
First Survey of the Sponge Community of a Semi-Submerged Marine Cave along the Adriatic Apulian Coast



Abstract
:1. Introduction
2. Materials and Methods
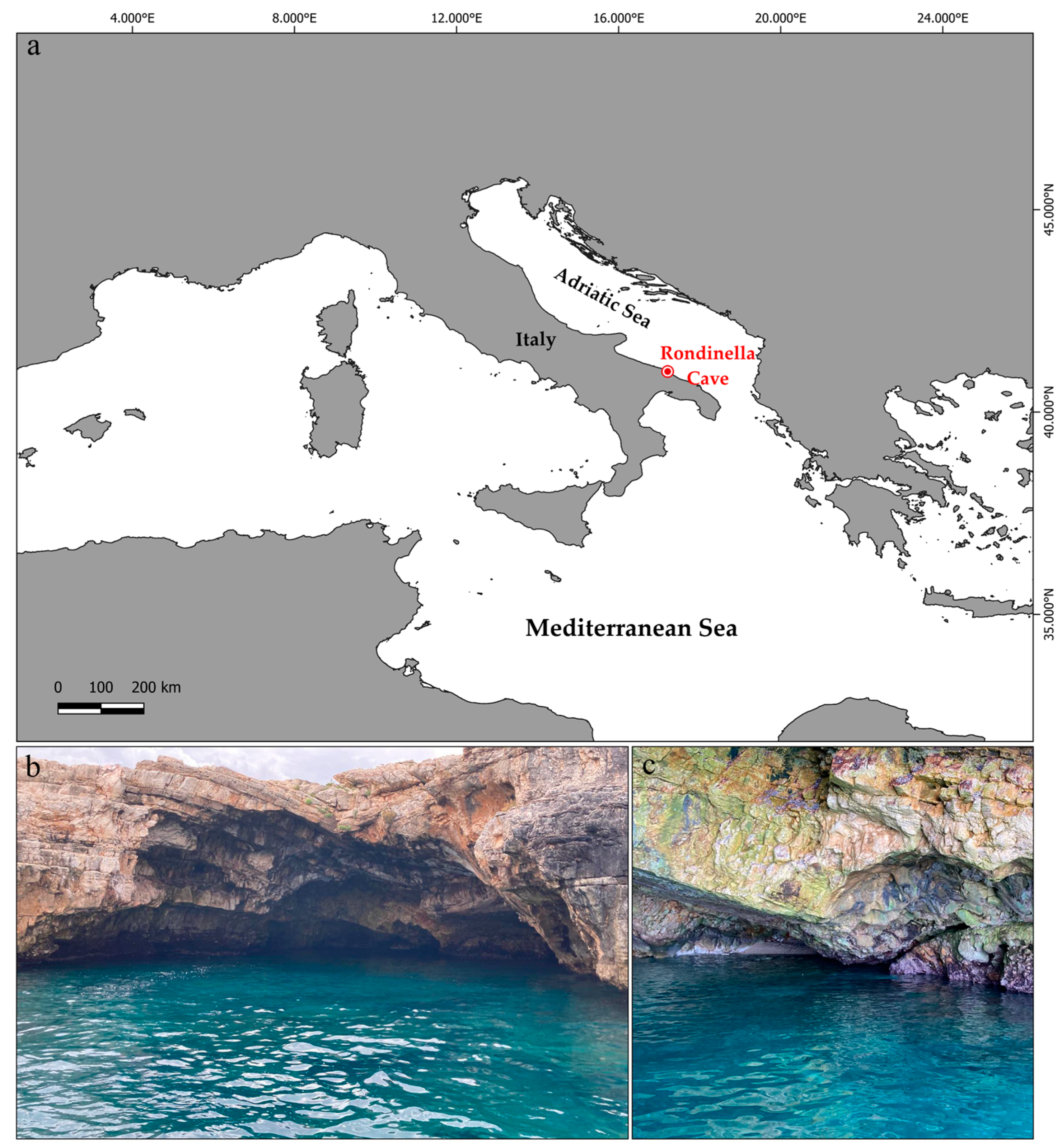
2.1. Study Area
2.2. Sampling
2.2.1. Sponge Cover
2.2.2. Taxonomic Analysis
2.2.3. Statistical Analysis
3. Results
3.1. Sponge Cover
3.2. Taxonomic Composition
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gerovasileiou, V.; Voultsiadou, E. Marine Caves of the Mediterranean Sea: A Sponge Biodiversity Reservoir within a Biodiversity Hotspot. PLoS ONE 2012, 7, e39873. [Google Scholar] [CrossRef] [PubMed]
- Sarà, M. Il Popolamento Delle Grotte Marine e Sua Protezione. In Proceedings of the Atti del IV Simposio Nazionale sulla Conservazione della Natura, Bari, Italy, 23–28 April 1974; Volume 1, pp. 51–59. [Google Scholar]
- Giakoumi, S.; Sini, M.; Gerovasileiou, V.; Mazor, T.; Beher, J.; Possingham, H.P.; Abdulla, A.; Çinar, M.E.; Dendrinos, P.; Gucu, A.C.; et al. Ecoregion-Based Conservation Planning in the Mediterranean: Dealing with Large-Scale Heterogeneity. PLoS ONE 2013, 8, e76449. [Google Scholar] [CrossRef] [PubMed]
- Nepote, E.; Bianchi, C.N.; Morri, C.; Ferrari, M.; Montefalcone, M. Impact of a Harbour Construction on the Benthic Community of Two Shallow Marine Caves. Mar. Pollut. Bull. 2017, 114, 35–45. [Google Scholar] [CrossRef] [PubMed]
- Gerovasileiou, V.; Dimitriadis, C.; Arvanitidis, C.; Voultsiadou, E. Taxonomic and Functional Surrogates of Sessile Benthic Diversity in Mediterranean Marine Caves. PLoS ONE 2017, 12, e0183707. [Google Scholar] [CrossRef] [PubMed]
- Harmelin, J.G. Les Grottes Sous-Marines Obscures: Un Milieu Extrême et Un Remarquable Biotope Refuge. Téthys 1985, 11, 214–229. [Google Scholar]
- Bianchi, C.N.; Morri, C. Southern Species in the Ligurian Sea (Northern Mediterranean): New Records and a Review. Boll. Mus. Ist. Biol. Univ. Genova 1994, 58, 181–197. [Google Scholar]
- Sarà, M.; Siribelli, L. La Fauna Di Poriferi Delle ‘Secche’Del Golfo Di Napoli. II. La Secca Di Benda Palummo. Annu. Ist. Mus. Zool. Univ. Napoli 1962, 14, 1–62. [Google Scholar]
- Ereskovsky, A.V.; Kovtun, O.A.; Pronin, K.K. Marine Cave Biota of the Tarkhankut Peninsula (Black Sea, Crimea), with Emphasis on Sponge Taxonomic Composition, Spatial Distribution and Ecological Particularities. J. Mar. Biol. Assoc. U. K. 2016, 96, 391–406. [Google Scholar] [CrossRef]
- Corriero, G.; Scalera Liaci, L.; Ruggiero, D.; Pansini, M. The Sponge Community of a Semi-Submerged Mediterranean Cave. Mar. Ecol. 2000, 21, 85–96. [Google Scholar] [CrossRef]
- Bell, J.J. The Sponge Community in a Semi-Submerged Temperate Sea Cave: Density, Diversity and Richness. Mar. Ecol. 2002, 23, 297–311. [Google Scholar] [CrossRef]
- Sarà, M. Stratification Des Peuplements d’éponges à Recouvrement Total Dans Certaines Grottes Du Niveau Superficiel. Rapp. Comm. Int. Mer. Médit. 1968, 19, 83–85. [Google Scholar]
- Bell, J.J.; Carballo, J.L. Patterns of Sponge Biodiversity and Abundance across Different Biogeographic Regions. Mar. Biol. 2008, 155, 563–570. [Google Scholar] [CrossRef]
- Sarà, M.; Vacelet, J. Ecologie de Demosponges. In Traite de Zoologie. Anatomie Systematiqui, Biologie; Grassé, P.P., Ed.; Masson et Cie: Paris, France, 1973. [Google Scholar]
- Gerovasileiou, V.; Chintiroglou, C.; Vafidis, D.; Koutsoubas, D.; Sini, M.; Dailianis, T.; Issaris, Y.; Akritopoulou, E.; Dimarchopoulou, D.; Voutsiadou, E. Census of Biodiversity in Marine Caves of the Eastern Mediterranean Sea. Med. Mar. Sci. 2015, 16, 245–265. [Google Scholar] [CrossRef]
- Barnes, D.K.A.; Bell, J.J. Coastal Sponge Communities of the West Indian Ocean: Morphological Richness and Diversity. Afr. J. Ecol. 2002, 40, 350–359. [Google Scholar] [CrossRef]
- Martí, R.; Uriz, M.J.; Ballesteros, E.; Turon, X. Benthic Assemblages in Two Mediterranean Caves: Species Diversity and Coverage as a Function of Abiotic Parameters and Geographic Distance. J. Mar. Biol. Assoc. U. K. 2004, 84, 557–572. [Google Scholar] [CrossRef]
- Bussotti, S.; Terlizzi, A.; Fraschetti, S.; Belmonte, G.; Boero, F. Spatial and Temporal Variability of Sessile Benthos in Shallow Mediterranean Marine Caves. Mar. Ecol. Prog. Ser. 2006, 325, 109–119. [Google Scholar] [CrossRef]
- Gerovasileiou, V.; Bianchi, C.N. Mediterranean Marine Caves: A Synthesis of Current Knowledge. In Oceanography and Marine Biology; CRC Press: Boca Raton, FL, USA, 2021; ISBN 978-1-00-313884-6. [Google Scholar]
- Longo, C.; Giménez, G.; Miscioscia, F.; Corriero, G. Sponge Fauna of the Apulian Marine Caves (Southern Italy): Current State of Knowledge. Diversity 2023, 15, 641. [Google Scholar] [CrossRef]
- Wulff, J. Assessing and Monitoring Coral Reef Sponges: Why and How? Bull. Mar. Sci. 2001, 69, 831–846. [Google Scholar]
- Grenier, M.; Ruiz, C.; Fourt, M.; Santonja, M.; Dubois, M.; Klautau, M.; Vacelet, J.; Boury-Esnault, N.; Perez, T. Sponge Inventory of the French Mediterranean Waters, with an Emphasis on Cave-Dwelling Species. Zootaxa 2018, 4466, 205–228. [Google Scholar] [CrossRef]
- Aspetti Naturalistici e Scientifici delle Grotte e delle Falesie di Polignano a Mare. Available online: https://ricerca.uniba.it/handle/11586/182931.1 (accessed on 18 March 2024).
- Forti, P. Processi Carsici e Speleogenesi. Prima Parte. Speleologia 1991, 24, 42–46. [Google Scholar]
- Forti, P. Processi Carsici e Speleogenesi. Seconda Parte. Speleologia 1992, 26, l. [Google Scholar]
- Favale, F.F.; Sauro, U. Le Grotte di Polignano: Studi in Memoria di Franco Orofino; Federazione Speleologica Pugliese: Taranto, Italy, 1994. [Google Scholar]
- Spaccavento, M.; Mastrototaro, F.; Tursi, A.; Montesanto, F.; Bottalico, A.; Longo, C.; Chimienti, G. A Non-Invasive Monitoring Method to Assess the Composition of Megabenthic Communities in Semi-Submerged Marine Caves. In Proceedings of the IEEE International Workshop on Metrology for the Sea, Learning to Measure Sea Health Parameters (MetroSea), Milazzo, Italy, 3–5 October 2022; pp. 257–261. [Google Scholar]
- Gimenez, G.; Corriero, G.; Beqiraj, S.; Lazaj, L.; Lazic, T.; Longo, C.; Mercurio, M.; Nonnis Marzano, C.; Zuccaro, M.; Zuna, V.; et al. Characterization of the Coralligenous Formations from the Marine Protected Area of Karaburun-Sazan, Albania. J. Mar. Sci. Eng. 2022, 10, 1458. [Google Scholar] [CrossRef]
- Trygonis, V.; Sini, M. photoQuad: A Dedicated Seabed Image Processing Software, and a Comparative Error Analysis of Four Photoquadrat Methods. J. Exp. Mar. Biol. Ecol. 2012, 424–425, 99–108. [Google Scholar] [CrossRef]
- Rützler, K. Sponges in Coral Reefs. In Coral Reefs: Research Methods: Monographs on Oceanographic Methodology; UNESCO: Paris, France, 1978; Volume 5. [Google Scholar]
- Hooper, J.N.A.; Van Soest, R.W.M. Systema Porifera. A Guide to the Classification of Sponges. In Systema Porifera: A Guide to the Classification of Sponges; Hooper, J.N.A., Van Soest, R.W.M., Willenz, P., Eds.; Springer US: Boston, MA, USA, 2002; pp. 1–7. ISBN 978-1-4615-0747-5. [Google Scholar]
- Pansini, M. Fauna d’Italia: Porifera; 1. Calcarea, Demospongiae (Partim), Hexactinellida, Homoscleromorpha; Calderini: Bologna, Italy, 2011. [Google Scholar]
- Morrow, C.; Cárdenas, P. Proposal for a Revised Classification of the Demospongiae (Porifera). Front. Zool. 2015, 12, 7. [Google Scholar] [CrossRef] [PubMed]
- de Voogd, N.; Alvarez, B.; Boury-Esnault, N.; Cárdenas, P.; Díaz, M.-C.; Dohrmann, M.; Downey, R.; Goodwin, C.; Hajdu, E.; Hooper, J.; et al. World Porifera Database. Available online: https://www.marinespecies.org/porifera (accessed on 18 March 2024).
- Gerovasileiou, V.; Martínez García, A.; Álvarez Noguera, F.; Boxshall, G.; Humphreys, W.F.; Jaume, D.; Becking, L.E.; Muricy, G.; van Hengstum, P.J.; Yamasaki, H.; et al. World Register of Marine Cave Species (WoRCS). Available online: Https://www.Marinespecies.Org/Worcs (accessed on 18 March 2024).
- Pansini, M.; Longo, C. Checklist Della Fauna Marina Italiana. Porifera. Biol. Mar. Med. 2008, 15, 44–70. [Google Scholar]
- Longo, C.; Cardone, F.; Pierri, C.; Mercurio, M.; Mucciolo, S.; Marzano, C.N.; Corriero, G. Sponges Associated with Coralligenous Formations along the Apulian Coasts. Mar. Biodivers. 2018, 48, 2151–2163. [Google Scholar] [CrossRef]
- Boury-Esnault, N.; Rützler, K. Thesaurus of Sponge Morphology. Smithson. Contrib. Zoöl. 1997, 596, 1–55. [Google Scholar] [CrossRef]
- Clarke, K.R.; Gorley, R.N. Primer; PRIMER-e: Plymouth, UK, 2006; Volume 866. [Google Scholar]
- Anderson, M. PERMANOVA+ for PRIMER: Guide to Software and Statistical Methods; Primer-E Ltd.: Plymouth, UK, 2008. [Google Scholar]
- Hammer, Ø.; Harper, D.A. Past: Paleontological Statistics Software Package for Education and Data Analysis. Palaeontol. Electron. 2001, 4, 1. [Google Scholar]
- Sarà, M. La Fauna Di Poriferi Delle Grotte Delle Isole Tremiti. Studio Ecologico e Sistematico. Arch. Zool. Ital. 1961, 46, 1–59. [Google Scholar]
- Pulitzer-Finali, G.; Pronzato, R. Report on a Collection of Sponges from the Bay of Naples; Pubblicazioni della Stazione Zoologica di Napoli: Marine Ecology: Napoli, Italy, 1970; Volume 38, pp. 328–354. [Google Scholar]
- Pulitzer-Finali, G. A Collection of Mediterranean Demospongiae (Porifera) with, in Appendix, a List of the Demospongiae Hitherto Recorded from the Mediterranean Sea. Ann. Mus. Civ. Stor. Nat. Giacomo Doria 1983, 84, 445–621. [Google Scholar]
- Corriero, G.; Scalera Liaci, L.; Pronzato, R. Two New Species of Dendroxea Griessinger (Porifera: Demospongiae) from the Mediterranean Sea. Bull. Inst. R. Sci. Nat. Belg. 1996, 66, 197–203. [Google Scholar]
- Pouliquen, L. Les Spongiaires Des Grottes Sous-Marines de La Région de Marseille: Écologie et Systématique. Téthys 1972, 3, 717–758. [Google Scholar]
- Costa, G.; Bavestrello, G.; Micaroni, V.; Pansini, M.; Strano, F.; Bertolino, M. Sponge Community Variation along the Apulian Coasts (Otranto Strait) over a Pluri-Decennial Time Span. Does Water Warming Drive a Sponge Diversity Increasing in the Mediterranean Sea? J. Mar. Biol. Assoc. U. K. 2019, 99, 1519–1534. [Google Scholar] [CrossRef]
- Calcinai, B.; Bavestrello, G.; Betti, F.; Bo, M.; Cerrano, C.; Di Camillo, C.G.; Tazioli, S. Storie Vitali Peculiari Del Benthos Di Substrato Duro Nei SIC Marini Marchigiani. Biol. Mar. Med. 2009, 16, 69–72. [Google Scholar]
- Mercurio, M.; Giménez, G.; Bavestrello, G.; Cardone, F.; Corriero, G.; Giampaoletti, J.; Gravina, M.F.; Pierri, C.; Longo, C.; Giangrande, A.; et al. A Dataset of Benthic Species from Mesophotic Bioconstructions on the Apulian Coast (Southeastern Italy, Mediterranean Sea). Data 2024, 9, 45. [Google Scholar] [CrossRef]
- Chevaldonné, P.; Lejeusne, C. Regional Warming-induced Species Shift in North-west Mediterranean Marine Caves. Ecol. Lett. 2003, 6, 371–379. [Google Scholar] [CrossRef]
- Parravicini, V.; Guidetti, P.; Morri, C.; Montefalcone, M.; Donato, M.; Bianchi, C.N. Consequences of Sea Water Temperature Anomalies on a Mediterranean Submarine Cave Ecosystem. Estuar. Coast. Shelf Sci. 2010, 86, 276–282. [Google Scholar] [CrossRef]
- Montefalcone, M.; De Falco, G.; Nepote, E.; Canessa, M.; Bertolino, M.; Bavestrello, G.; Morri, C.; Bianchi, C.N. Thirty Year Ecosystem Trajectories in a Submerged Marine Cave under Changing Pressure Regime. Mar. Environ. Res. 2018, 137, 98–110. [Google Scholar] [CrossRef] [PubMed]
- Pulido Mantas, T.; Roveta, C.; Calcinai, B.; Coppari, M.; Di Camillo, C.G.; Marchesi, V.; Marrocco, T.; Puce, S.; Cerrano, C. Photogrammetry as a Promising Tool to Unveil Marine Caves’ Benthic Assemblages. Sci. Rep. 2023, 13, 7587. [Google Scholar] [CrossRef]
- Costa, G.; Violi, B.; Bavestrello, G.; Pansini, M.; Bertolino, M. Aplysina aerophoba (Nardo, 1833) (Porifera, Demospongiae): An Unexpected Miniaturised Growth Form from the Tidal Zone of Mediterranean Caves: Morphology and DNA Barcoding. Eur. Zool. J. 2020, 87, 73–81. [Google Scholar] [CrossRef]
- Vacelet, J. Eponges de la Roche du Large et de l’Etage Bathyal de Méditerranée: Récoltes de la Soucoupe Plongeante Cousteau et Dragages; Éditions du Muséum: Paris, France, 1969; Volume 59. [Google Scholar]
- Cinelli, F.; Fresi, E.; Mazzella, L.; Pansini, M.; Pronzato, R.; Svoboda, A. Distribution of Benthic Phyto-and Zoocoenoses along a Light Gradient in a Superficial Marine Cave. In Biology of Benthic Organisms; Elsevier: Amsterdam, The Netherlands, 1977; pp. 173–183. [Google Scholar]
- Bibiloni, M.A.; Uriz, M.J.; Gili, J.M. Sponge Communities in Three Submarine Caves of the Balearic Islands (Western Mediterranean): Adaptations and Faunistic Composition. Mar. Ecol. 1989, 10, 317–334. [Google Scholar] [CrossRef]
- Pansini, M. Petrosia pulitzeri n.sp. (Porifera, Demospongiae) from Mediterranean Caves. Ital. J. Zool. 1996, 63, 169–172. [Google Scholar] [CrossRef]
- Gerovasileiou, V.; Voultsiadou, E. Sponge Diversity Gradients in Marine Caves of the Eastern Mediterranean. J. Mar. Biol. Assoc. U. K. 2016, 96, 407–416. [Google Scholar] [CrossRef]
- Knittweis, L.; Chevaldonné, P.A.; Ereskovsky, A.; Schembri, J.J.; Borg, J.A. A Preliminary Survey of Marine Cave Habitats in the Maltese Islands. Xjenza Online 2015, 3, 153–164. [Google Scholar]
- Dimarchopoulou, D.; Gerovasileiou, V.; Voultsiadou, E. Spatial Variability of Sessile Benthos in a Semi-Submerged Marine Cave of a Remote Aegean Island (Eastern Mediterranean Sea). Reg. Stud. Mar. Sci. 2018, 17, 102–111. [Google Scholar] [CrossRef]
- Digenis, M.; Arvanitidis, C.; Dailianis, T.; Gerovasileiou, V. Comparative Study of Marine Cave Communities in a Protected Area of the South-Eastern Aegean Sea, Greece. J. Mar. Sci. Eng. 2022, 10, 660. [Google Scholar] [CrossRef]
- Ereskovsky, A.; Kovtun, O.A.; Pronin, K.K.; Apostolov, A.; Erpenbeck, D.; Ivanenko, V. Sponge Community of the Western Black Sea Shallow Water Caves: Diversity and Spatial Distribution. PeerJ 2018, 6, e4596. [Google Scholar] [CrossRef] [PubMed]
- Balduzzi, A.; Bianchi, C.N.; Boero, F.; Cattaneo Vietti, R.; Pansini, M.; Sará, M. The Suspension-Feeder Communities of a Mediterranean Sea Cave. Sci. Mar. 1989, 53, 387–395. [Google Scholar]
- Russ, K.; Rützler, K. Zur Kenntnis Der Schwammfauna Unterseeischer Höhlen. Pubbl. Stn. Zool. Napoli 1959, 30, 756–787. [Google Scholar]
- Rastorgueff, P.-A.; Bellan-Santini, D.; Bianchi, C.N.; Bussotti, S.; Chevaldonné, P.; Guidetti, P.; Harmelin, J.-G.; Montefalcone, M.; Morri, C.; Perez, T.; et al. An Ecosystem-Based Approach to Evaluate the Ecological Quality of Mediterranean Undersea Caves. Ecol. Ind. 2015, 54, 137–152. [Google Scholar] [CrossRef]
- Díaz, J.A.; Ordines, F.; Massutí, E.; Cárdenas, P. From Caves to Seamounts: The Hidden Diversity of Tetractinellid Sponges from the Balearic Islands, with the Description of Eight New Species. PeerJ 2024, 12, e16584. [Google Scholar] [CrossRef]
- Vacelet, J.; Boury-Esnault, N.; Harmelin, J.-G. Hexactinellid Cave, a Unique Deep-Sea Habitat in the Scuba Zone. Deep. Sea Res. Part I Oceanogr. Res. Pap. 1994, 41, 965–973. [Google Scholar] [CrossRef]
- Relini, G. Le Specie Protette del Protocollo SPA/BIO (Convenzione di Barcellona) Presenti in Italia: Schede Descrittive per L’Identificazione; Erredi: Genoa, Italy, 2009. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Distance from Entrance (m) | GF | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| 0 | 5 | 10 | 15 | 20 | 25 | 30 | 35 | ||
| Calcarea | |||||||||
| Subclass: Calcinea | |||||||||
| Order: Clathrinidae | |||||||||
| Leucetta solida (Schmidt, 1862) | * | M | |||||||
| Subclass: Calcaronea | |||||||||
| Order: Leucosolenida | |||||||||
| Sycon raphanus Schmidt, 1862 # | * | Er | |||||||
| Homoscleromorpha | |||||||||
| Order: Homosclerophorida | |||||||||
| Oscarella lobularis (Schmidt, 1862) | * | M | |||||||
| Plakina trilopha Schulze, 1880 | * | En | |||||||
| Plakortis simplex Schulze, 1880 | * | * | En | ||||||
| Demospongiae | |||||||||
| Subclass: Verongimorpha | |||||||||
| Order: Chondrosida | |||||||||
| Chondrosia reniformis Nardo, 1847 | * | * | * | * | * | * | * | * | M |
| Order: Verongida | |||||||||
| Aplysina aerophoba (Nardo, 1833) | * | * | * | * | * | * | * | * | Er |
| Subclass: Keratosa | |||||||||
| Order: Dictyoceratida | |||||||||
| Ircinia oros (Schmidt, 1864) | * | M | |||||||
| Ircinia variabilis (Schmidt, 1862) | * | * | * | * | * | * | M | ||
| Sarcotragus spinosulus Schmidt, 1862 | * | * | M | ||||||
| Dysidea fragilis (Montagu, 1814) | * | En | |||||||
| Subclass: Heteroscleromorpha | |||||||||
| Order: Haplosclerida | |||||||||
| Dendroxea lenis (Topsent, 1892) | * | * | * | * | En | ||||
| Haliclona (Rhizoniera) rosea (Bowebank, 1866) | * | M | |||||||
| Petrosia (Petrosia) ficiformis (Poiret, 1789) | * | * | * | * | * | M | |||
| Petrosia (Strongylophora) cf. vansoesti Boury-Esnault, Pansini, and Uriz, 1994 | * | M | |||||||
| Siphonodictyon infestum (Johnson, 1899) #,§ | * | * | Ex | ||||||
| Order: Axinellida | |||||||||
| Eurypon sp. | * | En | |||||||
| Order: Bubarida | |||||||||
| Bubaris vermiculata (Bowerbank, 1866) | * | * | * | * | * | En | |||
| Dictyonella marsillii (Topsent, 1893) | * | M | |||||||
| Order: Biemnida | |||||||||
| Rhabderemia topsenti van Soest and Hooper, 1993 | * | * | En | ||||||
| Order: Tetractinellida | |||||||||
| Dercitus (Stoeba) plicatus (Schmidt, 1868) | * | * | In | ||||||
| Jaspis johnstonii (Schmidt, 1862) | * | Ex, In | |||||||
| Stelletta lactea Carter, 1871 #,Ø | * | * | En | ||||||
| Erylus discophorus (Schmidt, 1862) | * | * | * | * | * | * | En | ||
| Geodia cydonium (Linnaeus, 1767) | * | * | * | * | * | * | M | ||
| Penares helleri (Schmidt, 1864) | * | * | * | M | |||||
| Alectona millari Carter, 1879 | * | * | * | Ex | |||||
| Thoosa mollis Volz, 1939 | * | * | * | * | Ex | ||||
| Order: Poecilosclerida | |||||||||
| Crambe crambe (Schmidt, 1862) | * | En | |||||||
| Hamigera hamigera (Schmidt, 1862) #,Ø | * | En | |||||||
| Hymedesmia (Hymedesmia) peachii Bowerbank, 1882 | * | En | |||||||
| Hymedesmia sp. | * | * | * | En | |||||
| Phorbas fictitius (Bowerbank, 1866) | * | * | * | * | * | * | En | ||
| Phorbas tenacior (Topsent, 1925) | * | * | * | * | * | En | |||
| Clathria (Microciona) strepsitoxa (Hope, 1889) # | * | En | |||||||
| Mycale (Mycale) lingua (Bowerbank, 1866) | * | En | |||||||
| Tedania (Tedania) anhelans (Vio in Olivi, 1792) | * | * | En | ||||||
| Order: Clionaida | |||||||||
| Cliona celata Grant, 1826 | * | * | * | Ex | |||||
| Cliona rhodensis Rützler and Bromley, 1981 | * | * | * | * | * | * | Ex | ||
| Cliona schmidtii (Ridley, 1881) | * | * | * | * | * | * | * | * | Ex |
| Cliona vermifera Hancock, 1867 | * | * | * | Ex | |||||
| Cliona viridis (Schmidt, 1862) | * | * | * | * | Ex | ||||
| Cliona sp. | * | Ex | |||||||
| Spirastrella cunctatrix Schmidt, 1868 | * | * | * | * | * | En | |||
| Order: Tethyida | |||||||||
| Tethya aurantium (Pallas, 1766) | * | * | * | M | |||||
| Timea crassa (Topsent, 1900) #,Ø | * | En | |||||||
| Timea fasciata Topsent, 1934 | * | En | |||||||
| Timea stellata (Bowerbank,1866) | * | * | En | ||||||
| Order: Suberitida | |||||||||
| Aaptos aaptos (Schmidt, 1864) | * | * | * | * | * | En | |||
| Pseudosuberites sulphureus (Bowerbank, 1866) | * | En | |||||||
| Terpios gelatinosus (Bowerbank, 1866) | * | * | * | * | * | * | * | En | |
| Halichondria (Halichondria) contorta (Sarà, 1961) | * | En | |||||||
| Halichondriidae | * | En | |||||||
| Topsentia cf. lacazei (Schmidt, 1868) | * | En | |||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schiavo, A.; Oddenino, M.; Giménez, G.; Longo, C. First Survey of the Sponge Community of a Semi-Submerged Marine Cave along the Adriatic Apulian Coast. J. Mar. Sci. Eng. 2024, 12, 682. https://doi.org/10.3390/jmse12040682
Schiavo A, Oddenino M, Giménez G, Longo C. First Survey of the Sponge Community of a Semi-Submerged Marine Cave along the Adriatic Apulian Coast. Journal of Marine Science and Engineering. 2024; 12(4):682. https://doi.org/10.3390/jmse12040682
Chicago/Turabian StyleSchiavo, Antonella, Muriel Oddenino, Guadalupe Giménez, and Caterina Longo. 2024. "First Survey of the Sponge Community of a Semi-Submerged Marine Cave along the Adriatic Apulian Coast" Journal of Marine Science and Engineering 12, no. 4: 682. https://doi.org/10.3390/jmse12040682
APA StyleSchiavo, A., Oddenino, M., Giménez, G., & Longo, C. (2024). First Survey of the Sponge Community of a Semi-Submerged Marine Cave along the Adriatic Apulian Coast. Journal of Marine Science and Engineering, 12(4), 682. https://doi.org/10.3390/jmse12040682