


Sponge Communities of Submarine Caves and Tunnels on the Fernando de Noronha Archipelago, Northeast Brazil
 , , , , , ,
, , , , , ,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
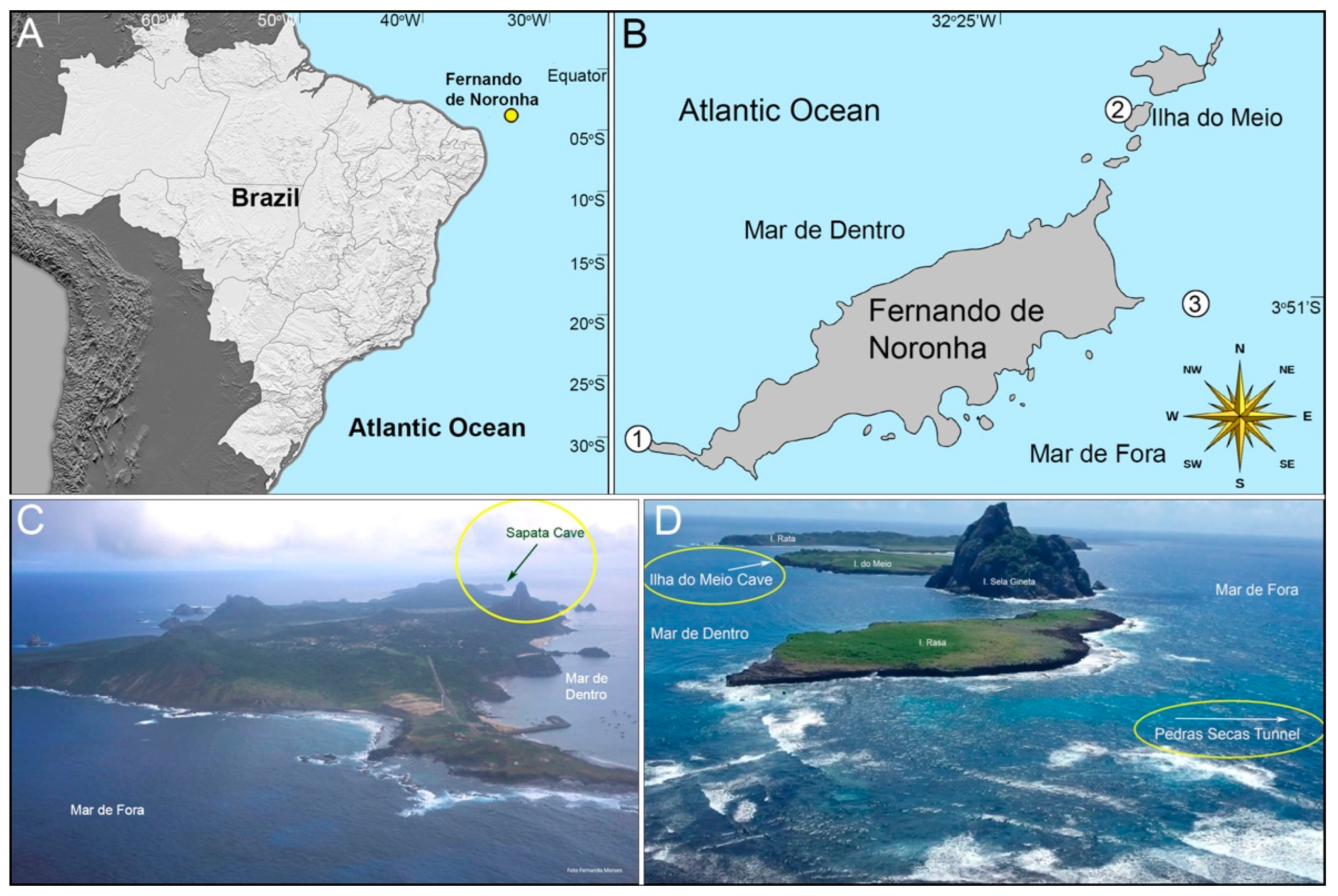
2.1. Study Area
2.2. Methods
3. Results
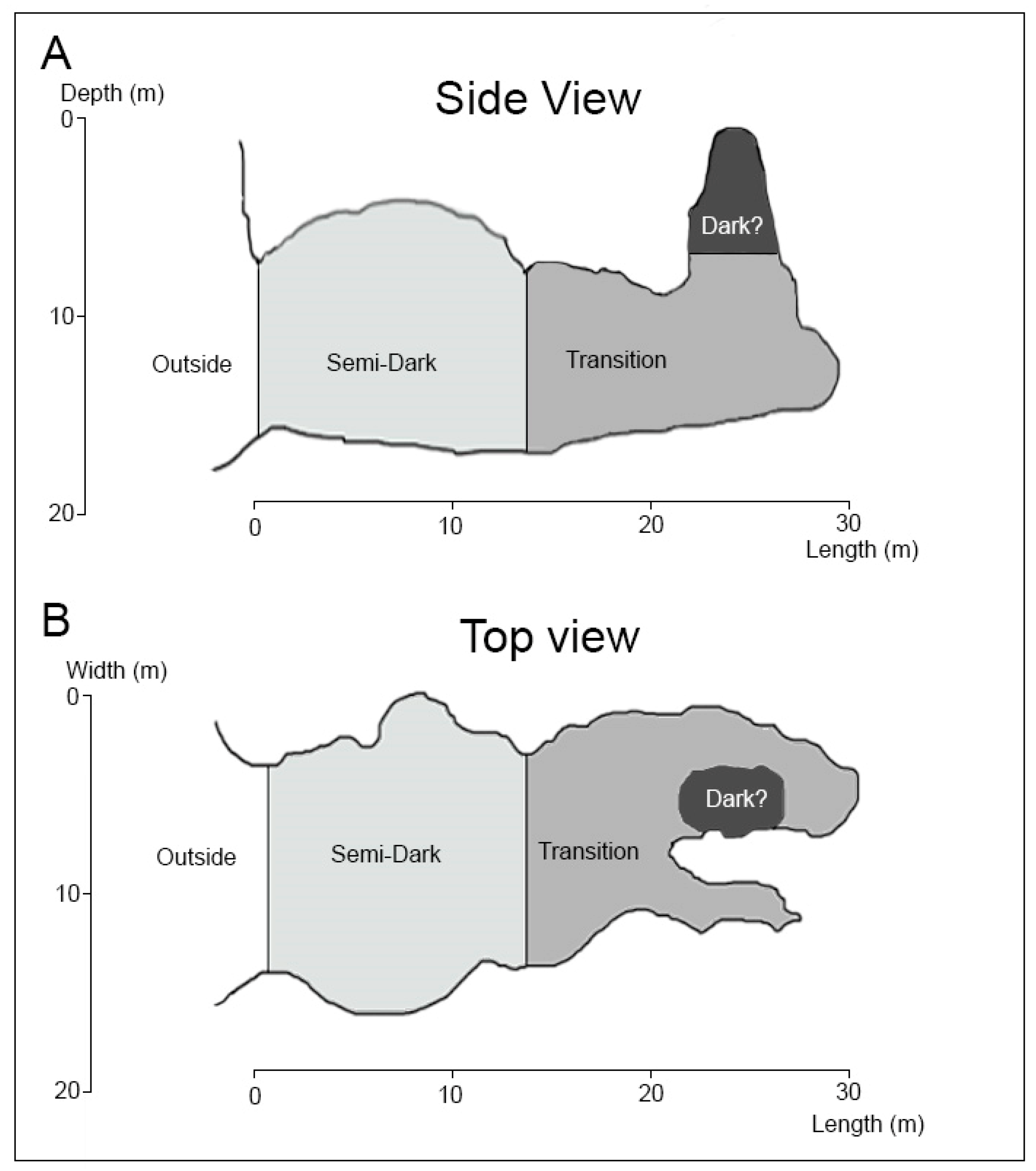
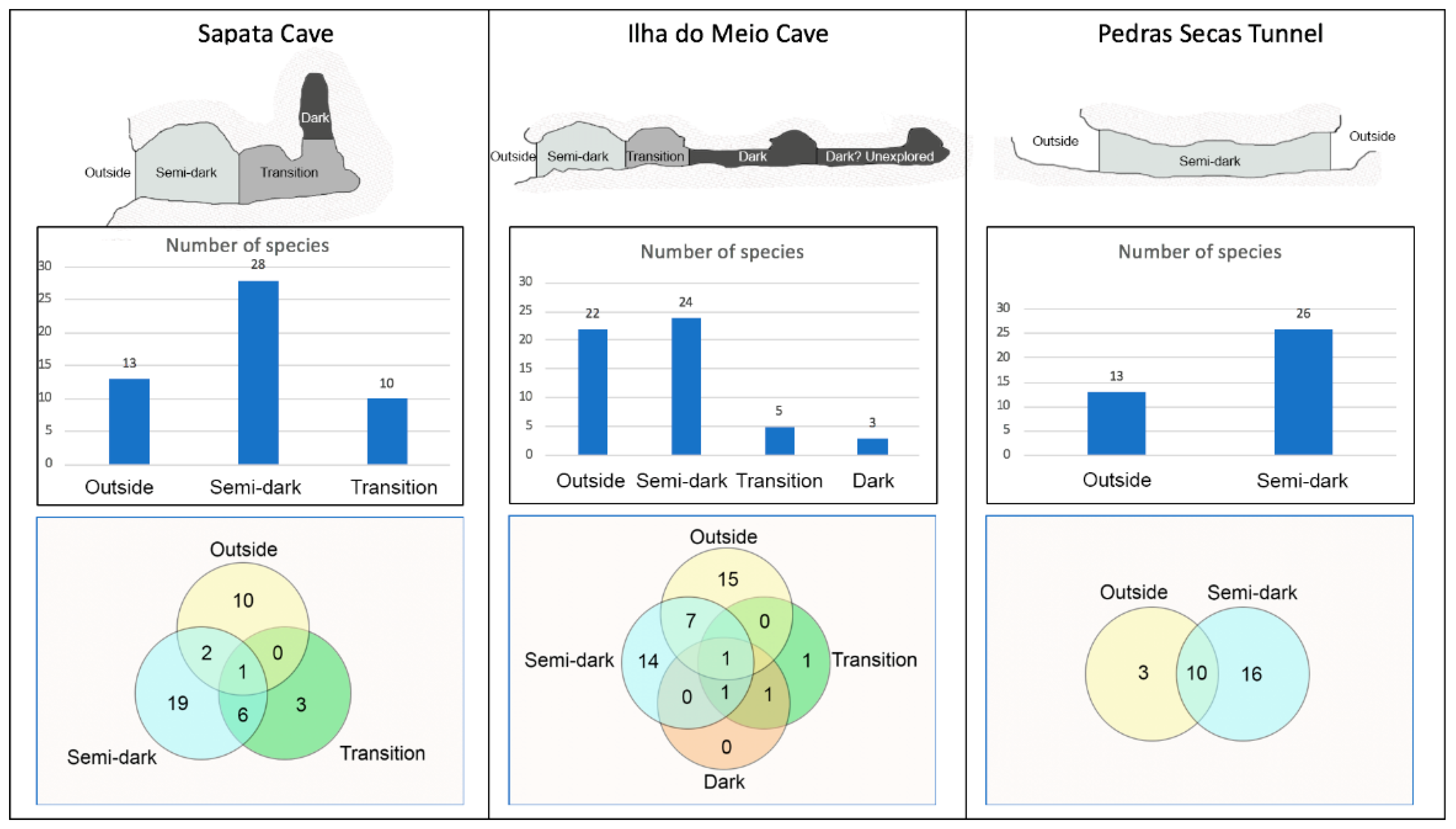
3.1. Sapata Cave
3.1.1. Description of the Cave
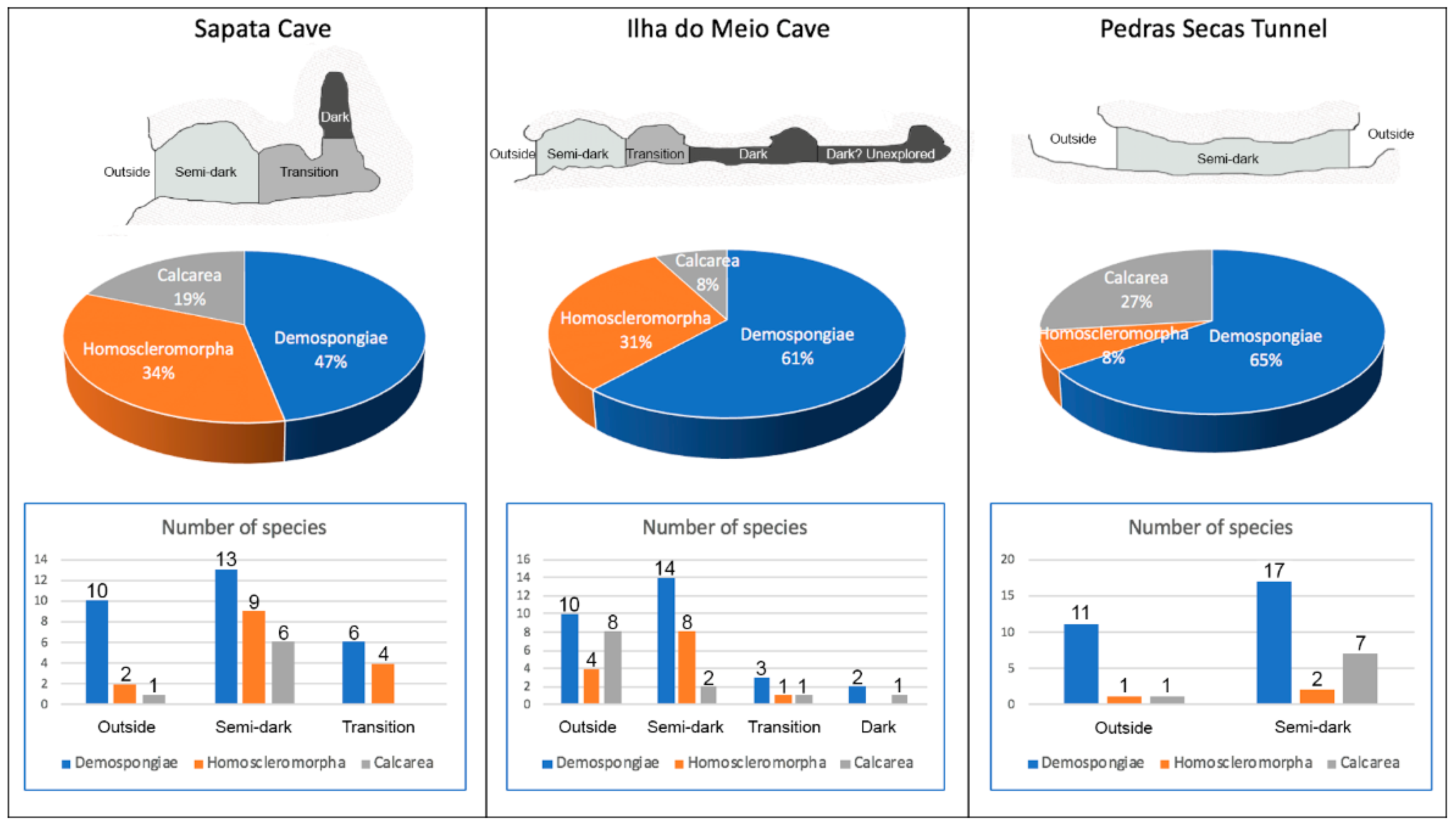
3.1.2. Sponge Communities of Sapata Cave
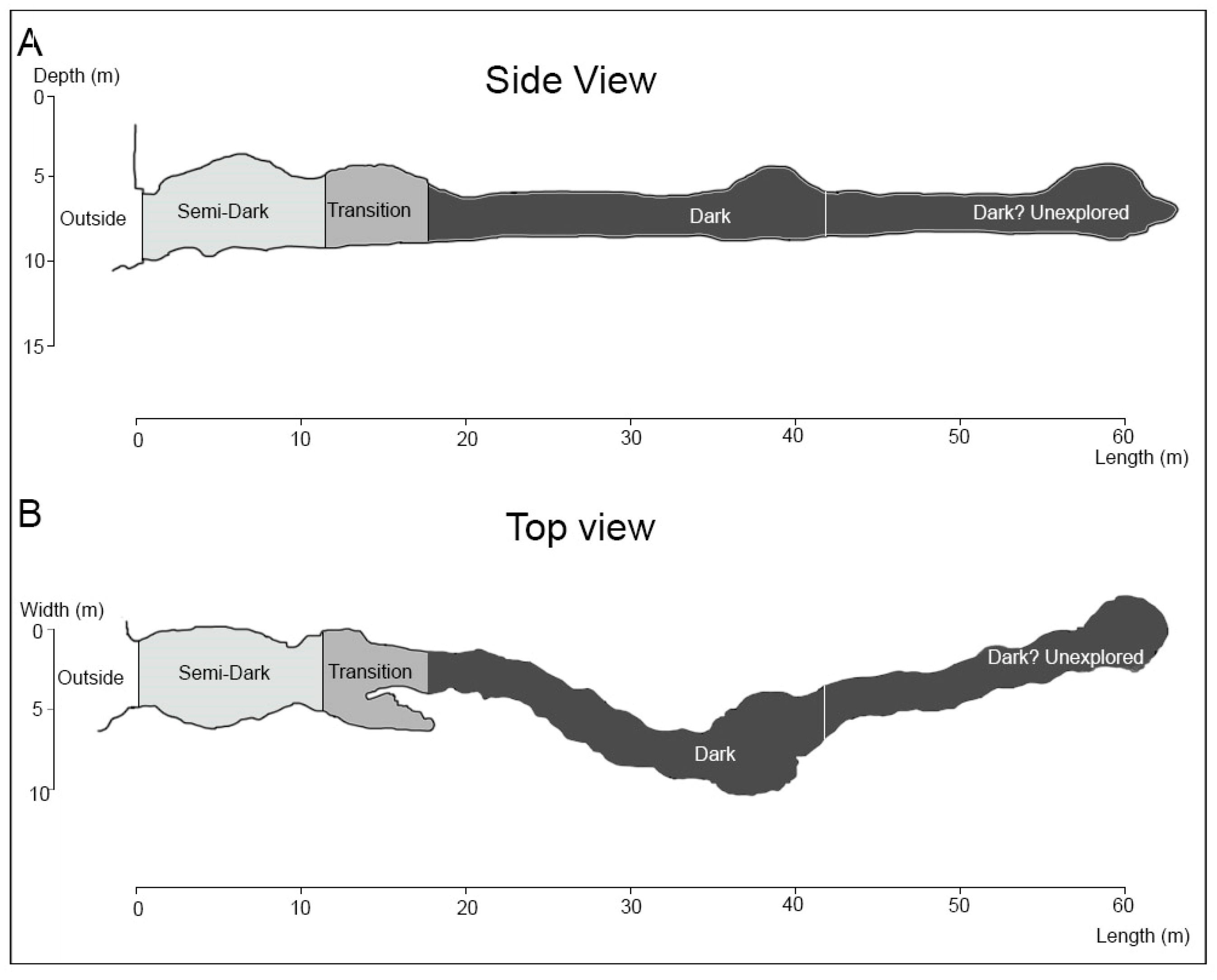
3.2. Ilha do Meio Cave
3.2.1. Description of the Cave
3.2.2. Sponge Communities of Ilha Do Meio Cave
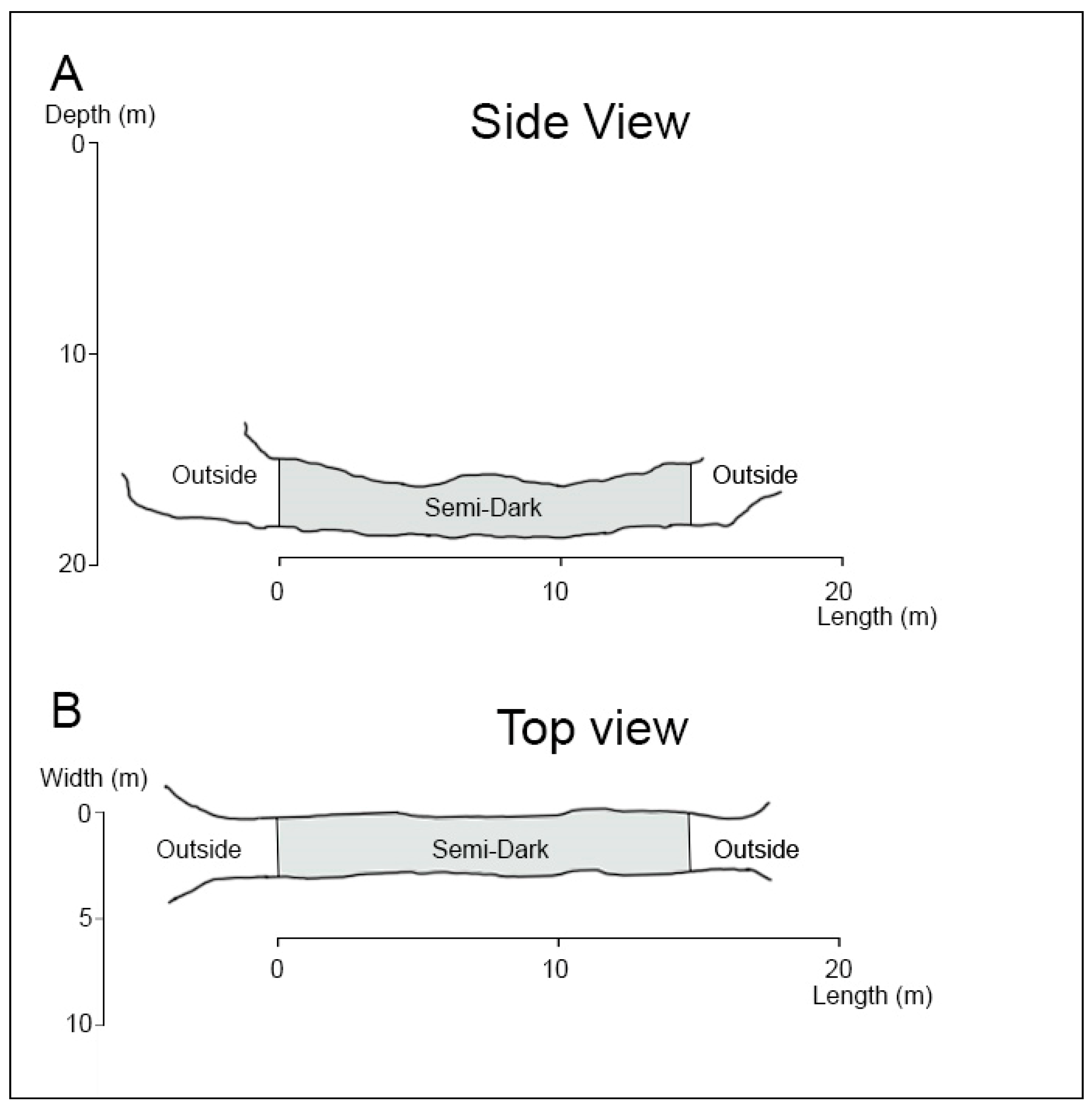
3.3. Pedras Secas Tunnel
3.3.1. Description of the Tunnel
3.3.2. Sponge Communities of Pedras Secas Tunnel
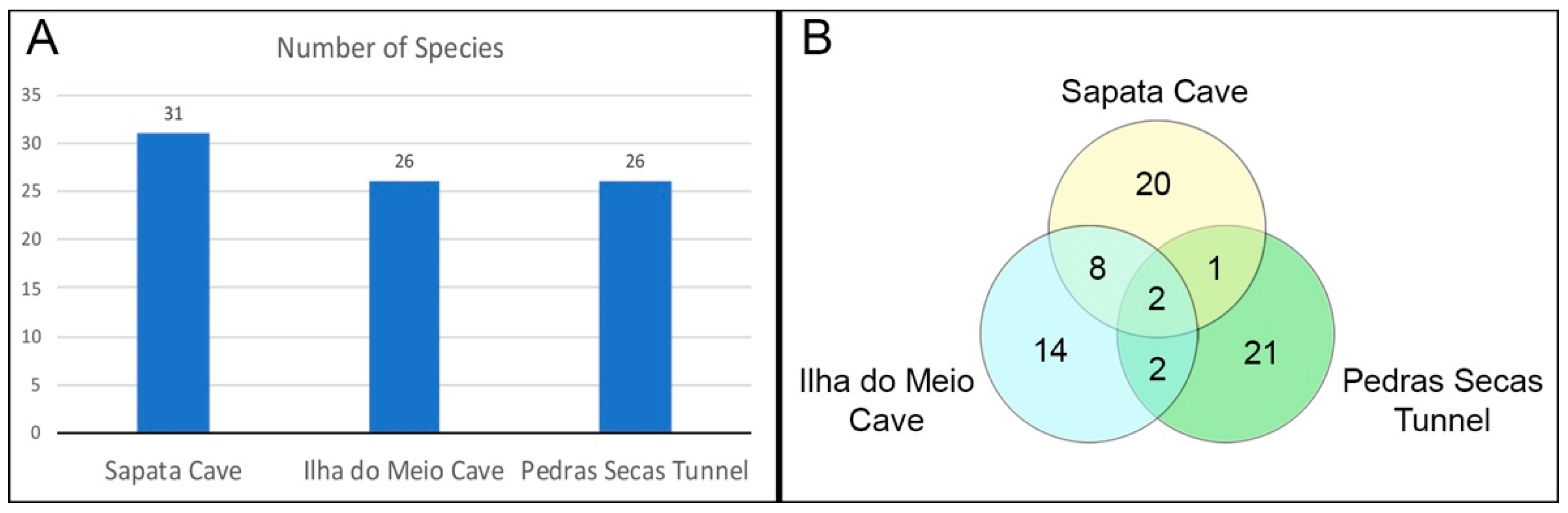
3.4. Species Richness and Composition
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kempe, S.; Rosendahl, W.; Wiegand, B.; Eisenhauer, A. New speleothem dates from caves in Germany and their importance for the Middle and Upper Pleistocene climate reconstruction. Acta Geol. Polón 2002, 52, 55–61. [Google Scholar]
- Culver, D.C.; Sket, B. Hotspots of subterranean biodiversity in caves and wells. J. Cave Karst Stud. 2005, 62, 11–17. [Google Scholar]
- Cigna, A.A.; Forti, P. Caves: The most important geotouristic feature in the world. Tour. Karst Ar. 2013, 6, 9–26. [Google Scholar]
- Zepon, T.; Bichouette, M.E. Influence of substrate on the richness and composition of neotropical cave fauna. An. Acad. Bras. Ciênc. 2017, 89, 1615–1628. [Google Scholar] [CrossRef]
- Trajano, E. Cave faunas in the Atlantic tropical rain forest: Composition, ecology and conservation. Biotropica 2000, 32, 882–893. [Google Scholar] [CrossRef]
- Gerovasileiou, V.; Bianchi, C.N. Mediterranean marine caves: A synthesis of current knowledge. Oceanogr. Mar. Biol. Ann. Rev. 2021, 59, 87. [Google Scholar] [CrossRef]
- Álvarez, F.; Durán, B.; Meacham, S. Anchialine fauna of the Yucatan Peninsula: Diversity and conservation challenges. In Mexican Fauna in the Anthropocene; Jones, R.W., Ornelas-García, C.P., Pineda-López, R., Álvarez, F., Eds.; Springer: Cham, Switzerland, 2023; pp. 287–301. [Google Scholar] [CrossRef]
- Laborel, J.; Vacelet, J. Étude des peuplements d’une grotte sous-marine du golfe de Marseille. Bull. Inst. Océanogr. 1958, 1120, 1–20. [Google Scholar]
- Gerovasileiou, V.; Voultsiadou, E. Marine caves of the Mediterranean Sea: A sponge biodiversity reservoir within a biodiversity hotspot. PLoS ONE 2012, 7, e39873. [Google Scholar] [CrossRef]
- Harmelin, J.G.; Vacelet, J.; Vasseur, P. Les grottes sous-marines obscures: Un milieu extrême et un remarquable biotope refuge. Téthys 1985, 11, 214–229. [Google Scholar]
- Rastorgueff, P.A.; Bellan-Santini, D.; Bianchi, C.N.; Bussotti, S.; Chevaldonné, P.; Guidetti, P.; Harmelin, J.G.; Montefalcone, M.; Morri, C.; Pérez, T.; et al. An ecosystem-based approach to evaluate the ecological quality of Mediterranean undersea caves. Ecol. Indic. 2015, 54, 137–152. [Google Scholar] [CrossRef]
- Vacelet, J.; Boury-Esnault, N.; Harmelin, J.-G. Hexactinellid cave, a unique deep-sea habitat in the scuba zone. Deep Sea Res. 1994, 41, 965–973. [Google Scholar] [CrossRef]
- Vacelet, J. Deep-sea sponges in a Mediterranean cave. In Deep-Sea and Extreme Shallow-Water Habitats: Affinities and Adaptations; Uiblein, F., Ott, J., Stachowitsch, M., Eds.; Österreichische Akademie der Wissenschaften: Vienna, Austria, 1996; Volume 11, pp. 299–312. [Google Scholar]
- Bell, J.J. The sponge community in a semi-submerged temperate sea cave: Density, diversity and richness. PSZN Mar. Ecol. 2002, 23, 297–311. [Google Scholar] [CrossRef]
- Gerovasileiou, V.; Voultsiadou, E. Sponge diversity gradients in marine caves of the eastern Mediterranean. J. Mar. Biol. Ass. UK 2016, 96, 407–416. [Google Scholar] [CrossRef]
- Cleary, D.F.R.; Yusheng, M.; Huang, Y.M.; Polónia, A.R.M.; van der Plas, M.; Gomes, N.C.M.; de Voogd, N.J. Sponges and their prokaryotic communities sampled from a remote karst ecosystem. Mar. Biodiv. 2024, 54, 8. [Google Scholar] [CrossRef]
- Manning, R.B.; Hart, C.W., Jr.; Iliffe, T.M. Mesozoic relicts in marine caves of Bermuda. Stygologia 1986, 2, 156–166. [Google Scholar]
- Iliffe, T.M.; Kornicker, L.S. Worldwide diving discoveries of living fossil animals from the depths of anchialine and marine caves. Smithson. Contrib. Mar. Sci. 2009, 38, 269–280. [Google Scholar]
- Vacelet, J. Recent Calcarea with reinforced skeleton (“Pharetronids”). In Fossil and Recent Sponges; Reitner, J., Keupp, H., Eds.; Springer: Berlin/Heidelberg, Germany, 1991; pp. 252–265. [Google Scholar] [CrossRef]
- Vacelet, J. Recent ‘Sphinctozoa’. Family Verticillitidae Steinmann, 1882. In Systema Porifera: A Guide to the Classification of Sponges; Hooper, J.N.A., Van Soest, R.W.M., Eds.; Kluwer Academic/Plenum Publishers: New York, NY, USA, 2002; pp. 1109–1110. [Google Scholar]
- Pisera, A.; Vacelet, J. Lithistid sponges from submarine caves in the Mediterranean: Taxonomy and affinities. Sci. Mar. 2011, 75, 17–40. [Google Scholar] [CrossRef]
- Hart, C.W., Jr.; Manning, R.B.; Iliffe, T.M. The fauna of Atlantic marine caves: Evidence of dispersal by sea floor spreading while maintaining ties to deep waters. Proc. Biol. Soc. Wash. 1985, 98, 288–292. [Google Scholar]
- Rützler, K.; Piantoni, C.; Van Soest, R.W.M.; Díaz, M.C. Diversity of sponges (Porifera) from cryptic habitats on the Belize barrier reef near Carrie Bow Cay. Zootaxa 2014, 3805, 1–129. [Google Scholar] [CrossRef]
- Iliffe, T.M.; Álvarez, F. Chapter 18. Research in anchialine caves. In Cave Ecology; Moldovan, O.T., Kováč, L., Halse, S., Eds.; Springer: Cham, Switzerland, 2018; Volume 235, pp. 383–397. [Google Scholar] [CrossRef]
- Ise, Y. Preliminary report of submarine cave sponges in Shimoji Islands, Miyako Islands, Okinawa. Taxa 2019, 46, 13–17. [Google Scholar]
- Gerovasileiou, V.; Martínez García, A.; Álvarez Noguera, F.; Boxshall, G.; Humphreys, W.F.; Jaume, D.; Becking, L.E.; Muricy, G.; van Hengstum, P.J.; Yamasaki, H.; et al. World Register of Marine Cave Species (WoRCS). Available online: https://www.marinespecies.org/worcs (accessed on 5 March 2024).
- Volkmer-Ribeiro, C.; Bichuette, M.E.; Machado, K.F. Racekiela cavernicola (Porifera: Demospongiae) new species and the first record of cave freshwater sponge from Brazil. Neotrop. Biol. Conserv. 2010, 5, 53–58. [Google Scholar] [CrossRef]
- Leite, T.S.; Pinheiro, I.E.; Berchez, F.; Bertoncini, A.A.; del Cima, O.M.; Demetrescu, I.E.; Francini-Filho, R.B.; Kikuchi, R.K.P.; Machado, A.A.; Maia-Nogueira, R.; et al. Scientific diving in Brazil: History, present and perspectives. Oc. Coast. Res. 2023, 71, e23045. [Google Scholar] [CrossRef]
- Bertoncini, A.A.; Moraes, F.; Borgonha, M.; Aguiar, A.; Duarte, B. Guia de Biodiversidade Marinha e Mergulho das Ilhas do Rio; Museu Nacional: Rio de Janeiro, Brazil, 2019; pp. 1–354. [Google Scholar]
- Muricy, G.; Hajdu, E.; Custódio, M.; Klautau, M.; Russo, C.A.M.; Peixinho, S. Sponge distribution at Arraial do Cabo, SE Brazil. In Proceedings of the Coastal Zone 1991. VII Symposium on Coastal and Ocean Management, Long Beach, CA, USA, 8–12 July 1991; Magoon, O.T., Ed.; ASCE Publications: Long Beach, CA, USA, 1991; Volume 2, pp. 1183–1196. [Google Scholar]
- Mothes, B.; Hajdu, E.; van Soest, R.W.M. Tedania brasiliensis new species (Demospongiae, Poecilosclerida, Tedaniidae) from Brazil, with some remarks about the genus Tedania in the tropical southwestern Atlantic. Bull. Mar. Sci. 2000, 66, 1–11. [Google Scholar]
- Esteves, E.L.; de Paula, T.S.; Lerner, C.; Lôbo-Hajdu, G.; Hajdu, E. Morphological and molecular systematics of the ‘Monanchora arbuscula complex’ (Poecilosclerida: Crambeidae), with the description of five new species and a biogeographic discussion of the genus in the Tropical Western Atlantic. Invert. Syst. 2018, 32, 457–503. [Google Scholar] [CrossRef]
- Muricy, G.; Moraes, F.C. Marine sponges of Pernambuco State, NE Brazil. Rev. Bras. Oceanogr. 1998, 46, 213–217. [Google Scholar] [CrossRef]
- Muricy, G.; Minervino, J.V. A new species of Gastrophanella from central western Atlantic, with discussion of the family Siphoniidae (Demospongiae: Lithistida). J. Mar. Biol. Ass. UK 2000, 80, 599–605. [Google Scholar] [CrossRef]
- Moraes, F.C.; Muricy, G. Taxonomy of Plakortis and Plakinastrella (Demospongiae: Plakinidae) from oceanic islands of North-Eastern Brazil, with description of three new species. J. Mar. Biol. Ass. UK 2003, 83, 385–397. [Google Scholar] [CrossRef]
- Moraes, F.C.; Muricy, G. A new species of Stoeba (Demospongiae: Astrophorida) from oceanic islands off north-eastern Brazil. J. Mar. Biol. Ass. UK 2007, 87, 1387–1393. [Google Scholar] [CrossRef]
- Moraes, F.C. Esponjas das Ilhas Oceânicas Brasileiras; Série Livros 44; Museu Nacional: Rio de Janeiro, Brazil, 2011; pp. 1–252. [Google Scholar]
- Domingos, C.; Moraes, F.; Muricy, G. Four new species of Plakinidae (Porifera: Homoscleromorpha) from Brazil. Zootaxa 2013, 3718, 530–544. [Google Scholar] [CrossRef] [PubMed]
- Azevedo, F.C.; Padua, A.Q.; Moraes, F.; Rossi, A.L.; Muricy, G.; Klautau, M. Taxonomy and phylogeny of calcareous sponges (Porifera: Calcarea: Calcinea) from Brazilian mid-shelf and oceanic islands. Zootaxa 2017, 4311, 301–344. [Google Scholar] [CrossRef]
- Cordani, U. Idade do vulcanismo no Oceano Atlântico Sul. Bol. Inst. Geociênc. Astron. 1970, 1, 9–75. [Google Scholar] [CrossRef]
- ICMBio. Fernando de Noronha. Available online: https://www.parnanoronha.com.br (accessed on 6 March 2024).
- IBGE. Fernando de Noronha. Available online: https://www.ibge.gov.br/cidades-e-estados/pe/fernando-de-noronha.html (accessed on 6 March 2024).
- Vale, T.F. A Gestão do Território e os Benefícios de um Geopark: Ações Visando a Implantação do Projeto Geopark Fernando de Noronha. Master’s Thesis, Universidade Estadual de Ponta Grossa, Ponta Grossa, Brazil, 2017. [Google Scholar]
- Moreira, J.C.; Silva Júnior, J.M. Trilhas subaquáticas em Fernando de Noronha—PE: A importância da interpretação ambiental relacionada à geodiversidade. In Proceedings of the Congresso Nacional de Planejamento e Manejo de Trilhas and Colóquio Brasileiro da Red Latino Americana de Senderismo, Rio de Janeiro, Brazil, 16–18 October 2013; pp. 1203–1219. [Google Scholar]
- Hildebrand, S.F.; Schroeder, W.C. Fishes of Chesapeake Bay. Bull. US. Bur. Fish. 1928, 43, 1–366. [Google Scholar]
- Bertoncini, A.; Giglio, V.; Alves, J.; Grossman, A.; Matheus, Z.; Ferreira, B. Projeto Meros do Brasil Photo-ID: First insights of Goliath Grouper (Epinephelus itajara, Epinephelidae) population and movements at Fernando de Noronha National Marine Park. In Proceedings of the Gulf and Caribbean Fisheries Institute 66th Annual Conference, Corpus Christi, TX, USA, 4–8 November 2013; pp. 297–300. [Google Scholar]
- Schmidt, O. Grundzüge Einer Spongien-Fauna des Atlantischen Gebietes; Wilhelm Engelmann: Leipzig, Germany, 1870; pp. 1–88. [Google Scholar]
- Duchassaing de Fonbressin, P.; Michelotti, G. Spongiaires de la mer Caraïbe. Natuurk. Verhand. Holl. Maatsch. Wetensch. Haarlem 1864, 21, 1–124. [Google Scholar]
- Hyatt, A. Revision of the North American Poriferae; with remarks upon foreign species. Part I. Mem. Boston Soc. Nat. Hist. 1875, 2, 399–408. [Google Scholar]
- Pallas, P.S. Elenchus Zoophytorum Sistens Generum Adumbrations Generaliores et Specierum Cognitarum Succinctas Descriptiones cum Selectis Auctorum Synonymis; Petrus van Cleef: The Hague, The Netherlands, 1766; pp. 1–451. [Google Scholar]
- Rützler, K.; Duran, S.; Piantoni, C. Adaptation of reef and mangrove sponges to stress: Evidence for ecological speciation exemplified by Chondrilla caribensis new species (Demospongiae, Chondrosida). Mar. Ecol. 2007, 28 (Suppl. S1), 95–111. [Google Scholar] [CrossRef]
- Soest, R.W.M. van. Marine sponges from Curaçao and other Caribbean localities. Part III. Poecilosclerida. Stud. Fau. Curaçao Caribb. Isl. 1984, 199, 1–167. [Google Scholar]
- Pulitzer-Finali, G. A collection of West Indian Demospongiae (Porifera). In appendix, a list of the Demospongiae hitherto recorded from the West Indies. Ann. Mus. Civ. Stor. Nat. Genova 1986, 18, 65–216. [Google Scholar]
- Hechtel, G.J. A systematic study of the Demospongiae of Port Royal, Jamaica. Bull. Peabody Mus. Nat. Hist. 1965, 20, 1–94. [Google Scholar]
- Ridley, S.O.; Dendy, A. Preliminary report on the Monaxonida collected by H.M.S. ‘Challenger’. Ann. Mag. Nat. Hist. 1886, 18, 470–493. [Google Scholar] [CrossRef]
- Sollas, W.J. Preliminary account of the Tetractinellid sponges dredged by H.M.S. ‘Challenger 1872–76. Part I. The Choristida. Sci. Proc. Roy. Dublin Soc. 1886, 5, 177–199. [Google Scholar]
- Vacelet, J.; Donadey, C. A new species of Halisarca (Porifera, Demospongiae) from the Caribbean, with remarks on the cytology and affinities of the genus. In European Contributions to the Taxonomy of Sponges; Jones, W.C., Ed.; Sherkin Island Marine Station: Cork, Ireland, 1987; Volume 1, pp. 5–12. [Google Scholar]
- Lamarck, J.B.P.; de Monet, C. Histoire Naturelle des Animaux sans Vertèbres, Présentant les Caractères Généraux et Particuliers de ces Animaux, leur Distribution, leurs Classes, leurs Familles, leurs Genres, et la Citation des Principales Espèces qui s’y Rapportent; Verdière: Paris, France, 1816; Volume 2, pp. 1–568. [Google Scholar]
- Soest, R.W.M. van. Marine sponges from Curaçao and other Caribbean localities. Part II. Haplosclerida. Stud. Fau. Curaçao Caribb. Isl. 1980, 62, 1–173. [Google Scholar]
- Wiedenmayer, F. Shallow-water sponges of the western Bahamas. Experientia 1977, 28, 1–287. [Google Scholar]
- Boury-Esnault, N.; Klautau, M.; Bézac, C.; Wulff, J.; Solé-Cava, A.M. Comparative study of putative conspecific sponge populations from both sides of the Isthmus of Panama. J. Mar. Biol. Ass. UK 1999, 79, 39–50. [Google Scholar] [CrossRef]
- de Laubenfels, M.W. New sponges from the Puerto Rican deep. Smithson. Misc. Coll. 1934, 91, 1–28. [Google Scholar]
- Lanna, E.; Rossi, A.L.; Cavalcanti, F.F.; Hajdu, E.; Klautau, M. Calcareous sponges from São Paulo State, Brazil (Porifera: Calcarea: Calcinea), with the description of two new species. J. Mar. Biol. Ass. UK 2007, 87, 1553–1561. [Google Scholar] [CrossRef]
- Thacker, A.G. On collections of the Cape Verde Islands fauna made by Cyril Crossland, M.A. The calcareous sponges. Proc. Zool. Soc. Lond. 1908, 49, 757–782. [Google Scholar] [CrossRef]
- Solé-Cava, A.M.; Klautau, M.; Boury-Esnault, N.; Borojevic, R.; Thorpe, J.P. Genetic evidence for cryptic speciation in allopatric populations of two cosmopolitan species of the calcareous sponge Clathrina Mar. Biol. 1991, 111, 381–386. [Google Scholar] [CrossRef]
- Wörheide, G.; Hooper, J. Calcarea from the Great Barrier Reef. I: Cryptic Calcinea from Heron Island and Wistari Reef (Capricorn-Bunker group). Mem. Queensl. Mus. 1999, 43, 859–891. [Google Scholar]
- Fontana, T.; Cóndor-Luján, B.; Azevedo, F.; Pérez, T.; Klautau, M. Diversity and distribution patterns of Calcareous sponges (subclass Calcinea) from Martinique. Zootaxa 2018, 4410, 331–369. [Google Scholar] [CrossRef] [PubMed]
- Pérez, T.; Ruiz, C. Description of the first Caribbean Oscarellidae (Porifera: Homoscleromorpha). Zootaxa 2018, 4369, 501–514. [Google Scholar] [CrossRef] [PubMed]
- Cedro, V.R.; Hajdu, E.; Correia, M.D. Three new intertidal sponges (Porifera: Demospongiae) from Brazil’s fringing urban reefs (Maceió, Alagoas, Brazil), and support for Rhabderemia ‘s exclusion from Poecilosclerida. J. Nat. Hist. 2013, 47, 2151–2174. [Google Scholar] [CrossRef]
- Carter, H.J. Contributions to our knowledge of the Spongida. Ann. Mag. Nat. Hist. 1879, 3, 343–360. [Google Scholar] [CrossRef]
- Olivier, A.G. Suite de l’Introduction à l’Histoire Naturelle des Insectes. Palèmon. In Encyclopédie Méthodique. Histoire Naturelle. Insectes; Olivier, A.G., Ed.; H. Agasse, Imprimeur-Libraire: Paris, France, 1811; Volume 8, pp. 656–670. [Google Scholar]
- Rathbun, M.J. A new Scyllarides from Brazil. Proc. Biol. Soc. Wash. 1906, 19, 113. [Google Scholar]
- Martí, R.; Uriz, M.J.; Ballesteros, E.; Turon, X. Benthic assemblages in two Mediterranean caves: Species diversity and coverage as a function of abiotic parameters and geographic distance. J. Mar. Biol. Ass. UK 2004, 84, 557–572. [Google Scholar] [CrossRef]
- Onorato, M.; Belmonte, G. Submarine caves of the Salento Pensinsula: Faunal aspects. Thalassia Sal. 2017, 39, 47–72. [Google Scholar] [CrossRef]
- Bussotti, S.; Terlizzi, A.; Fraschetti, S.; Belmonte, G.; Boero, F. Spatial and temporal variability of sessile benthos in shallow Mediterranean marine caves. Mar. Ecol. Prog. Ser. 2006, 325, 109–119. [Google Scholar] [CrossRef]
- Denitto, F.; Terlizzi, A.; Belmonte, G. Settlement and primary succession in a shallow submarine cave: Spatial and temporal benthic assemblage distinctness. Mar. Ecol. 2007, 28 (Suppl. S1), 35–46. [Google Scholar] [CrossRef]
- Maldonado, M.; Young, C.M. Bathymetric patterns of sponge distribution on the Bahamian slope. Deep-Sea Res. I 1996, 43, 897–915. [Google Scholar] [CrossRef]
- Calle, S.R. Ecological aspects of sponges in mesophotic coral ecosystems. Master’s Thesis, University of Puerto Rico, Mayagüez, PR, USA, 2010. [Google Scholar]
- Pisera, A.; Gerovasileiou, V. Lithistid Demosponges of deep-water origin in marine caves of the north-eastern Mediterranean Sea. Front. Mar. Sci. 2021, 8, 630900. [Google Scholar] [CrossRef]
- Gerovasileiou, V.; Trygonis, V.; Sini, M.; Koutsoubas, D.; Voultsiadou, E. Three-dimensional mapping of marine caves using a handheld echosounder. Mar. Ecol. Progr. Ser. 2013, 486, 13–22. [Google Scholar] [CrossRef]
- Mantas, T.P.; Roveta, C.; Calcinai, B.; Coppari, M.; di Camillo, C.G.; Marchesi, V.; Marrocco, T.; Puce, S.; Cerrano, C. Photogrammetry as a promising tool to unveil marine caves’ benthic assemblages. Sci. Rep. 2023, 13, 7587. [Google Scholar] [CrossRef]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sapata Cave | Ilha do Meio Cave | Pedras Secas Tunnel | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Species | Outside | Semi-Dark Zone | Transition Zone | Outside | Semi-Dark Zone | Transition Zone | Dark Zone | Outside | Semi-Dark Zone |
| CLASS DEMOSPONGIAE | |||||||||
| Agelas clathrodes [47] | x | xx | xxx | ||||||
| Agelas dispar [48] | xx | xxx | x | xx | xxx | ||||
| Agelas sp. 1 | x | ||||||||
| Agelas sp. 2 | x | ||||||||
| Aiolochroia crassa [49] | xx | xx | x | ||||||
| Amphimedon compressa [48] | xx | x | x | ||||||
| Aplysina fulva [50] | x | ||||||||
| Batzella sp. 1 | x | x | |||||||
| Batzella sp. 2 | x | ||||||||
| Chondrilla caribensis [51] | x | ||||||||
| Chondrosia collectrix [47] | x | ||||||||
| Chondrosia sp. | x | ||||||||
| Clathria (Thalysias) minuta [52] | xx | x | |||||||
| Dercitus (Halinastra) luteus [53] | x | ||||||||
| Dercitus (Stoeba) latex [36] | x | ||||||||
| Dictyonella sp. | x | ||||||||
| Diplastrella megastellata [54] | xxx | xx | xx | x | |||||
| Dragmacidon reticulatum [55] | x | ||||||||
| Dysidea sp. | x | ||||||||
| Ectyoplasia ferox [48] | xx | xx | |||||||
| Erylus formosus [56] | x | ||||||||
| Gastrophanella cavernicola [34] | xx | xx | |||||||
| Haliclona sp. | x | ||||||||
| Halisarca caerulea [57] | x | ||||||||
| Hyrtios proteus [48] | x | ||||||||
| Ircinia felix [48] | x | x | xx | ||||||
| Ircinia strobilina [58] | xx | xxx | xx | x | |||||
| Ircinia sp. 1 | x | ||||||||
| Ircinia sp. 2 | x | ||||||||
| Jaspis sp. | x | x | |||||||
| Monanchora arbuscula [48] | xx | xxx | xx | x | |||||
| Neopetrosia sp. | x | ||||||||
| Niphates amorpha [59] | x | ||||||||
| Oceanapia sp. | x | x | xx | ||||||
| Petrosia sp. | x | ||||||||
| Scopalina ruetzleri [60] | x | xx | x | x | |||||
| Spirastrella hartmani [61] | x | xx | x | xx | xxx | x | x | ||
| Suberites sp. | x | ||||||||
| Thorecta sp. | x | ||||||||
| Topsentia ophiraphidites [62] | x | x | xx | x | x | xx | |||
| Verongida unidentified | x | ||||||||
| Xestospongia muta [47] | x | ||||||||
| Xestospongia sp. | x | ||||||||
| CLASS CALCAREA | |||||||||
| Arturia cf. alcatraziensis [63] | x | ||||||||
| Ascandra cf. atlantica [64] | x | ||||||||
| Ascandra sp. | x | xx | x | x | x | ||||
| Calcarea unidentified | x | ||||||||
| Clathrina aurea [65] | x | x | x | ||||||
| Clathrina aff. luteoculcitella [66] | x | x | |||||||
| Clathrina delicata [67] | x | ||||||||
| Clathrina insularis [39] | x | x | |||||||
| Clathrina sp. 1 | x | ||||||||
| Clathrina sp. 2 | x | x | |||||||
| Clathrina sp. 3 | x | ||||||||
| Clathrina sp. 4 | x | ||||||||
| Clathrina sp. 5 | x | ||||||||
| Clathrina sp. 6 | x | ||||||||
| Janusya sp. | x | ||||||||
| Neoernsta sp. 1 | x | ||||||||
| Neoernsta sp. 2 | x | ||||||||
| Neoernsta sp. 3 | x | ||||||||
| CLASS HOMOSCLEROMORPHA | |||||||||
| cf. Aspiculortis sp. | x | ||||||||
| Oscarella cf. zoranja [68] | xx | x | xx | ||||||
| Oscarella sp. 1 | x | xx | xx | ||||||
| Oscarella sp. 2 | x | x | x | ||||||
| Oscarella sp. 3 | x | ||||||||
| cf. Oscarella sp. | x | ||||||||
| Plakina coerulea [69] | x | ||||||||
| Plakina sp. 1 | x | x | |||||||
| Plakina sp. 2 | x | ||||||||
| Plakinastrella microspiculifera [35] | x | xxx | xx | ||||||
| Plakinastrella sp. 1 | x | ||||||||
| Plakinastrella sp. 2 | x | ||||||||
| Plakortis angulospiculatus [70] | xx | xx | x | xxx | |||||
| Plakortis insularis [35] | xxx | x | xx | ||||||
| Plakortis microrhabdifera [35] | x | ||||||||
| Plakortis spinalis [38] | x | ||||||||
| Plakortis sp. | xx | x | |||||||
| Number of species | 13 | 28 | 10 | 22 | 24 | 5 | 3 | 13 | 26 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Muricy, G.; Lage, A.; Sandes, J.; Klautau, M.; Pinheiro, U.; Laport, M.S.; de Oliveira, B.F.R.; Pequeno, C.B.; Lopes, M.V. Sponge Communities of Submarine Caves and Tunnels on the Fernando de Noronha Archipelago, Northeast Brazil. J. Mar. Sci. Eng. 2024, 12, 657. https://doi.org/10.3390/jmse12040657
Muricy G, Lage A, Sandes J, Klautau M, Pinheiro U, Laport MS, de Oliveira BFR, Pequeno CB, Lopes MV. Sponge Communities of Submarine Caves and Tunnels on the Fernando de Noronha Archipelago, Northeast Brazil. Journal of Marine Science and Engineering. 2024; 12(4):657. https://doi.org/10.3390/jmse12040657
Chicago/Turabian StyleMuricy, Guilherme, Anaíra Lage, Joana Sandes, Michelle Klautau, Ulisses Pinheiro, Marinella Silva Laport, Bruno Francesco Rodrigues de Oliveira, Carolline Braga Pequeno, and Matheus Vieira Lopes. 2024. "Sponge Communities of Submarine Caves and Tunnels on the Fernando de Noronha Archipelago, Northeast Brazil" Journal of Marine Science and Engineering 12, no. 4: 657. https://doi.org/10.3390/jmse12040657