
Changes in Carbon, Nitrogen, and Oxygen Stable Isotope Ratios and Mercury Concentrations in Killer Whales (Orcinus orca) during and after Lactation

 , ,
, ,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Samples of Killer and Common Minke Whales
2.2. Chemical Analyses
2.3. Statistical Analyses
3. Results
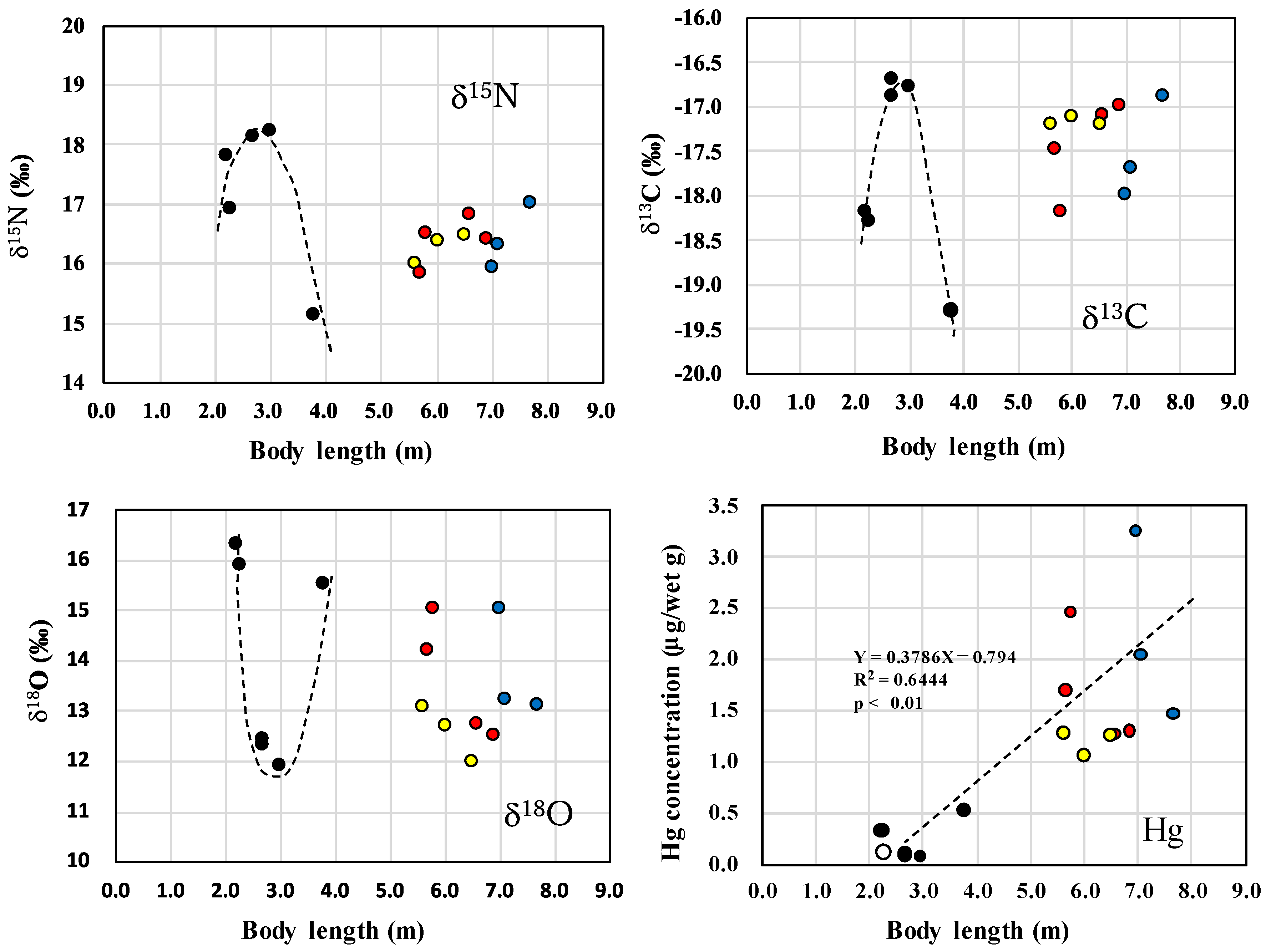
3.1. Changes in δ13C, δ15N, δ18O Values and Hg Concentrations during and after Lactation in Killer Whales
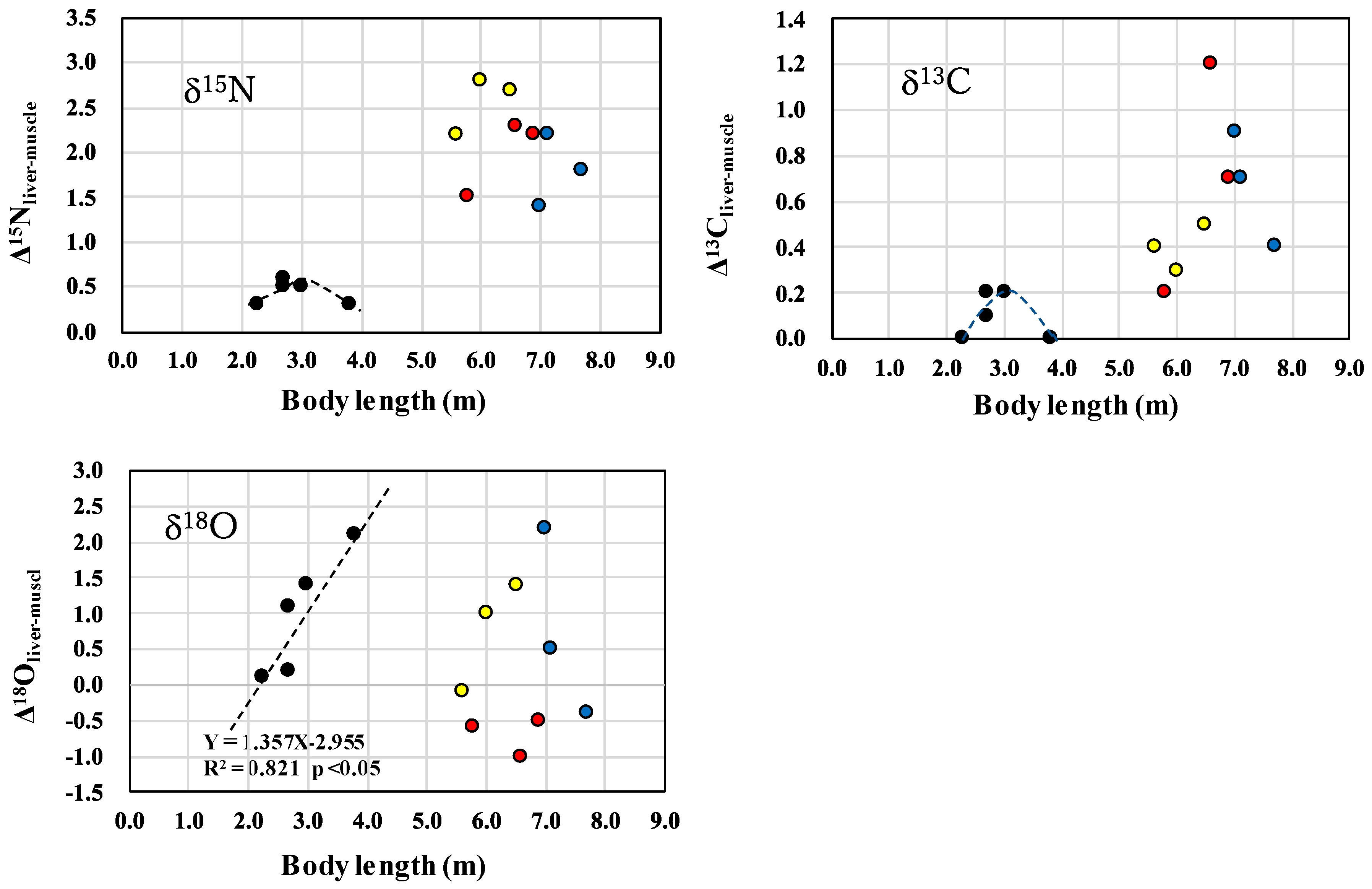
3.2. Comparison of δ13Cliver-muscle, δ15Nliver-muscle, and δ18Oliver-muscle Levels in Calves and Mature Animals
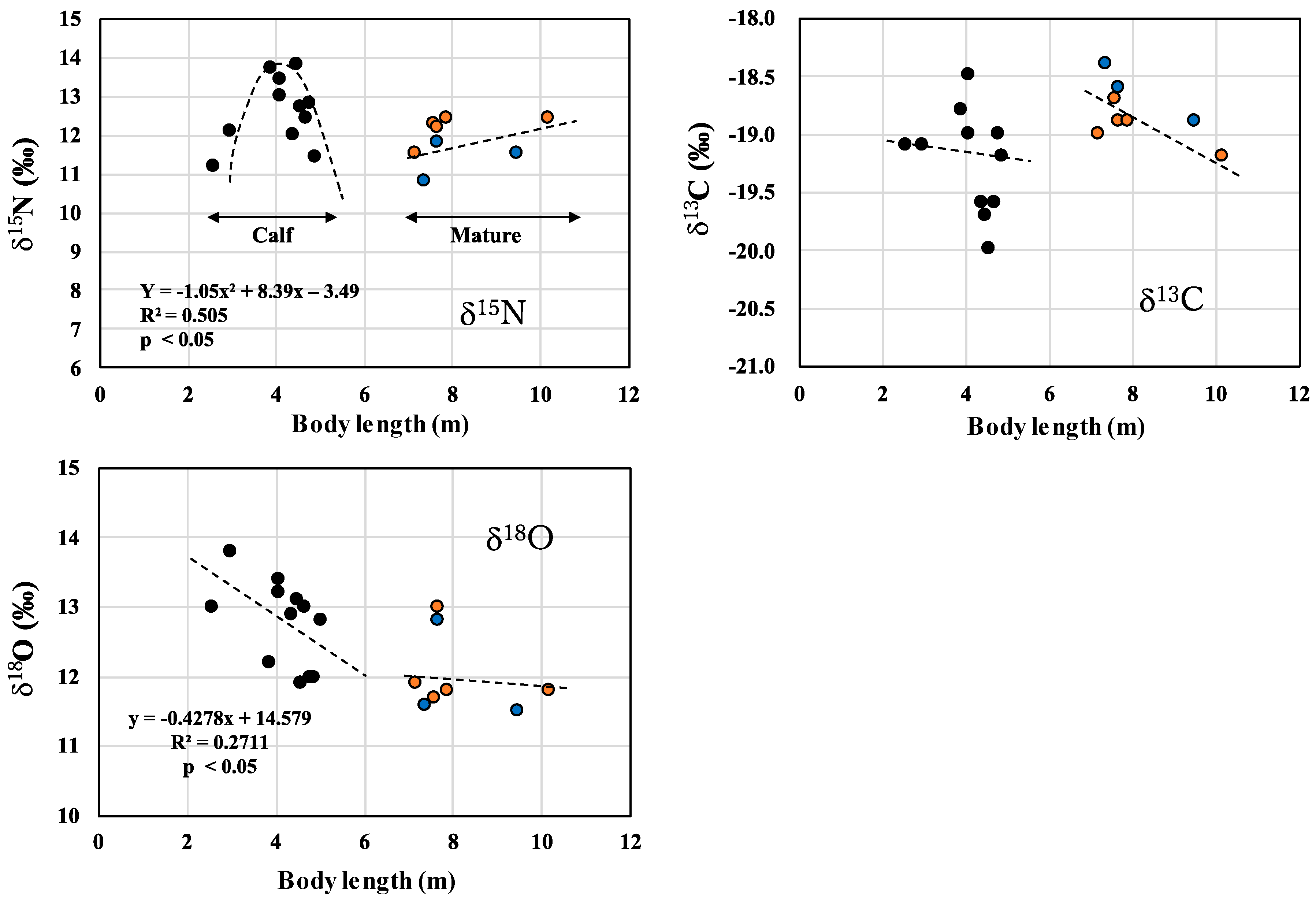
3.3. Changes in δ18O Values during Lactation in Common Minke Whales
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Endo, T.; Kobayashi, M. Typical changes in carbon and nitrogen stable isotope ratios and mercury concentration during the lactation of marine mammals. In Marine Mammals; Kaoud, H.A.E., Ed.; IntechOpen: London, UK, 2022; ISBN 978-1-80355-490-7. [Google Scholar]
- Fuller, B.; Fuller, J.; Harris, D.; Hedges, R. Detection of breastfeeding and weaning in modern human infants with carbon and nitrogen stable isotope ratios. Am. J. Phys. Anthr. 2006, 129, 279–293. [Google Scholar] [CrossRef] [PubMed]
- Newsome, S.D.; Koch, P.L.; Etnier, M.A.; Aurioles-Gamboa, D. Using Carbon and nitrogen isotope values to investigate maternal strategies in northeast pacific otariids. Mar. Mammal Sci. 2006, 22, 556–572. [Google Scholar] [CrossRef]
- Borrell, A.; Gómez-Campos, E.; Aguilar, A. Influence of Reproduction on Stable-Isotope Ratios: Nitrogen and Carbon Isotope Discrimination between Mothers, Fetuses, and Milk in the Fin Whale, a Capital Breeder. Physiol. Biochem. Zool. 2016, 89, 41–50. [Google Scholar] [CrossRef] [PubMed]
- Vales, D.; Cardona, L.; García, N.; Zenteno, L.; Crespo, E. Ontogenetic dietary changes in male South American fur seals Arctocephalus australis in Patagonia. Mar. Ecol. Prog. Ser. 2015, 525, 245–260. [Google Scholar] [CrossRef]
- Endo, T.; Kimura, O.; Terasaki, M.; Kobayashi, M. Body length, stable carbon, and nitrogen isotope ratios and mercury levels in common minke whales (Balaenoptera acutorostrata) stranded along the coast of Hokkaido, Japan. Aquat. Mamm. 2021, 47, 86–95. [Google Scholar] [CrossRef]
- Endo, T.; Terasaki, M.; Kimura, O. Stable isotope ratios of carbon, nitrogen, oxygen, and mercury concentrations in north pacific baleen whales and the comparison of their calves with toothed whale calves. J. Veter.-Sci. Med. 2022, 10, 1–10. [Google Scholar] [CrossRef]
- Oftedal, O.T. Lactation in Whales and Dolphins: Evidence of Divergence Between Baleen- and Toothed-Species. J. Mammary Gland. Biol. Neoplasia 1997, 2, 205–230. [Google Scholar] [CrossRef] [PubMed]
- Tsutaya, T.; Yoneda, M. Reconstruction of breastfeeding and weaning practices using stable isotope and trace element analyses: A review. Am. J. Phys. Anthr. 2015, 156, 2–21. [Google Scholar] [CrossRef]
- Oftedal, O.T. Milk of marine mammals. In Encyclopedia of Dairy Sciences, 2nd ed.; Fuquay, J.W., Ed.; Academic Press: Cambridge, MA, USA, 2011; pp. 563–580. [Google Scholar] [CrossRef]
- Kalish, J.M. 13C and 18O isotopic disequilibria in fish otoliths: Metabolic and kinetic effects. Mar. Ecol. Prog. Ser. 1991, 75, 191–203. [Google Scholar] [CrossRef]
- Clementz, M.T.; Koch, P.L. Differentiating aquatic mammal habitat and foraging ecology with stable isotopes in tooth enamel. Oecologia 2001, 129, 461–472. [Google Scholar] [CrossRef]
- Liu, A.G.S.C.; Seiffert, E.R.; Simons, E.L. Stable isotope evidence for an amphibious phase in early proboscidean evolution. Proc. Natl. Acad. Sci. USA 2008, 105, 5786–5791. [Google Scholar] [CrossRef]
- Newsome, S.D.; Clementz, M.T.; Koch, P.L. Using stable isotope biogeochemistry to study marine mammal ecology. Mar. Mammal Sci. 2010, 26, 509–572. [Google Scholar] [CrossRef]
- Lin, H.-Y.; Shiao, J.-C.; Chen, Y.-G.; Iizuka, Y. Ontogenetic vertical migration of grenadiers revealed by otolith microstructures and stable isotopic composition. Deep. Sea Res. Part I Oceanogr. Res. Pap. 2012, 61, 123–130. [Google Scholar] [CrossRef]
- Matthews, C.J.D.; Longstaffe, F.J.; Ferguson, S.H. Dentine oxygen isotopes (δ18O) as a proxy for odontocete distributions and movements. Ecol. Evol. 2016, 6, 4643–4653. [Google Scholar] [CrossRef]
- Matthews, C.J.D.; Longstaffe, F.J.; Lawson, J.W.; Ferguson, S.H. Distributions of Arctic and Northwest Atlantic killer whales inferred from oxygen isotopes. Sci. Rep. 2021, 11, 6739. [Google Scholar] [CrossRef]
- Camin, F.; Bontempo, L.; Perini, M.; Piasentier, E. Stable Isotope Ratio Analysis for Assessing the Authenticity of Food of Animal Origin. Compr. Rev. Food Sci. Food Saf. 2016, 15, 868–877. [Google Scholar] [CrossRef]
- Yoshida, N.; Miyazaki, N. Oxygen isotope correlation of cetacean bone phosphate with environmental water. J. Geophys. Res. Ocean. 1991, 96, 815–820. [Google Scholar] [CrossRef]
- Wright, L.E.; Schwarcz, H.P. Stable carbon and oxygen isotopes in human tooth enamel: Identifying breastfeeding and weaning in prehistory. Am. J. Physiol. Anthropol. 1998, 106, 1–18. [Google Scholar] [CrossRef]
- Andvik, C.; Jourdain, E.; Ruus, A.; Lyche, J.L.; Karoliussen, R.; Borgå, K. Preying on seals pushes killer whales from Norway above pollution effects thresholds. Sci. Rep. 2020, 10, 11888. [Google Scholar] [CrossRef]
- Power, M.; Klein, G.M.; Guiguer, K.R.R.A.; Kwan, M.K.H. Mercury accumulation in the fish community of a sub-Arctic lake in relation to trophic position and carbon sources. J. Appl. Ecol. 2002, 39, 819–830. [Google Scholar] [CrossRef]
- Chouvelon, T.; Spitz, J.; Caurant, F.; Mèndez-Fernandez, P.; Autier, J.; Lassus-Débat, A.; Chappuis, A.; Bustamante, P. Enhanced bioaccumulation of mercury in deep-sea fauna from the Bay of Biscay (north-east Atlantic) in relation to trophic positions identified by analysis of carbon and nitrogen stable isotopes. Deep. Sea Res. Part I Oceanogr. Res. Pap. 2012, 65, 113–124. [Google Scholar] [CrossRef]
- Chouvelon, T.; Caurant, F.; Cherel, Y.; Simon-Bouhet, B.; Spitz, J.; Bustamante, P. Species- and size-related patterns in stable isotopes and mercury concentrations in fish help refine marine ecosystem indicators and provide evidence for distinct management units for hake in the Northeast Atlantic. ICES J. Mar. Sci. 2014, 71, 1073–1087. [Google Scholar] [CrossRef]
- Pethybridge, H.; Butler, E.; Cossa, D.; Daley, R.; Boudou, A. Trophic structure and biomagnification of mercury in an assemblage of deepwater chondrichthyans from southeastern Australia. Mar. Ecol. Prog. Ser. 2012, 451, 163–174. [Google Scholar] [CrossRef]
- Honda, K. Contamination of heavy metals in marine mammals. In Biology of Marine Mammals; Miyazaki, N., Kasuya, T., Eds.; Scientist Inc.: Tokyo, Japan, 1990; pp. 242–253. [Google Scholar]
- Endo, T.; Kimura, O.; Hisamichi, Y.; Minoshima, Y.; Haraguchi, K. Age-dependent accumulation of heavy metals in a pod of killer whales (Orcinus orca) stranded in the northern area of Japan. Chemosphere 2007, 67, 51–59. [Google Scholar] [CrossRef] [PubMed]
- Endo, T.; Hisamichi, Y.; Kimura, O.; Haraguchi, K.; Baker, C.S. Contamination levels of mercury and cadmium in melon-headed whales (Peponocephala electra) from a mass stranding on the Japanese coast. Sci. Total. Environ. 2008, 401, 73–80. [Google Scholar] [CrossRef] [PubMed]
- Sakamoto, M.; Itai, T.; Murata, K. Effects of Prenatal Methylmercury Exposure: From Minamata Disease to Environmental Health Studies. Nippon. Eiseigaku Zasshi Jpn. J. Hyg. 2017, 72, 140–148. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, M.L.K.; Ellis, S.; Towers, J.R.; Doniol-Valcroze, T.; Franks, D.W.; Cant, M.A.; Weiss, M.N.; Johnstone, R.A.; Balcomb, K.C., 3rd; Ellifrit, D.K.; et al. A long postreproductive life span is a shared trait among genetically distinct killer whale populations. Ecol. Evol. 2021, 11, 9123–9136. [Google Scholar] [CrossRef] [PubMed]
- Ciner, B.; Wang, Y.; Parker, W. Oxygen isotopic variations in modern cetacean teeth and bones: Implications for ecological, paleoecological, and paleoclimatic studies. Sci. Bull. 2016, 61, 92–104. [Google Scholar] [CrossRef]
- Morin, P.A.; Archer, F.I.; Foote, A.D.; Vilstrup, J.; Allen, E.E.; Wade, P.; Durban, J.; Parsons, K.; Pitman, R.; Li, L.; et al. Complete mitochondrial genome phylogeographic analysis of killer whales (Orcinus orca) indicates multiple species. Genome Res. 2010, 20, 908–916. [Google Scholar] [CrossRef]
- Mitani, Y.; Kita, Y.F.; Saino, S.; Yoshioka, M.; Ohizumi, H.; Nakahara, F. Mitochondrial DNA haplotypes of killer whales around Hokkaido, Japan. Mammal Study 2021, 46, 205–211. [Google Scholar] [CrossRef]
- Endo, T.; Kimura, O.; Sato, R.; Kobayashi, M.; Matsuda, A.; Matsuishi, T.; Haraguchi, K. Stable isotope ratios of carbon, nitrogen and oxygen in killer whales (Orcinus orca) stranded on the coast of Hokkaido, Japan. Mar. Pollut. Bull. 2014, 86, 238–243. [Google Scholar] [CrossRef] [PubMed]
- Kastelein, R.A.; Kershaw, J.; Berghout, E.; Wiepkema, P.R. Food consumption and suckling in Killer whales. Int. Zoo Yearb. 2003, 38, 204–218. [Google Scholar] [CrossRef]
- Kasuya, T. Biology and Conservation of Small Cetaceans around Japan; University of Tokyo Press: Tokyo, Japan, 2019; ISBN 978-4-13-060238-9. [Google Scholar]
- Heyning, J.E. Presence of solid food in a young calf killer whale (Orcinus orca). Mar. Mamm. Sci. 1988, 4, 68–71. [Google Scholar] [CrossRef]
- Clark, S.T.; Odell, D.K.; Lacinak, C.T. Aspects of growth in captive killer whales (Orcinus orca). Mar. Mammal Sci. 2000, 16, 110–123. [Google Scholar] [CrossRef]
- Newsome, S.; Etnier, M.; Monson, D.; Fogel, M. Retrospective characterization of ontogenetic shifts in killer whale diets via δ13C and δ15N analysis of teeth. Mar. Ecol. Prog. Ser. 2009, 374, 229–242. [Google Scholar] [CrossRef]
- Logan, J.M.; Lutcavage, M.E. A comparison of carbon and nitrogen stable isotope ratios of fish tissues following lipid extractions with non-polar and traditional chloroform/methanol solvent systems. Rapid Commun. Mass Spectrom. 2008, 22, 1081–1086. [Google Scholar] [CrossRef] [PubMed]
- Ogasawara, H.; Hayasaka, M.; Maemoto, A.; Furukawa, S.; Ito, T.; Kimura, O.; Endo, T. Stable isotope ratios of carbon, nitrogen and selenium concentration in the scalp hair of Crohn’s disease patients who ingested the elemental diet Elental®. Rapid Commun. Mass Spectrom. 2018, 33, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Baird, R.W. Killer Whales of the World: Natural History and Conservation; Voyager Press: McGregor, MN, USA, 2002. [Google Scholar]
- Dietz, R.; Letcher, R.J.; Desforges, J.-P.; Eulaers, I.; Sonne, C.; Wilson, S.; Andersen-Ranberg, E.; Basu, N.; Barst, B.D.; Bustnes, J.O.; et al. Current state of knowledge on biological effects from contaminants on arctic wildlife and fish. Sci. Total. Environ. 2019, 696, 133792. [Google Scholar] [CrossRef]
- de Armas, Y.C.; Mavridou, A.-M.; Domínguez, J.G.; Hanson, K.; Laffoon, J. Tracking breastfeeding and weaning practices in ancient populations by combining carbon, nitrogen and oxygen stable isotopes from multiple non-adult tissues. PLoS ONE 2022, 17, e0262435. [Google Scholar] [CrossRef]
- O’Sullivan, R.; Schmidt, O.; Monahan, F.J. Stable isotope ratio analysis for the authentication of milk and dairy ingredients: A review. Int. Dairy J. 2021, 126, 105268. [Google Scholar] [CrossRef]
- Yamamoto, M.; Tanaka, N.; Tsunogai, S. Okhotsk Sea intermediate water formation deduced from oxygen isotope systematics. J. Geophys. Res. Oceans 2001, 106, 31075–31084. [Google Scholar] [CrossRef]
- Oda, M.; Tetsu, T.; Sakai, S.; Ishimura, T. Discrimination of the migration for Japanese sardine in Pacific stock by using mi-croscale stable isotopic analytical technique. Bull. Jpn. Scoc. Fish Oceanogr. 2016, 80, 48–55. [Google Scholar]
- Boecklen, W.J.; Yarnes, C.T.; Cook, B.A.; James, A.C. On the Use of Stable Isotopes in Trophic Ecology. Annu. Rev. Ecol. Evol. Syst. 2011, 42, 411–440. [Google Scholar] [CrossRef]
- O’Brien, D.M. Stable Isotope Ratios as Biomarkers of Diet for Health Research. Annu. Rev. Nutr. 2015, 35, 565–594. [Google Scholar] [CrossRef] [PubMed]
- Trueman, C.N.; McGill, R.A.R.; Guyard, P.H. The effect of growth rate on tissue-diet isotopic spacing in rapidly growing animals. An experimental study with Atlantic salmon (Salmo salar). Rapid Commun. Mass Spectrom. 2005, 19, 3239–3247. [Google Scholar] [CrossRef] [PubMed]
- Zanden, M.J.; Clayton, M.K.; Moody, E.K.; Solomon, C.T.; Weidel, B.C. Stable Isotope Turnover and Half-Life in Animal Tissues: A Literature Synthesis. PLoS ONE 2015, 10, e0116182. [Google Scholar] [CrossRef] [PubMed]
- Caut, S.; Angulo, E.; Courchamp, F. Variation in discrimination factors (Δ15N and Δ13C): The effect of diet isotopic values and applications for diet reconstruction. J. Appl. Ecol. 2009, 46, 443–453. [Google Scholar] [CrossRef]
- Drago, M.; Franco-Trecu, V.; Cardona, L.; Inchausti, P. Diet-to-female and female-to-pup isotopic discrimination in South American sea lions. Rapid Commun. Mass Spectrom. 2015, 29, 1513–1520. [Google Scholar] [CrossRef] [PubMed]
- Borrell, A.; Abad-Oliva, N.; Gómez-Campos, E.; Giménez, J.; Aguilar, A. Discrimination of stable isotopes in fin whale tissues and application to diet assessment in cetaceans. Rapid Commun. Mass Spectrom. 2012, 26, 1596–1602. [Google Scholar] [CrossRef]
- Honda, K.; Tatsukawa, R.; Itano, K.; Miyazaki, N.; Fujiyama, T. Heavy metal concentrations in muscle, liver and kidney tissue of striped dolphin, Stenella coeruleoalba, and their variations with body length, weight, age and sex. Agric. Biol. Chem. 1983, 47, 1219–1228. [Google Scholar] [CrossRef]
- Jourdain, E.; Andvik, C.; Karoliussen, R.; Ruus, A.; Vongraven, D.; Borgå, K. Isotopic niche differs between seal and fish-eating killer whales (Orcinus orca) in northern Norway. Ecol. Evol. 2020, 10, 4115–4127. [Google Scholar] [CrossRef] [PubMed]
- Itano, K.; Kawai, S.; Miyazaki, N.; Tatsukawa, R.; Fujiyama, T. Mercury and selenium levels at the fetal and suckling stages of striped dolphin, Stenella coeruleoalba. Agric. Biol. Chem. 1984, 48, 1691–1698. [Google Scholar] [CrossRef][Green Version]
- Endo, T.; Kimura, O.; Terasaki, M.; Nakagun, S.; Kato, Y.; Fujii, Y.; Haraguchi, K.; Baker, C.S. Carbon, nitrogen, and oxygen stable isotope ratios of striped dolphins and short-finned pilot whales stranded in Hokkaido, northern Japan, compared with those of other cetaceans stranded and hunted in Japan. Isot. Environ. Health Stud. 2023, 59, 230–247. [Google Scholar] [CrossRef] [PubMed]
- Caon, G.; Secchi, E.R.; Capp, E.; Kucharski, L.C. Milk composition of franciscana dolphin (Pontoporia blainvillei) from Rio Grande do Sul, southern Brazil. J. Mar. Biol. Assoc. United Kingd. 2008, 88, 1099–1101. [Google Scholar] [CrossRef]

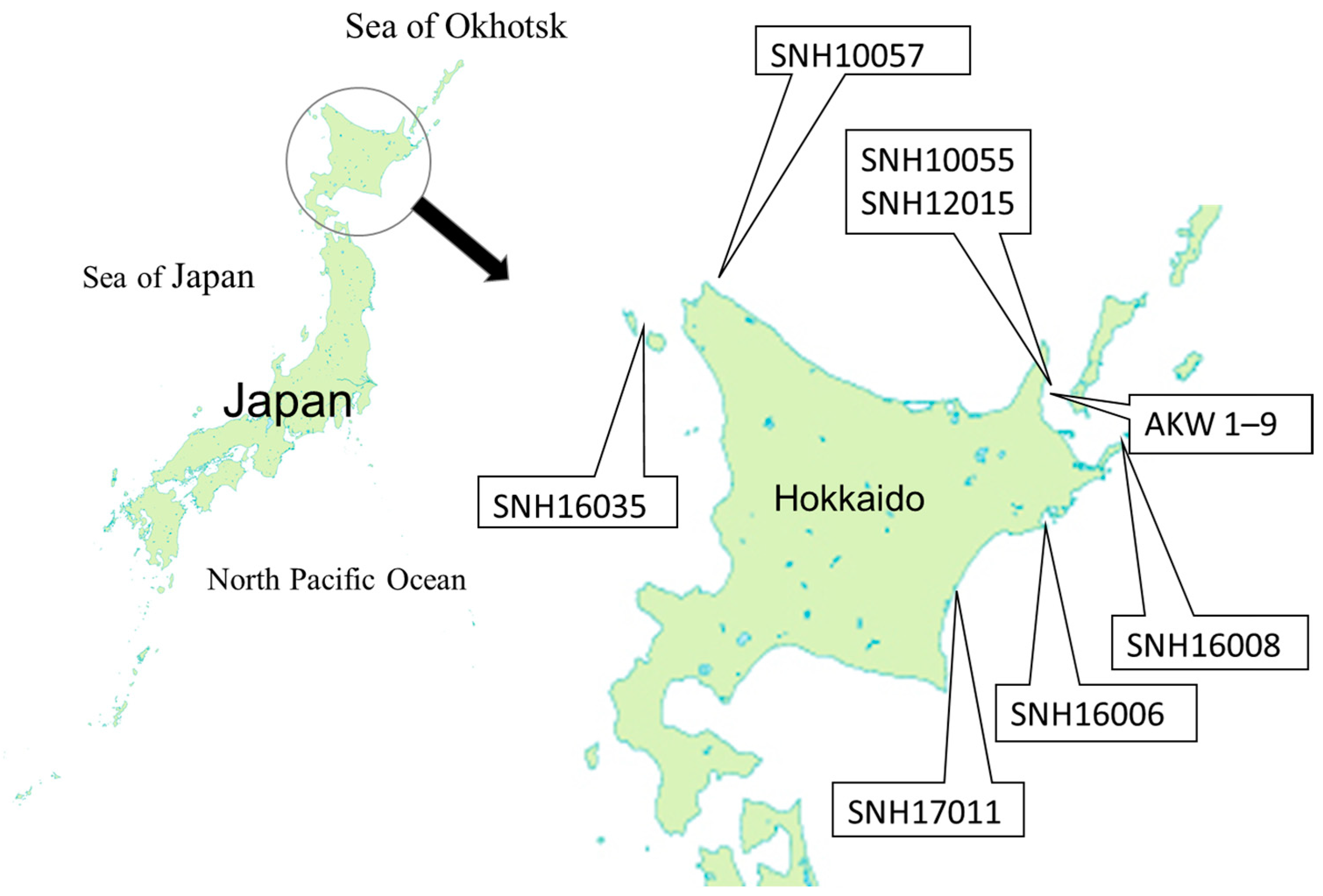
 ), lactating female (
), lactating female ( ), non-lactating females (
), non-lactating females ( ) and mature males (
) and mature males ( ) of killer whales.
), lactating female (), non-lactating females () and mature males () of killer whales.
) of killer whales.
), lactating female (), non-lactating females () and mature males () of killer whales. ), lactating female (), non-lactating females (), and mature males ().
), lactating female (), non-lactating females (), and mature males ().
), lactating female (), non-lactating females (), and mature males ().
), lactating female (), non-lactating females (), and mature males (). ), mature males (), and mature females (). The upper figures showing δ15N and δ13C values were reprinted from Endo et al. [6] with permission from Aquatic Mammals.
), mature males (), and mature females (). The upper figures showing δ15N and δ13C values were reprinted from Endo et al. [6] with permission from Aquatic Mammals.
), mature males (), and mature females (). The upper figures showing δ15N and δ13C values were reprinted from Endo et al. [6] with permission from Aquatic Mammals.
), mature males (), and mature females (). The upper figures showing δ15N and δ13C values were reprinted from Endo et al. [6] with permission from Aquatic Mammals.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Muscle | Liver | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Body Length (m) | Age (y) | δ13C (‰) | δ15N (‰) | δ18O (‰) | Hg (μg/ wet g) | δ13C (‰) | δ15N (‰) | δ18O (‰) | Hg (μg/ wet g) | ||
| SNH16006 * | Calf male | 2.2 | <1 | −18.2 | 17.8 | 16.3 | 0.22 | ND | ND | ND | ND |
| SNH16035 * | Calf female | 2.3 | <1 | −18.3 | 16.9 | 15.9 | 0.21 | −18.3 | 17.2 | 16.0 | 0.48 |
| AKW3 ** | Calf male a | 2.7 | <1 | −16.7 | 18.1 | 12.4 | 0.10 | −16.6 | 18.6 | 12.6 | 0.30 |
| AKW8 ** | Calf female b | 2.7 | <1 | −16.9 | 18.1 | 12.3 | 0.08 | −16.7 | 18.7 | 13.4 | 0.50 |
| AKW7 ** | Calf female c | 3.0 | <1 | −16.8 | 18.2 | 11.9 | 0.07 | −16.6 | 18.7 | 13.3 | 0.30 |
| SNH12015 ** | Weaning calf d | 3.8 | 3 | −19.3 | 15.1 | 15.5 | 0.52 | −19.3 | 15.4 | 17.6 | 2.36 |
| AKW2 ** | Mature female e | 5.6 | 13 | −17.2 | 16.0 | 13.1 | 1.27 | −16.8 | 18.2 | 13.0 | 57.4 |
| SNH16008 * | Mature female | 5.7 | ND | −17.5 | 15.8 | 14.2 | 1.68 | ND | ND | ND | ND |
| SNH10055 ** | Mature female | 5.8 | ND | −18.2 | 16.5 | 15.0 | 2.45 | −18.0 | 18.0 | 14.4 | 107.6 |
| AKW6 ** | Mature female f | 6.0 | 17 | −17.1 | 16.4 | 12.7 | 1.06 | −16.8 | 19.2 | 13.7 | 38.0 |
| AKW9 ** | Mature female g | 6.5 | 29 | −17.2 | 16.5 | 12.0 | 1.26 | −16.7 | 19.2 | 13.4 | 62.4 |
| AKW4 ** | Mature female | 6.6 | 24 | −17.1 | 16.8 | 12.7 | 1.25 | −15.9 | 19.1 | 11.7 | 55.4 |
| AKW5 ** | Mature female | 6.9 | 59 | −17.0 | 16.4 | 12.5 | 1.30 | −16.3 | 18.6 | 12.0 | 97.8 |
| SNH10057 ** | Mature male | 7.0 | ND | −18.0 | 15.9 | 15.0 | 3.24 | −17.1 | 17.3 | 17.2 | 53.3 |
| SNH17011 * | Mature male | 7.1 | ND | −17.7 | 16.3 | 13.5 | 2.03 | −17.0 | 18.5 | 13.7 | 41.0 |
| AKW1 ** | Mature male | 7.7 | 34 | −16.9 | 17.0 | 13.1 | 1.46 | −16.5 | 18.8 | 12.7 | 35.1 |
| Samples | BL | δ18O | δ13C * | δ15N * | Hg * |
|---|---|---|---|---|---|
| (m) | (‰) | (‰) | (‰) | (μg/wet g) | |
| 1 | 2.6 (C) | 13.0 | −9.1 | 11.2 | 0.03 |
| 2 | 3.0 (C) | 13.8 | −19.1 | 12.1 | 0.01 |
| 3 | 3.9 (C) | 12.2 | −18.8 | 13.7 | 0.04 |
| 4 | 4.1 (C) | 13.4 | −18.5 | 13.0 | 0.01 |
| 5 | 4.1 (C) | 13.2 | −19.0 | 13.4 | 0.02 |
| 6 | 4.4 (C) | 12.9 | −19.6 | 12.0 | 0.06 |
| 7 | 4.5 (C) | 13.1 | −19.7 | 13.8 | 0.02 |
| 8 | 4.6 (C) | 11.9 | −20.0 | 12.7 | 0.01 |
| 9 | 4.7 (C) | 13.0 | −19.6 | 12.4 | 0.07 |
| 10 | 4.8 (C) | 12.0 | −19.0 | 12.8 | 0.02 |
| 11 | 4.9 (C) | 12.0 | −19.2 | 11.4 | 0.06 |
| 12 | 5.0 (C) | 12.8 | −18.3 | 12.9 | 0.03 |
| 13 | 7.2 (M) | 11.9 | −19.0 | 11.5 | 0.12 |
| 14 | 7.4 (M) | 12.8 | −18.4 | 10.8 | 0.11 |
| 15 | 7.6 (F) | 12.5 | −18.7 | 12.3 | 0.14 |
| 16 | 7.7 (M) | 12.8 | −18.6 | 11.8 | 0.13 |
| 17 | 7.7 (F) | 13.0 | −18.9 | 12.2 | 0.15 |
| 18 | 7.9 (F) | 11.8 | −18.9 | 12.4 | 0.07 |
| 19 | 9.5 (M) | 12.0 | −18.9 | 11.5 | 0.17 |
| 20 | 10.2 (F) | 11.8 | −19.2 | 12.4 | 0.18 |
| δ13C (‰) | δ15N (‰) | δ18O (‰) | ||
|---|---|---|---|---|
| Calves | Muscle (n = 6) | −17.7 ± 1.1 | 17.4 ± 1.2 | 14.1 ± 2.1 |
| Liver (n = 5) | −17.5 ± 1.2 | 17.7 ± 1.4 | 14.6 ± 2.1 | |
| Mature animals | Muscle (n =10) | −17.4 ± 0.4 | 16.4 ± 0.4 | 13.4 ± 1.0 |
| Liver (n = 9) | −16.8 ± 0.6 | 18.5 ± 0.6 * | 13.5 ± 1.6 | |
| Species | Hg concentration in Liver (μg/wet g) | δ15N in Muscle (‰) |
|---|---|---|
| Killer whale (mature animals) of this study | 60.9 ± 25.6 (n = 9) a | 16.4 ± 0.4 (n = 10) a |
| all animals of this study | 39.4 ± 36.0 (n = 16) a | 16.7 ± 0.9 (n = 16) a |
| Dall’s porpoises (Phocoenoides dalli) | 8.62 ± 10.60 (n = 52) b | 13.1 ± 0.9 (n = 56) b |
| Harbor porpoises (Phocoena phocoena) | 6.80 ± 14.9 (n = 45) b | 13.2 ± 1.0 (n = 49) b |
| Spotted seals (Phoca largha) | 1.93 and 0.34 b | 14.5 and 15.1 b |
| Harbor seals (Phoca vitulina) | 2.96 ± 3.26 (n = 32) b | 15.6 ± 0.52 (n = 32) b |
| Common minke whales (Balaenoptera acutorostrata) | 0.103 ± 0.100 (n = 8) b | 12.3 ± 0.8 (n = 20) c |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Endo, T.; Kimura, O.; Terasaki, M.; Kato, Y.; Fujii, Y.; Haraguchi, K. Changes in Carbon, Nitrogen, and Oxygen Stable Isotope Ratios and Mercury Concentrations in Killer Whales (Orcinus orca) during and after Lactation. J. Mar. Sci. Eng. 2024, 12, 623. https://doi.org/10.3390/jmse12040623
Endo T, Kimura O, Terasaki M, Kato Y, Fujii Y, Haraguchi K. Changes in Carbon, Nitrogen, and Oxygen Stable Isotope Ratios and Mercury Concentrations in Killer Whales (Orcinus orca) during and after Lactation. Journal of Marine Science and Engineering. 2024; 12(4):623. https://doi.org/10.3390/jmse12040623
Chicago/Turabian StyleEndo, Tetsuya, Osamu Kimura, Masaru Terasaki, Yoshihisa Kato, Yukiko Fujii, and Koichi Haraguchi. 2024. "Changes in Carbon, Nitrogen, and Oxygen Stable Isotope Ratios and Mercury Concentrations in Killer Whales (Orcinus orca) during and after Lactation" Journal of Marine Science and Engineering 12, no. 4: 623. https://doi.org/10.3390/jmse12040623
APA StyleEndo, T., Kimura, O., Terasaki, M., Kato, Y., Fujii, Y., & Haraguchi, K. (2024). Changes in Carbon, Nitrogen, and Oxygen Stable Isotope Ratios and Mercury Concentrations in Killer Whales (Orcinus orca) during and after Lactation. Journal of Marine Science and Engineering, 12(4), 623. https://doi.org/10.3390/jmse12040623