia from contrast agro-ecological regions of Nepal. Syst. Appl. Microbiol. 2012, 35, 45–53. [Google Scholar] [CrossRef] [PubMed]
- Risal, C.P.; Yokoyama, T.; Ohkama-Ohtsu, N.; Djedidi, S.; Sekimoto, H. Genetic diversity of native soybean bradyrhizobia from different topographical regions along the southern slopes of the Himalayan Mountains in Nepal. Syst. Appl. Microbiol. 2010, 33, 416–425. [Google Scholar] [CrossRef] [PubMed]
- Zengeni, R.; Mpepereki, S.; Giller, K.E. Manure and soil properties affect survival and persistence of soyabean nodulating rhizobia in smallholder soils of Zimbabwe. Appl. Soil Ecol. 2006, 32, 232–242. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site/Year | Parameter | B + Mo (T2) | 9030 Rhizobia (T3) | 9030 Rhizobia + B + Mo (T4) | Local Rhizobia (T5) | Local Rhizobia + B + Mo (T6) | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| + | - | No Effect | + | - | No Effect | + | - | No Effect | + | - | No Effect | + | - | No Effect | ||
| Total | Nodule # | 20 | 14 | 5 | 25 | 7 | 7 | 17 | 18 | 4 | 22 | 14 | 3 | 12 | 6 | 1 |
| N = 39 | Root DWT | 20 | 15 | 4 | 20 | 14 | 5 | 26 | 10 | 3 | 23 | 11 | 1 | 13 | 6 | 0 |
| Shoot DWT | 16 | 21 | 2 | 24 | 13 | 2 | 29 | 8 | 2 | 22 | 14 | 3 | 10 | 9 | 0 | |
| Shoot total N | 22 | 10 | 7 | 29 | 9 | 1 | 29 | 6 | 4 | 25 | 6 | 8 | 15 | 3 | 1 | |
| Grain total N | 9 | 9 | 1 | 10 | 7 | 2 | 11 | 8 | 0 | 9 | 8 | 2 | 10 | 5 | 4 | |
| Grain yield | 22 | 13 | 4 | 26 | 9 | 4 | 23 | 11 | 5 | 23 | 13 | 3 | 11 | 5 | 3 | |
| Kaski | Nodule # | 4 | 4 | 2 | 6 | 1 | 3 | 3 | 6 | 1 | 4 | 5 | 1 | n/a | n/a | n/a |
| 2015 | Root DWT | 7 | 2 | 1 | 8 | 2 | 0 | 6 | 3 | 1 | 4 | 4 | 2 | n/a | n/a | n/a |
| N = 10 | Shoot DWT | 4 | 5 | 1 | 6 | 3 | 1 | 9 | 0 | 1 | 7 | 1 | 2 | n/a | n/a | n/a |
| Shoot total N | 3 | 5 | 2 | 7 | 3 | 0 | 8 | 0 | 2 | 7 | 2 | 1 | n/a | n/a | n/a | |
| Grain yield | 6 | 4 | 0 | 7 | 1 | 2 | 6 | 1 | 3 | 8 | 1 | 1 | n/a | n/a | n/a | |
| Kaski | Nodule # | 7 | 2 | 0 | 6 | 2 | 1 | 6 | 3 | 0 | 7 | 2 | 0 | 6 | 3 | 0 |
| 2016 | Root DWT | 6 | 1 | 2 | 3 | 6 | 0 | 9 | 0 | 0 | 7 | 2 | 0 | 6 | 3 | 0 |
| N = 9 | Shoot DWT | 6 | 2 | 1 | 7 | 2 | 0 | 9 | 0 | 0 | 6 | 2 | 1 | 6 | 3 | 0 |
| Shoot total N | 9 | 0 | 0 | 6 | 2 | 1 | 9 | 0 | 0 | 7 | 1 | 1 | 8 | 1 | 0 | |
| Grain total N | 3 | 6 | 0 | 4 | 5 | 0 | 8 | 1 | 0 | 4 | 4 | 1 | 5 | 1 | 3 | |
| Grain yield | 5 | 3 | 1 | 5 | 3 | 1 | 8 | 0 | 1 | 4 | 4 | 1 | 6 | 1 | 2 | |
| Dhading | Nodule # | 3 | 4 | 3 | 6 | 1 | 3 | 4 | 4 | 2 | 5 | 3 | 2 | n/a | n/a | n/a |
| 2015 | Root DWT | 3 | 6 | 1 | 3 | 4 | 3 | 4 | 4 | 2 | 7 | 2 | 1 | n/a | n/a | n/a |
| N = 10 | Shoot DWT | 4 | 6 | 0 | 8 | 2 | 0 | 6 | 3 | 1 | 8 | 2 | 0 | n/a | n/a | n/a |
| Shoot total N | 5 | 4 | 1 | 8 | 2 | 0 | 7 | 3 | 0 | 6 | 3 | 1 | n/a | n/a | n/a | |
| Grain yield | 8 | 1 | 1 | 8 | 2 | 0 | 6 | 3 | 1 | 7 | 3 | 0 | n/a | n/a | n/a | |
| Dhading | Nodule # | 6 | 4 | 0 | 7 | 3 | 0 | 4 | 5 | 1 | 6 | 4 | 0 | 6 | 3 | 1 |
| 2016 | Root DWT | 4 | 6 | 0 | 6 | 2 | 2 | 7 | 3 | 0 | 5 | 5 | 0 | 7 | 3 | 0 |
| N = 10 | Shoot DWT | 2 | 8 | 0 | 3 | 6 | 1 | 5 | 5 | 0 | 1 | 9 | 0 | 4 | 6 | 0 |
| Shoot total N | 5 | 1 | 4 | 8 | 2 | 0 | 5 | 3 | 2 | 5 | 0 | 5 | 7 | 2 | 1 | |
| Grain total N | 6 | 3 | 1 | 6 | 2 | 2 | 3 | 7 | 0 | 5 | 4 | 1 | 5 | 4 | 1 | |
| Grain yield | 3 | 5 | 2 | 6 | 3 | 1 | 3 | 7 | 0 | 4 | 5 | 1 | 5 | 4 | 1 | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Thilakarathna, M.S.; Chapagain, T.; Ghimire, B.; Pudasaini, R.; Tamang, B.B.; Gurung, K.; Choi, K.; Rai, L.; Magar, S.; BK, B.; et al. Evaluating the Effectiveness of Rhizobium Inoculants and Micronutrients as Technologies for Nepalese Common Bean Smallholder Farmers in the Real-World Context of Highly Variable Hillside Environments and Indigenous Farming Practices. Agriculture 2019, 9, 20. https://doi.org/10.3390/agriculture9010020
Thilakarathna MS, Chapagain T, Ghimire B, Pudasaini R, Tamang BB, Gurung K, Choi K, Rai L, Magar S, BK B, et al. Evaluating the Effectiveness of Rhizobium Inoculants and Micronutrients as Technologies for Nepalese Common Bean Smallholder Farmers in the Real-World Context of Highly Variable Hillside Environments and Indigenous Farming Practices. Agriculture. 2019; 9(1):20. https://doi.org/10.3390/agriculture9010020
Chicago/Turabian StyleThilakarathna, Malinda S., Tejendra Chapagain, Bhawana Ghimire, Roshan Pudasaini, Bir Bhahadur Tamang, Khem Gurung, Khem Choi, Laxmi Rai, Samjhana Magar, Bishnu BK, and et al. 2019. "Evaluating the Effectiveness of Rhizobium Inoculants and Micronutrients as Technologies for Nepalese Common Bean Smallholder Farmers in the Real-World Context of Highly Variable Hillside Environments and Indigenous Farming Practices" Agriculture 9, no. 1: 20. https://doi.org/10.3390/agriculture9010020
APA StyleThilakarathna, M. S., Chapagain, T., Ghimire, B., Pudasaini, R., Tamang, B. B., Gurung, K., Choi, K., Rai, L., Magar, S., BK, B., Gaire, S., & Raizada, M. N. (2019). Evaluating the Effectiveness of Rhizobium Inoculants and Micronutrients as Technologies for Nepalese Common Bean Smallholder Farmers in the Real-World Context of Highly Variable Hillside Environments and Indigenous Farming Practices. Agriculture, 9(1), 20. https://doi.org/10.3390/agriculture9010020