
Untangling Coastal Diversity: How Habitat Complexity Shapes Demersal and Benthopelagic Assemblages in NW Iberia
 , , ,
, , ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. BRUVS Apparatus and Sampling Protocol
2.3. Video Analysis
2.4. Data Analyses
3. Results
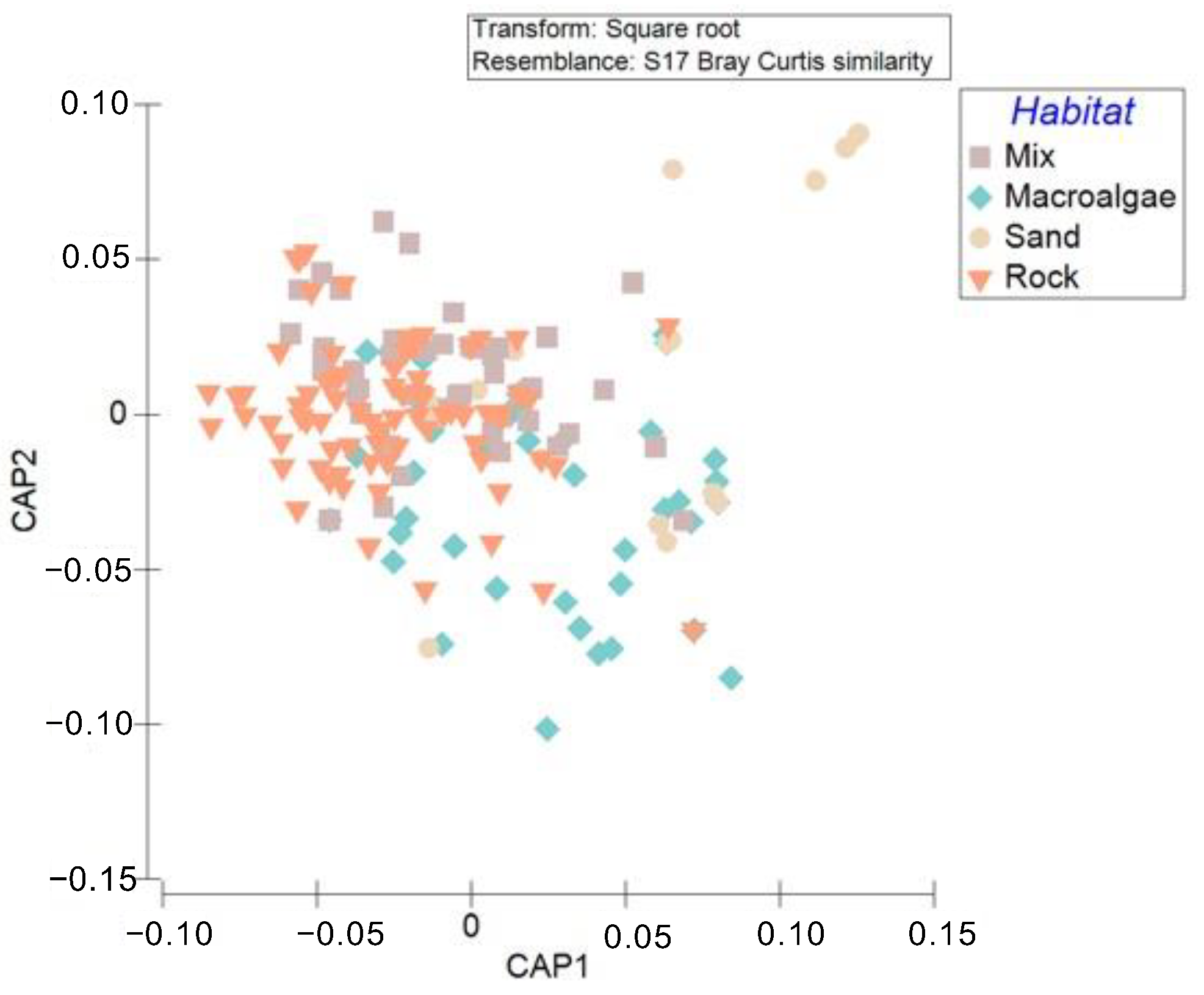
3.1. Taxonomic Diversity in Coastal Habitats
3.2. Demersal and Benthopelagic Assemblages Composition
3.3. Demersal and Benthopelagic Diversity Patterns
4. Discussion
4.1. Diversity of the Demersal and Benthopelagic Species in Coastal Habitats
4.2. Influence of Habitat Complexity on Biodiversity
4.3. Conservation Implications and Future Research Directions
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phylum | Family | Specie |
|---|---|---|
| Chordata | Gadidae | Trisopterus luscus |
| Chordata | Sparidae | Diplodus vulgaris |
| Chordata | Labridae | Ctenolabrus rupestris |
| Chordata | Moronidae | Dicentrarchus labrax |
| Chordata | Ammodytidae | Ammodytes tobianus |
| Chordata | Labridae | Labrus bergylta |
| Arthropoda | Portunidae | Carcinus maenas |
| Chordata | Labridae | Symphodus bailloni |
| Chordata | Serranidae | Serranus cabrilla |
| Chordata | Blenniidae | Parablennius gattorugine |
| Chordata | Sparidae | Diplodus sargus |
| Chordata | Labridae | Coris julis |
| Chordata | Labridae | Symphodus melops |
| Chordata | Mugilidae | Chelon auratus |
| Chordata | Congridae | Conger conger |
| Chordata | Mugilidae | Chelon labrosus |
| Chordata | Mullidae | Mullus surmuletus |
| Mollusca | Octopodidae | Octopus vulgaris |
| Chordata | Sparidae | Spondyliosoma cantharus |
| Chordata | Blenniidae | Parablennius pilicornis |
| Chordata | Sparidae | Sarpa salpa |
| Chordata | Triglidae | Chelidonichthys lucerna |
| Chordata | Phycidae | Phycis phycis |
| Arthropoda | Portunidae | Necora puber |
| Chordata | Ammodytidae | Hyperoplus lanceolatus |
| Chordata | Scophthalmidae | Zeugopterus punctatus |
| Arthropoda | Nephropidae | Homarus gammarus |
| Chordata | Rajidae | Raja undulata |
| Mollusca | Sepiidae | Sepia officinalis |
| Chordata | Sparidae | Diplodus cervinus |
| Chordata | Balistidae | Balistes capriscus |
| Estimate Std. | Error z | Z Value | Pr (>|z|) | |
|---|---|---|---|---|
| Species richness | ||||
| Intercept | 0.30250 | 0.50691 | 0.597 | 0.5507 |
| Habitat sand | −0.43151 | 0.20374 | −2.118 | 0.0342 |
| Habitat mix | 0.05181 | 0.15057 | 0.344 | 0.7308 |
| Habitat rock | 0.26723 | 0.13000 | 2.056 | 0.0398 |
| TRI 1.12 | 1.14071 | 0.56890 | 2.005 | 0.0449 |
| TRI 2.62 | 1.30535 | 0.56406 | 2.314 | 0.0207 |
| TRI 3.75 | 1.27210 | 0.54090 | 2.352 | 0.0187 |
| TRI 8.125 | 1.50971 | 0.61659 | 2.449 | 0.0143 |
| Pair-wise habitat | ||||
| sand-algae | −0.43151 | 0.20374 | −2.118 | 0.14277 |
| mix-algae | 0.05181 | 0.15057 | 0.344 | 0.98559 |
| rock-algae | 0.26723 | 0.13000 | 2.056 | 0.16272 |
| mix-sand | 0.48332 | 0.20807 | 2.323 | 0.08950 |
| rock-sand | 0.69874 | 0.20339 | 3.435 | 0.00309 |
| rock-mix | 0.21541 | 0.15087 | 1.428 | 0.47421 |
| Abundance | ||||
| Intercept | 0.57574 | 0.37098 | 1.552 | 0.123209 |
| Habitat sand | −0.22525 | 0.17066 | −1.320 | 0.189301 |
| Habitat mix | 0.24731 | 0.14804 | 1.671 | 0.097306 |
| Habitat rock | 0.38228 | 0.13024 | 2.935 | 0.003968 |
| TRI 1.12 | 1.81298 | 0.47500 | 3.817 | 0.000212 |
| TRI 2.62 | 0.96752 | 0.46352 | 2.087 | 0.038887 |
| TRI 3.75 | 1.31991 | 0.43299 | 3.048 | 0.002809 |
| TRI 5.75 | 0.85355 | 0.42902 | 1.990 | 0.048826 |
| TRI 7.625 | 1.29056 | 0.64736 | 1.994 | 0.048375 |
| TRI 8.12 | 1.30965 | 0.64696 | 2.024 | 0.045069 |
| Pair-wise habitat | ||||
| sand-algae | −0.2252 | 0.1707 | −1.320 | 0.54661 |
| mix-algae | 0.2473 | 0.1480 | 1.671 | 0.33584 |
| rock-algae | 0.3823 | 0.1302 | 2.935 | 0.01712 |
| mix-sand | 0.4726 | 0.1803 | 2.621 | 0.04310 |
| rock-sand | 0.6075 | 0.1723 | 3.526 | 0.00232 |
| rock-mix | 0.1350 | 0.1475 | 0.915 | 0.79443 |
| Simpson diversity | ||||
| Intercept | 0.260279 | 0.269267 | 0.967 | 0.335600 |
| Habitat sand | −0.436706 | 0.129872 | −3.363 | 0.001025 |
| Habitat mix | −0.002609 | 0.117799 | −0.022 | 0.982366 |
| Habitat rock | 0.291935 | 0.103534 | 2.820 | 0.005591 |
| TRI 0.75 | 0.947355 | 0.350629 | 2.702 | 0.007853 |
| TRI 1 | 0.948522 | 0.372574 | 2.546 | 0.012116 |
| TRI 1.12 | 0.921093 | 0.372186 | 2.475 | 0.014670 |
| TRI 1.25 | 0.928816 | 0.372287 | 2.495 | 0.013904 |
| TRI 2 | 0.886297 | 0.309524 | 2.863 | 0.004916 |
| TRI 2.25 | 1.039249 | 0.334634 | 3.106 | 0.002350 |
| TRI 2.37 | 0.727429 | 0.306913 | 2.370 | 0.019311 |
| TRI 2.5 | 1.020710 | 0.372287 | 2.742 | 0.007009 |
| TRI 2.62 | 1.372803 | 0.351153 | 3.909 | 0.000151 |
| TRI 3.125 | 0.798455 | 0.323704 | 2.467 | 0.014994 |
| TRI 3.25 | 1.094760 | 0.375573 | 2.915 | 0.004217 |
| TRI 3.5 | 0.717641 | 0.348111 | 2.062 | 0.041325 |
| TRI 3.75 | 1.208433 | 0.328587 | 3.678 | 0.000348 |
| TRI 4.375 | 0.852410 | 0.375573 | 2.270 | 0.024944 |
| TRI 4.75 | 1.157256 | 0.528916 | 2.188 | 0.030531 |
| TRI 5 | 1.040476 | 0.418589 | 2.486 | 0.014251 |
| TRI 5.75 | 0.981637 | 0.323165 | 3.038 | 0.002904 |
| TRI 7.375 | 1.349159 | 0.526761 | 2.561 | 0.011618 |
| TRI 8.125 | 1.498214 | 0.526976 | 2.843 | 0.005221 |
| TRI 8.625 | 1.375141 | 0.526976 | 2.609 | 0.010174 |
| TRI 19.25 | 1.294577 | 0.532970 | 2.429 | 0.016563 |
| Pair-wise habitat | ||||
| sand-algae | −0.436706 | 0.129872 | −3.363 | 0.00417 |
| mix-algae | −0.002609 | 0.117799 | −0.022 | 1.00000 |
| rock-algae | 0.291935 | 0.103534 | 2.820 | 0.02461 |
| mix-sand | 0.434097 | 0.139524 | 3.111 | 0.00969 |
| rock-sand | 0.728641 | 0.132459 | 5.501 | <0.001 |
| rock-mix | 0.294544 | 0.118241 | 2.491 | 0.06038 |
| Evenness | ||||
| Intercept | −2.715 × 102 | 5.59 × 10−2 | −0.486 | 0.628065 |
| Habitat sand | 1.542 × 10−2 | 2.69 × 10−2 | 0.572 | 0.568514 |
| Habitat mix | −2.252× 10−2 | 2.44 × 10−2 | −0.921 | 0.358976 |
| Habitat rock | −3.808 × 10−2 | 2.15 × 10−2 | −1.771 | 0.078954 |
| TRI 1.125 | −2.295 × 10−1 | 7.72 × 10−2 | −2.969 | 0.003578 |
| TRI 5.375 | −2.989 × 10−1 | 8.84 × 10−1 | −3.378 | 0.000975 |
References
- Rönnbäck, P.; Kautsky, N.; Pihl, L.; Troell, M.; Söderqvist, T.; Wennhage, W. Ecosystem Goods and Services from Swedish Coastal Habitats: Identification, Valuation, and Implications of Ecosystem Shifts. AMBIO J. Hum. Environ. 2007, 36, 534–544. [Google Scholar] [CrossRef]
- Davidson, N.C.; d’ALaffoley, D.; Doody, J.P.; Way, L.S.; Gordon, J.; Key, R.; Pienkowski, M.W.; Mitchell, R.; Duff, K.L. Nature Conservation and Estuaries in Great Britain; Nature Conservancy Council: Peterborough, UK, 1991. [Google Scholar]
- ICES. ICES Draft ICES Science Plan (2009–2013); ICES: Copenhagen, Denmark, 2008. [Google Scholar]
- Seitz, R.D.; Wennhage, H.; Bergström, U.; Lipcius, R.N.; Ysebaert, T. Ecological Value of Coastal Habitats for Commercially and Ecologically Important Species. ICES J. Mar. Sci. 2014, 71, 648–665. [Google Scholar] [CrossRef]
- Choat, J.H. Fish Feeding and the Structure of Benthic Communities in Temperate Waters. Annu. Rev. Ecol. Syst. 1982, 13, 423–449. [Google Scholar] [CrossRef]
- Caddy, J.F. Why Do Assessments of Demersal Stocks Largely Ignore Habitat? ICES J. Mar. Sci. 2014, 71, 2114–2126. [Google Scholar] [CrossRef]
- Gwinn, D.C.; Allen, M.S.; Johnston, F.D.; Brown, P.; Todd, C.R.; Arlinghaus, R. Rethinking Length-based Fisheries Regulations: The Value of Protecting Old and Large Fish with Harvest Slots. Fish Fish. 2015, 16, 259–281. [Google Scholar] [CrossRef]
- Worm, B.; Hilborn, R.; Baum, J.K.; Branch, T.A.; Collie, J.S.; Costello, C.; Fogarty, M.J.; Fulton, E.A.; Hutchings, J.A.; Jennings, S.; et al. Rebuilding Global Fisheries. Science 2009, 325, 578–585. [Google Scholar] [CrossRef] [PubMed]
- De Raedemaecker, F.; Brophy, D.; O’Connor, I.; Comerford, S. Habitat Characteristics Promoting High Density and Condition of Juvenile Flatfish at Nursery Grounds on the West Coast of Ireland. J. Sea Res. 2012, 73, 7–17. [Google Scholar] [CrossRef]
- Rabaut, M.; Audfroid Calderón, M.; Van de Moortel, L.; van Dalfsen, J.; Vincx, M.; Degraer, S.; Desroy, N. The Role of Structuring Benthos for Juvenile Flatfish. J. Sea Res. 2013, 84, 70–76. [Google Scholar] [CrossRef]
- Evans, R.D.; Wilson, S.K.; Field, S.N.; Moore, J.A.Y. Importance of Macroalgal Fields as Coral Reef Fish Nursery Habitat in North-West Australia. Mar. Biol. 2014, 161, 599–607. [Google Scholar] [CrossRef]
- Wilson, S.K.; Depczynski, M.; Fisher, R.; Holmes, T.H.; O’Leary, R.A.; Tinkler, P. Habitat Associations of Juvenile Fish at Ningaloo Reef, Western Australia: The Importance of Coral and Algae. PLoS ONE 2010, 5, e15185. [Google Scholar] [CrossRef]
- Eggertsen, M.; Chacin, D.H.; Van Lier, J.; Eggertsen, L.; Fulton, C.J.; Wilson, S.; Halling, C.; Berkström, C. Seascape Configuration and Fine-Scale Habitat Complexity Shape Parrotfish Distribution and Function across a Coral Reef Lagoon. Diversity 2020, 12, 391. [Google Scholar] [CrossRef]
- Fulton, C.J.; Abesamis, R.A.; Berkström, C.; Depczynski, M.; Graham, N.A.J.; Holmes, T.H.; Kulbicki, M.; Noble, M.M.; Radford, B.T.; Tano, S.; et al. Form and Function of Tropical Macroalgal Reefs in the Anthropocene. Funct. Ecol. 2019, 33, 989–999. [Google Scholar] [CrossRef]
- Sievers, K.T.; McClure, E.C.; Abesamis, R.A.; Russ, G.R. Non-reef Habitats in a Tropical Seascape Affect Density and Biomass of Fishes on Coral Reefs. Ecol. Evol. 2020, 10, 13673–13686. [Google Scholar] [CrossRef] [PubMed]
- Bonsdorff, E.; Blomqvist, E.M. Biotic Coupling On Shallow Water Soft Bottoms—Examples from the Northern Baltic Sea. Oceanogr. Mar. Biol. Annu. Rev. 1993, 31, 153–176. [Google Scholar]
- Hall, A.E.; Kingsford, M.J. Habitat Type and Complexity Drive Fish Assemblages in a Tropical Seascape. J. Fish Biol. 2021, 99, 1364–1379. [Google Scholar] [CrossRef] [PubMed]
- Jones, G.P. Ecology of Rocky Reef Fish of North-eastern New Zealand: A Review. N. Z. J. Mar. Freshw. Res. 1988, 22, 445–462. [Google Scholar] [CrossRef]
- Navarrete-Fernández, T.; Landaeta, M.F.; Bustos, C.A.; Pérez-Matus, A. Nest Building and Description of Parental Care Behavior in a Temperate Reef Fish, Chromis Crusma (Pisces: Pomacentridae). Rev. Chil. Hist. Nat. 2014, 87, 30. [Google Scholar] [CrossRef]
- Navarte, M.; González, R.; Storero, L.; Fernández, M. Effects of Competition and Egg Predation on Shelter Use by Octopus Tehuelchus Females. Mar. Ecol. Prog. Ser. 2013, 482, 141–151. [Google Scholar] [CrossRef]
- Sayer, M.D.J.; Gibson, R.N.; Atkinson, R.J.A. Distribution and Density of Populations of Goldsinny Wrasse (Ctenolabrus rupestris) on the West Coast of Scotland. J. Fish Biol. 1993, 43, 157–167. [Google Scholar] [CrossRef]
- Friedlander, A.M.; Brown, E.K.; Jokiel, P.L.; Smith, W.R.; Rodgers, K.S. Effects of Habitat, Wave Exposure, and Marine Protected Area Status on Coral Reef Fish Assemblages in the Hawaiian Archipelago. Coral Reefs 2003, 22, 291–305. [Google Scholar] [CrossRef]
- Graham, N.A.J.; Nash, K.L. The Importance of Structural Complexity in Coral Reef Ecosystems. Coral Reefs 2013, 32, 315–326. [Google Scholar] [CrossRef]
- McClure, E.C.; Hoey, A.S.; Sievers, K.T.; Abesamis, R.A.; Russ, G.R. Relative Influence of Environmental Factors and Fishing on Coral Reef Fish Assemblages. Conserv. Biol. 2021, 35, 976–990. [Google Scholar] [CrossRef]
- Perspectives on Ecosystem-Based Approaches to the Management of Marine Resources. Mar. Ecol. Prog. Ser. 2004, 274, 269–303. [CrossRef]
- Harvey, E.S.; Cappo, M.; Kendrick, G.A.; McLean, D.L. Coastal Fish Assemblages Reflect Geological and Oceanographic Gradients Within An Australian Zootone. PLoS ONE 2013, 8, e80955. [Google Scholar] [CrossRef] [PubMed]
- Reis-Filho, J.A.; Schmid, K.; Harvey, E.S.; Giarrizzo, T. Coastal Fish Assemblages Reflect Marine Habitat Connectivity and Ontogenetic Shifts in an Estuary-Bay-Continental Shelf Gradient. Mar. Environ. Res. 2019, 148, 57–66. [Google Scholar] [CrossRef] [PubMed]
- Dias, J.M.A.; Jouanneau, J.M.; Gonzalez, R.; Araújo, M.F.; Drago, T.; Garcia, C.; Oliveira, A.; Rodrigues, A.; Vitorino, J.; Weber, O. Present Day Sedimentary Processes on the Northern Iberian Shelf. Prog. Oceanogr. 2002, 52, 249–259. [Google Scholar] [CrossRef]
- Rosa-Santos, P.; Veloso-Gomes, F.; Taveira-Pinto, F.; Silva, R.; Pais-Barbosa, J. Evolution of Coastal Works in Portugal and Their Interference with Local Morphodynamics. J. Coast. Res. 2009, 1, 757–761. [Google Scholar]
- Rey, D.; Álvarez-Iglesias, P.; Jo, M.F.A.; Bernabeu, A.M.; Comas, M.; Decastro, M.; Druet, M.; Silva, E.F.D.; Ferrín, A.; Gesteira, M.; et al. The NW Iberian Continental Shelf. Geol. Soc. Mem. 2014, 41, 91–108. [Google Scholar] [CrossRef]
- Veiga, P.; Rubal, M.; Sousa-Pinto, I. Structural Complexity of Macroalgae Influences Epifaunal Assemblages Associated with Native and Invasive Species. Mar. Environ. Res. 2014, 101, 115–123. [Google Scholar] [CrossRef] [PubMed]
- Lemos, R.T.; Pires, H.O. The Upwelling Regime off the West Portuguese Coast, 1941–2000: PORTUGUESE COASTAL UPWELLING. Int. J. Climatol. 2004, 24, 511–524. [Google Scholar] [CrossRef]
- Pérez, F.F.; Padín, X.A.; Pazos, Y.; Gilcoto, M.; Cabanas, M.; Pardo, P.C.; Doval, M.D.; Farina-Busto, L. Plankton Response to Weakening of the Iberian Coastal Upwelling. Glob. Change Biol. 2010, 16, 1258–1267. [Google Scholar] [CrossRef]
- Alvarez, I.; Gomez-Gesteira, M.; deCastro, M.; Lorenzo, M.N.; Crespo, A.J.C.; Dias, J.M. Comparative Analysis of Upwelling Influence between the Western and Northern Coast of the Iberian Peninsula. Cont. Shelf Res. 2011, 31, 388–399. [Google Scholar] [CrossRef]
- Priede, I.G.; Bagley, P.M.; Smith, A.; Creasey, S.; Merrett, N.R. Scavenging Deep Demersal Fishes of the Porcupine Seabight, North-East Atlantic: Observations by Baited Camera, Trap and Trawl. J. Mar. Biol. Assoc. U. K. 1994, 74, 481–498. [Google Scholar] [CrossRef]
- Cappo, M.; Harvey, E.; Malcolm, H.; Speare, P. Potential of Video Tecniques to Monitor Diversity, Abundance and Size of Fish in Studies of Marine Protected Areas. In Aquatic Protected Areas: What Works Bestand How Do We Know? Proceedings of the World Congress on Aquatic Protected Areas, Cairns, Australia, 14–17 August 2002; Australian Society of Fish Biology: Cairns, Austalia, 2003; pp. 455–464. [Google Scholar]
- Cappo, M.; Speare, P.; De’ath, G. Comparison of Baited Remote Underwater Video Stations (BRUVS) and Prawn (Shrimp) Trawls for Assessments of Fish Biodiversity in Inter-Reefal Areas of the Great Barrier Reef Marine Park. J. Exp. Mar. Biol. Ecol. 2004, 302, 123–152. [Google Scholar] [CrossRef]
- Whitmarsh, S.K.; Fairweather, P.G.; Huveneers, C. What Is Big BRUVver up to? Methods and Uses of Baited Underwater Video. Rev. Fish Biol. Fish. 2017, 27, 53–73. [Google Scholar] [CrossRef]
- Willis, T.J.; Babcock, R.C. A Baited Underwater Video System for the Determination of Relative Density of Carnivorous Reef Fish. Mar. Freshw. Res. 2000, 51, 755. [Google Scholar] [CrossRef]
- Airoldi, L.; Beck, M.W. Loss, Status and Trends for Coastal Marine Habitats of Europe. Oceanogr. Mar. Biol. Ann. Rev. 2007, 45, 345–405. Available online: https://www.scirp.org/reference/ReferencesPapers?ReferenceID=1728187 (accessed on 15 February 2024).
- Riley, J.S.; Degloria, D.S.; Elliot, D.S. Terrain Ruggedness Index That Quantifies Topographic Heterogeneity. Available online: https://www.researchgate.net/publication/259011943_A_Terrain_Ruggedness_Index_that_Quantifies_Topographic_Heterogeneity (accessed on 15 February 2024).
- QGIS QGIS.Org QGIS Geographic Information System. 2023. Available online: https://www.qgis.org/en/site/ (accessed on 15 February 2024).
- GDAL/OGR Contributors. Available online: https://github.com/OSGeo/gdal/blob/master/CITATION (accessed on 15 February 2024).
- Romoth, K.; Darr, A.; Papenmeier, S.; Zettler, M.L.; Gogina, M. Substrate Heterogeneity as a Trigger for Species Diversity in Marine Benthic Assemblages. Biology 2023, 12, 825. [Google Scholar] [CrossRef]
- Clarke, K.R.; Gorley, R.N. User Manual/Tutorial; Primer-E Ltd.: Plymouth, UK, 2006. [Google Scholar]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Package ‘Vegan’: Community Ecology Package. R Package Version 2.4–3. 2017. Available online: https://CRAN.R-Project.Org/Package=vegan (accessed on 20 November 2023).
- Flávio, H.; Seitz, R.; Eggleston, D.; Svendsen, J.C.; Støttrup, J. Hard-Bottom Habitats Support Commercially Important Fish Species: A Systematic Review for the North Atlantic Ocean and Baltic Sea. PeerJ 2023, 11, e14681. [Google Scholar] [CrossRef] [PubMed]
- Kovalenko, K.E.; Thomaz, S.M.; Warfe, D.M. Habitat Complexity: Approaches and Future Directions. Hydrobiologia 2012, 685, 1–17. [Google Scholar] [CrossRef]
- Wehkamp, S.; Fischer, P. Impact of Hard-Bottom Substrata on the Small-Scale Distribution of Fish and Decapods in Shallow Subtidal Temperate Waters. Helgol. Mar. Res. 2013, 67, 59–72. [Google Scholar] [CrossRef][Green Version]
- Wilhelmsson, D.; Malm, T.; Öhman, M.C. The Influence of Offshore Windpower on Demersal Fish. ICES J. Mar. Sci. 2006, 63, 775–784. [Google Scholar] [CrossRef]
- Hixon, M.A.; Menge, B.A. Species Diversity: Prey Refuges Modify the Interactive Effects of Predation and Competition. Theor. Popul. Biol. 1991, 39, 178–200. [Google Scholar] [CrossRef]
- Gotceitas, V.; Fraser, S.; Brown, J.A. Habitat Use by Juvenile Atlantic Cod (Gadus morhua) in the Presence of an Actively Foraging and Non-Foraging Predator. Mar. Biol. 1995, 123, 421–430. [Google Scholar] [CrossRef]
- Mesa, G.L.; Muccio, S.D.; Vacchi, M. Structure of a Mediterranean Cryptobenthic Fish Community and Its Relationships with Habitat Characteristics. Mar. Biol. 2006, 149, 149–167. [Google Scholar] [CrossRef]
- Macarthur, R.; Levins, R. The Limiting Similarity, Convergence, and Divergence of Coexisting Species. Am. Nat. 1967, 101, 377–385. [Google Scholar] [CrossRef]
- Murdoch, W.W.; Oaten, A. Predation and Population Stability. In Advances in Ecological Research; MacFadyen, A., Ed.; Academic Press: Cambridge, MA, USA, 1975; Volume 9, pp. 1–131. [Google Scholar]
- Almany, G.R. Differential Effects of Habitat Complexity, Predators and Competitors on Abundance of Juvenile and Adult Coral Reef Fishes. Oecologia 2004, 141, 105–113. [Google Scholar] [CrossRef]
- Holbrook, S.J.; Schmitt, R.J. The Combined Effects of Predation Risk and Food Reward on Patch Selection. Ecology 1988, 69, 125–134. [Google Scholar] [CrossRef]
- Pierce, C.L. Predator Avoidance, Microhabitat Shift, and Risk-Sensitive Foraging in Larval Dragonflies. Oecologia 1988, 77, 81–90. [Google Scholar] [CrossRef]
- Schneider, K.J. Dominance, Predation, and Optimal Foraging in White-Throated Sparrow Flocks. Ecology 1984, 65, 1820–1827. [Google Scholar] [CrossRef]
- Hilldén, N.-O. Territoriality and Reproductive Behaviour in the Goldsinny, Ctenolabrus rupestris L. Behav. Process. 1981, 6, 207–221. [Google Scholar] [CrossRef]
- Costello, M.J. Review of the Biology of Wrasse (Labridae: Pisces) in Northern Europe. Prog. Underw. Sci. 1991, 16, 29–51. [Google Scholar]
- Andersson, M.H.; Berggren, M.; Wilhelmsson, D.; Öhman, M.C. Epibenthic Colonization of Concrete and Steel Pilings in a Cold-Temperate Embayment: A Field Experiment. Helgol. Mar. Res. 2009, 63, 249–260. [Google Scholar] [CrossRef]
- Gjøsaeter, J. Distribution and Density of Goldsinny Wrasse (Ctenolabrus rupestris) (Labridae) in the Risør and Arendal Areas along the Norwegian Skagerrak Coast. Sarsia 2002, 87, 75–82. [Google Scholar] [CrossRef]
- Haines, L.J.; Côté, I.M. Homing Decisions Reveal Lack of Risk Perception by Caribbean Damselfish of Invasive Lionfish. Biol. Invasions 2019, 21, 1657–1668. [Google Scholar] [CrossRef]
- Shulman, M.J. Recruitment of Coral Reef Fishes: Effects of Distribution of Predators and Shelter. Ecology 1985, 66, 1056–1066. [Google Scholar] [CrossRef]
- Turgeon, K.; Robillard, A.; Goire, J.G.; Duclos, V.; Kramer, D.L. Functional Connectivity from a Reef FIsh Perspective: Behavioral Tactics for Moving in a Fragmented Landscape. Ecology 2010, 91, 3332–3342. [Google Scholar] [CrossRef] [PubMed]
- Cohen, D.M.; Inada, T.; Iwamoto, T.; Scialabba, N. Fishery and Aquaculture Economics and Policy Division. FAO Species Catalogue. Vol.10. Gadiform Fishes of the World (Order Gadiformes). An Annotated and Illustrated Catalogue of Cods, Hakes, Grenadiers and Other Gadiform Fishes Known to Date; FAO Fisheries Synopsis; FAO: Rome, Italy, 1990; ISBN 978-92-5-102890-2. [Google Scholar]
- Castro, N.; Costa, J.L.; Domingos, I.; Angélico, M.M. Trophic Ecology of a Coastal Fish Assemblage in Portuguese Waters. J. Mar. Biol. Assoc. U. K. 2013, 93, 1151–1161. [Google Scholar] [CrossRef]
- Garrido, S.; Santos, A.M.P.; Dos Santos, A.; Ré, P. Spatial Distribution and Vertical Migrations of Fish Larvae Communities off Northwestern Iberia Sampled with LHPR and Bongo Nets. Estuar. Coast. Shelf Sci. 2009, 84, 463–475. [Google Scholar] [CrossRef]
- Guerreiro, M.A.; Martinho, F.; Baptista, J.; Costa, F.; Pardal, M.Â.; Primo, A.L. Function of Estuaries and Coastal Areas as Nursery Grounds for Marine Fish Early Life Stages. Mar. Environ. Res. 2021, 170, 105408. [Google Scholar] [CrossRef] [PubMed]
- Reubens, J.T.; Degraer, S.; Vincx, M. Aggregation and Feeding Behaviour of Pouting (Trisopterus luscus) at Wind Turbines in the Belgian Part of the North Sea. Fish. Res. 2011, 108, 223–227. [Google Scholar] [CrossRef]
- Alonso-Fernández, A.; Domínguez-Petit, R.; Bao, M.; Rivas, C.; Saborido-Rey, F. Spawning Pattern and Reproductive Strategy of Female Pouting Trisopterus luscus (Gadidae) on the Galician Shelf of North-Western Spain. Aquat. Living Resour. 2008, 21, 383–393. [Google Scholar] [CrossRef]
- Olsen, E.; Hansen, C.; Nilsen, I.; Perryman, H.; Vikebø, F. Ecological Effects and Ecosystem Shifts Caused by Mass Mortality Events on Early Life Stages of Fish. Front. Mar. Sci. 2019, 6, 477196. [Google Scholar] [CrossRef]
- Dufour, V.; Cantou, M.; Lecomte, F. Identification of Sea Bass (Dicentrarchus labrax) Nursery Areas in the North-Western Mediterranean Sea. J. Mar. Biol. Assoc. U. K. 2009, 89, 1367–1374. [Google Scholar] [CrossRef]
- Moksnes, P.-O. The Relative Importance of Habitat-Specific Settlement, Predation and Juvenile Dispersal for Distribution and Abundance of Young Juvenile Shore Crabs Carcinus maenas L. J. Exp. Mar. Biol. Ecol. 2002, 271, 41–73. [Google Scholar] [CrossRef]
- Sambrook, K.; Hoey, A.S.; Andréfouët, S.; Cumming, G.S.; Duce, S.; Bonin, M.C. Beyond the Reef: The Widespread Use of Non-reef Habitats by Coral Reef Fishes. Fish Fish. 2019, 20, 903–920. [Google Scholar] [CrossRef]
- Bradley, M.; Baker, R.; Sheaves, M. Hidden Components in Tropical Seascapes: Deep-Estuary Habitats Support Unique Fish Assemblages. Estuaries Coasts 2017, 40, 1195–1206. [Google Scholar] [CrossRef]
- García-Charton, J.A.; Pérez-Ruzafa, Á. Spatial Pattern and the Habitat Structure of a Mediterranean Rocky Reef Fish Local Assemblage. Mar. Biol. 2001, 138, 917–934. [Google Scholar] [CrossRef]
- Gilby, B.L.; Tibbetts, I.R.; Olds, A.D.; Maxwell, P.S.; Stevens, T. Seascape Context and Predators Override Water Quality Effects on Inshore Coral Reef Fish Communities. Coral Reefs 2016, 35, 979–990. [Google Scholar] [CrossRef]
- Connell, S.D.; Jones, G.P. The Influence of Habitat Complexity on Postrecruitment Processes in a Temperate Reef Fish Population. J. Exp. Mar. Biol. Ecol. 1991, 151, 271–294. [Google Scholar] [CrossRef]
- Rice, J.; Arvanitidis, C.; Borja, A.; Frid, C.; Hiddink, J.G.; Krause, J.; Lorance, P.; Ragnarsson, S.Á.; Sköld, M.; Trabucco, B.; et al. Indicators for Sea-floor Integrity under the European Marine Strategy Framework Directive. Ecol. Indic. 2012, 12, 174–184. [Google Scholar] [CrossRef]
- Bryars, S.; Rogers, P. Movements in the Harlequin Fish (Othos dentex), a Large Predatory Serranid from Southern Australia: Implications for Detection and Protection. N. Z. J. Mar. Freshw. Res. 2016, 50, 600–618. [Google Scholar] [CrossRef]
- Martins, G.; Faria, J.; Rubal, M.; Neto, A.I. Linkages between Rocky Reefs and Soft-Bottom Habitats: Effects of Predation and Granulometry on Sandy Macrofaunal Assemblages. J. Sea Res. 2013, 81, 1–9. [Google Scholar] [CrossRef]




| Variable | Estimation Method | Levels and Definition |
|---|---|---|
| Habitat | Categorised during video analysis and based on the underlying habitat structure in the BRUVS field of view. | Rock: sea bottom with bedrock as the underlying substrate. Mix: areas where rocky reefs are intermingled with sand beds in equal proportions. Macroalgae formations: dominance of algae (primarily brown algae). Sand Bed: presence of a sandy substrate as the dominant underlying surface. |
| Topographic complexity (Terrain Ruggedness Index—TRI) | Calculated in software QGIS using GDAL tools. | Continuous; low values represent flat areas; high values represent rugged ones. |
| Habitat | |||||
|---|---|---|---|---|---|
| Phylum | Family | Sand | Macroalgae | Mix | Rock |
| Chordata | Labridae | 4 | 5 | 5 | 5 |
| Chordata | Sparidae | 1 | 3 | 3 | 4 |
| Chordata | Blenniidae | 1 | 2 | 2 | 2 |
| Chordata | Congridae | 1 | 1 | 1 | 1 |
| Chordata | Gadidae | 1 | 1 | 1 | 1 |
| Chordata | Moronidae | 1 | 1 | 1 | 1 |
| Mollusca | Octopodidae | 1 | 1 | 1 | 1 |
| Chordata | Mugilidae | - | 2 | 2 | 2 |
| Chordata | Serranidae | - | 1 | 1 | 1 |
| Arthropoda | Portunidae | 1 | - | 2 | - |
| Chordata | Ammodytidae | - | - | 1 | 2 |
| Chordata | Rajidae | - | 1 | - | - |
| Mollusca | Sepiidae | - | 1 | - | - |
| Arthropoda | Nephropidae | - | - | - | 1 |
| Chordata | Balistidae | - | - | 1 | - |
| Chordata | Physidae | - | - | - | 1 |
| Chordata | Scophthalmidae | - | - | - | 1 |
| Chordata | Triglidae | 1 | - | - | - |
| Chordata | Mullidae | - | - | 1 | - |
| Species Total | 12 | 19 | 22 | 23 | |
| Source | df | SS | MS | Pseudo-F | P (perm) | Unique Perms |
|---|---|---|---|---|---|---|
| Habitat | 3 | 37,232 | 12,411 | 4.3557 | 0.0001 | 9936 |
| Complexity | 66 | 1.8855 × 105 | 2856.8 | 1.0026 | 0.4828 | 9703 |
| Habitat*Complexity | 50 | 1.5309 × 105 | 3061.8 | 1.0746 | 0.2527 | 9761 |
| Residual | 75 | 2.137 × 105 | 2849.3 | |||
| Total | 194 | 6.1278 × 105 | ||||
| PERMDISP (Habitat) | F: 13.517 | P (perm): 0.0001 | ||||
| PERMDISP (Complexity) | F: 8.1968 | P (perm): 0.0001 | ||||
| Groups | t | P (Perm) | Unique Perms | |
|---|---|---|---|---|
| Habitat | Macroalgae, Sand | 1.9386 | 0.0024 | 9942 |
| Macroalgae, Mix | 1.3927 | 0.0818 | 9937 | |
| Macroalgae, Rock | 2.2907 | 0.0001 | 9937 | |
| Sand, Mix | 1.8025 | 0.0081 | 9929 | |
| Sand, Rock | 2.839 | 0.0001 | 9934 | |
| Mix, Rock | 0.74279 | 0.7512 | 9950 | |
| Species | Average Abundance | Av.Diss | Diss/SD | Contrib% | Cum% | |
|---|---|---|---|---|---|---|
| Macroalgae | Sand | |||||
| Carcinus maenas | 0.00 | 0.74 | 12.96 | 0.61 | 15.07 | 15.07 |
| Trisopterus luscus | 0.60 | 0.46 | 12.83 | 0.90 | 14.92 | 29.99 |
| Diplodus vulgaris | 0.85 | 0.41 | 12.69 | 1.00 | 14.76 | 44.75 |
| Dicentrarchus labrax | 0.53 | 0.31 | 10.58 | 0.80 | 12.30 | 57.05 |
| Symphodus melops | 0.25 | 0.04 | 5.61 | 0.51 | 6.52 | 63.57 |
| Ctenolabrus rupestris | 0.27 | 0.13 | 5.18 | 0.57 | 6.03 | 69.60 |
| Symphodus bailloni | 0.27 | 0.06 | 4.93 | 0.51 | 5.73 | 75.33 |
| Labrus bergylta | 0.23 | 0.13 | 4.76 | 0.55 | 5.53 | 80.86 |
| Parablennius gattorugine | 0.16 | 0.04 | 2.82 | 0.44 | 3.27 | 84.13 |
| Chelon auratus | 0.13 | 0.00 | 2.16 | 0.29 | 2.51 | 86.64 |
| Chelidonichthys lucerna | 0.00 | 0.08 | 1.84 | 0.27 | 2.14 | 88.78 |
| Conger conger | 0.05 | 0.04 | 1.61 | 0.26 | 1.88 | 90.66 |
| Macroalgae | Rock | |||||
| Trisopterus luscus | 0.60 | 1.16 | 13.29 | 1.11 | 17.24 | 17.24 |
| Diplodus vulgaris | 0.85 | 0.73 | 10.81 | 1.03 | 14.03 | 31.27 |
| Ctenolabrus rupestris | 0.27 | 0.81 | 10.00 | 1.03 | 12.97 | 44.24 |
| Dicentrarchus labrax | 0.53 | 0.13 | 6.64 | 0.76 | 8.61 | 52.85 |
| Labrus bergylta | 0.23 | 0.40 | 6.08 | 0.73 | 7.89 | 60.74 |
| Symphodus bailloni | 0.27 | 0.31 | 5.34 | 0.70 | 6.93 | 67.67 |
| Serranus cabrilla | 0.05 | 0.39 | 4.83 | 0.73 | 6.27 | 73.93 |
| Parablennius gattorugine | 0.16 | 0.27 | 4.33 | 0.63 | 5.61 | 79.54 |
| Symphodus melops | 0.25 | 0.06 | 4.23 | 0.52 | 5.49 | 85.04 |
| Chelon auratus | 0.13 | 0.04 | 1.97 | 0.33 | 2.55 | 87.59 |
| Conger conger | 0.05 | 0.08 | 1.55 | 0.32 | 2.01 | 89.60 |
| Coris julis | 0.05 | 0.07 | 1.27 | 0.31 | 1.65 | 91.25 |
| Sand | Mix | |||||
| Trisopterus luscus | 0.46 | 1.36 | 19.04 | 1.02 | 21.98 | 21.98 |
| Carcinus maenas | 0.74 | 0.02 | 12.29 | 0.60 | 14.20 | 36.18 |
| Diplodus vulgaris | 0.41 | 0.68 | 11.07 | 0.85 | 12.78 | 48.96 |
| Ctenolabrus rupestris | 0.13 | 0.57 | 7.79 | 0.79 | 8.99 | 57.95 |
| Dicentrarchus labrax | 0.31 | 0.22 | 7.39 | 0.56 | 8.53 | 66.49 |
| Labrus bergylta | 0.13 | 0.32 | 6.16 | 0.58 | 7.11 | 73.60 |
| Serranus cabrilla | 0.00 | 0.29 | 4.05 | 0.53 | 4.68 | 78.27 |
| Symphodus bailloni | 0.06 | 0.18 | 3.40 | 0.44 | 3.93 | 82.20 |
| Coris julis | 0.00 | 0.22 | 3.12 | 0.41 | 3.61 | 85.81 |
| Chelidonichthys lucerna | 0.08 | 0.00 | 1.73 | 0.26 | 1.99 | 87.80 |
| Parablennius gattorugine | 0.04 | 0.10 | 1.55 | 0.33 | 1.79 | 89.59 |
| Mullus surmuletus | 0.00 | 0.14 | 1.52 | 0.31 | 1.76 | 91.35 |
| Sand | Rock | |||||
| Trisopterus luscus | 0.46 | 1.16 | 15.89 | 1.18 | 18.66 | 18.66 |
| Ctenolabrus rupestris | 0.13 | 0.81 | 11.05 | 1.03 | 12.97 | 31.63 |
| Diplodus vulgaris | 0.41 | 0.73 | 10.97 | 0.90 | 12.88 | 44.51 |
| Carcinus maenas | 0.74 | 0.00 | 10.88 | 0.60 | 12.77 | 57.28 |
| Labrus bergylta | 0.13 | 0.40 | 6.59 | 0.71 | 7.73 | 65.01 |
| Dicentrarchus labrax | 0.31 | 0.13 | 5.83 | 0.56 | 6.84 | 71.85 |
| Serranus cabrilla | 0.00 | 0.39 | 5.33 | 0.72 | 6.26 | 78.11 |
| Symphodus bailloni | 0.06 | 0.31 | 4.36 | 0.61 | 5.12 | 83.23 |
| Parablennius gattorugine | 0.04 | 0.27 | 4.00 | 0.55 | 4.69 | 87.92 |
| Conger conger | 0.04 | 0.08 | 1.50 | 0.34 | 1.77 | 89.69 |
| Chelidonichthys lucerna | 0.08 | 0.00 | 1.49 | 0.27 | 1.75 | 91.44 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gomes, M.A.; Alves, C.M.; Faria, F.; Troncoso, J.S.; Gomes, P.T. Untangling Coastal Diversity: How Habitat Complexity Shapes Demersal and Benthopelagic Assemblages in NW Iberia. J. Mar. Sci. Eng. 2024, 12, 538. https://doi.org/10.3390/jmse12040538
Gomes MA, Alves CM, Faria F, Troncoso JS, Gomes PT. Untangling Coastal Diversity: How Habitat Complexity Shapes Demersal and Benthopelagic Assemblages in NW Iberia. Journal of Marine Science and Engineering. 2024; 12(4):538. https://doi.org/10.3390/jmse12040538
Chicago/Turabian StyleGomes, Marisa A., Catarina M. Alves, Fábio Faria, Jesus S. Troncoso, and Pedro T. Gomes. 2024. "Untangling Coastal Diversity: How Habitat Complexity Shapes Demersal and Benthopelagic Assemblages in NW Iberia" Journal of Marine Science and Engineering 12, no. 4: 538. https://doi.org/10.3390/jmse12040538
APA StyleGomes, M. A., Alves, C. M., Faria, F., Troncoso, J. S., & Gomes, P. T. (2024). Untangling Coastal Diversity: How Habitat Complexity Shapes Demersal and Benthopelagic Assemblages in NW Iberia. Journal of Marine Science and Engineering, 12(4), 538. https://doi.org/10.3390/jmse12040538