Abstract
Idunella spinifera (Dauvin and Gentil, 1983) (Crustacea: Amphipoda: Liljeborgiidae) is reported for the first time after the original description that was based on one immature female. Specimens were collected in the Ría de Muros (NW Iberian Peninsula) during the course of a study on macrofauna diversity in shallow sublittoral biogenic sands along a one-year period. The male and the ovigerous female are fully described, and the data on ecology and temporal variation of abundance is provided as well.
1. Introduction
The Order Amphipoda is the largest group within the superorder Peracarida (Crustacea: Malacostraca). Amphipods can inhabit terrestrial habitats, but they are mainly found on aquatic environments, on fully marine conditions (from intertidal to deep sea), transitional systems (e.g., estuaries and lagoons), and fresh water (e.g., rivers, pounds, and wells) [1]. However, 79% of the known amphipod species are found in marine environments, including soft bottoms, rocky shores, phytal, and pelagic habitats [2]. In marine habitats, amphipods can be one of the most important components of the macrobenthic assemblages, both in terms of abundance and diversity [3,4]. Moreover, amphipods play a key role in food webs by processing primary production and detritus, and establish a link in the transfer of energy to higher trophic levels [2]. Thus, amphipods are globally crucial in marine ecosystems by supporting and linking trophic levels [2,5]. Their ecological relevance is explained by the great functional diversity as they embrace different life styles (e.g., burrowing, bentho–pelagic, epibenthic, and pelagic) [6,7] and multiple feeding strategies (detritivores, filter-feeders, grazers, predators, and scavengers) [8,9]. In this way, amphipods can act as benthic–pelagic coupling facilitators [2]. Moreover, many amphipods (burrowers or tube-builders) are bioturbators that have a relevant role in structuring benthic assemblages [10,11]. Considering the great abundance and diversity of amphipods in benthic and pelagic communities, they also play an important role in disturbance events [12]. However, there are still many knowledge gaps in multiple issues of amphipod biology [13,14] and taxonomy; it is estimated that only one third of extant amphipod species have been described until now [15]. Thus, nowadays, their biological and ecological role is underestimated and understudied [12,15].
The cosmopolitan family Liljeborgiidae Stebbing, 1899, currently includes three valid genera and about 120 species [16,17]. During the first decade of this century, the collection of new material from different parts of Europe was allowed to review and clarify the identity of many species and genera of this family [16]. Among European species, Idunella spinifera (Dauvin and Gentil, 1983) had not been reported after the original description [18]. The only known specimen corresponds to an immature female collected in biogenic coarse sands at 15 m depth in Roscoff (France) [18]. Therefore, the description lacks information about ovigerous females and adult males, and whether the latter are similar to those of similar European species [16].
In the framework of a study aimed to characterise the benthic fauna in biogenic sands at the Ría de Muros (Galicia, NW Spain), several individuals of I. spinifera were collected; those included males and fully mature ovigerous females. Therefore, this work provides a detailed description of this new material to complement the original description as well as ecological information about habitat features and temporal variation in abundance of I. spinifera.
2. Materials and Methods
2.1. Study Area
Sampling was conducted in Ría de Muros, an extensive coastal embayment in Galicia (NW coast of Iberian Peninsula), characterised as a partially mixed estuary with a positive circulation according to thermohaline circulation and dynamics [19]; fresh water influence is mostly limited to the inner part. Ría de Muros is under the wind-driven coastal upwelling of the NW Africa upwelling system [20], thus showing high primary productivity. It also shows a variety of sedimentary habitats with a high benthos diversity [21].
2.2. Sampling and Sample Processing
Samples were collected from one site on the north margin of the Ría de Muros (42°46′16.23″ N; 8° 58′40.61″ W) at 16 m depth. The site was visited in March, June, September, and December 2016. At each date, thirteen sediment samples were collected using a Van Veen grab (sampling surface 0.05 m2). An additional grab was deployed to collect sediment destined to granulometry analysis and to measure environmental parameters. Samples that were destined to study macrobenthos were immediately washed on board over a 1 mm mesh sieve. The retained macrofauna was then preserved in 4% neutralised formaldehyde solution with Rose Bengal in labelled plastic bags until posterior sorting in the laboratory under a stereomicroscope. Samples used for sedimentary study were frozen. At each date, the surface water temperature, salinity, and redox potential (Eh) of sediment were measured in situ.
Structural features of the sediment (e.g., median grain size, sorting (QDΦ), and silt and clay content) were determined for each sample. The sediment grain size was analysed by using a mechanical shaker and then dry sieving through a tower of sieves (from 4 to 0.063 mm mesh) and classified according to the Wentworth scale. Total carbon (TC), inorganic carbon (IC), and total organic carbon (TOC) were measured after a previously dried and powdered portion of the sediment sample by means of a LECO CNS-2000 elemental macro analyser.
A total of 40 specimens of I. spinifera were examined, corresponding to 5 mature males, 6 ovigerous females, and 29 non-ovigerous females. Several males and ovigerous females were dissected, and their appendages were mounted on slides and examined under the microscope. Line drawings were conducted by means of a camera lucida connected to a compound microscope. Selected specimens are deposited at the Museo Nacional de Ciencias Naturales (MNCN, Madrid), and the remaining material is kept in the collection of the authors.
3. Results
3.1. Environmental Variables
Water temperature ranged from 12 °C in March to 16.2 °C in September. Salinity also showed some temporal variability with values that ranged from 31 psu in September to 34 psu in June. Sediment Eh values were always positive, and ranged from 340 mV in December to 10 mV in June. Sediment type was stable across all sampling periods, and corresponded to coarse sand. Sorting was always moderately sorted. The amount of TC ranged from 0.21 mg g−1 in September to 0.12 mg g−1 in March. Values of IC ranged from 0.12 mg g−1 in June to 0.08 mg g−1 in March. The amount of TOC ranged from 0.08 mg g−1 in June to 0.03 mg g−1 in March.
3.2. Abundance and Sizes of I. spinifera
The abundance of I. spinifera was variable in time, with low values in March that increased to achieve the maximum value of abundance in September to decrease again in December (Figure 1A). Females were present at each of the studied dates. However, males and ovigerous females were only found in June and September (Figure 1B).
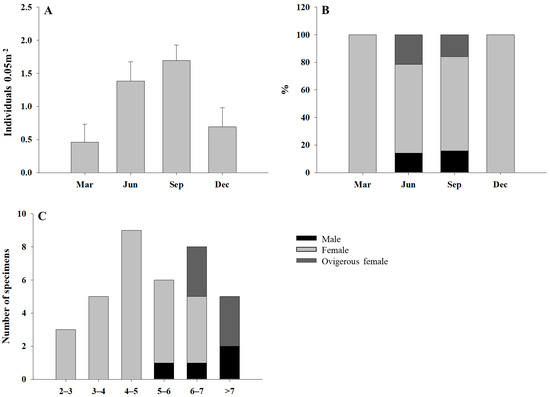
Figure 1.
Mean abundance (+SE; n = 13) of I. spinifera (A) per month; % of males, females, and ovigerous females at each month (B); size distribution (total body length, mm) of males, females, and ovigerous females (C).
Specimens longer than 4 mm (total body length, BL) comprised 77.7% of the total (n = 36 complete specimens) (Figure 1C). However, smaller and juvenile specimens were likely to be washed away through the 1 mm sieve used in the field, and therefore their presence might be underestimated. Larger specimens (>6 mm BL) mostly corresponded to ovigerous females and males.
3.3. Taxonomy
Family Liljeborgiidae Stebbing, 1899.
Subfamily Idunellinae d’Udekem d’Acoz, 2010.
Genus Idunella G.O. Sars, 1894.
Idunella spinifera (Dauvin and Gentil, 1983).
Listriella spinifera Dauvin and Gentil, 1983. Cah. Biol. Mar., 24: 436–440, Figures 3 and 4.
3.4. Description of Male
Five males examined (one broken at pleosome-urosome level), 6.0–7.1 mm BL (n = 4).
Head (Figure 2 and Figure 3A): 1.8 times as long as pereonite 1 and 8% of body length. Eyes well developed, ommatidia covering most of eye, strong dark pigmentation; rostrum short; ventral margin produced anteriorly and rounded.
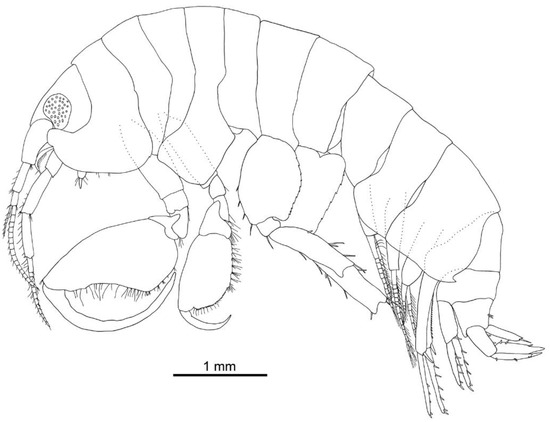
Figure 2.
Idunella spinifera (Dauvin and Gentil, 1983). Male, Ría de Muros (NW Spain), lateral view.
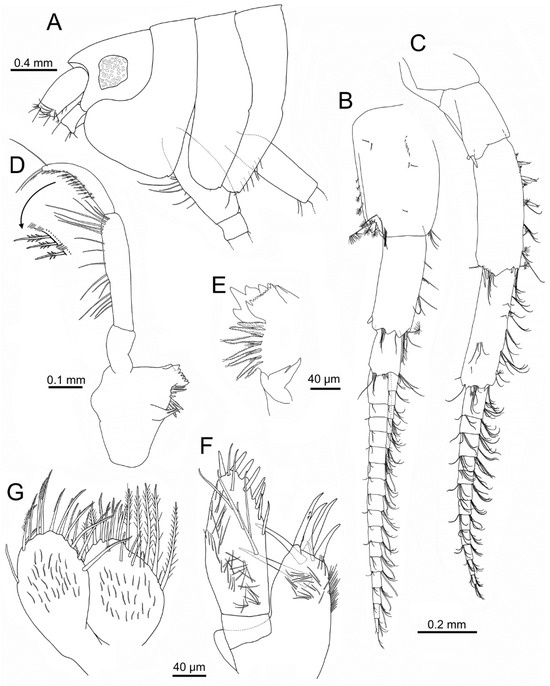
Figure 3.
Idunella spinifera (Dauvin and Gentil, 1983). Male, Ría de Muros (NW Spain). (A) head and anterior thoracic segments, lateral view. (B) antenna 1. (C) antenna 2. (D) left mandible (palp upturned) and detail of rows of D3 (inferior) and B3-setae (superior; only some clusters of setae illustrated). (E) right mandible, distomedial part. (F) right maxilla 1. (G) left maxilla 2. (B,C,F,G) same scale.
Antenna 1 (Figure 3B): peduncle article 1 1.6 times as long as wide, with several simple short setae on external surface, plumose setae on distal half of posterior margin, cluster of setae on anterodistal margin; article 2 0.8 times as long as article 1, with one spine on posterodistal margin and clusters of setae on anterior and distal margins, one plumose seta on anterodistal corner; article 3 0.46 times as long as article 2, with setae on distal margin. Flagellum with twelve to seventeen articles, each provided with two clusters of setae and one aesthetasc on anterodistal margin; accessory flagellum with five to six articles, with several setae mostly on distal margin or minute distal article; ratio accessory flagellum/flagellum: 0.3; ratio flagellum/peduncle article 1: 2.0.
Antenna 2 (Figure 3C): subequal in length to antenna 1. Peduncle article 3 with one to two posterodistal spines; article 4 about three times as long as wide, with a row of five spines on anterior margin each associated with simple and plumose setae; two spines on anterodistal margin, and three spines on posterodistal margin, plus several associated setae; article 5 3.5 times as long as wide, with several clusters of setae on anterior margin and external face on distal third, one spine on anterodistal corner, and several terminal plumose and simple setae. Flagellum about 0.6 times as long as peduncle, with ten to twelve articles, each with two clusters of setae on distal margin.
Mandible (Figure 3D,E): distolateral corner not forming a definite tooth; left lacinia mobilis with six triangular teeth; right lacinia mobilis with anterior margin with sharp small teeth (>10) and two larger acute teeth; incisor process with five triangular teeth, one much longer than others, six to seven raker robust setae with many setules, with several shorter setae in between; molar process distinct, with two distally acute setae. Left and right mandibular palp similar; palp article 1 about 1.6 times as long as wide and 0.4 times as long as article 2, lacking setae; article 2 4.3 times as long as wide, with a row of medial and subdistal setae; article 3 curved, widest at mid-length, 0.7 times as long as article 2, with posterior row of D3-setae not increasing in length and with a parallel row of short, very thin B3-setae grouped in clusters of four or five, one long subdistal B3-seta, and two to three E3-setae.
Maxilla 1 (Figure 3F): palp article 1 about 0.25 times as long as article 2; article 2 with eight stout setae of increasing thickness on mesial and apical margins, facial setae including six long setae and numerous thinner and shorter setae. Outer plate with seven distal simple and serrate stout setae, one much shorter than others, and numerous shorter setae on surface of distal half.
Maxilla 2 (Figure 3G): inner plate slightly wider than outer plate; both plates with long setae on distal part, most plumose; inner plate with several facial setae near anterior margin, and numerous shorter setae along surfaces of both plates.
Maxilliped (Figure 4A–C): palp length ratio of article 1–4 about 1:3:1.8:1.7; article 1 with one seta; article 2 with long medial setae, and cluster of setae on distolateral corner; article 3 with medial setae on distal half and cluster of setae on distolateral corner; article four slender, slightly curved, unguis about 0.25 times as long as dactylus, with two basal setae. Outer plate reaching proximal third of palp article 2, with five to seven lateral setae and five distal spines of growing thickness. Inner plate small, with four simple setae, one spine, and two plumose setae on margin of distal half, posterodistal margin with one to two longer setae.
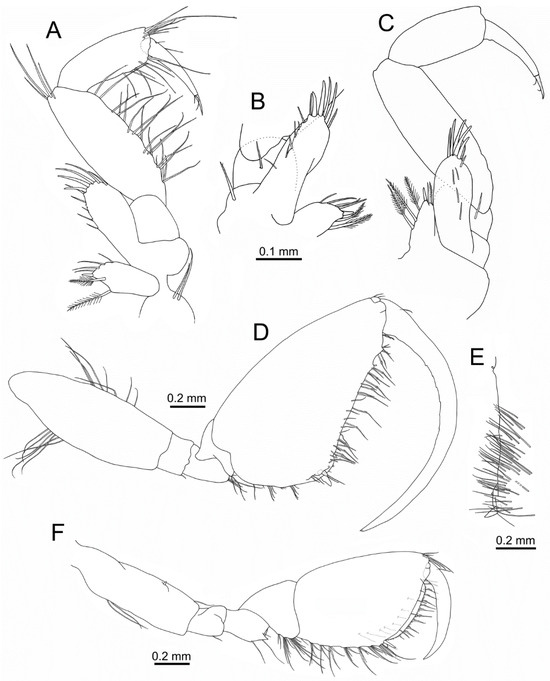
Figure 4.
Idunella spinifera (Dauvin and Gentil, 1983). Male, Ría de Muros (NW Spain). (A) right maxilliped (palp upturned). (B) right maxilliped, proximal part (opposite face). (C) left maxilliped (palp upturned; opposite face). (D) gnathopod 1. (E) gnathopod 1 propodus, palm inner face. (F) gnathopod 2. (A–C) same scale. (C) palp setae not illustrated.
Gnathopod 1 (Figure 3A and Figure 4D,E): larger than gnathopod 2. Coxa broad, 1.2 times as long as wide, with a posterodistal tooth, anterior margin rounded. Length ratio of articles from basis to dactylus about 5.4:1:1.5:1.5:6.6:7.2. Basis with row of setae on anterior (second third) and posterior margin (proximal third). Ischium with one small posterodistal seta. Merus with a posterodistal cluster of setae, lacking distal tooth. Carpus with a posterodistal cluster of setae. Propodus roughly triangular in shape, 1.7 times as long as wide, length ratio of anterior, posterior and palmar margins about 2.25:1:1.6; with four clusters of setae along posterior margin; palm defined by two spines of different length associated with several setae, palm margin almost straight with two processes on distal third near to dactylus, one lateral row of simple, shorter setae and medial row of longer setae with several spinules, posterodistal palmar margin with one simple seta and one pappose seta, inner face of palm proximal half with several rows of simple setae; two anterodistal small setae near dactylus. Dactylus strongly curved, dactylar teeth as small indentations on second quarter; anterior margin with one proximal seta.
Gnathopod 2 (Figure 3A and Figure 4F): coxa 1.6 times as long as wide, anterior margin slightly convex and posterior margin nearly straight, with a posterodistal small tooth and one seta. Length ratio of articles from basis to dactylus about 3.6:1:1.2:1.5:3.6:2.5. Basis with two setae on posterior margin, anterior margin with one acute process on distal third. Ischium with one posterior subdistal seta. Merus posterodistal corner with conspicuous triangular tooth and several setae of different length. Carpus with three clusters of setae along posterior margin. Propodus with anterior and posterior border nearly straight, 1.4 times as long as wide, length ratio of anterior, posterior and palmar margins about 1.4:1:0.9; posterior margin border crested, with seven clusters of setae on posterior margin, each cluster arising from one crest; palm defined by two spines of different length, palm border smooth, with four lateral setae widely spaced, two medial rows of about twenty-five (outer row) and ten to twelve (inner) hooked stout setae regularly spaced and medial row of setae with spinules, inner face with several setae; six anterodistal setae near dactylus. Dactylus lacking teeth; anterior margin with one proximal seta.
Pereopod 3 (Figure 5A): coxa nearly rectangular, 2.3 times as long as wide, with several setae on posterior margin and small tooth with one seta on posterodistal corner. Length ratio of articles from basis to dactylus about 4.7:1:2.4:1.7:2.7:1.4. Basis with several setae on distal third of anterior margin. Merus and carpus with several anterodistal and posterodistal setae, carpus also with two setae on posterior margin. Propodus with one anterodistal seta and six setae on posterior margin. Dactylus slender, slightly curved along distal half, without setae.
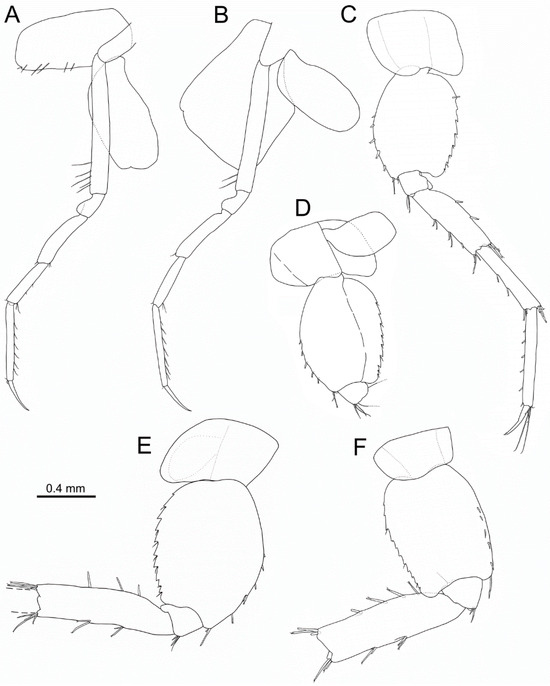
Figure 5.
Idunella spinifera (Dauvin and Gentil, 1983). Male, Ría de Muros (NW Spain). Pereopods and coxae. (A) right pereopod 3, inner face. (B) right pereopod 4, inner face. (C) left pereopod 5, outer face. (D) right pereopod 5, proximal part, inner face. (E) right pereopod 6, outer face. (F) right pereopod 7, outer face. (A–F) same scale bar. (E,F) broken at the carpus-merus level.
Pereopod 4 (Figure 5B): coxa broad, 1.25 times as long as wide, anterior and posterior margins parallel, with notch on anterodistal corner. Length ratio of articles from basis to dactylus about 4.7:1:2.3:2.1:3.1:1.7. Basis with two setae on distal third of anterior margin. Merus with one anterodistal seta. Carpus with one anterodistal seta and two posterodistal setae. Propodus with one anterodistal seta and several setae along posterior margin. Dactylus slender, slightly curved along distal half, without setae.
Pereopod 5 (Figure 5C,D): coxa 0.7 times as long as wide; anterior and posterior margins convex to slightly convex, lacking tooth. Length ratio of articles from basis to dactylus about 3.6:1:2.9:3:3.3:1.4. Basis broad, 1.35 times as long as wide, anterior margin strongly convex, posterior margin regularly convex, anterior margin with seven conical stout spines along distal half and three distal setae, posterior margin with seven to eight serrations along ¾ of distal half, each serration usually with one seta, distal corner rounded and produced into a lobe. Ischium with three anterodistal setae and one small spine. Merus 3.2 times as long as broad, with two to three pairs of setae on anterior margin, two spines on posterior margin on second third, three anterodistal setae, posterodistal corner with two setae and one spine about 0.25 times as long as merus. Carpus anterior margin with two setae on second third, anterodistal and posterodistal setae similar to merus. Propodus with 6 setae on anterior margin, propodal apical tuft with five anterodistal setae (two clearly longer than dactylus). Dactylus slender, slightly curved along distal half, with subterminal short seta.
Pereopod 6 (Figure 5E): broken in all specimens at carpus-merus level. Coxa 0.65 times as long as wide; anterior margin almost straight, posterior border convex, lacking tooth. Length ratio of articles from basis to merus about 4.9:1:4.5. Basis broad, 1.35 times as long as wide, anterior and posterior margins slightly and regularly convex, anterior margin with five spines along distal half and two distal setae, ten serrations all along posterior margin, each serration usually with one seta, distal corner rounded and produced into a small lobe. Ischium with three anterodistal setae, without spine. Merus 3.4 times as long as broad, with two pairs of setae on anterior margin, three spines on posterior margin on proximal ⅔, four anterodistal, and four posterodistal setae.
Pereopod 7 (Figure 5F): broken in all specimens at carpus-merus level. Coxa 0.6 times as long as wide; anterior margin almost straight, posterior margin slightly convex, lacking tooth. Length ratio of articles from basis to merus about 4.3:1:4.7. Basis broad, 1.5 times as long as wide, anterior and posterior margins almost parallel, anterior margin with five spines and one distal longer seta, posterior margin with 11 serrations, some accompanied by one small seta. Ischium with two anterodistal setae, without spine. Merus 3.4 times as long as broad, with two pairs of setae on anterior margin, four spines on posterior margin on proximal ⅔, three anterodistal, and three posterodistal setae.
Pleonites (Figure 6A): all pleonites lacking posterodistal teeth. Pleonite 1 posterior margin weakly convex, posteroventral corner slightly acute. Pleonite 2 posterior margin slightly convex, posteroventral corner with small tooth. Pleonite 3 posterior margin clearly convex on inferior half, posteroventral corner produced into a well-developed tooth.
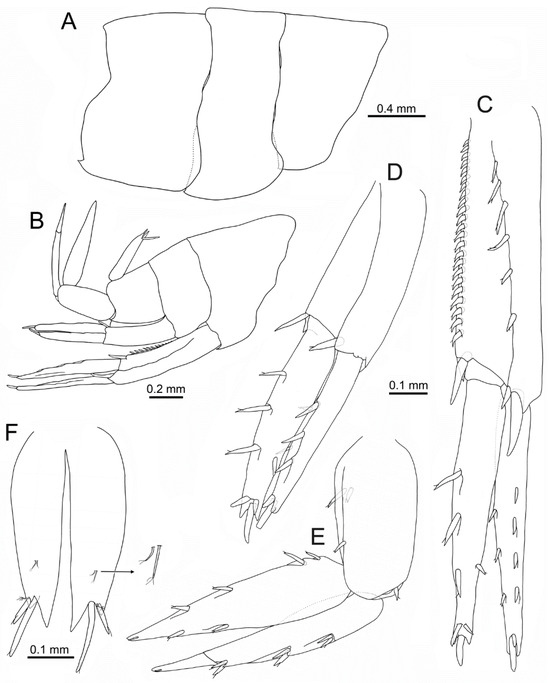
Figure 6.
Idunella spinifera (Dauvin and Gentil, 1983). Male, Ría de Muros (NW Spain). (A) pleonites, right side, lateral view. (B) urosomites, uropods, and telson, right side, lateral view. (C) right uropod 1, laterodorsal view. (D) right uropod 2, laterodorsal view. (E) right uropod 3, lateral view. (F) telson, dorsal view. (C–E) same scale. (B) only some setae illustrated.
Urosomites (Figure 6B): all lacking posterodorsal tooth; urosomite 1 with posteroventral acute tooth.
Uropod 1 (Figure 6C): peduncle 1.2 times as long as rami, five dorsolateral spines and a pair of distal spines consisting of a small one and a long spine pointing backwards, dorsomedial border with a row of sixteen to twenty-three spines and a pair distal of spines similar to dorsolateral ones; outer ramus with four outer and three medial spines, two spines on tip (one much longer); inner ramus with two outer spines and four medial longer and thicker spines, tip with large spine.
Uropod 2 (Figure 6D): peduncle subequal to rami, one distal dorsolateral spine and one distal dorsomedial spine; outer ramus with three outer and two medial spines, two spines on tip (one much longer); inner ramus with one outer and three medial spines, tip with three spines (central much longer).
Uropod 3 (Figure 6E): peduncle 0.7 times as long as rami, one dorsolateral and two dorsomedial spines, one anterodistal spine; outer and inner ramus subequal, outer ramus 2-articulated, article 2 0.4 times as long as article 1, article 1 with three pairs of spines on outer side; inner ramus with two outer and four medial spines on medial border, and one subdistal seta.
Telson (Figure 6B,F): medial cleft 0.9 times as long as telson total length; medial tooth of each lobe about twice as long as outer tooth; each lobe with two spines, medial the longest and about 2–2.2 times as long as outer spine, each pair accompanied by plumose seta; pair of short seta with two to threeapical setules on each lobe dorsal face at distal third.
3.5. Description of Ovigerous Female
Six ovigerous females examined, 6.75–8.75 mm BL (n = 6).
Head: 1.85 times as long as pereonite 1, and 8.5% of body length. Eyes well developed, similar to male.
Antenna 1 (Figure 7A): peduncle article 1 1.7 times as long as wide; article 2 0.65 times as long as article 1; article 3 0.5 times as long as article 2. Flagellum with 13–18 articles; accessory flagellum with 5–7 articles, minute distal article; ratio accessory flagellum/flagellum: 0.3; ratio flagellum/peduncle article 1: 2.0.
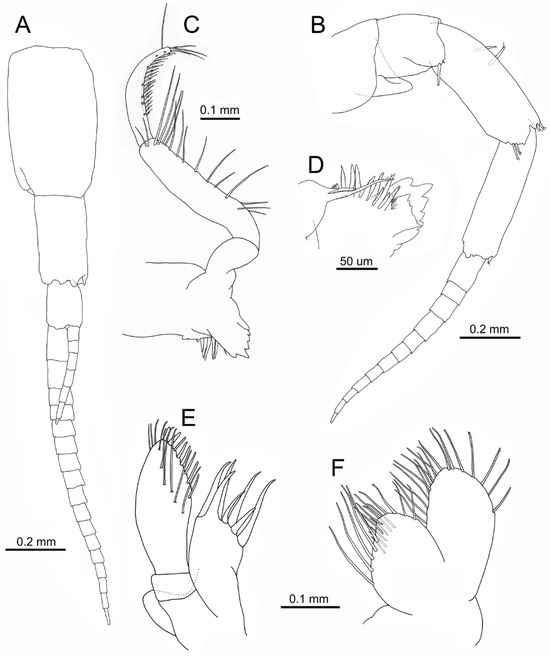
Figure 7.
Idunella spinifera (Dauvin and Gentil, 1983). Ovigerous female, Ría de Muros (NW Spain). (A) antenna 1. (B) antenna 2. (C) right mandible. (D) right mandible, distomedial part, inner face. (E) right maxilla 1. (F) right maxilla 2. (E,F) same scale. (A,B,E,F) only some setae illustrated.
Antenna 2 (Figure 7B): subequal in length to antenna 1. Peduncle article 3 with one long spine; article 4 2.8 times as long as wide, with two spines on inner face near anterior margin proximal third, two spines on the anterodistal margin, and two spines on the posterodistal margin; article 5 3.6 times as long as wide, one spine on anterodistal corner. Flagellum about 0.6 times as long as peduncle, with 9–12 articles.
Mandible (Figure 7C,D): distolateral corner not forming a definite tooth; mandible (laciniae, processes) similar to male. Similar left and right mandibular palp; palp article 1 about 1.6 times as long as wide and 0.35 times as long as article 2, lacking setae; article 2 4.35 times as long as wide; article 3 strongly curved at distal half, widest at mid-length, 0.7 times as long as article 2.
Maxilla 1 (Figure 7E): palp article 1 about 0.23 times as long as article 2. Outer plate similar to male.
Maxilla 2 (Figure 7F): similar to male.
Maxilliped (Figure 8A,B): palp length ratio of article 1–4 about 1:2.5:1.1:1.2; article 1 with two setae; article 3 setae similar to male, also including three facial setae; article 4 slender, slightly curved, unguis 0.25 times as long as dactylus, with one basal seta. Outer plate reaching proximal third of palp article 2, five distal spines of growing thickness. Inner plate small, including three plumose setae, posterodistal margin with four to five setae.
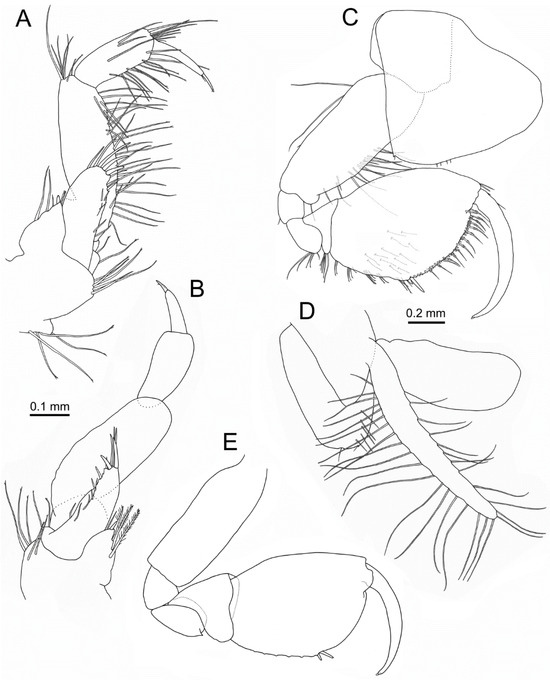
Figure 8.
Idunella spinifera (Dauvin and Gentil, 1983). Ovigerous female, Ría de Muros (NW Spain). (A) left maxilliped. (B) right maxilliped. (C) gnathopod 1. (D) gnathopod 2 coxa, gill, and oostegite, inner face. (E) gnathopod 2. (B) palp setae, not illustrated. (E) only some setae illustrated. (A,B,C,E) same scale.
Gnathopod 1 (Figure 8C): different to male gnathopod 1. Coxa broad, about as long as wide, with a posterodistal tooth (sometimes indistinct) and one seta, anterior margin rounded. Length ratio of articles from basis to dactylus about 4.3:1:1.2:1.2:3.6:3.4. Basis with row of setae on second third anterior margin, and two setae on proximal third posterior margin. Ischium with one small posterodistal seta. Merus with a posterodistal cluster of setae, lacking distal tooth. Carpus with a posterodistal cluster of setae. Propodus oval in shape, 1.6 times as long as wide; length ratio of anterior, posterior, and palmar margins about 2:1:1.15, with five clusters of setae along posterior margin; palm defined by two spines of different length associated with several setae, palm margin with two medial rows of hooked stout setae (>30), and medial row of setae with several spinules; three anterodistal simple setae near dactylus; inner face with several setae. Dactylus gently curved along its length, lacking teeth; anterior margin with one proximal seta.
Gnathopod 2 (Figure 8D,E): similar to gnathopod 1. Coxa oval, wider at base, 1.5 times as long as wide, margins nearly straight, with a posterodistal small tooth and anteroventral notch each with one seta, several setae on inner face near posterodistolateral margin; oostegite well developed 1.5 times as long as coxa, with long setae on anterior and posterior margins. Length ratio of articles from basis to dactylus about 2.9:1:1.1:1.25:2.75:2.3. Merus posterodistal corner with conspicuous triangular tooth. Propodus posterior margin border with small crests, with eight clusters of setae. Palm defined by two spines of different length, palm border smooth, setae and spines similar to gnathopod 1. Dactylus gently curved along length, lacking teeth.
Pereopod 3 (Figure 9A): coxa nearly rectangular, 2.1 times as long as wide, with a posterodistal small tooth and anteroventral notch each with one seta, several setae on inner face near posterodistolateral margin; oostegite well developed, 1.4 times as long as coxa, with long setae on anterior and posterior margins. Length ratio of articles from basis to dactylus about 4.6:1:2:1.6:3:1.5. Dactylus slender, almost straight, without setae.
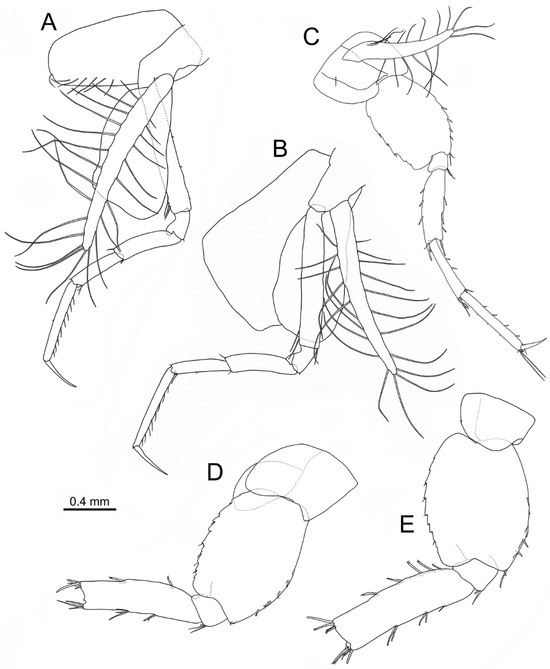
Figure 9.
Idunella spinifera (Dauvin and Gentil, 1983). Ovigerous female, Ría de Muros (NW Spain). Pereopods and coxae. (A) right pereopod 3, inner face. (B) right pereopod 4, inner face. (C) left pereopod 5, inner face. (D) right pereopod 6, outer face. (E) right pereopod 7, outer face. (A–E) same scale bar. (D,E) broken at the carpus-merus level.
Pereopod 4 (Figure 9B): coxa broad, 1.3 times as long as wide, anterior and posterior margins almost straight; oostegite well developed 1.25 times as long as coxa, with long setae on anterior and posterior margins. Length ratio of articles from basis to dactylus about 4.5:1:2.25:1.75:2.75:1.5. Basis with two setae on distal third of anterior margin and one posterodistal seta. Dactylus slender, almost straight, without setae.
Pereopod 5 (Figure 9C): coxa 0.8 times as long as wide; anterior and posterior margins convex to slightly convex, lacking tooth; oostegite well developed, 1.8 times as long as coxa, with long setae on anterior and posterior margins. Length ratio of articles from basis to dactylus about 5:1:3.75:3.1:4.1:1.5. Basis broad, 1.5 times as long as wide, anterior margin convex, posterior margin slightly convex, anterior margin with four conical stout spines along distal half and two distal setae, posterior margin with five to six serrations all along distal half, each serration usually with one seta, distal corner rounded and produced into a small lobe. Merus 3.2 times as long as broad, with two to three pairs of setae on anterior margin, three spines on posterior margin, two anterodistal setae, posterodistal corner with two setae and one spine about 0.25 times as long as merus. Propodus with four setae on anterior margin, propodal apical tuft with five anterodistal setae (two clearly longer than dactylus). Dactylus slender, slightly curved along distal half.
Pereopod 6 (Figure 9D): broken in all specimens at carpus-merus level. Coxa 0.95 times as long as wide; anterior margin almost straight, posterior border convex, lacking tooth. Length ratio of articles from basis to merus about 4.8:1:4.5. Basis broad, 1.35 times as long as wide, anterior and posterior margins slightly and regularly convex, anterior margin with five spines along distal half and two distal setae, ten serrations all along posterior margin, each serration usually with one seta, distal corner rounded and produced into a small lobe. Ischium with three anterodistal setae, without spine. Merus 3.4 times as long as broad, with two pairs of setae on anterior margin, three spines on posterior margin on proximal two thirds, four anterodistal and four posterodistal setae.
Pereopod 7 (Figure 9E): broken in all specimens at carpus-merus level. Coxa 0.7 times as long as wide; anterior margin almost straight, posterior margin convex, lacking tooth. Length ratio of articles from basis to merus about 4.5:1:4.9. Basis broad, 1.35 times as long as wide, anterior and posterior margins convex, anterior margin with five spines and two distal setae, posterior margin with nine serrations, some accompanied by one small seta. Merus 3.3 times as long as broad, with two to three pairs of setae on anterior margin, five spines on posterior margin on proximal ⅔, five anterodistal and five posterodistal setae.
Pleonites (Figure 10A): all pleonites lacking posterodistal teeth. Pleonite 1 posterior margin almost straight, posteroventral corner rounded to slightly pointed. Pleonite 2 posterior margin straight, posteroventral corner with small tooth. Pleonite 3 posterior margin clearly convex on inferior half, posteroventral corner produced into a well-developed tooth.
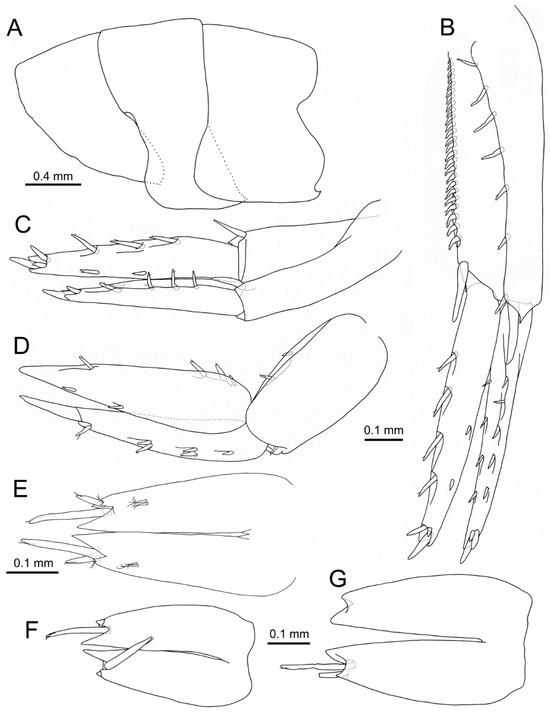
Figure 10.
Idunella spinifera (Dauvin and Gentil, 1983). Ovigerous female (A–E), non-ovigerous females (F,G), Ría de Muros (NW Spain). (A) pleonites, left side, lateral view. (B) right uropod 1, laterodorsal view. (C) right uropod 2, laterodorsal view. (D) right uropod 3, lateral view. (E–G) telson, dorsal view. (B–D,F,G) same scale. (F,G) dorsal and terminal setae, with setules not illustrated.
Urosomites: similar to male.
Uropod 1 (Figure 10B): peduncle 1.1 times as long as rami, six dorsolateral spines and a pair of distal spines consisting of a small one and a long spine pointing backwards, dorsomedial border with a row of nineteen to twenty-five spines and one distal spine; outer ramus with five outer and four medial spines, three spines on tip (one much longer); inner ramus with two outer spines and four medial longer and thicker spines, tip with three spines similar to outer ramus.
Uropod 2 (Figure 10C): peduncle subequal to rami, one distal dorsolateral spine and one distal dorsomedial spine; outer ramus with four outer and two medial spines, two spines on tip (one much longer); inner ramus with two outer and three medial spines, tip with three spines (central much longer).
Uropod 3 (Figure 10D): peduncle 0.7 times as long as rami, one anterodistal and two dorsomedial spines; outer and inner ramus subequal, outer ramus 2-articulated, article 2 0.35 times as long as article 1, article 1 with one spine and three pairs of spines on outer side; inner ramus with two outer and four medial spines on medial border.
Telson (Figure 10E): medial cleft 0.85 times as long as telson length; tooth and setae similar to male.
3.6. Variability of Selected Characters in Non-Ovigerous Females
Specimens measured 2.9–6.1 mm BL (n = 27). Accessory flagellum had 3–5 articles; antenna 1 flagellum had 7–13 articles. Antenna 2 flagellum had 4–10 articles. Dorsomedial border of uropod 1 peduncle had 10–21 spines. Telson, in most specimens, had two pairs of distal spines (n = 11), some with one pair (n = 2) (Figure 10F), often broken (Figure 10G).
4. Discussion
This second record of I. spinifera at the Ría de Muros increases its range of distribution 6° to the south into the NW Iberian Peninsula. Specimens were found in moderately sorted fully marine shallow sublittoral coarse biogenic sand, a habitat that is similar to that of the type locality [18]. The presence of males and ovigerous females between June and September suggest that the reproductive period of this species ranges from late spring to early autumn. Other liljeborgiids reported from the same geographic area are Idunella picta (Norman, 1899), Idunella nana (Schiecke, 1973), Liljeborgia pallida (Bate, 1857), and Sextonia longirostris Chevreux, 1820, mostly in medium and fine sands at shallow depths [22,23,24].
I. spinifera was described after one immature female [18], and was characterised by having an accessory antenna 1 flagellum with four articles, about twenty spines on the dorsomedial border of the uropod 1 peduncle, the outer ramus of uropod three being composed of two articles, and the telson bearing only one pair of distal spines. Examination of the Iberian specimens revealed that the number of accessory flagellum articles ranges from 3 in females, about 2.9–3.6 mm BL to 7 in larger ovigerous females, and 6 in males. Similarly, the number of dorsomedial spines in the uropod 1 peduncle increases from 10–11 in the smallest specimens to up to 23–25 in mature males and females. In contrast with original description, most specimens bear a telson that is provided with two pairs of distal spines instead of one; however, it seems that these spines are easily broken, and a few specimens bore only one pair.
The keys provided by [18,25] for European Atlantic and Mediterranean species separate I. spinifera, I. aeqvicornis G.O. Sars, 1877, and I. pirata Krapp-Schickel, 1975, regarding the presence of more than 10 dorsomedial spines on the uropod 1 peduncle, a short article 1 on the mandibular palp, a palp article 2 of maxilliped that bears dorsolateral setae, and urosomite 1 that lacks a posterodorsal tooth. I. spinifera differs from these species in the number of dorsomedial spines (up to 20–25) and in its lack of distolateral teeth in all its pleonites.
The male of I. spinifera is described here for the first time, and presents the diagnostic characters of the species. The most remarkable feature is the different appearance and size of gnathopod 1 when compared to gnathopod 2, the latter being similar to gnathopods 1–2 of the females. Males of I. aeqvicornis and I. pirata also bear a gnathopod 1 that is larger than gnathopod 2; however, in I. aeqvicornis and I. spinifera, gnathopod 1 has a triangular shape, whereas in I. pirata, gnathopod 1 and 2 are similar in shape to each other, with a palm that is regularly convex. Males of I. aeqvicornis mainly differ from those of I. spinifera in that the palm is regularly concave and is defined proximally by a large triangular process, and the dactylus lacks teeth.
Author Contributions
Conceptualization, J.M., M.R. and P.V.; methodology, J.M., M.R. and P.V.; investigation, J.M., M.R. and P.V.; writing—original draft preparation, J.M.; writing—review and editing, M.R. and P.V. All authors have read and agreed to the published version of the manuscript.
Funding
This study had the support of Fundação para a Ciência e Tecnologia (FCT) through LA/P/0069/2020 granted to the Associate Laboratory ARNET, DOI 10.54499/UIDB/04050/2020 granted to the CBMA (University of Minho) and P Veiga was funded by 10.54499/CEECIND/03893/2018/CP1574/CT0004.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The raw data supporting the conclusions of this article will be made available by the authors on request.
Acknowledgments
We are grateful to three anonymous referees for all the helpful comments and suggestions, which greatly improved this paper. Authors want to thank Fernando Aneiros, Jesús López, Jesús S. Troncoso, Marina Gómez, and Mariano Lastra for their valuable support in sampling.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Hughes, L.E.; Ahyong, S.T. Collecting and processing amphipods. J. Crustac. Biol. 2016, 36, 584–588. [Google Scholar] [CrossRef]
- Ritter, C.J.; Bourne, D.G. Marine amphipods as integral members of global ocean ecosystems. J. Exp. Mar. Biol. Ecol. 2024, 572, 151985. [Google Scholar] [CrossRef]
- Baldo, F.; Arias, A.M.; Drake, P. The macrobenthic community of the Guadalquivir estuary. Bol. Inst. Esp. Oceanogr. 2001, 17, 137–148. [Google Scholar]
- De-La-Ossa-Carretero, J.A.; Dauvin, J.-C.; Del-Pilar-Ruso, Y.; Giménez-Casalduero, F.; Sánchez-Lizaso, J.L. Inventory of benthic amphipods from fine sand community of the Iberian Peninsula east coast (Spain), western Mediterranean, with new records. Mar. Biodivers. Rec. 2010, 3, e119. [Google Scholar] [CrossRef]
- Legeżyńska, J.; Kędra, M.; Walkusz, W. When season does not matter: Summer and winter trophic ecology of Arctic amphipods. Hydrobiologia 2012, 684, 189–214. [Google Scholar] [CrossRef]
- Srinivas, T.; Sukumaran, S.; Neetu, S.; Ramesh Babu, K. Diversity and functional patterns of benthic amphipods in the coralline intertidal zones of a Marine National Park, India. Front. Mar. Sci. 2020, 7, 589195. [Google Scholar] [CrossRef]
- Manokaran, S.; Joydas, T.V.; Qurban, M.A.; Cheruvathur, L.L.; Kariyathil, T.J.; Basali, A.U.; Khan, S.A.; Al-Suwailem, A. Baseline patterns of structural and functional diversity of benthic amphipods in the western Arabian Gulf. Mar. Pollut. Bull. 2021, 164, 112054. [Google Scholar] [CrossRef] [PubMed]
- Guerra-García, J.M.; Tierno de Figueroa, J.M.; Navarro-Barranco, C.; Ros, M.; Sánchez-Moyano, J.E.; Moreira, J. Dietary analysis of the marine Amphipoda (Crustacea: Peracarida) from the Iberian Peninsula. J. Sea Res. 2014, 85, 508–517. [Google Scholar] [CrossRef]
- Nyssen, F.; Brey, T.; Lepoint, G.; Bouquegneau, J.-M.; De Broyer, C.; Dauby, P. A stable isotope approach to the eastern Weddell Sea trophic web: Focus on benthic amphipods. Polar Biol. 2002, 25, 280–287. [Google Scholar] [CrossRef][Green Version]
- Rigolet, C.; Dubois, S.F.; Thiébaut, E. Benthic control freaks: Effects of the tubiculous amphipod Haploops nirae on the specific diversity and functional structure of benthic communities. J. Sea Res. 2014, 85, 413–427. [Google Scholar] [CrossRef][Green Version]
- Burd, B.J.; Barnes, P.A.G.; Wright, C.A.; Thomson, R.E. A review of subtidal benthic habitats and invertebrate biota of the strait of Georgia, British Columbia. Mar. Environ. Res. 2008, 66, S3–S38. [Google Scholar] [CrossRef]
- Conlan, K.E. Amphipod crustaceans and environmental disturbance: A review. J. Nat. Hist. 1994, 28, 519–554. [Google Scholar] [CrossRef]
- Mancinelli, G.; Rossi, L. The influence of allochthonous leaf detritus on the occurrence of crustacean detritivores in the soft-bottom macrobenthos of the Po River Delta area (northwestern Adriatic Sea). Estuar. Coast. Shelf Sci. 2002, 54, 849–861. [Google Scholar] [CrossRef]
- Farlin, J.P.; Lewis, L.S.; Anderson, T.W.; Lai, C.T. Functional diversity in amphipods revealed by stable isotopes in an eelgrass ecosystem. Mar. Ecol. Prog. Ser. 2010, 420, 277–281. [Google Scholar] [CrossRef]
- Arfianti, T.; Wilson, S.; Costello, M.J. Progress in the discovery of amphipod crustaceans. PeerJ 2018, 6, e5187. [Google Scholar] [CrossRef] [PubMed]
- d’Udekem d’Acoz, C. New records of Liljeborgia from Antarctic and sub-Antarctic seas, with the description of two new species (Crustacea, Amphipoda, Liljeborgiidae). Bull. Inst. R. Sci. Nat. Belg. 2009, 79, 243–304. [Google Scholar]
- WoRMS Editorial Board. World Register of Marine Species; VLIZ: Oostende, Belgium, 2020; Available online: http://www.marinespecies.org (accessed on 19 May 2024).
- Dauvin, J.C.; Gentil, F. Description de deux nouvelles espèces de Liljeborgiidae des côtes françaises (Crustacea-Amphipoda) Listriella dentipalma n. sp. et L. spinifera n. sp. Cah. Biol. Mar. 1983, 24, 429–442. [Google Scholar]
- Souto, C.; Gilcoto, M.; Fariña-Busto, L.; Pérez, F.F. Modelling the residual circulation of a coastal embayment affected by wind-driven upwelling: Circulation of the Ría de Vigo (NW Spain). J. Geophys. Res. 2003, 108, 3340. [Google Scholar] [CrossRef]
- Figueiras, F.G.; Labarta, U.; Fernández Reiriz, M.J. Coastal upwelling, primary production and mussel growth in the Rías Baixas of Galicia. Hydrobiologia 2002, 484, 121–131. [Google Scholar] [CrossRef]
- López-Jamar, E. Spatial distribution of the infaunal benthic communities of the Ria de Muros, North-West Spain. Mar. Biol. 1981, 63, 29–37. [Google Scholar] [CrossRef]
- Garmendia, J.M.; Sánchez Mata, A.; Mora, J. Inventario de la macrofauna bentónica de sustratos blandos submareales de la Ría de Ares y Betanzos (NO de la Península Ibérica). Nova Acta Cient. Comp. 1998, 8, 209–231. [Google Scholar]
- Moreira, J.; Troncoso, J.S. Inventario de la macrofauna bentónica de sedimentos submareales de la Ensenada de Baiona (Galicia, NO Península Ibérica). Nova Acta Cient. Comp. 2007, 16, 101–128. [Google Scholar]
- Lourido, A.; Sorbe, J.C.; Troncoso, J.S. Inventario de los crustáceos bentónicos de sedimentos infralitorales de la Ría de Aldán (Galicia, NO Península Ibérica). Nova Acta Cient. Comp. 2008, 17, 149–168. [Google Scholar]
- Gouillieux, B.; Bonifacio, P.; Lavesque, N. Idunella bacheleti sp. nov., a new Liljeborgiidae species (Crustacea: Amphipoda) from the Capbreton Canyon (Bay of Biscay, NE Atlantic Ocean). Cah. Biol. Mar. 2020, 61, 311–322. [Google Scholar] [CrossRef]










Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).