Abstract
Artificial structures act as points of entry for non-indigenous species (NIS) in port areas and may support higher abundance and richness of them. The studies about NIS are increasing, but studies focusing on the variations in temporal recruitment and ecological mechanisms are still scarce. Thus, the aim of this work was to determine the colonization and development of non-indigenous sessile fouling species over two types of substrates (electrolytic carbonated and steel) during 12 months of immersion in the Alicante harbor. The biofouling communities of both substrates were analyzed in terms of abundance and species richness by status (native, cryptogenic, and NIS), and NIS assemblages of both substrates were studied by means of multivariate analyses. In total, 53 different species were identified, 38 in steel (six NIS and six cryptogenic) and 50 in the carbonated substrate (six NIS and 10 cryptogenic). Most NIS were more abundant and diverse after 9 months of immersion and had a preference for carbonated substrates. Furthermore, most of them were positively correlated in both substrates (mainly in steel) and it is noted that the number of NIS in the port of Alicante is increasing as new records have been detected.
1. Introduction
Biodiversity loss is one of the problems the world is facing nowadays. It is declining worldwide due to different reasons: global warming, overexploitation, changes in land and sea use, habitat destruction, and bioinvasions [1,2,3]. Bioinvasions are the introduction of non-indigenous species (hereafter NIS) in a different biogeographic region due to human activity, in both direct and indirect ways. In fact, the introduction of NIS and habitat modification are considered the most important drivers of biodiversity loss in coastal areas [4,5]. Almost all seas of the world have experienced the introduction of NIS [6,7,8], with the Mediterranean Sea being the most invaded one, with over 700 NIS [9,10].
During recent years, studies about NIS have increased, but studies focusing on the variations in temporal recruitment and ecological mechanisms are still scarce [11,12]. NIS has several effects on the host habitat, as they compete with native species for resources, such as food and space, and they can change the stablished community structure and alter the ecosystem functions by the substitution of species, up to the point of eliminating the native species [13,14]. Hence, they promote the biotic homogenization of biodiversity and functionality [13,15,16].
Biofouling and ballast waters are among the main human-mediated dispersal pathways for aquatic NIS, which make port areas, especially large ports, hotspots for the primary introduction of NIS [16,17]. In fact, those areas are the ones with a high number of human-made artificial infrastructures submerged in the marine environment. Those structures are made of both natural and artificial materials, but the artificial ones are more common, such as pontoons, buoys, and dikes. It has been seen that there are differences in biofouling assemblages within different substrates (e.g., [18,19,20]), and one of the reasons is that NIS are more frequent in artificial substrates than in naturally occurring ones [20,21,22]. As such, these artificial structures act as points of entry for NIS in port areas and may support more abundance and richness of them [23,24,25]. This is because artificial materials and port areas are an anthropologically altered environment, in which NIS can compete better than native ones, contrary to what happens in natural habitats [14,20,24].
In this context, there is an urgent need to determine the ecological and evolutionary mechanisms that promote NIS in order to make appropriate management and conservation strategies [2]. This is especially important when new structures are submerged in the marine environment, as it is aimed to recruit the least possible number of NIS [24,26]. Moreover, given the important role of port structures in the introduction and establishment of NIS, these areas are pivotal to assess the early detection of NIS for further management strategies [16].
Hence, we aimed to observe the colonization of sessile macrofouling from Alicante’s port on two types of substrates (electrolytic carbonated and steel) over 12 months. These materials were used as, contrary to concrete or PVC (the most commonly used materials), they do not have any negative impacts on the marine environment as they do not release any substances from them. In addition, as they are made out of the salts dissolved in the sea water, they are natural substrates and are similar in composition to natural limestone [27]. The colonization stages of biofouling species on these types of substrates have been analyzed in a previous work [28]. However, this study focused on the non-indigenous and cryptogenic species in order to observe how their development vary among substrates. It was hypothesized that the NIS would be more abundant on bare steel as it is a more artificial substrate. Moreover, we intended to observe the development and succession patterns of NIS within the different substrates.
2. Materials and Methods
2.1. Study Site
The study was conducted in the port of Alicante (SE Spain) in the 9th dock (N 38°20′09.23″–W 00°29′05.81″) for 12 months between 15 October 2019 and 13 October 2020. The dock is located in the outer quay of the harbor (Figure 1) and has a maximum depth of 10 m. During the study, the surface seawater mean temperature varied as shown, with a range from 14.1 to 28.3 °C (according to https://www.puertos.es/es-es accessed on 22 October 2022).

Figure 1.
Alicante’s harbor location and ninth dock position (white circle). Images adapted from Google Earth via QGIS 3.16.7.
2.2. Sampling
In order to study the development of non-indigenous and cryptogenic species over 12 months of succession, 24 carbon steel meshes of 15 × 15 cm2 were used: 12 with an electrolytically precipitated carbonated layer as in [29] and the other 12 only bare steel, acting as controls. All the meshes were arranged, alternately, in 3 profiles of 8 meshes each and were placed at 2 m depth, 1 m away from the dock. The meshes were separated 20 cm from each other, and the profiles were 5 m apart. Each mesh consisted of a quadrat of 15 × 15 cm2, without any subquadrat inside, as stated in [30]. To follow the colonization process according to the immersion time, every 3 months, 6 meshes (3 for each type of material) were collected.
In the lab, the samples were fixed with formalin at 10% with seawater for 48 h. Organisms were identified to the lowest taxon possible (based on the World Register of Marine Species: https://www.marinespecies.org accessed on 29 May 2024). For this study, only sessile macrofouling species were used. Moreover, the status of introduction (native, cryptogenic, or NIS) of each identified species was assessed by NIS inventories for the Mediterranean Sea [10,31,32,33,34] and specialized databases (World Register of Introduced Marine Species WRiMS: http://www.marinespecies.org/introduced/ accessed on 29 May 2024, National Exotic Marine and Estuarine Species Information System NEMESIS: http://invasions.si.edu/nemesis/ accessed on 29 May 2024) and European Alien Species Information Network (EASIN: https://easin.jrc.ec.europa.eu accessed on 29 May 2024).
2.3. Data Treatment
Both univariate and multivariate analyses were performed in order to evaluate the effect of the substrate and the immersion time over the sessile macrofouling. In all cases, a two-way test was carried out with substrate (orthogonal and fixed, with 2 levels) and the immersion time (orthogonal and fixed, with 4 levels) as factors. It has to be noted that the third replicate of the carbonated treatment in the last immersion time was lost, so the mean value between the other two was used as its value. Moreover, only taxa identified at the species level were used for the analysis.
In order to observe the effects of the substrates and the immersion time over the diversity and total abundance of the biofouling assemblages, a permutational ANOVA with Euclidean distance was performed [35]. This was carried out for the abundance (measured in organisms/cm2 or colonies/cm2) and for species richness detected on each sample. The analysis was performed by the status (native, non-indigenous and cryptogenic) independently. A permutational ANOVA was carried out instead of conventional ANOVA as the first one is not a parametric test and to assess the p-value with Monte Carlo method [36].
On the other hand, to determine the effect only in the NIS assemblages, a PERMANOVA was performed. The dataset was constructed with the abundances of NIS with a prior 4th root transformation and the Bray–Curtis similarity was used [37]. To supplement PERMANOVA, a non-metric multidimensional scaling (nMDS), also with the Bray–Curtis similarity, was performed to observe the results and the species with a correlation higher than 0.5 were graphed as vectors on it. As there were replicates without any NIS, they were erased to make the nMDS in order to correctly see the distribution and patterns (Figure S1). In addition, a PERMDISP was used to test whether the differences were caused by the factor or by the dispersion of the data [36]. Moreover, a Pearson correlation test was carried out among the NIS in order to detect facilitation or competence patterns among the species [38].
PERMANOVA, nMDS, and PERMDISP were conducted in PRIMERv6+PERMANOVA v1.0.3 software [39], whereas the correlation test was conducted in R 4.1.2 [40].
3. Results
3.1. Diversity
In this study, 61 different taxa were identified, most of them (53) to species level. Among them, 36 were natives, 7 were NIS, and 10 were cryptogenic (Table S1). Focusing on the different substrates, out of the 38 species of the control treatment, 26 (68.4%) were natives, 6 (15.8%) were NIS, and 6 (15.8%) were cryptogenic, whereas out of the 50 species of the carbonated treatment, 34 (68%) were natives, 6 (12%) were NIS, and 10 (20%) were cryptogenic. The recorded NIS were: Balanus trigonus, Botrylloides cf niger, Branchiomma bairdi, Hydroides dirampha, Hydroides elegans, Spirobranchus cf tetraceros, and Styela plicata. Moreover, B. cf. niger was only found on the carbonate substrate and S.cf. tetraceros only on the control treatment. The species recognized as cryptogenic were: Amphibalanus amphitrite, Bugula neritina, Bugulina fulva, Bugulina stolonifera, Ciona intestinalis, Diplosoma listerianum, Savignyella lafontii, Schizoporella errata, Styela canopus, and Watersipora subtorquata. Furthermore, out of the 10 known non-indigenous and nine cryptogenic sessile species in the Alicante harbor, three NIS and six cryptogenic were detected, highlighting the new records of the four NIS (H. dirampha, S. cf. tetraceros, B.trigonus, and B. cf. niger), and four cryptogenic species (B. fulva, B. stolonifera, S. lafontii, and W. subtorquata) (Table 1) for this area.

Table 1.
List of sessile non-indigenous and cryptogenic species recorded in the port of Alicante based on bibliography and this study. Status: (NIS) non-indigenous species; (Cryp) cryptogenic.
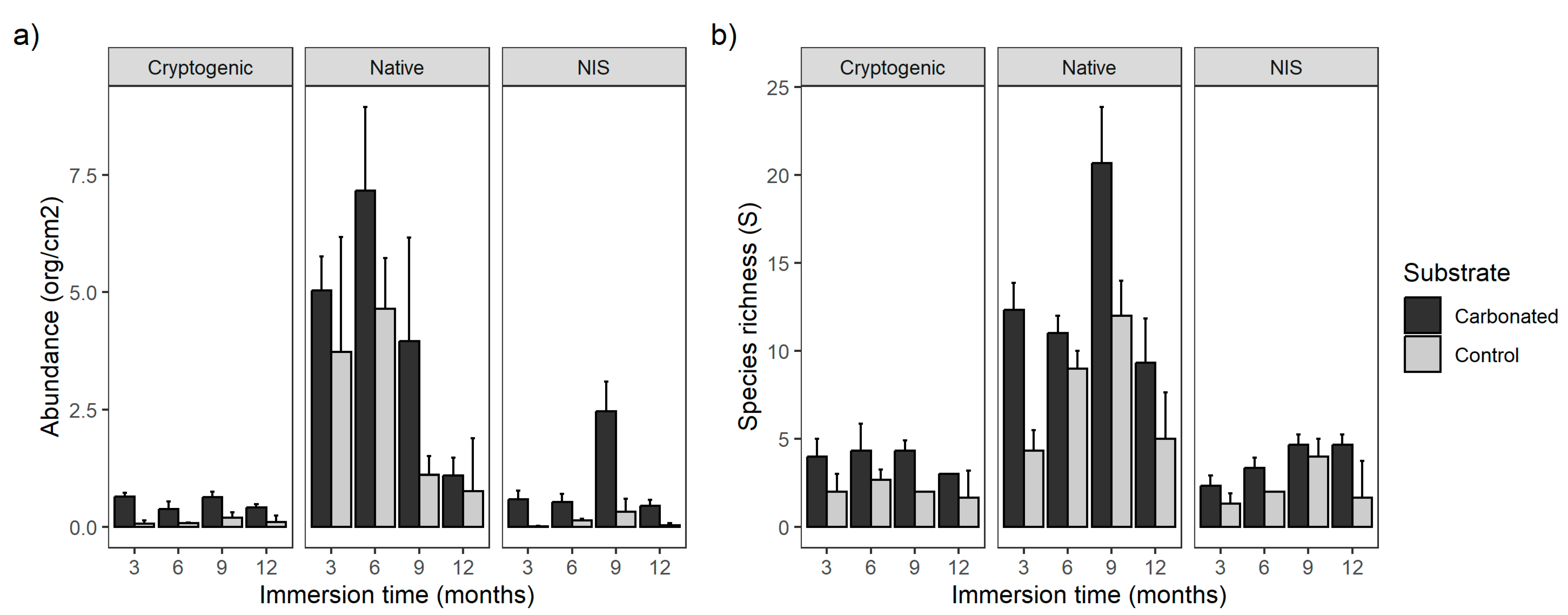
All species, independent of their status of introduction, were more abundant in the carbonated substrate (Figure 2a, Table 2) and the immersion time was significant, but only for NIS and cryptogenic species was the interaction of both factors significant. The native species were more abundant during the first two periods of immersion and after that they decreased, whereas NIS were more abundant after 9 months and then decreased, coinciding with the reduction of natives. Furthermore, NIS were more abundant in carbonated treatment during all immersion times. In the case of cryptogenic species, they were more abundant on the carbonated substrate after 3 months of immersion, but not during the other periods.
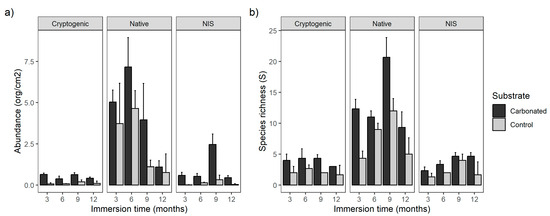
Figure 2.
Bar graph for abundance (a) and species richness (b) of each status of introduction type among substrates and immersion time.
Table 2.
Two-way crossed permutational ANOVA results for the abundance (organisms/cm2 or colonies/cm2) of native, NIS, and cryptogenic species for the substrate (=S) and immersion time (=T).
Referring to the richness, the trend was similar. All of them had their highest species richness on the carbonated treatment and the more diverse ones were natives, but in this case, all of them were affected by the immersion time (Figure 2b, Table 3), except for the cryptogenic species as their number was similar over all the study period with a small decreasing trend. Both native and NIS had their highest richness during the period of 9 months and the lowest after 3 months.
Table 3.
Two-way crossed permutational ANOVA results for the species richness of native, NIS, and cryptogenic species for the substrate (=S) and immersion time (=T). Notes: C = carbonated treatment, I = iron treatment.
3.2. Non-Indigenous and Cryptogenic Species
Over the seven recorded NIS, most of them were influenced by the substrate material, the immersion time, and the interaction (Table S2). Only one species (Spirobranchus cf. tetraceros) was not found to be influenced by the substrate material and the immersion time, so their recruitment, settlement, and development seemed to be influenced by other factors. Moreover, there were cases in which species did not appear in every period. This means that their appearance was linked to the immersion duration, as they needed a developed community before being recruited, or they needed a submersion season with specific environmental conditions to have a higher available propagule. This was the case of Branchiomma bairdi, Styela plicata, and Botrylloides cf. niger.
In the case of Balanus trigonus, B. bairdi, S. plicata, and B. cf. niger were significantly more abundant in the carbonated treatment, but this trend was not found in all the periods of immersion. Balanus trigonus was more abundant on the carbonated treatment until the last period, in which there were no differences, and S. plicata was more abundant after 9 and 12 months of immersion. A special case was B. bairdi, as it was more abundant in steel treatment after 6 months of immersion, but during the next periods was more abundant in carbonated treatment. On the other hand, both Hydroides dirampha and H. elegans were influenced by the immersion time and their interaction, but not by the material. In addition, they showed their higher abundances after 9 months, but this trend was not maintained after 12.
Five cryptogenic species were influenced by the substrate material or the immersion time (Table S3). In fact, four were influenced by the interaction of both factors. These species were Amphibalanus amphitrite, Watersipora subtorquata, Styela canopus, and Schizoporella errata. All of them were more abundant in the carbonated treatment: A. amphitrite was statistically more abundant after 3 and 9 months of immersion, H. elegans was more abundant only after 3 months, S. canopus after 9 and 12 months, and S. errata after 3, 6, and 12 months. Note that Bugulina fulva showed significant differences for the interactions, but the post hoc test was not capable of detecting those differences because of its low abundance. On the other hand, S. lafontii was clearly affected by the immersion time as it only appeared after 12 months.
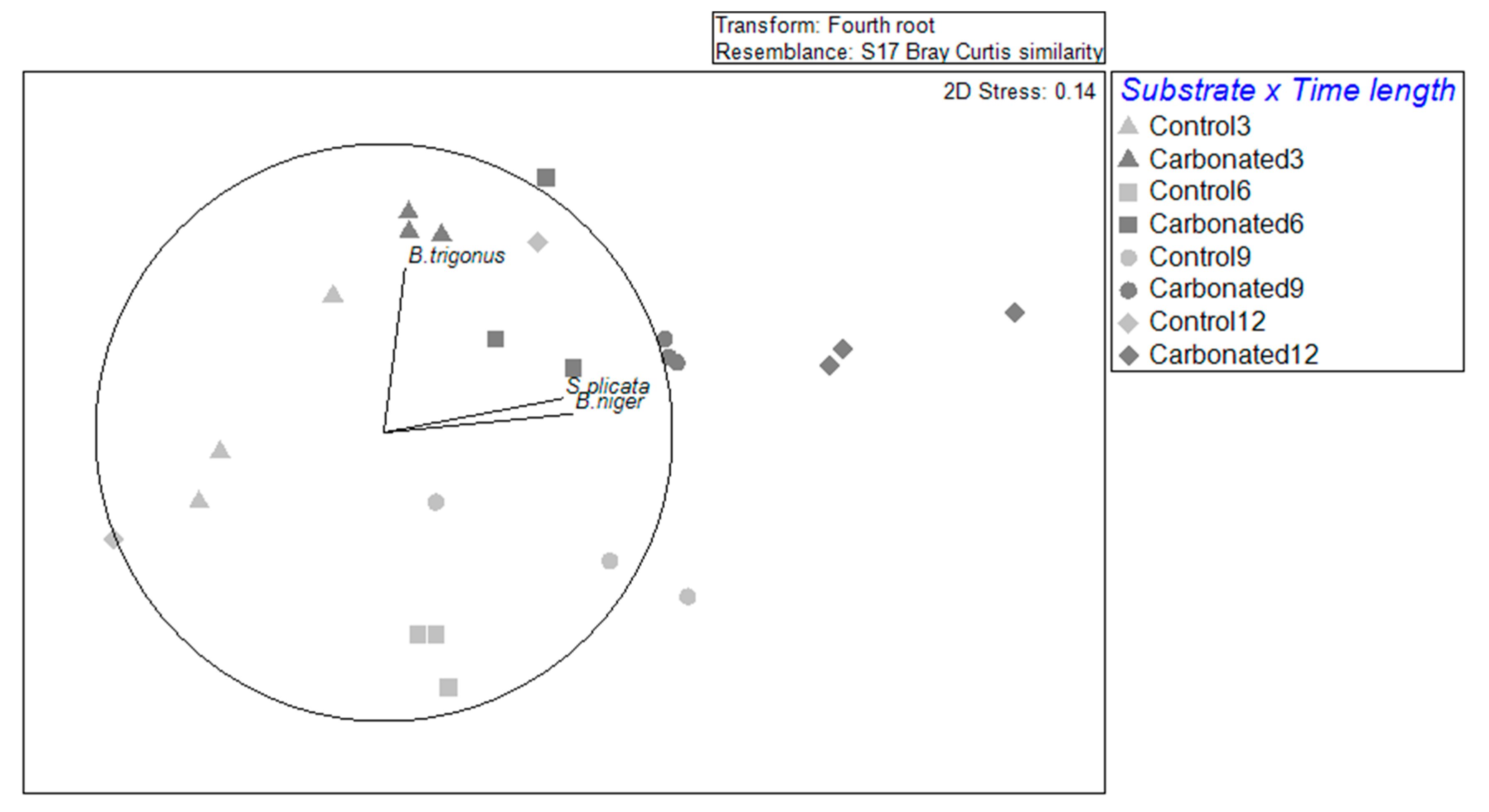
Focusing on the effect of NIS in the community, it is shown that these were affected by both substrate and immersion time and the interaction (Table 4). The dispersion was only significant (PERMDISP > 0.05) for the substrate, concretely in the steel substrate, as the samples were clearly more separated (Figure 3). In addition, by looking at the immersion time by substrate, samples were close to each other within the treatments, except for the carbonated treatment at 6 months and for the steel treatment at 3 and 12 months. In fact, it shows that there are species that highly correlated with substrates and immersion times. Ascidians S. plicata and B. cf. niger highly correlated with the 9 and 12 months in the carbonated substrate. In the case of B. bairdi, there was a correlation with the control treatment after 6 months, and B. trigonus correlated with the 3-month carbonated treatment.
Table 4.
PERMANOVA results for the abundance of macrofouling NIS assemblages for the different substrates and immersion time.
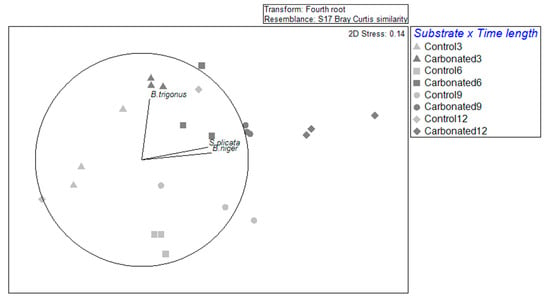
Figure 3.
A non-metric multidimensional scaling based on Bray–Curtis similarity of NIS biofouling assemblages for each immersion time and substrate type. The vectors represent the NIS with a correlation over 0.5.
Moreover, by looking at Pearson correlations (Table 5 and Table 6), almost all NIS (except for B. trigonus in the control and H. elegans and S. plicata in the carbonated) significantly correlated with, at least, one species. In the control treatment, all of them were positively correlated and a group made of H. dirampha, H. elegans, B. bairdi, and S. plicata formed as they were correlated together with values over 0.6. Regarding the carbonated treatment, B. trigonus and B. cf. niger were strongly negatively correlated and a group was formed by H. dirampha and B. bairdi, which correlated positively with values over 0.66.
Table 5.
Pearson correlation matrix of NIS appeared in the control treatment. Significant correlations are marked with *.
Table 6.
Pearson correlation matrix of NIS appeared in the carbonated treatment. Significant correlations are marked with *.
4. Discussion
The abundance of NIS, native, and cryptogenic species was higher in the carbonated treatment than in the steel one. Moreover, it was influenced by the immersion time, especially for the native species and NIS. Those two were offset during the succession, as at 6 months the natives showed their maximum abundance and NIS decreased, but after 9 months NIS abundance increased almost up to the native values, and they kept almost the same relative abundance to each other in the last period, although both abundances decreased. Similarly to the abundance, the same occurred for the species richness, except both the native species and NIS had their higher number of species at the third period (9 months). These results show that species were clearly influenced by seasonal larval supply and seasonality [12,49,50]. In fact, according to what was expected, the NIS had their highest abundance and species richness values after 9 months, from later spring to early–mid-summer (May–July), as they have been shown to have higher peaks in warmer months [49,51,52]. Some authors, such as Fargione et al. [53], have suggested that the success of NIS over natives is due to a non-overlapping recruitment period, but this trend does not have to be like the one experienced by [12] in the Central Mediterranean. Indeed, in our case, the trend was not completely clear, as abundances did not overlap in time, whereas the peak of species richness for both natives and NIS occurred at the same period, so more studies should be carried out in order to determine the trend in the port of Alicante.
Referring to the effect of the substrate over the species, contrary to expectations, both abundance and species richness of NIS were higher in the carbonated treatment. This is peculiar as NIS have a preference or they can compete better in artificial substrates than native ones, and because the carbonated material is very similar to naturally occurring limestone, so it was expected to have more NIS in the control treatment. Nevertheless, by looking at the control treatment, it is clear that NIS became more abundant than natives in their peak of recruitment (third period), agreeing with other studies [14,20,24]. The electrolytic carbonated substrate is so similar to natural rock that it imitates the home range substrate of a large number of species, independently of their status of introduction. This could explain the reason for having a high number of both natives and NIS, and why it enhanced the recruitment of them over the control treatment, where the expected trend occurred.
In port environments, it is not always clear what is the naturally occurring community as most of the substrates are artificial, it is an altered system [54], and there are (when there are) few previous studies about the inhabiting species. For the case of Alicante’s port, there are a few studies, of which, we highlight a study from [34,44,47], in which they found 10 NIS by rapid assessment protocol and both mobile and sessile macrofouling species were considered (Styela plicata, Celleporaria brunnea, Watersipora arcuata, Oculina patagonica, Caprella scaura, Mesanthura cf. romulea, Paranthura japonica, Branchiomma bairdi, Ficopomatus enigmaticus, and Hydroides elegans). In our study, a similar number of NIS (seven spp.) were found, but with some differences, as we did not find O. patagonica, F. enigmaticus, W. arcuata, and C. brunnea, while Spirobranchus cf. tetraceros and Botrylloides cf. niger where not found in [34,44]. It has to be highlighted that S.cf. tetraceros was only recorded in Spain in the port of Valencia [55] until this study. Although only one specimen has been found, it could mean that the population from Valencia is spreading, probably thanks to shipping, and it reached Alicante’s port. Moreover, this is the first record for B. cf. niger in the Spanish Mediterranean. However, one of the authors (AARE) probably detected this in 1979 and identified it as Botrylloides leachii in the port of Alicante during summer months [47]. Furthermore, there are some other new records for Alicante’s harbor that can have been overlooked due to the lack of studies, such as the case of Hydroides dirampha that has been detected since 1923 in Valencia [56], which could have arrived some years ago. Serpulids, Bryozoans, and Cirripedia are typical fouling species, and it is possible that the species recorded here have been in the study area for decades. In addition, it should be noted that cryptogenic and non-indigenous species abundance in the port of Alicante and recorded since the 1970s [45,47], such as the bryozoan Amathia verticillata (delle Chiaje, 1822), and the ascidians Clavelina lepadiformis (Müller, 1776), Botryllus schlosseri (Pallas, 1766), and Microcosmus squamiger Michaelsen, 1927 (as M. exasperatus), have not been collected on the meshes (carbonated and steel). So, it shows the need for a greater effort in temporal monitoring of present NIS in order to prevent further bioinvasions and local spreading. In fact, this monitoring should be a standardized method to remove the bias.
Referring to each NIS, it can be seen that there are three groups: (i) the ones not influenced by any of the studied factors (S. cf. tetraceros), (ii) ones influenced by both factors and interaction (B. bairdi, B. trigonus, S. plicata), and (iii) ones affected by immersion time and/or interaction (H. dirampha and H. elegans). As Hydroides spp. are the ones that dominate worldwide fouling communities [8,57], it makes sense for these to be time-dependent more than substrate-material-dependent, as they are gregarious species and grow over other serpulid tubes. In fact, H. elegans was only more abundant in the carbonated treatment in the first period, although it was abundant during the study, which means that it is a pioneer species, but it does not disappear. In the case of other species, this preference for the electrolytic carbonated substrate could be because of its similarity to limestone, as it has been said. Nevertheless, in all cases, the seasonality of species was demonstrated as most of them had their highest abundances after 9 months of immersion (third period), corresponding to warmer months. There was a case where high abundances were found, especially in the carbonated treatment, in the first months (Balanus trignous and H. elegans). Moreover, there were non-indigenous species that appeared only in the warmer periods, as in the case of B. cf. niger, and others appeared during the cold months and were maintained along the study (B. bairdi and S. plicata). This could be due to the propagule supply and because they need a specific development stage of the community.
Among cryptogenic species, Savignyella lafontii presented a specific development stage of the community as it only appeared in the last period, and Amphibalanus amphitrite, S. plicata, S. errata, and Watersipora subtorquata were influenced by the substrate and immersion length. A. amphitrite was more abundant in the carbonated treatment after 3 and 9 months, and after 12 months it did not appear alive, only the shell. This could mean that A. amphitrite acts as a pioneer species and, as the community develops it leaves the space to other species, such as S. lafontii, as it cannot compete against them. One of those species could be S. errata, which has a laminar growth and can grow over the shell, limiting or blocking its feeding capacity. In fact, this could be crucial in the development of the community, as it generates more available hard substrate for the colonization of other species, as happens with the shells of dead cirripedes. Moreover, S. errata was more abundant in the carbonated treatment for almost every sample, which goes along with the fact that this substrate has more space (carbonated layer plus other hard bodied organisms) to be colonized. Furthermore, W. subtorquata could have a similar behavior to S. errata as it also has this laminar growth, but this species seems to be influenced by the immersion period. Nevertheless, there are some species, mainly ascidians, that have not been recorded during the study and they are known for the port. This implies that these substrates are not suitable for their settlement, or they need a more developed community to correctly settle and develop.
Regarding NIS assemblages, it was observed that they were differentiated by substrate and by immersion time, as there are two groups: 3 to 6 months and 9 to 12 months. This trend was especially marked in the control treatment, and it matched the separation of months by their temperature: the colder months and the warmer ones. In the carbonated treatment, this separation could be seen between the first three months and the last 9 and 12, being the stage of 6 months a transitory and dispersed time. These results go along with what was observed, studying the whole fouling community in these typed of substrates [28]. In addition, it also suggests a development of the assemblages, which could be seen in the carbonated treatment, for which B. trigonus will be a pioneer species and, as far as the immersion duration goes ahead, ascidians will appear, and they will be more important in that step of succession.
All non-indigenous species were positively correlated between them in the steel treatment. This result, and the fact that steel is an artificial substrate, will support the idea that NIS can enhance the recruitment of each other, being the reason for appearing as better colonizers than natives in artificial materials. On the other hand, in the carbonated treatment, there were species with negative correlations. These were B. cf. niger with B. trigonus. These negative correlations could be caused by the encrusting growth of B. cf. niger as it could grow over the barnacle, hindering its capacity to feed. In fact, B. cf. niger negatively correlated with the rest of the species, except for S. plicata, supporting the idea that its growth type prevents the growth, feeding, and/or settlement of other species.
In addition, all tube-forming species (order Sabellida) correlated together, which would indicate that the species belonging to the taxa tend to recruit together. These results could mean that they not only do not compete with each other for the space, but also, they can co-occur within the same habitat and enhance the recruitment of others. In fact, this was seen with W. subtorquata in the carbonated treatment, which used the formed new substrate to grow, and with S. plicata in the control treatment. In this last case, both taxa benefitted from each other, as the ascidian could use the new substrata and generate more substrate with its tunic to the growth of other species [58]. Moreover, these species are the typical ones with high abundance and rapid growth (the case of serpulids, principally) in port areas, as well as other artificial substrates and perturbed areas [57,59], which could be a good explanation.
Taking all of this into account, most of the NIS have demonstrated a preference for the electrolytic carbonated substrate and the NIS that appeared are the typical ones in the fouling communities worldwide. It has been shown that the trend in artificial substrates in the recruitment of NIS is not always happening, so there is a need to determine which types of substrates recruit more NIS. In addition, the number of NIS present in the port of Alicante is increasing over the years and there is no control over it, so, there is a need for monitoring port environments in order to assess and to control new bioinvasions and the spreading of occurring NIS. Thus, as NIS preferred the electrolytic carbonated substrate, it would be a suitable material as an early detection tool of NIS in port areas. Actually, even though NIS were more abundant on it, native ones were too and the relative abundance between them was higher than in steel, so it can be considered a better construction material for those areas.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/jmse12071188/s1, Figure S1: A non-metric multidimensional scaling based on Bray–Curtis similarity of NIS biofouling assemblages for each immersion time and substrate type.; Table S1: Average abundance (X+SE) of the observed taxons (organisms/cm2) by moth and substrata: (C) carbonated; (S) steel; Table S2. Univariate PERMANOVA results for the abundance of biofouling non-indigenous species over different substrates (S) and immersion times (T). Note: na = not appeared; Table S3. Univariate PERMANOVA results for the abundance of biofouling cryptogenic species over different substrates (S) and immersion times (T). Note: na = not appeared.
Author Contributions
Conceptualization, M.-Á.C. and A.A.R.-E.; methodology, A.C.-R., C.A., M.-Á.C., P.G., V.M., E.A.-M. and A.A.R.-E.; formal analysis, A.C.-R.; investigation, A.C.-R., C.A., M.-Á.C., P.G., V.M., E.A.-M. and A.A.R.-E.; writing—original draft preparation, A.C.-R.; writing—review and editing, C.A., M.-Á.C., P.G., V.M., E.A.-M. and A.A.R.-E.; supervision, M.-Á.C. and A.A.R.-E.; project administration, A.A.R.-E.; funding acquisition, A.A.R.-E. All authors have read and agreed to the published version of the manuscript.
Funding
This research forms part of the ThinkInAzul program and was funded by Ministerio de Ciencia e Innovación with funding from European Union NextGenerationEU (PRTR-C17.I1) and by Generalitat Valenciana (THINKINAZUL/2021/014).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available on request from the corresponding authors.
Acknowledgments
The authors would like to express their gratitude to the Port Authority of Alicante for their support and for facilitating the work in the port, as well as to Fran Ramil from the University of Vigo for helping us in the identification of hydrozoans.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Catford, J.A.; Bode, M.; Tilman, D. Introduced Species That Overcome Life History Tradeoffs Can Cause Native Extinctions. Nat. Commun. 2018, 9, 2131. [Google Scholar] [CrossRef] [PubMed]
- Leclerc, J.; Viard, F.; González Sepúlveda, E.; Díaz, C.; Neira Hinojosa, J.; Pérez Araneda, K.; Silva, F.; Brante, A. Habitat Type Drives the Distribution of Non-indigenous Species in Fouling Communities Regardless of Associated Maritime Traffic. Divers Distrib. 2020, 26, 62–75. [Google Scholar] [CrossRef]
- Chaudhary, C.; Richardson, A.J.; Schoeman, D.S.; Costello, M.J. Global Warming Is Causing a More Pronounced Dip in Marine Species Richness around the Equator. Proc. Natl. Acad. Sci. USA 2021, 118, e2015094118. [Google Scholar] [CrossRef] [PubMed]
- Airoldi, L.; Bulleri, F. Anthropogenic Disturbance Can Determine the Magnitude of Opportunistic Species Responses on Marine Urban Infrastructures. PLoS ONE 2011, 6, e22985. [Google Scholar] [CrossRef] [PubMed]
- Anton, A.; Geraldi, N.R.; Lovelock, C.E.; Apostolaki, E.T.; Bennett, S.; Cebrian, J.; Krause-Jensen, D.; Marbà, N.; Martinetto, P.; Pandolfi, J.M.; et al. Global Ecological Impacts of Marine Exotic Species. Nat. Ecol. Evol. 2019, 3, 787–800. [Google Scholar] [CrossRef] [PubMed]
- Robinson, T.; Griffiths, C.; McQuaid, C.; Rius, M. Marine Alien Species of South Africa—Status and Impacts. Afr. J. Mar. Sci. 2005, 27, 297–306. [Google Scholar] [CrossRef]
- Smith, C.R.; Grange, L.J.; Honig, D.L.; Naudts, L.; Huber, B.; Guidi, L.; Domack, E. A Large Population of King Crabs in Palmer Deep on the West Antarctic Peninsula Shelf and Potential Invasive Impacts. Proc. R. Soc. B Biol. Sci. 2012, 279, 1017–1026. [Google Scholar] [CrossRef]
- Albano, M.J.; Obenat, S.M. Fouling Assemblages of Native, Non-Indigenous and Cryptogenic Species on Artificial Structures, Depths and Temporal Variation. J. Sea Res. 2019, 144, 1–15. [Google Scholar] [CrossRef]
- Galil, B.; Marchini, A.; Occhipinti-Ambrogi, A.; Ojaveer, H. The Enlargement of the Suez Canal—Erythraean Introductions and Management Challenges. Manag. Biol. Invasions 2017, 8, 141–152. [Google Scholar] [CrossRef]
- Zenetos, A.; Albano, P.G.; López García, E.; Stern, N.; Tsiamis, K.; Galanidi, M. Established Non-Indigenous Species Increased by 40% in 11 Years in the Mediterranean Sea. Mediterr. Mar. Sci. 2022, 23, 196–212. [Google Scholar] [CrossRef]
- Stachowicz, J.J.; Terwin, J.R.; Whitlatch, R.B.; Osman, R.W. Linking Climate Change and Biological Invasions: Ocean Warming Facilitates Nonindigenous Species Invasions. Proc. Natl. Acad. Sci. USA 2002, 99, 15497–15500. [Google Scholar] [CrossRef] [PubMed]
- Lezzi, M.; Del Pasqua, M.; Pierri, C.; Giangrande, A. Seasonal Non-Indigenous Species Succession in a Marine Macrofouling Invertebrate Community. Biol. Invasions 2018, 20, 937–961. [Google Scholar] [CrossRef]
- Levine, J.M. Species Diversity and Biological Invasions: Relating Local Process to Community Pattern. Science 2000, 288, 852–854. [Google Scholar] [CrossRef] [PubMed]
- Byers, J.E. Impact of Non-indigenous Species on Natives Enhanced by Anthropogenic Alteration of Selection Regimes. Oikos 2002, 97, 449–458. [Google Scholar] [CrossRef]
- Capinha, C.; Essl, F.; Seebens, H.; Moser, D.; Pereira, H.M. The Dispersal of Alien Species Redefines Biogeography in the Anthropocene. Science 2015, 348, 1248–1251. [Google Scholar] [CrossRef] [PubMed]
- Marraffini, M.; Ashton, G.; Brown, C.; Chang, A.; Ruiz, G. Settlement Plates as Monitoring Devices for Non-Indigenous Species in Marine Fouling Communities. Manag. Biol. Invasions 2017, 8, 559–566. [Google Scholar] [CrossRef]
- Outinen, O.; Puntila-Dodd, R.; Barda, I.; Brzana, R.; Hegele-Drywa, J.; Kalnina, M.; Kostanda, M.; Lindqvist, A.; Normant-Saremba, M.; Ścibik, M.; et al. The Role of Marinas in the Establishment and Spread of Non-Indigenous Species in Baltic Sea Fouling Communities. Biofouling 2021, 37, 984–997. [Google Scholar] [CrossRef] [PubMed]
- Brown, C.J. Epifaunal Colonization of the Loch Linnhe Artificial Reef: Influence of Substratum on Epifaunal Assemblage Structure. Biofouling 2005, 21, 73–85. [Google Scholar] [CrossRef] [PubMed]
- Vaz-Pinto, F.; Torrontegi, O.; Prestes, A.C.L.; Álvaro, N.V.; Neto, A.I.; Martins, G.M. Invasion Success and Development of Benthic Assemblages: Effect of Timing, Duration of Submersion and Substrate Type. Mar. Environ. Res. 2014, 94, 72–79. [Google Scholar] [CrossRef]
- Chase, A.L.; Dijkstra, J.A.; Harris, L.G. The Influence of Substrate Material on Ascidian Larval Settlement. Mar. Pollut. Bull. 2016, 106, 35–42. [Google Scholar] [CrossRef]
- Airoldi, L.; Turon, X.; Perkol-Finkel, S.; Rius, M. Corridors for Aliens but Not for Natives: Effects of Marine Urban Sprawl at a Regional Scale. Divers Distrib. 2015, 21, 755–768. [Google Scholar] [CrossRef]
- Megina, C.; González-Duarte, M.M.; López-González, P.J. Benthic Assemblages, Biodiversity and Invasiveness in Marinas and Commercial Harbours: An Investigation Using a Bioindicator Group. Biofouling 2016, 32, 465–475. [Google Scholar] [CrossRef] [PubMed]
- Glasby, T.M.; Connell, S.D.; Holloway, M.G.; Hewitt, C.L. Nonindigenous Biota on Artificial Structures: Could Habitat Creation Facilitate Biological Invasions? Mar. Biol. 2007, 151, 887–895. [Google Scholar] [CrossRef]
- Tyrrell, M.C.; Byers, J.E. Do Artificial Substrates Favor Nonindigenous Fouling Species over Native Species? J. Exp. Mar. Biol. Ecol. 2007, 342, 54–60. [Google Scholar] [CrossRef]
- Dafforn, K.A.; Glasby, T.M.; Johnston, E.L. Comparing the Invasibility of Experimental “Reefs” with Field Observations of Natural Reefs and Artificial Structures. PLoS ONE 2012, 7, e38124. [Google Scholar] [CrossRef] [PubMed]
- Cangussu, L.C.; Altvater, L.; Haddad, M.A.; Cabral, A.C.; Heyse, H.L.; Rocha, R.M. Substrate Type as a Selective Tool against Colonization by Non-Native Sessile Invertebrates. Braz. J. Oceanogr. 2010, 58, 219–231. [Google Scholar] [CrossRef]
- Farfan, G.A.; Apprill, A.; Cohen, A.; Decarlo, T.M.; Post, J.E.; Waller, R.G.; Hansel, C.M. Crystallographic and Chemical Signatures in Coral Skeletal Aragonite. Coral. Reefs 2022, 41, 19–34. [Google Scholar] [CrossRef]
- Carmona-Rodríguez, A.; Antón, C.; Climent, M.-Á.; Garcés, P.; Montiel, V.; Ramos-Esplá, A.A. Sessile Biofouling on Electrolytic Carbonated Structures: Stages of Colonization and Succession. J. Mar. Sci. Eng. 2024, 12, 443. [Google Scholar] [CrossRef]
- Hilbertz, W.H. Electrodeposition of Minerals in Sea Water: Experiments and Applications. IEEE J. Ocean. Eng. 1979, 4, 94–113. [Google Scholar] [CrossRef]
- Antón, C.; Carmona, A.; Climent, M.Á.; Garcés, P.; Montiel, V.; Ramos-Esplá, A.Á. Sistema Para La Formación de Arrecifes Marinos Artificiales y Estructuras Submarinas Con Recubrimiento Calcáreo Inducido Por Electrólisis. Rev. Digit. Del Cedex 2024, 203, 33–42. [Google Scholar]
- Zenetos, A.; Çinar, M.E.; Crocetta, F.; Golani, D.; Rosso, A.; Servello, G.; Shenkar, N.; Turon, X.; Verlaque, M. Uncertainties and Validation of Alien Species Catalogues: The Mediterranean as an Example. Estuar. Coast Shelf. Sci. 2017, 191, 171–187. [Google Scholar] [CrossRef]
- Zenetos, A.; Gofas, S.; Verlaque, M.; Cinar, M.E.; García Raso, J.E.; Bianchi, C.N.; Morri, C.; Azzurro, E.; Bilecenoglu, M.; Froglia, C.; et al. Alien Species in the Mediterranean Sea by 2010. A Contribution to the Application of European Union’s Marine Strategy Framework Directive (MSFD). Part I. Spatial Distribution. Mediterr. Mar. Sci. 2010, 11, 381. [Google Scholar] [CrossRef]
- Zenetos, A.; Gofas, S.; Morri, C.; Rosso, A.; Violanti, D.; García Raso, J.E.; Cinar, M.E.; Almogi-Labin, A.; Ates, A.S.; Azurro, E.; et al. Alien Species in the Mediterranean Sea by 2012. A Contribution to the Application of European Union’s Marine Strategy Framework Directive (MSFD). Part 2. Introduction Trends and Pathways. Mediterr. Mar. Sci. 2012, 13, 328–352. [Google Scholar] [CrossRef]
- Ulman, A.; Ferrario, J.; Occhpinti-Ambrogi, A.; Arvanitidis, C.; Bandi, A.; Bertolino, M.; Bogi, C.; Chatzigeorgiou, G.; Çiçek, B.A.; Deidun, A.; et al. A Massive Update of Non-Indigenous Species Records in Mediterranean Marinas. PeerJ 2017, 5, e3954. [Google Scholar] [CrossRef] [PubMed]
- McArdle, B.H.; Anderson, M.J. Fitting Multivariate Models to Community Data: A Comment on Distance-based Redundancy Analysis. Ecology 2001, 8, 290–297. [Google Scholar] [CrossRef]
- Anderson, M.J. Permutational Multivariate Analysis of Variance (PERMANOVA). In Wiley StatsRef: Statistics Reference Online; Wiley: Hoboken, NJ, USA, 2017; pp. 1–15. [Google Scholar]
- Clarke, K.; Gorley, R.; Somerfield, P.; Warwick, R. Change in Marine Communities: An Approach to Statistical Analysis and Interpretation, 3rd ed.; Primer-E Ltd.: Auckland, New Zealand, 2014. [Google Scholar]
- Hollander, M.; Wolfe, D.A.; Chicken, E. Nonparametric Statistical Methods; Sheahart, W.A., Wilks, S.S., Eds.; John Wiley & Sons: New York, NY, USA, 1973. [Google Scholar]
- Anderson, M.J.; Gorley, R.N.; Clarke, K.R. PERMANOVA+ for PRIMER: Guide to Software and Statistical Methods; Primer-E Ltd.: Auckland, New Zealand, 2008; pp. 1–224. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Core Team: Vienna, Austria, 2021. [Google Scholar]
- Dailianis, T.; Akyol, O.; Babali, N.; Bariche, M.; Crocetta, F.; Gerovasileau, V.; Chanem, R.; Gökoglu, M.; Hasiotis, T.; Izquierdo-Muñoz, A.; et al. New Mediterranean Biodiversity Records (July 2016). Mediterr. Mar. Sci. 2016, 17, 608. [Google Scholar] [CrossRef]
- Zibrowius, H.; Ramos, A.A. Oculina Patagonica, Scléractiniaire Exotiqueen Méditerranée Nouvelles Observations Dans Le Sud-Est de l’Espagne. Rep. Int. Comm. Mediterr. Sea 1983, 28. [Google Scholar]
- Del-Pilar-Ruso, Y.; San Martín, G.; Giménez-Casalduero, F.; López, E.; de-la-Ossa-Carretero, J.A.; Ramos-Esplá, A.A.; Sánchez-Lizaso, J.L. Interesting Polychaeta Species in Alicante Bay (W Mediterranean): Syllidae and Sabellidae. In Proceedings of the 11th International Polychaete Conference, Sidney, Australia, 4–9 August 2013. [Google Scholar]
- Ulman, A. Recreational Boating as a Major Vector of Spread of Non-Indigenous Species around the Mediterranean. Ph.D. Thesis, University of Sorbonne and University of Pavia, Paris, France, 2018. [Google Scholar]
- Katsanevakis, S. Unpublished Mediterranean Records of Marine Alien and Cryptogenic Species. Bioinvasions Rec. 2020, 9, 165–182. [Google Scholar] [CrossRef]
- Zibrowius, H. Ongoing Modification of the Mediterranean Marine Fauna and Flora by the Establishment of Exotic Species. Mésogée 1991, 5, 83–107. [Google Scholar]
- Ramos-Esplá, A.A. Ascidias Litorales Del Mediterráneo Ibérico: Faunística, Ecología y Biogeografía; Universidad de Barcelona: Barcelona, Spain, 1988. [Google Scholar]
- Cornet, C.; Ramos-Esplá, A.A. Ascidiacea. In Estudios Sistemático y Ecológico de las Esponjas y Ascidias del Mediterráneo Español; Fundación Juan March: Madrid, Spain, 1980; pp. 362–499. [Google Scholar]
- Cifuentes, M.; Krueger, I.; Dumont, C.P.; Lenz, M.; Thiel, M. Does Primary Colonization or Community Structure Determine the Succession of Fouling Communities? J. Exp. Mar. Biol. Ecol. 2010, 395, 10–20. [Google Scholar] [CrossRef]
- Lara-Romero, C.; de la Cruz, M.; Escribano-Ávila, G.; García-Fernández, A.; Iriondo, J.M. What Causes Conspecific Plant Aggregation? Disentangling the Role of Dispersal, Habitat Heterogeneity and Plant–Plant Interactions. Oikos 2016, 125, 1304–1313. [Google Scholar] [CrossRef]
- Lu, L.; Wu, R.S.S. Seasonal Effects on Recolonization of Macrobenthos in Defaunated Sediment: A Series of Field Experiments. J. Exp. Mar. Biol. Ecol. 2007, 351, 199–210. [Google Scholar] [CrossRef]
- Fortič, A.; Mavrič, B.; Pitacco, V.; Lipej, L. Temporal Changes of a Fouling Community: Colonization Patterns of the Benthic Epifauna in the Shallow Northern Adriatic Sea. Reg. Stud. Mar. Sci. 2021, 45, 101818. [Google Scholar] [CrossRef]
- Fargione, J.; Brown, C.S.; Tilman, D. Community Assembly and Invasion: An Experimental Test of Neutral versus Niche Processes. Proc. Natl. Acad. Sci. USA 2003, 100, 8916–8920. [Google Scholar] [CrossRef] [PubMed]
- Tempesti, J.; Langeneck, J.; Maltagliati, F.; Castelli, A. Macrobenthic Fouling Assemblages and NIS Success in a Mediterranean Port: The Role of Use Destination. Mar. Pollut. Bull. 2020, 150, 110768. [Google Scholar] [CrossRef] [PubMed]
- Palero, F.; Torrado, H.; Perry, O.; Kupriyanova, E.; Ulman, A.; Genis-Armero, R.; Ten Hove, H.A.; Capaccioni-Azzati, R. Following the Phoenician Example: Western Mediterranean Colonization by Spirobranchus Cf. Tetraceros(Annelida: Serpulidae). Sci. Mar. 2020, 84, 83. [Google Scholar] [CrossRef]
- Zibrowius, H. Remarques Sur Trois Espèces de Serpulidae Acclimatées En Méditerranée: Hydroides Dianthus (Verrill, 1873), Hydroides Dirampha Mörch, 1863, et Hydroides Elegans (Haswell, 1883). Rep. Int. Comm. Mediterr. Sea 1973, 21, 683–686. [Google Scholar]
- Gollasch, S.; Haydar, D.; Minchin, D.; Wolff, W.J.; Reise, K. Introduced Aquatic Species of the North Sea Coasts and Adjacent Brackish Waters. In Biological Invasions in Marine Ecosystems: Ecological, Management, and Geographic Perspectives; Springer: Berlin/Heidelberg, Germany, 2009; pp. 507–528. [Google Scholar]
- Castilla, J.C.; Lagos, N.A.; Cerda, M. Marine Ecosystem Engineering by the Alien Ascidian Pyura Praeputialis on a Mid-Intertidal Rocky Shore. Mar. Ecol. Prog. Ser. 2004, 268, 119–130. [Google Scholar] [CrossRef]
- Giachetti, C.B.; Battini, N.; Castro, K.L.; Schwindt, E. Invasive Ascidians: How Predators Reduce Their Dominance in Artificial Structures in Cold Temperate Areas. J. Exp. Mar. Biol. Ecol. 2020, 533, 151459. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).