

Towards Fish Welfare in the Presence of Robots: Zebrafish Case
 ,
,  , ,
, ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fish and Accommodation Conditions
2.2. Robots Tested
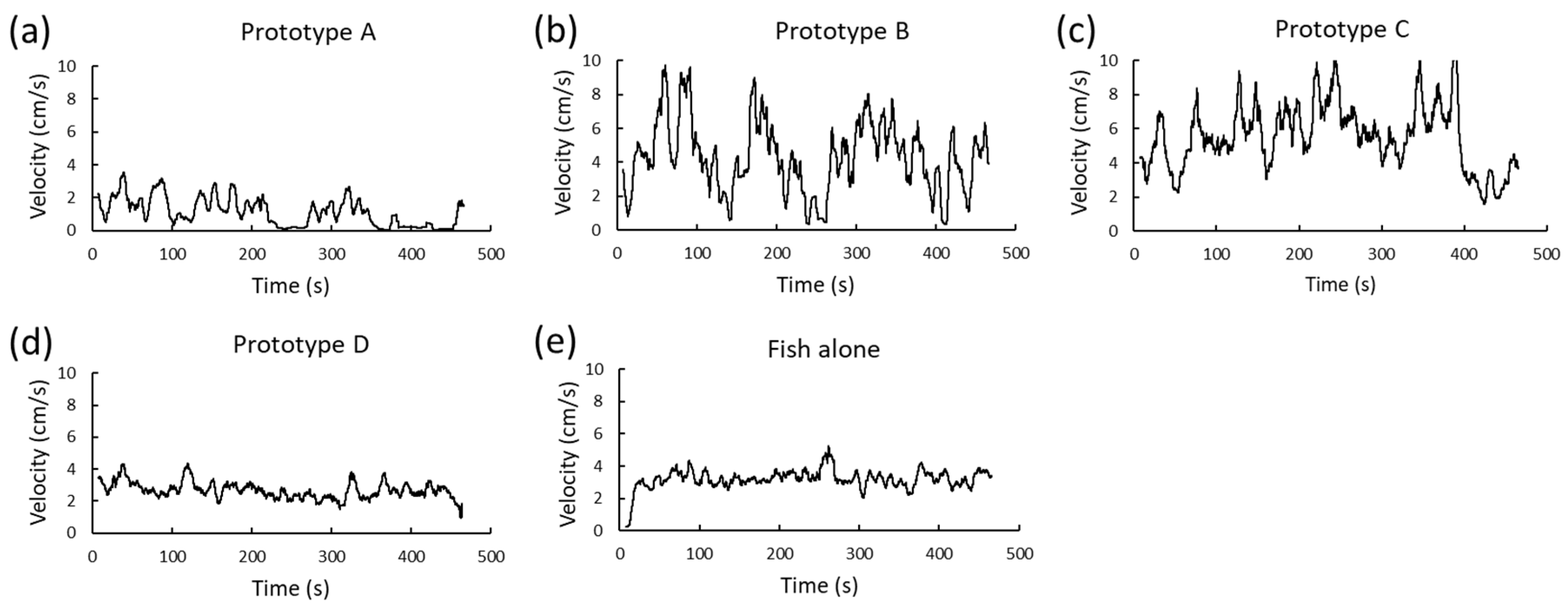
- Prototype A is powered by a 0.3 g electromagnetic actuator (provided by Shuaichi, CN), model DIY RC Aircraft, measuring 10 × 10 × 2 mm, with a resistance of 60 ohms, an operating voltage of 3.7–4.2 V, and an operating current of 55 mAh, which is connected to an acetate tail. This propulsion system generates a tail movement characterized by oscillatory beats, the frequency of which can be adjusted in advance. Consequently, changes in the robot’s speed and direction are achieved. The electronic system of this prototype includes a rechargeable 2.7 V, 30 mAh lithium battery, model 450909, and a mini-PCB (Figure 3a). The dimensions of this prototype are as follows: length—6.5 cm; height—2 cm; and thickness—1.2 cm (Prototype A in Figure 1). Upon contact with water, the circuit is automatically closed activating the prototype. However, it should be noted that Prototype A only swims on the water’s surface and is unable to go deeper than 2 cm below the surface level within the tank environment.
- Prototype B is a replica of the previous model which also includes a red LED light (Prototype B in Figure 1). This red LED is included in the mini-PCB commercially acquired and flashes intermittently at the same frequency as the tail, from within the housing, illuminating the entire body of the prototype. Given that the experiments are intended to be conducted at a maximum depth of 15 cm and a maximum distance of 35 cm, and that the wavelength of the red LED can be seen by both the cameras and individuals at these distances, this LED is used to simplify the composition of the prototypes.
- Prototype C is actuated by a planetary gear motor (provided by Zhaowei, CN), model ZWPD006006 to 420 rpm, with a weight of 1.6 g, a working torque of 40 g·cm, and a stall torque of 90 g·cm. It measures 6 mm in diameter and 21 mm in length and is linked to a 1.2 cm diameter propeller designed and manufactured following the same process as the outer housings. This propulsion system offers continuous rotation resulting in constant speed and advancement exclusively in the frontal direction. Additionally, the electronic system includes a rechargeable 4.2 V lithium battery and a magnetic switch that allows the system to be actuated by an external magnet which (Figure 3b), in turn, serves as a counterweight to achieve neutral buoyancy. The prototype measures 5.5 cm long, 2 cm tall, and 1.2 cm thick (Prototype C in Figure 1). This prototype can submerge due to the thrust generated by the propeller.
- Prototype D is identical to prototypes A and B, yet all electronic components were removed, resulting in a motionless prototype that can only float or remain stationary at the bottom, depending on its buoyancy (Prototype D in Figure 1). This model allows us to study whether the effects generated by the movement, sounds, and waves of the electronic components of the robots are significant and allows us to analyze whether the presence of a foreign object in the tank, its aesthetics, or size are influential in perceiving the prototypes as stressful.
2.3. Behavioral Quantification
2.4. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Huang, H.; Tang, Q.; Li, J.; Zhang, W.; Bao, X.; Zhu, H.; Wang, G. A Review on Underwater Autonomous Environmental Perception and Target Grasp, the Challenge of Robotic Organism Capture. Ocean. Eng. 2020, 195, 106644. [Google Scholar] [CrossRef]
- Ranapurwala, S.I.; Kucera, K.L.; Denoble, P.J. The Healthy Diver: A Cross-Sectional Survey to Evaluate the Health Status of Recreational Scuba Diver Members of Divers Alert Network (DAN). PLoS ONE 2018, 13, e0194380. [Google Scholar] [CrossRef]
- Bannister, J.; Sievers, M.; Bush, F.; Bloecher, N. Biofouling in Marine Aquaculture: A Review of Recent Research and Developments. Biofouling 2019, 35, 631–648. [Google Scholar] [CrossRef]
- Amundsen, H.B.; Caharija, W.; Pettersen, K.Y. Autonomous ROV Inspections of Aquaculture Net Pens Using DVL. IEEE J. Ocean. Eng. 2022, 47, 1–19. [Google Scholar] [CrossRef]
- Thinkinazul. Available online: https://thinkinazul.es/en/ (accessed on 25 May 2024).
- López-Barajas, S.; Sanz, P.J.; Marín-Prades, R.; Gómez-Espinosa, A.; González-García, J.; Echagüe, J. Inspection Operations and Hole Detection in Fish Net Cages through a Hybrid Underwater Intervention System Using Deep Learning Techniques. J. Mar. Sci. Eng. 2024, 12, 80. [Google Scholar] [CrossRef]
- Kruusmaa, M.; Gkliva, R.; Tuhtan, J.A.; Tuvikene, A.; Alfredsen, J.A. Salmon Behavioural Response to Robots in an Aquaculture Sea Cage. R. Soc. Open Sci. 2020, 7, 191220. [Google Scholar] [CrossRef]
- Vidyarthi, P.K.; Mukherjee, K.; Roy, B.K. Fish-Like Robots and Applications of Sensor-A Review. In Proceedings of the 4th International Conference on Energy, Power, and Environment, ICEPE 2022, Shillong, India, 29 April–1 May 2022. [Google Scholar]
- Free, B.A.; Lee, J.; Paley, D.A. Bioinspired Pursuit with a Swimming Robot Using Feedback Control of an Internal Rotor. Bioinspir. Biomim. 2020, 15, 035005. [Google Scholar] [CrossRef]
- Li, L.; Ravi, S.; Xie, G.; Couzin, I.D. Using a Robotic Platform to Study the Influence of Relative Tailbeat Phase on the Energetic Costs of Side-by-Side Swimming in Fish. Proc. R. Soc. A Math. Phys. Eng. Sci. 2021, 477, 20200810. [Google Scholar] [CrossRef]
- Van Den Berg, S.C.; Scharff, R.B.N.; Rusák, Z.; Wu, J. OpenFish: Biomimetic Design of a Soft Robotic Fish for High Speed Locomotion. HardwareX 2022, 12, e00320. [Google Scholar] [CrossRef]
- Katzschmann, R.K.; Delpreto, J.; Maccurdy, R.; Rus, D. Exploration of Underwater Life with an Acoustically Controlled Soft Robotic Fish. Sci. Robot. 2018, 3, eaar3449. [Google Scholar] [CrossRef]
- Jarque, A.P.; Solis, A.; Vidal, R.; Marín, R.; Cerdá-Reverter, J.M.; Sanz, P.J. Towards Precision Aquaculture: “The UJI Micro Robot Fish Prototype”. In Proceedings of the OCEANS 2023-Limerick, OCEANS Limerick 2023, Limerick, Ireland, 5–8 June 2023. [Google Scholar]
- Chiara, V.; Kim, S.Y. AnimalTA: A Highly Flexible and Easy-to-Use Program for Tracking and Analysing Animal Movement in Different Environments. Methods Ecol. Evol. 2023, 14, 1699–1707. [Google Scholar] [CrossRef]
- Maertens, A.P.; Triantafyllou, M.S.; Yue, D.K.P. Efficiency of Fish Propulsion. Bioinspir. Biomim. 2015, 10, 046013. [Google Scholar] [CrossRef]
- Tschirren, L.; Bachmann, D.; Güler, A.C.; Blaser, O.; Rhyner, N.; Seitz, A.; Zbinden, E.; Wahli, T.; Segner, H.; Refardt, D. Myfishcheck: A Model to Assess Fish Welfare in Aquaculture. Animals 2021, 11, 145. [Google Scholar] [CrossRef]
- Blanchard, D.C.; Blanchard, R.J. Chapter 2.4 Defensive Behaviors, Fear, and Anxiety. In Handbook of Anxiety and Fear; Academic Press: Boston, MA, USA; Amsterdam, The Netherlands, 2008; Volume 17, ISBN 0-444-53065-7. [Google Scholar]
- Kopman, V.; Laut, J.; Polverino, G.; Porfiri, M. Closed-Loop Control of Zebrafish Response Using a Bioinspired Robotic-Fish in a Preference Test. J. R. Soc. Interface 2013, 10, 20120540. [Google Scholar] [CrossRef]
- Bierbach, D.; Landgraf, T.; Romanczuk, P.; Lukas, J.; Nguyen, H.; Wolf, M.; Krause, J. Using a Robotic Fish to Investigate Individual Differences in Social Responsiveness in the Guppy. R. Soc. Open Sci. 2018, 5, 181026. [Google Scholar] [CrossRef]
- Kruusmaa, M.; Rieucau, G.; Montoya, J.C.C.; Markna, R.; Handegard, N.O. Collective Responses of a Large Mackerel School Depend on the Size and Speed of a Robotic Fish but Not on Tail Motion. Bioinspir. Biomim. 2016, 11, 056020. [Google Scholar] [CrossRef]
- Phamduy, P.; Polverino, G.; Fuller, R.C.; Porfiri, M. Fish and Robot Dancing Together: Bluefin Killifish Females Respond Differently to the Courtship of a Robot with Varying Color Morphs. Bioinspir. Biomim. 2014, 9, 036021. [Google Scholar] [CrossRef]
- Bierbach, D.; Mönck, H.J.; Lukas, J.; Habedank, M.; Romanczuk, P.; Landgraf, T.; Krause, J. Guppies Prefer to Follow Large (Robot) Leaders Irrespective of Own Size. Front. Bioeng. Biotechnol. 2020, 8, 441. [Google Scholar] [CrossRef]
- Ruchin, A.B. Effect of Illumination on Fish and Amphibian: Development, Growth, Physiological and Biochemical Processes. Rev. Aquac. 2021, 13, 567–600. [Google Scholar] [CrossRef]
- Glaze, J.; Salazar, R.; Vasconcellos, R.; Abdelkefi, A. Comparative Design, Hydrodynamic Analysis, and Physical Performance of Fish-like Robots. Appl. Ocean. Res. 2021, 106, 102443. [Google Scholar] [CrossRef]
- Dadswell, M.J.; Spares, A.D.; Mclean, M.F.; Harris, P.J.; Rulifson, R.A. Long-Term Effect of a Tidal, Hydroelectric Propeller Turbine on the Populations of Three Anadromous Fish Species. J. Fish Biol. 2018, 93, 192–206. [Google Scholar] [CrossRef]
- Maximino, C.; de Brito, T.M.; da Silva Batista, A.W.; Herculano, A.M.; Morato, S.; Gouveia, A. Measuring Anxiety in Zebrafish: A Critical Review. Behav. Brain Res. 2010, 214, 157–171. [Google Scholar] [CrossRef]
- Stewart, A.; Gaikwad, S.; Kyzar, E.; Green, J.; Roth, A.; Kalueff, A.V. Modeling Anxiety Using Adult Zebrafish: A Conceptual Review. Neuropharmacology 2012, 62, 135–143. [Google Scholar] [CrossRef]
- Johnson, A.; Loh, E.; Verbitsky, R.; Slessor, J.; Franczak, B.C.; Schalomon, M.; Hamilton, T.J. Examining Behavioural Test Sensitivity and Locomotor Proxies of Anxiety-like Behaviour in Zebrafish. Sci. Rep. 2023, 13, 3768. [Google Scholar] [CrossRef]
- Phamduy, P. Robotic Fish to Aid Animal Behavior Studies and Informal Science Learning. Ph.D. Thesis, New York University, New York, NY, USA, 2017. [Google Scholar]
- Blackstock, J.M.; Covington, M.D.; Perne, M.; Myre, J.M. Monitoring Atmospheric, Soil, and Dissolved CO2 Using a Low-Cost, Arduino Monitoring Platform (CO2-LAMP): Theory, Fabrication, and Operation. Front. Earth Sci. 2019, 7, 313. [Google Scholar] [CrossRef]
- Kalueff, A.V.; Gebhardt, M.; Stewart, A.M.; Cachat, J.M.; Brimmer, M.; Chawla, J.S.; Craddock, C.; Kyzar, E.J.; Roth, A.; Landsman, S.; et al. Towards a Comprehensive Catalog of Zebrafish Behavior 1.0 and Beyond. Zebrafish 2013, 10, 70–86. [Google Scholar] [CrossRef]
- Romenskyy, M.; Herbert-Read, J.E.; Ward, A.J.W.; Sumpter, D.J.T. Body Size Affects the Strength of Social Interactions and Spatial Organization of a Schooling Fish (Pseudomugil signifer). R. Soc. Open Sci. 2017, 4, 161056. [Google Scholar] [CrossRef]
- Polverino, G.; Porfiri, M. Mosquitofish (Gambusia affinis) Responds Differentially to a Robotic Fish of Varying Swimming Depth and Aspect Ratio. Behav. Brain Res. 2013, 250, 133–138. [Google Scholar] [CrossRef]
- Landgraf, T.; Bierbach, D.; Nguyen, H.; Muggelberg, N.; Romanczuk, P.; Krause, J. RoboFish: Increased Acceptance of Interactive Robotic Fish with Realistic Eyes and Natural Motion Patterns by Live Trinidadian Guppies. Bioinspir. Biomim. 2016, 11, 15001. [Google Scholar] [CrossRef]
- Rocha, A.; Godino-Gimeno, A.; Rotllant, J.; Cerdá-Reverter, J.M. Agouti-Signalling Protein Overexpression Reduces Aggressiveness in Zebrafish. Biology 2023, 12, 712. [Google Scholar] [CrossRef]
- Spinello, C.; Yang, Y.; Macrì, S.; Porfiri, M. Zebrafish Adjust Their Behavior in Response to an Interactive Robotic Predator. Front. Robot. AI 2019, 1, 38. [Google Scholar] [CrossRef]
- Polverino, G.; Phamduy, P.; Porfiri, M. Fish and Robots Swimming Together in a Water Tunnel: Robot Color and Tail-Beat Frequency Influence Fish Behavior. PLoS ONE 2013, 8, e77589. [Google Scholar] [CrossRef]
- Butail, S.; Polverino, G.; Phamduy, P.; Del Sette, F.; Porfiri, M. Influence of Robotic Shoal Size, Configuration, and Activity on Zebrafish Behavior in a Free-Swimming Environment. Behav. Brain Res. 2014, 275, 269–280. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Analytical Model | Evaluation Measures for Determining the Presence of Stress |
|---|---|
| Conditioned Alarm Reaction | Time in Bottom Zone |
| Total Path Length | |
| Freeze Time | |
| Fast Swimming | |
| Inhibitory Avoidance | Time Until Visiting Aversive Zone |
| Time in Aversive Zone | |
| Predator Response | Burst Swimming |
| Freeze | |
| Bottom Dwell Time | |
| Distance to Predator | |
| Inspection of Novel Objects | Distance Between Fish and Object |
| Time Near the Object |
| 95% Confidence Interval for Mean | |||||||
|---|---|---|---|---|---|---|---|
| Parameter | Model | Mean | Std. Deviation | Lower Bound | Upper Bound | Normality Sig. | Homogeneity of Variances Sig. * |
| Top time proportion | A | 30.51 | 18.37 | 13.52 | 47.51 | ||
| B | 56.20 | 20.55 | 37.20 | 75.21 | |||
| C | 28.92 | 30.08 | 3.77 | 54.06 | 0.005 | 0.23 | |
| D | 22.67 | 29.93 | −5.01 | 50.35 | |||
| E | 15.13 | 10.65 | 5.28 | 24.98 | |||
| Nº of visits to surface | A | 2.81 | 2.21 | 0.77 | 4.86 | ||
| B | 7.25 | 2.80 | 4.66 | 9.84 | |||
| C | 7.94 | 8.45 | 0.88 | 15.00 | 0.002 | 0.440 | |
| D | 2.48 | 2.70 | −0.02 | 4.98 | |||
| E | 5.33 | 4.43 | 1.23 | 9.43 | |||
| Perimeter time proportion | A | 12.83 | 8.43 | 5.04 | 20.63 | ||
| B | 24.48 | 11.70 | 13.66 | 35.30 | |||
| C | 43.29 | 22.38 | 24.58 | 62.01 | 0.111 | 0.281 | |
| D | 23.22 | 15.39 | 8.98 | 37.45 | |||
| E | 31.40 | 10.92 | 21.29 | 41.50 | |||
| Freezing time proportion | A | 21.70 | 18.84 | 4.28 | 39.13 | ||
| B | 6.88 | 14.45 | −6.49 | 20.25 | |||
| C | 10.96 | 15.70 | −2.16 | 24.07 | <0.001 | 0.373 | |
| D | 12.52 | 28.67 | −13.99 | 39.04 | |||
| E | 7.08 | 9.30 | −1.53 | 15.69 | |||
| Tracking average distance | A | 275.34 | 160.96 | 126.48 | 424.2 | ||
| B | 418.24 | 140.34 | 288.45 | 548.03 | |||
| C | 470.95 | 255.39 | 257.44 | 684.47 | 0.200 | 0.140 | |
| D | 302.38 | 127.81 | 184.17 | 420.58 | |||
| E | 401.09 | 61.83 | 343.91 | 458.27 | |||
| Velocity absolute deviation | A | 3.39 | 1.62 | 1.90 | 4.88 | ||
| B | 4.15 | 0.73 | 3.48 | 4.82 | |||
| C | 4.69 | 2.84 | 2.32 | 7.06 | 0.019 | 0.005 | |
| D | 3.19 | 0.73 | 2.52 | 3.87 | |||
| E | 2.23 | 0.68 | 1.60 | 2.86 | |||
| Acceleration absolute deviation | A | 100.41 | 69.72 | 35.93 | 164.90 | ||
| B | 119.52 | 49.13 | 74.08 | 164.96 | |||
| C | 69.46 | 26.26 | 47.51 | 91.41 | 0.056 | 0.020 | |
| D | 61.31 | 34.35 | 29.54 | 93.08 | |||
| E | 40.99 | 12.26 | 29.65 | 52.33 | |||
| Velocity Average | A | 2.97 | 1.74 | 1.36 | 4.57 | ||
| B | 3.2 | 1.55 | 1.76 | 4.63 | |||
| C | 4.97 | 2.9 | 2.55 | 7.39 | 0.200 | 0.047 | |
| D | 3.55 | 1.84 | 1.85 | 5.26 | |||
| E | 4.42 | 0.93 | 3.56 | 5.28 | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pino, A.; Vidal, R.; Tormos, E.; Cerdà-Reverter, J.M.; Marín Prades, R.; Sanz, P.J. Towards Fish Welfare in the Presence of Robots: Zebrafish Case. J. Mar. Sci. Eng. 2024, 12, 932. https://doi.org/10.3390/jmse12060932
Pino A, Vidal R, Tormos E, Cerdà-Reverter JM, Marín Prades R, Sanz PJ. Towards Fish Welfare in the Presence of Robots: Zebrafish Case. Journal of Marine Science and Engineering. 2024; 12(6):932. https://doi.org/10.3390/jmse12060932
Chicago/Turabian StylePino, Andrea, Rosario Vidal, Elisabeth Tormos, José Miguel Cerdà-Reverter, Raúl Marín Prades, and Pedro J. Sanz. 2024. "Towards Fish Welfare in the Presence of Robots: Zebrafish Case" Journal of Marine Science and Engineering 12, no. 6: 932. https://doi.org/10.3390/jmse12060932