
Numerical Studies on the Hydrodynamic Patterns and Energy-Saving Advantages of Fish Swimming in Vortical Flows of an Upstream Cylinder

Abstract
:1. Introduction
2. Physical and Numerical Models
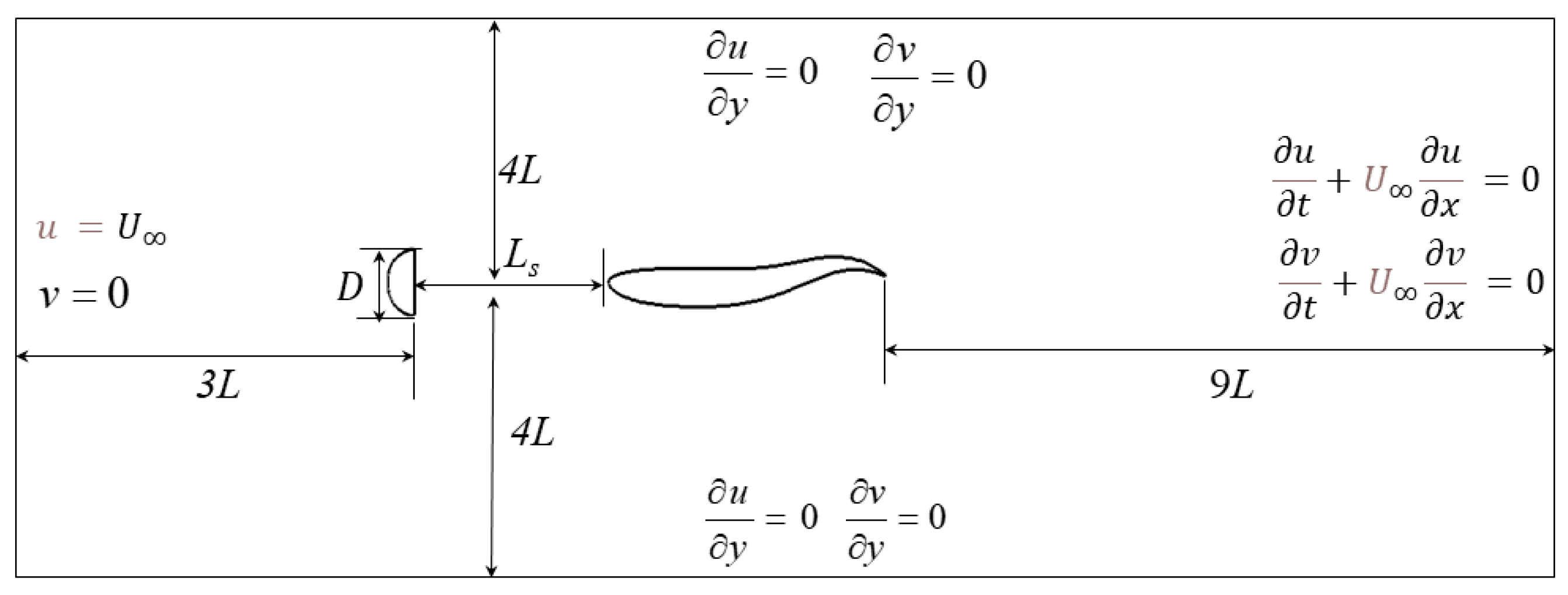
2.1. Problem Description
2.2. Numerical Model
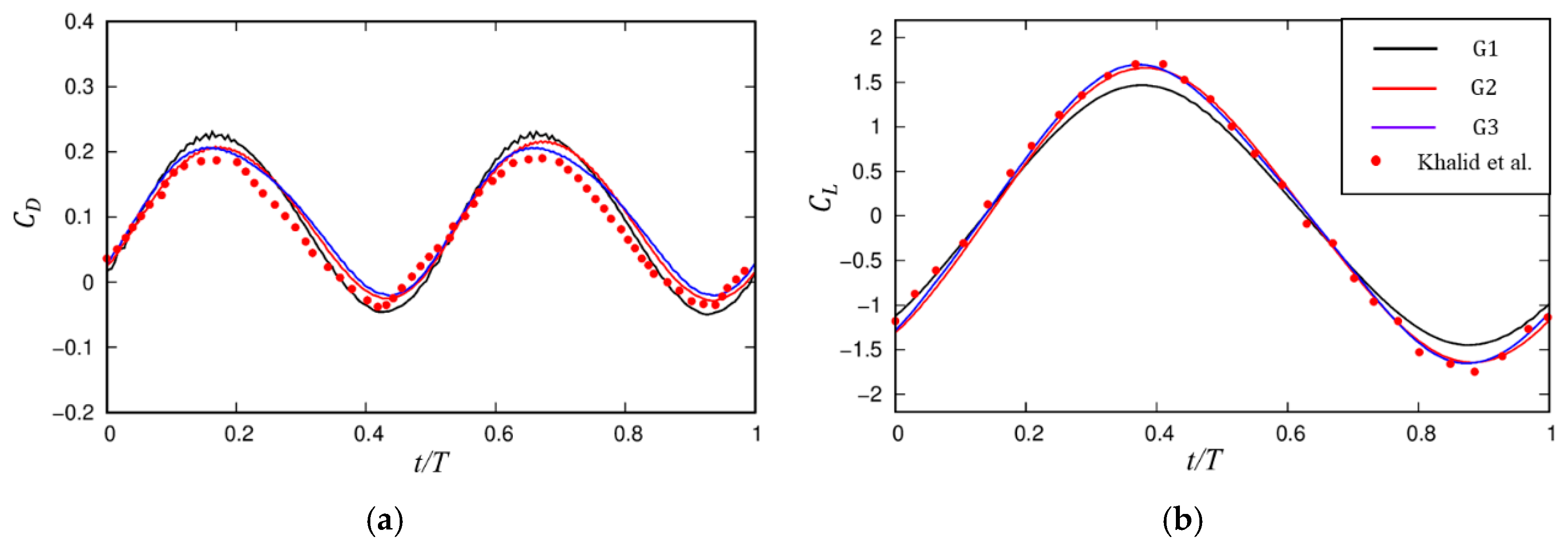
2.3. Numerical Validation
3. Numerical Results and Discussion
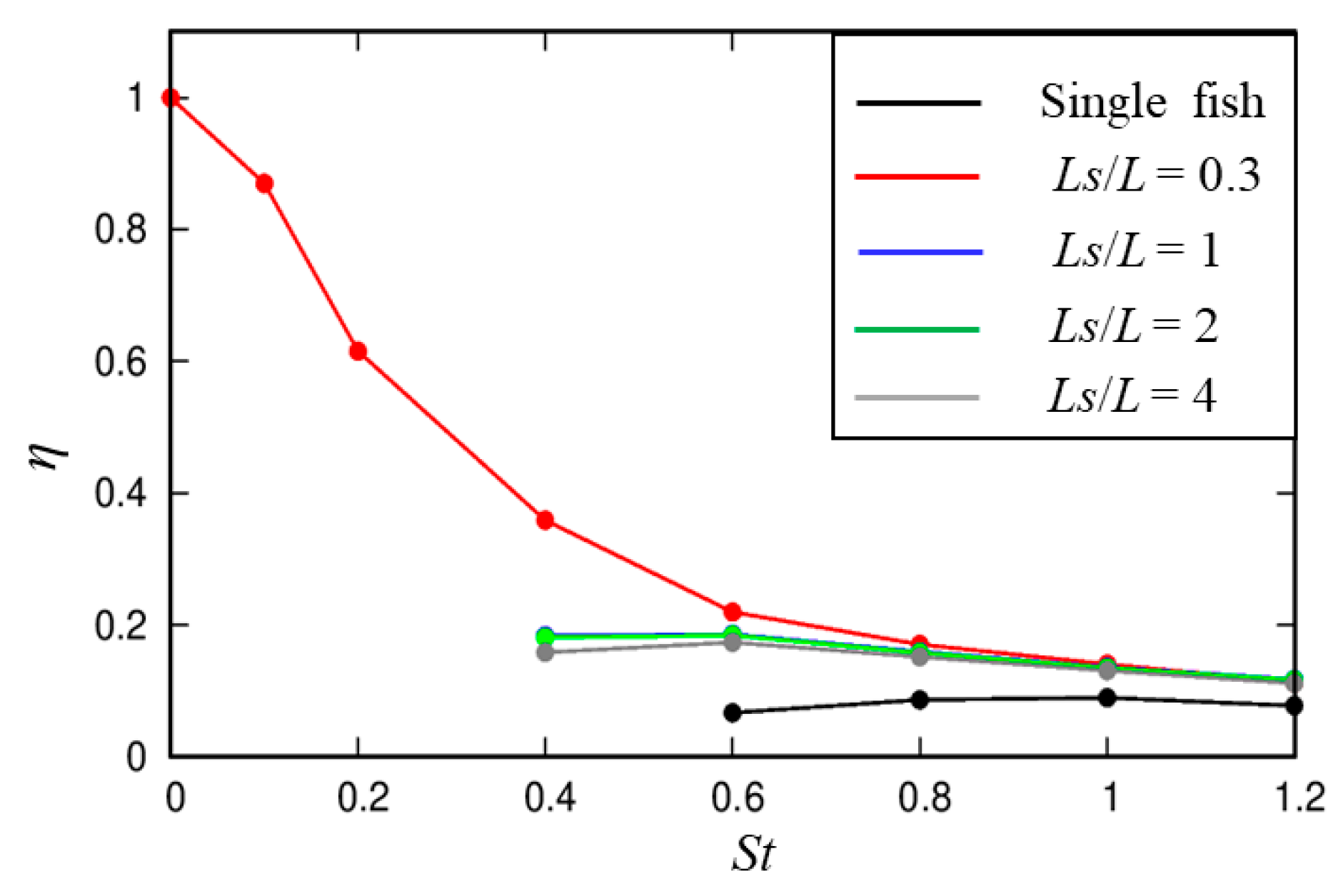
3.1. Effects of the Location of the Cylinder
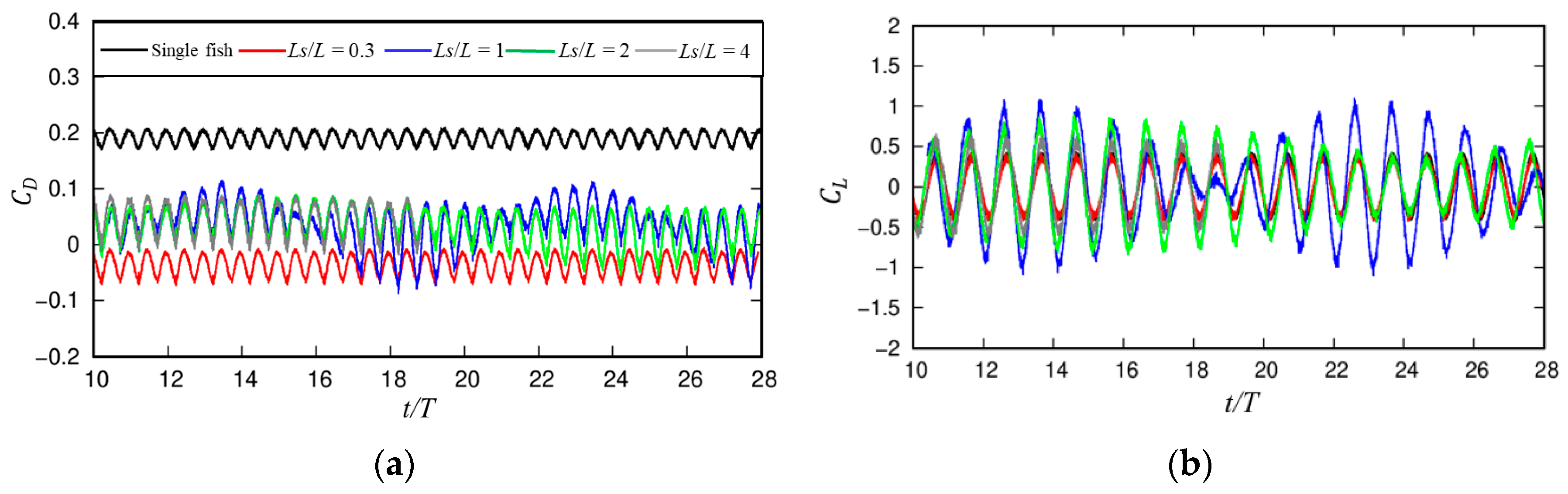
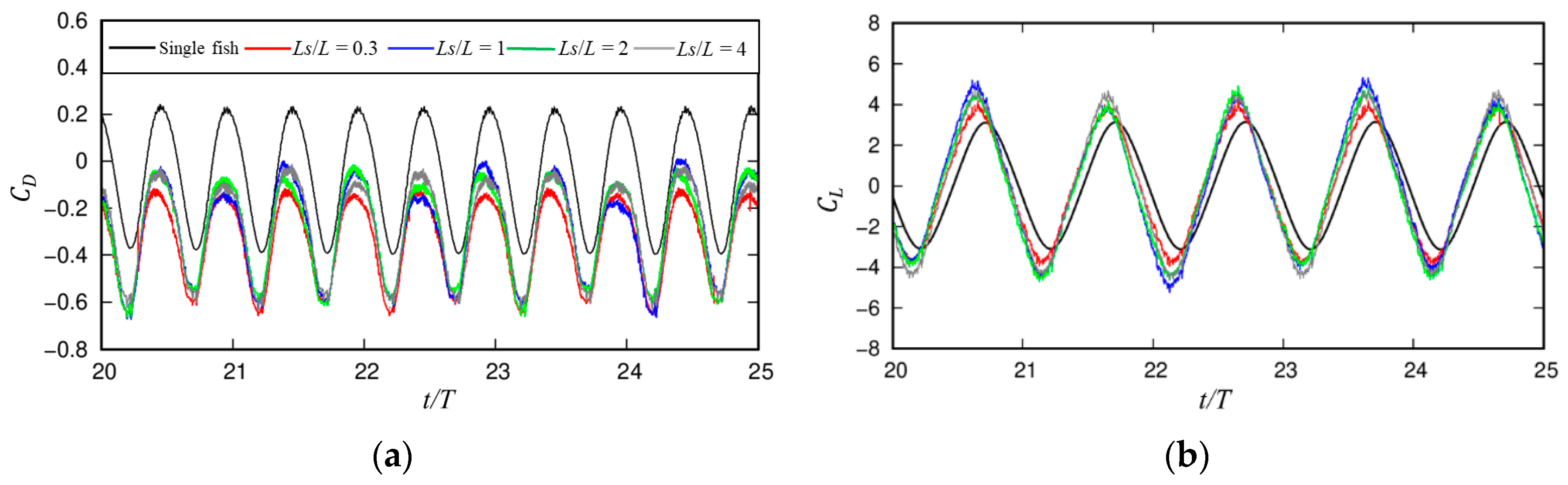
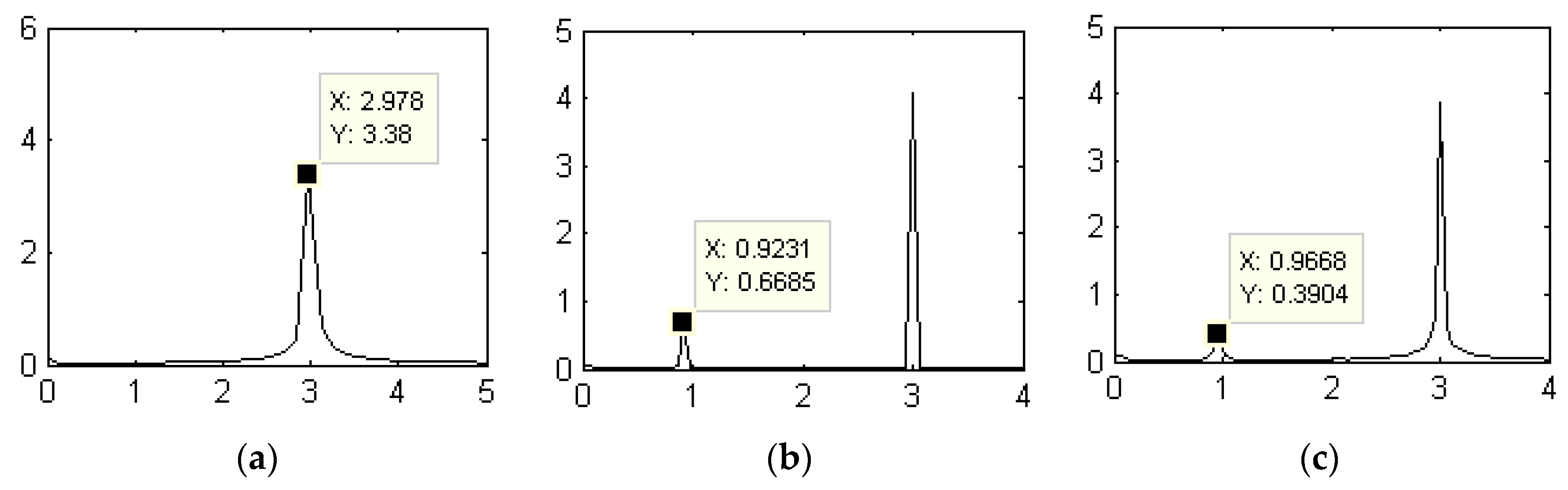
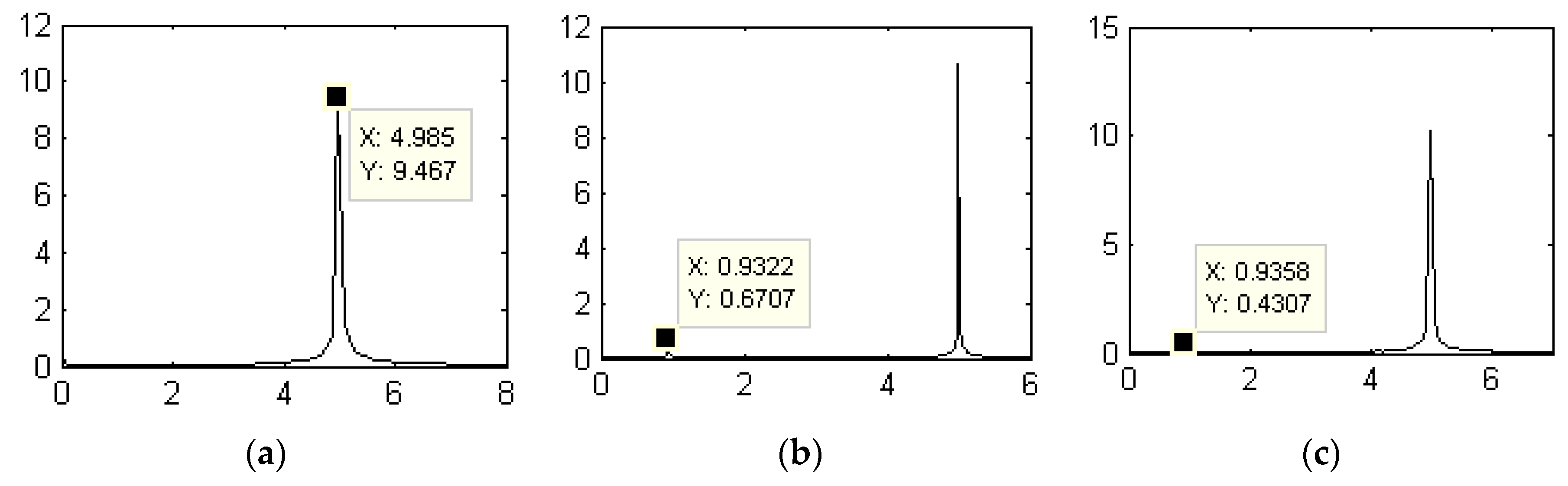
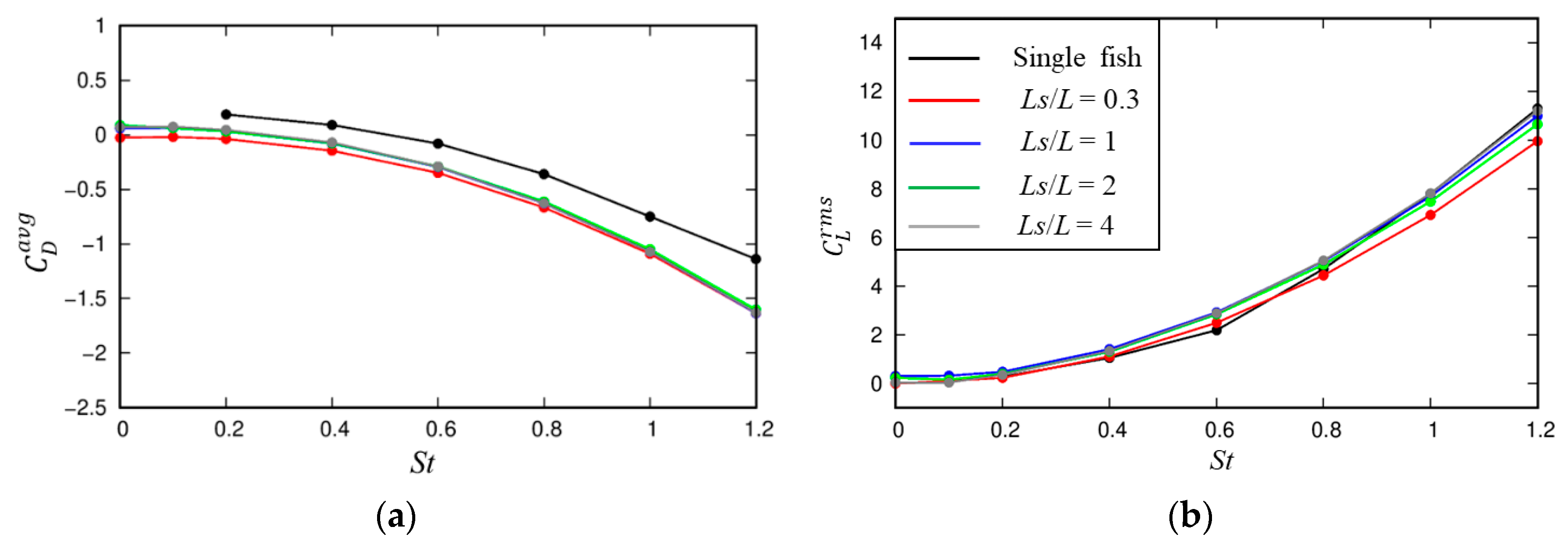
3.1.1. Dynamic Characteristics
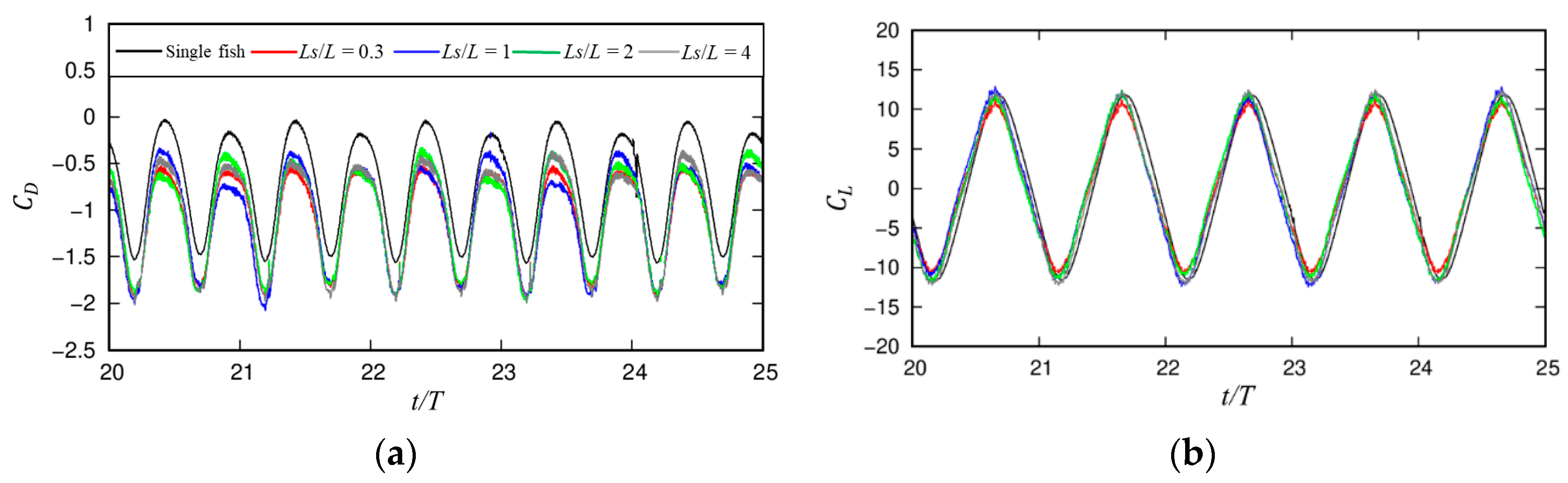
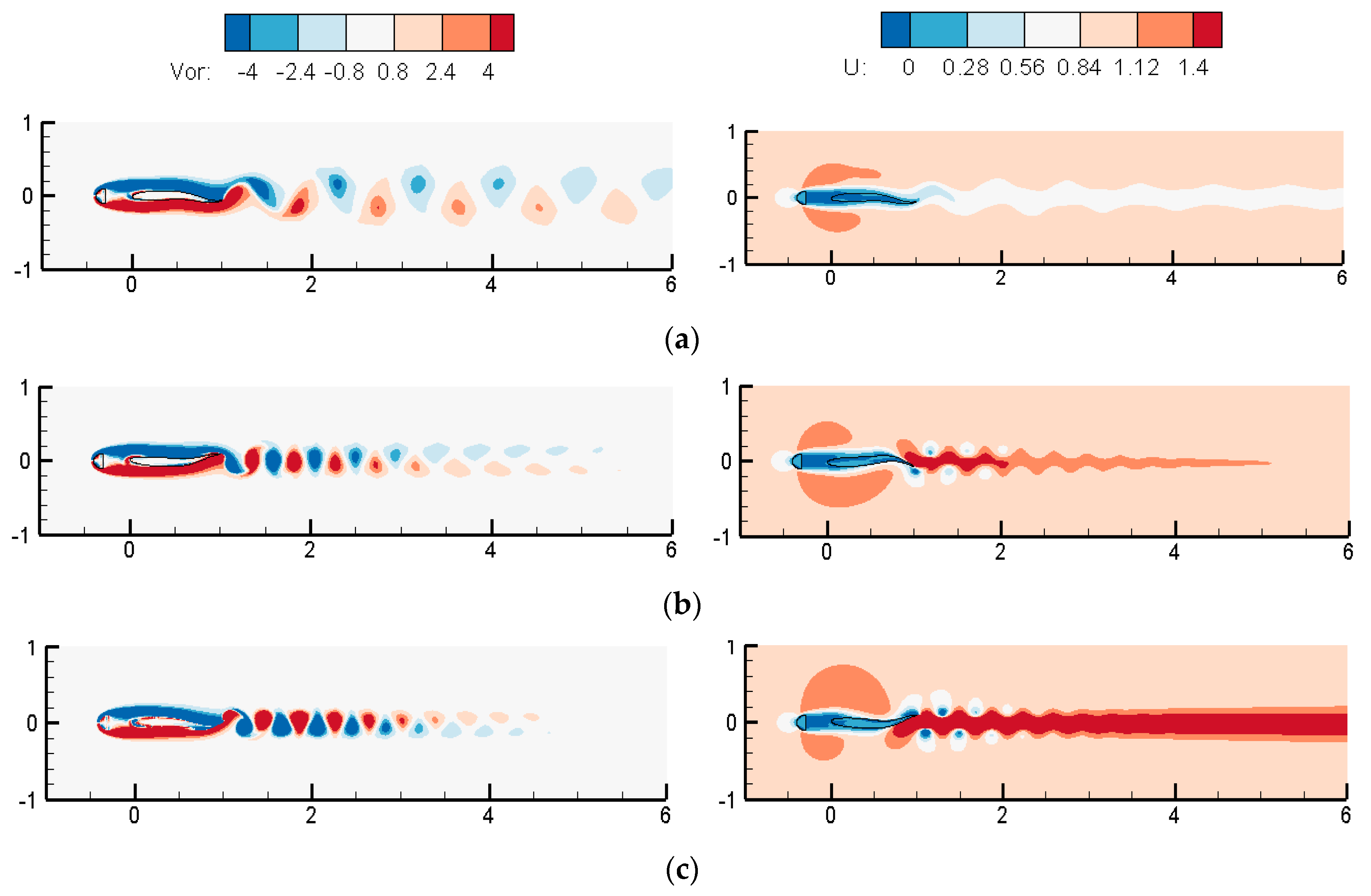
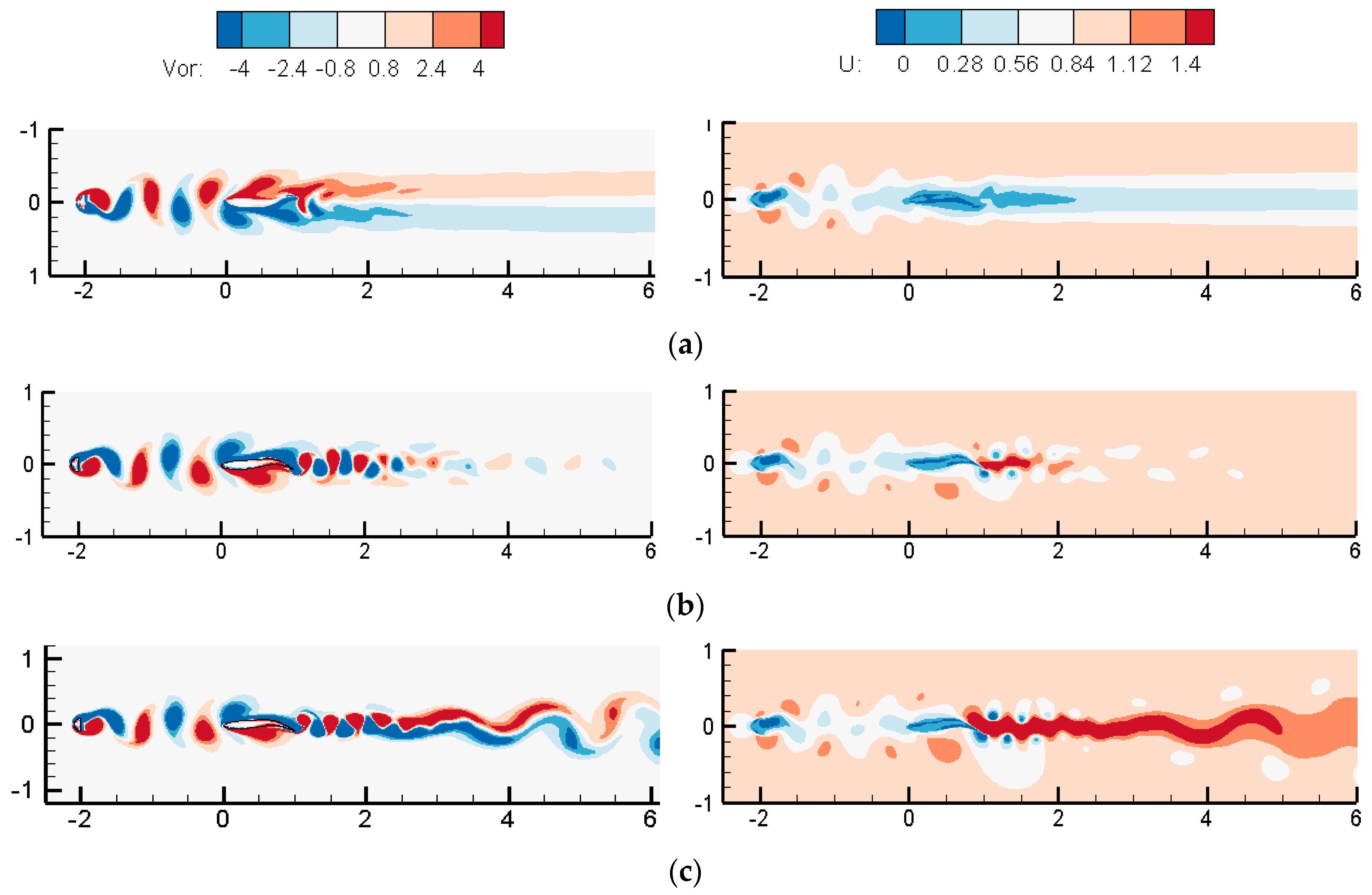
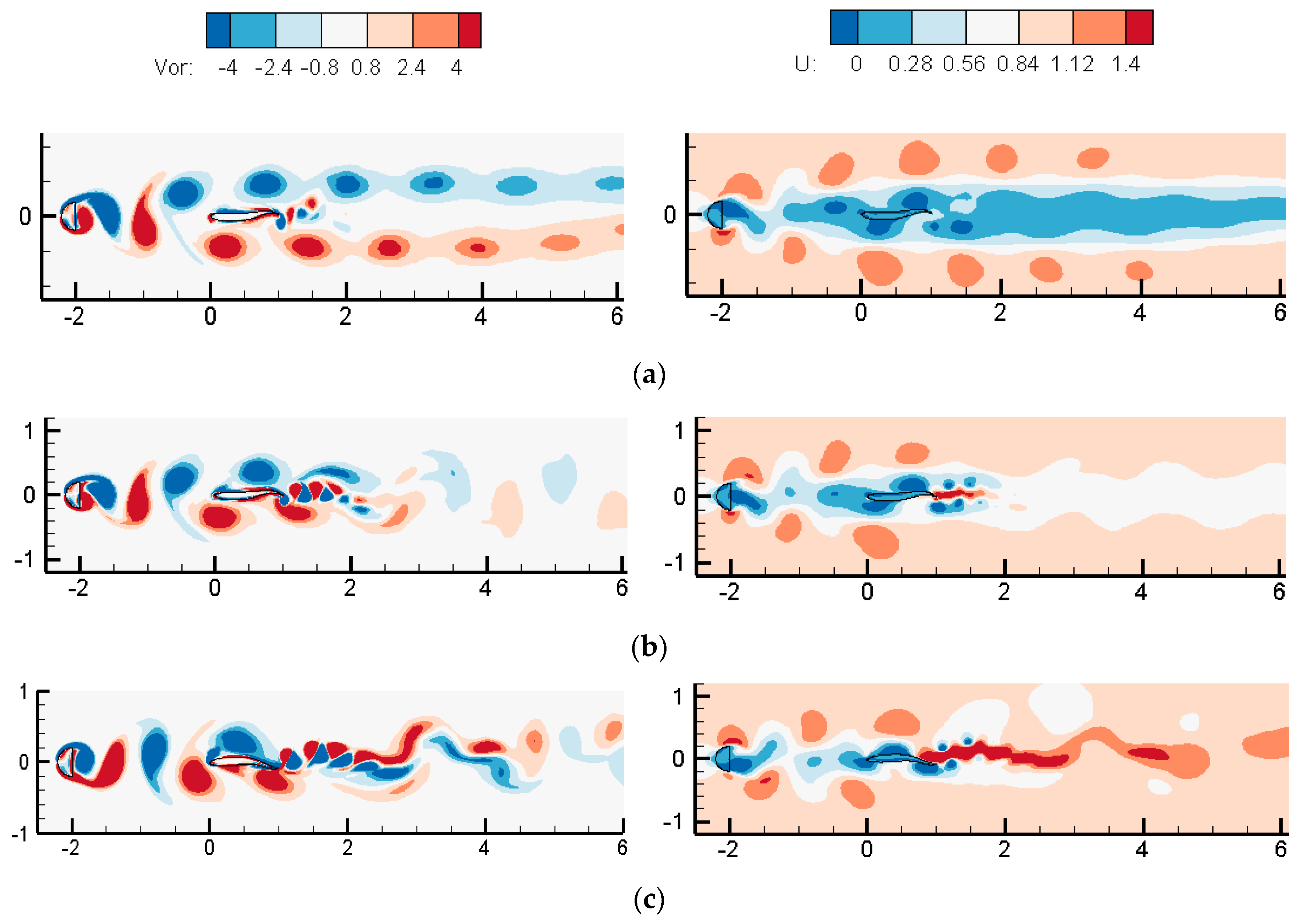
3.1.2. Wake Dynamics
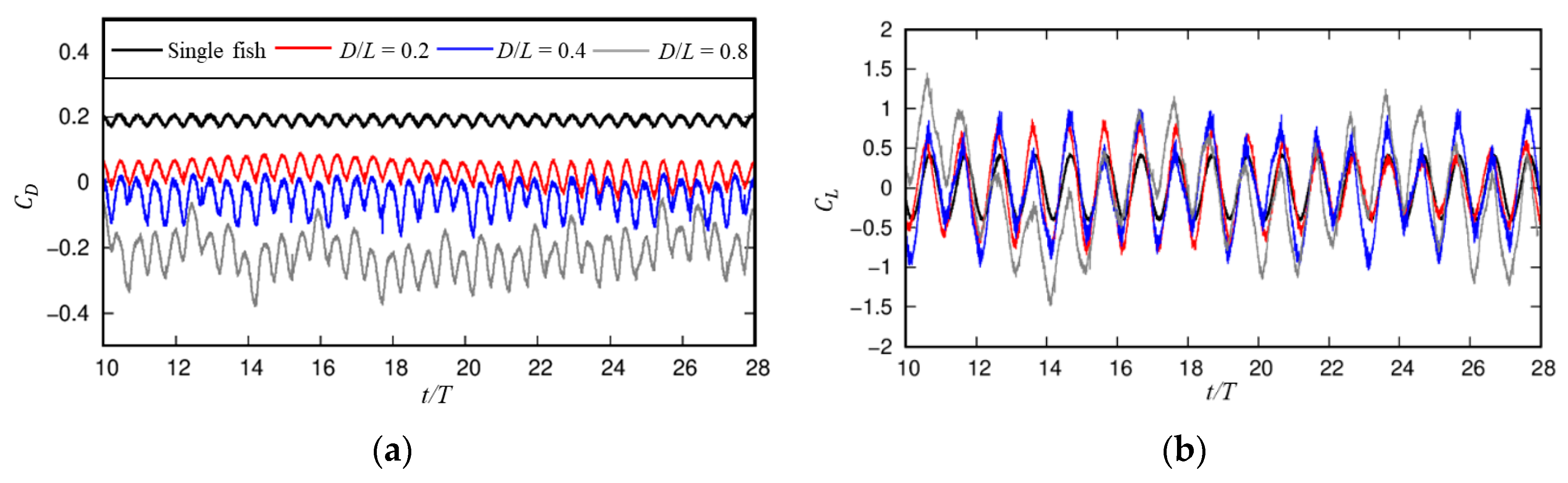
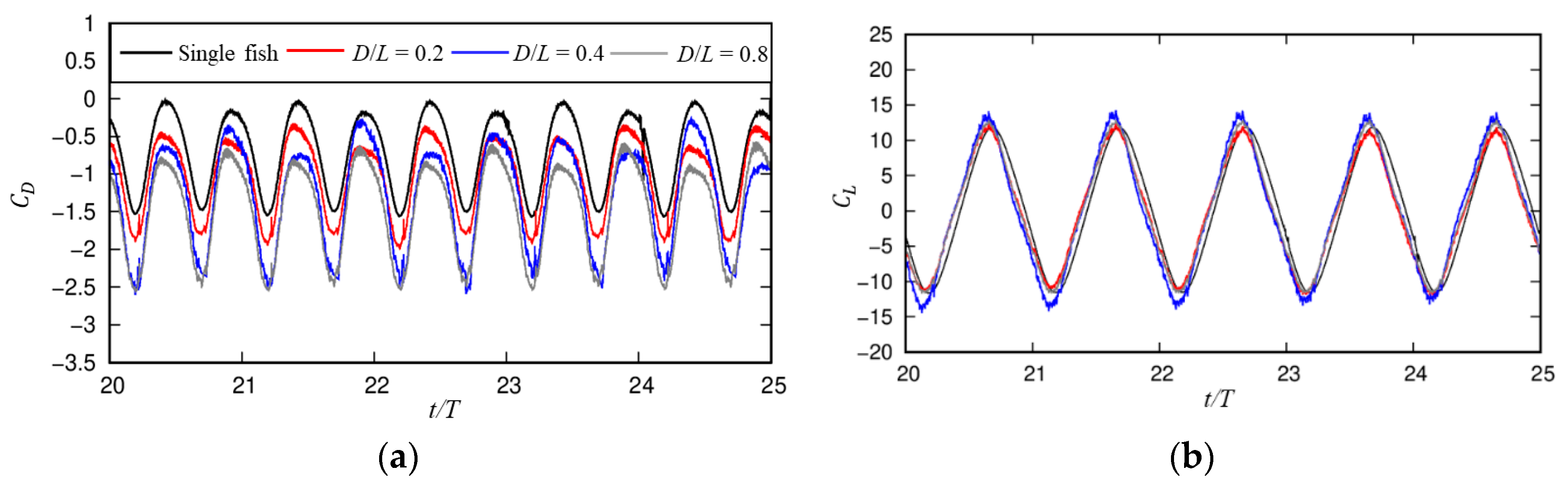
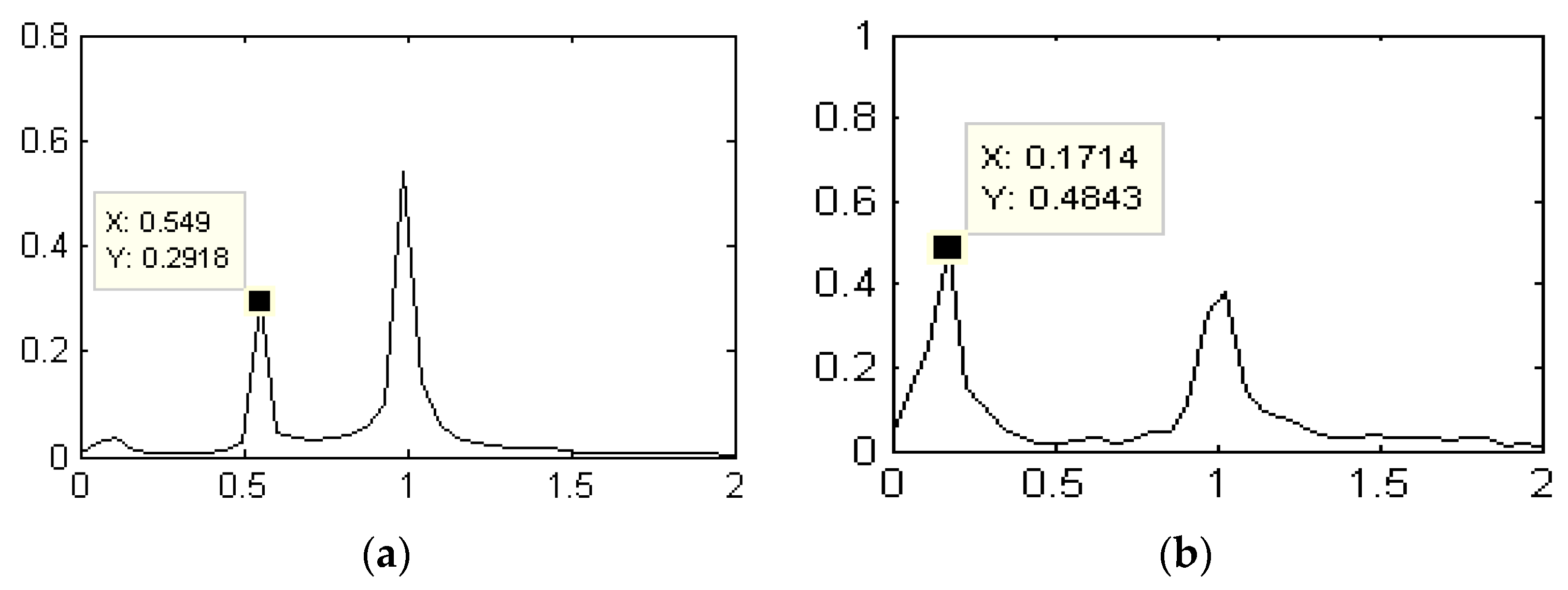
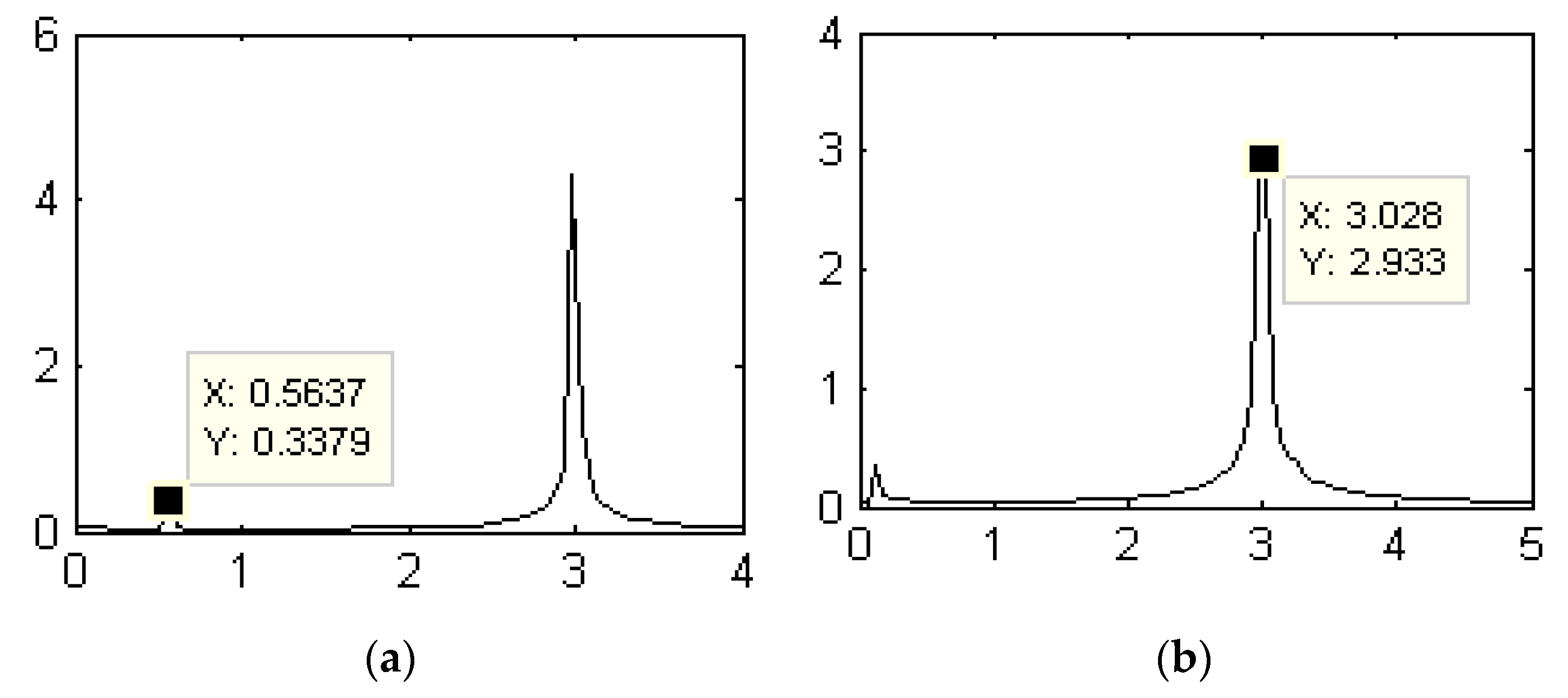
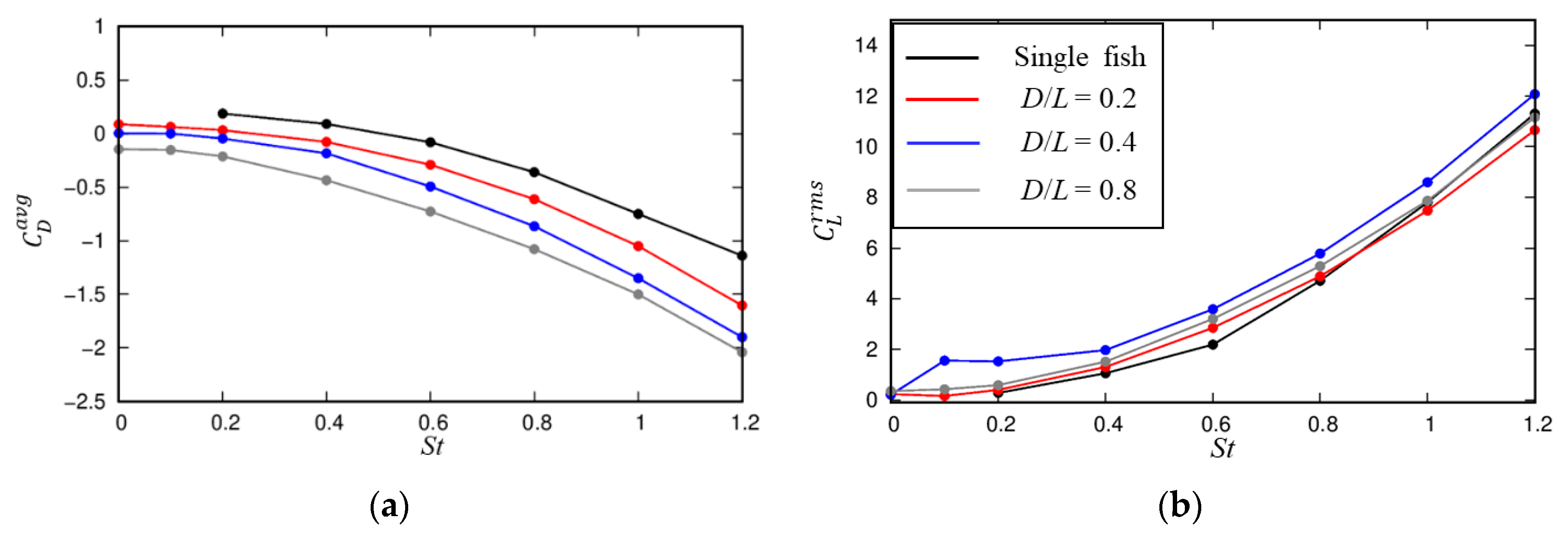
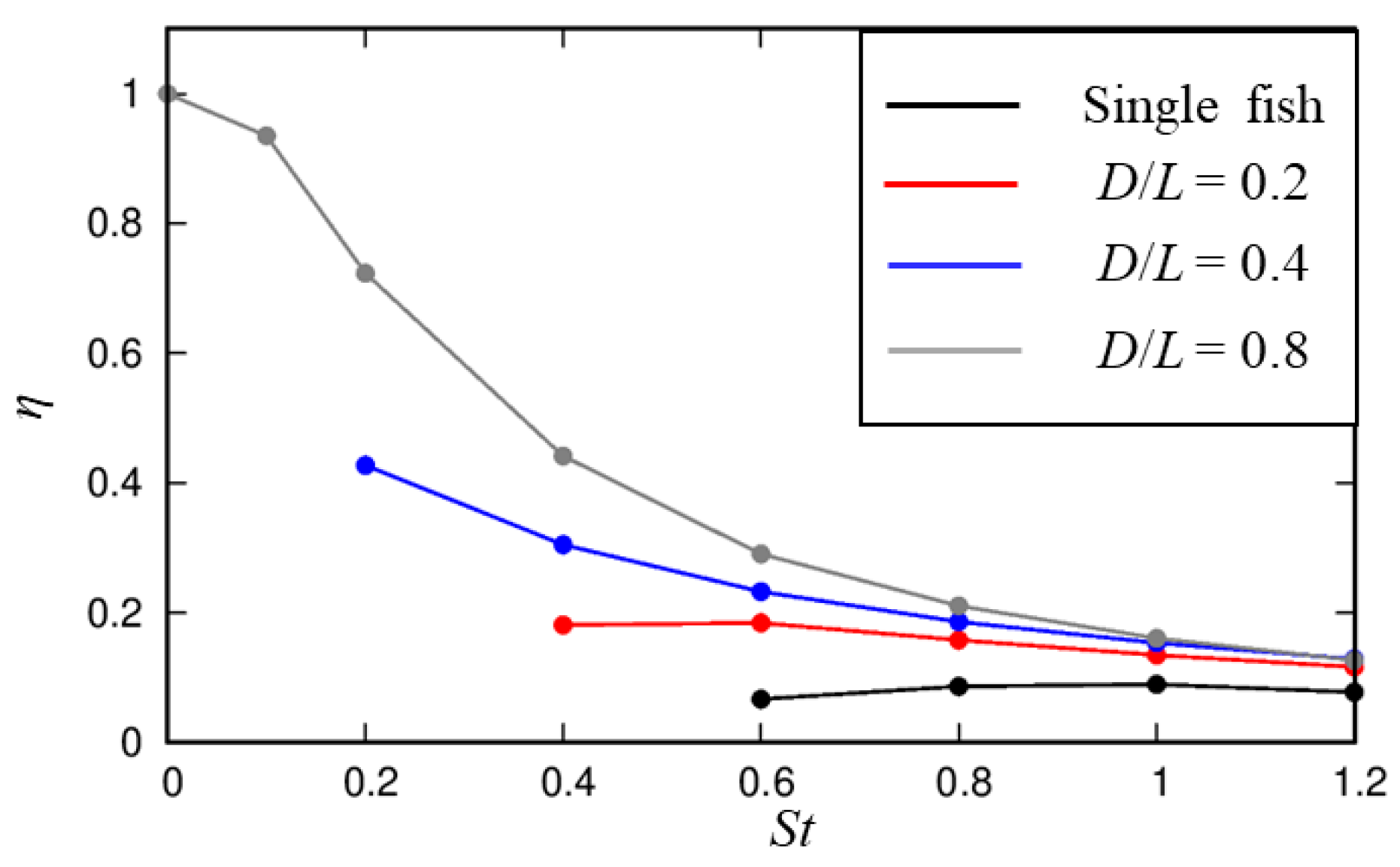
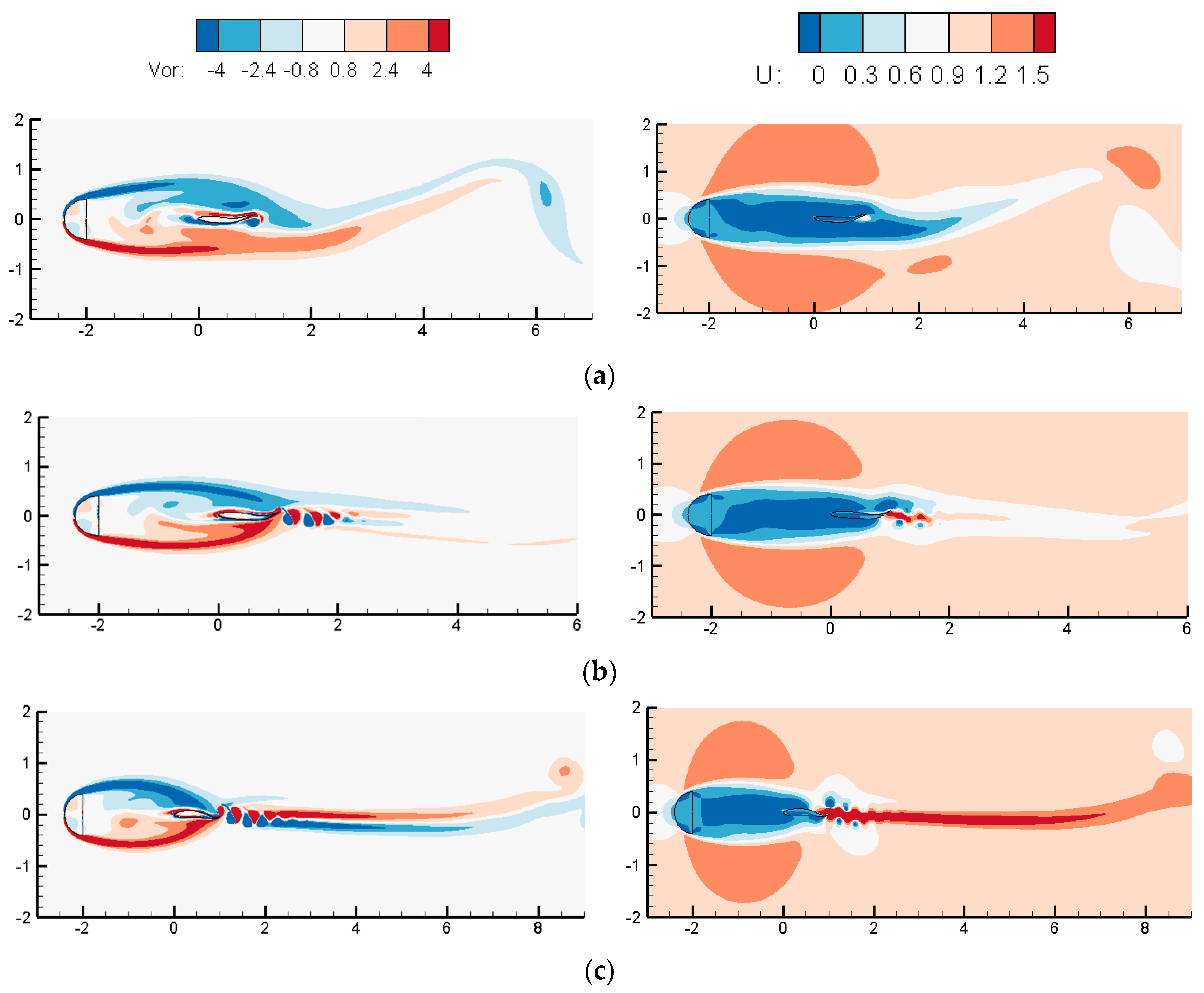
3.2. Influences of the Cylindrical Diameter
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Castro-Santos, T.; Goerig, E.; He, P.; Lauder, G.V. Applied Aspects of Locomotion and Biomechanics; Academic Press Fish Physiology: Cambridge, MA, USA, 2022; pp. 91–140. [Google Scholar]
- Lighthill, M.J. Large-amplitude elongated-body theory of fish locomotion. Proc. R. Soc. Lond. 1971, 179, 125–138. [Google Scholar]
- Wu, Y.T. Swimming of a waving plate. J. Fluid Mech. 1961, 10, 321–344. [Google Scholar] [CrossRef]
- Cheng, J.Y.; Zhuang, L.X.; Tong, B.G. Analysis of Swimming Three-dimensional Waving Plates. J. Fluid Mech. 1991, 232, 341–355. [Google Scholar] [CrossRef]
- Eloy, C. Optimal Strouhal number for swimming animals. J. Fluids Struct. 2011, 30, 205–218. [Google Scholar] [CrossRef]
- Wu, Y.T. Fish swimming and bird/insect flight. Annu. Rev. Fluid Mech. 2011, 43, 25–58. [Google Scholar] [CrossRef]
- Tu, H.; Wang, F.; Wang, H.; Gao, Q.; Wei, R. Experimental study on wake flows of a live fish with time-resolved tomographic PIV and pressure reconstruction. Exp. Fluids 2022, 63, 25. [Google Scholar] [CrossRef]
- Piskur, P.; Szymak, P.; Przybylski, M.; Naus, K.; Jaskólski, K.; Żokowski, M. Innovative energy-saving propulsion system for low-speed biomimetic underwater vehicles. Energies 2021, 14, 8418. [Google Scholar] [CrossRef]
- Piskur, P. Strouhal Number Measurement for Novel Biomimetic Folding Fins Using an Image Processing Method. J. Mar. Sci. Eng. 2022, 10, 484. [Google Scholar] [CrossRef]
- Wei, C.; Hu, Q.; Liu, Y.; Yin, S.; Ji, X. Performance evaluation and optimization for two-dimensional fish-like propulsion. Ocean Eng. 2021, 233, 109191. [Google Scholar] [CrossRef]
- Zhao, Z.; Dou, L. Effects of the structural relationships between the fish body and caudal fin on the propulsive performance of fish. Ocean Eng. 2019, 186, 106117. [Google Scholar] [CrossRef]
- Lauder, G.V.; Han, P. Hydrodynamics of median-fin interactions in fish-like locomotion: Effects of fin shape and movement. Phys. Fluids 2020, 32, 011902. [Google Scholar]
- Li, N.Y.; Zhuang, J.Y.; Zhu, Y.Z.; Su, G.S.; Yu, Y.M. Fluid dynamics of a self-propelled biomimetic underwater vehicle with pectoral fins. J. Ocean Eng. Sci. 2021, 6, 160–169. [Google Scholar] [CrossRef]
- Sun, P.N.; Colagrossi, A.; Zhang, A.M. Numerical simulation of the self-propulsive motion of a fishlike swimming foil using the δ+-sph model. Theor. Appl. Mech. Lett. 2018, 8, 115–125. [Google Scholar] [CrossRef]
- Verzicco, R. Immersed Boundary Methods: Historical Perspective and Future Outlook. Annu. Rev. Fluid Mech. 2023, 55, 129–155. [Google Scholar] [CrossRef]
- Huang, W.X.; Tian, F.B. Recent trends and progress in the immersed boundary method. J. Mech. Eng. Sci. 2019, 233, 095440621984260. [Google Scholar] [CrossRef]
- Wang, W.; Yan, Y.; Tian, F.B. Numerical study on hydrodynamics for a non-sinusoidal forced oscillating hydrofoil based on an immersed boundary method. Ocean Eng. 2017, 147, 606–620. [Google Scholar] [CrossRef]
- Ghommem, M.; Bourantas, G.C.; Wittek, A.; Miller, K.; Hajj, M.R. Hydrodynamic modeling and performance analysis of bio-inspired swimming. Ocean Eng. 2020, 197, 106897. [Google Scholar] [CrossRef]
- Romano, D.; Stefanini, C. Individual neon tetras (Paracheirodon innesi, myers) optimise their position in the group depending on external selective contexts: Lesson learned from a fish-robot hybrid school. Biosyst. Eng. 2021, 204, 170–180. [Google Scholar] [CrossRef]
- Deng, J.; Shao, X.M.; Yu, Z.S. Hydrodynamic studies on two traveling wavy foils in tandem arrangement. Phys. Fluids 2007, 19, 305–371. [Google Scholar] [CrossRef]
- Khalid, M.S.U.; Akhtar, I.; Dong, H. Hydrodynamics of a tandem fish school with asynchronous undulation of individuals. J. Fluids Struct. 2016, 66, 19–35. [Google Scholar] [CrossRef]
- Dai, L.Z.; He, G.W.; Zhang, X.; Zhang, X. Stable formations of self-propelled fish-like swimmers induced by hydrodynamic interactions. J. R. Soc. Interface 2018, 15, 20180490. [Google Scholar] [CrossRef] [PubMed]
- Kurt, M.; Mivehchi, A.; Moored, K.W. Two-dimensionally stable self-organization arises in simple schooling swimmers through hydrodynamic interactions. Physics 2021, 10, 14. [Google Scholar]
- Feng, Y.K.; Liu, H.X.; Su, Y.Y.; Su, Y.M. Numerical study on the hydrodynamics of c-turn maneuvering of a tuna-like fish body under self-propulsion. J. Fluid Struct. 2020, 94, 102954. [Google Scholar] [CrossRef]
- Hemelrijk, C.; Reid, D.; Hildenbrandt, H.; Padding, J. The increased efficiency of fish swimming in a school. Fish Fish. 2015, 16, 12072. [Google Scholar] [CrossRef]
- Liao, J.C. Fish exploiting vortices decrease muscle activity. Science 2003, 302, 1566–1569. [Google Scholar] [CrossRef] [PubMed]
- Liao, J.; Beal, D.; Lauder, G.; Triantafyllou, M. The Kármán gait: Novel body kinematics of rainbow trout swimming in a vortex street. J. Exp. Biol. 2003, 206, 1059–1073. [Google Scholar] [CrossRef] [PubMed]
- Beal, D.N.; Hover, F.S.; Triantafyllou, M.S.; Liao, J.C.; Lauder, G.V. Passive propulsion in vortex wakes. J. Fluid Mech. 2006, 549, 385–402. [Google Scholar] [CrossRef]
- Sahu, M.M.S. Numerical study of flow-induced vibration of a circular cylinder with attached flexible splitter plate at low re. J. Fluid Mech. 2019, 880, 551–593. [Google Scholar] [CrossRef]
- Mao, Q.; Zhao, J.; Liu, Y.; Sung, H.J. Drag reduction by a rotationally oscillating cylinder with a flexible filament. Phys. Fluids 2022, 34, 041910. [Google Scholar] [CrossRef]
- Sun, X.; Suh, C.S.; Ye, Z.H.; Yu, B. Dynamics of a circular cylinder with an attached splitter plate in laminar flow: A transition from vortex-induced vibration to galloping. Phys. Fluids 2020, 32, 027104. [Google Scholar] [CrossRef]
- Hu, L.C.; Liu, N.S.; Hang, D.; Lu, X.Y. Study on flapping modes of a flexible filament in the wake of an upstream rotating cylinder. Sci. Sin. Phys. Mech. Astron. 2018, 48, 094706. [Google Scholar] [CrossRef]
- Shi, F.L.; Xin, J.J.; Yin, P.D.; Dong, Y.H. A GPU accelerated three-dimensional ghost cell method with an improved implicit surface representation for complex rigid or flexible boundary flows. Comput. Phys. Commun. 2024, 298, 109098. [Google Scholar] [CrossRef]
- Xin, J.J.; Shi, F.L.; Jin, Q.; Lin, C. A radial basis function based ghost cell method with improved mass conservation for complex moving boundary flows. Comput. Fluids 2018, 176, 210–225. [Google Scholar] [CrossRef]
- Dong, G.J.; Lu, X.Y. Characteristics of flow over traveling wavy foils in a side-by-side arrangement. Phys. Fluids 2007, 19, 057107. [Google Scholar] [CrossRef]
- Paraz, F.; Schouveiler, L.; Eloy, C. Thrust generation by a heaving flexible foil: Resonance, nonlinearities, and optimality. Phys. Fluids 2016, 28, 011903. [Google Scholar] [CrossRef]
- Liao, J.C. Fish swimming efficiency. Curr. Biol. 2022, 32, R589–R683. [Google Scholar] [CrossRef]
- Borazjani, I.; Sotiropoulos, F. On the role of form and kinematics on the hydrodynamics of self-propelled body/caudal fin swimming. J. Exp. Biol. 2010, 213, 89–107. [Google Scholar] [CrossRef] [PubMed]
- Triantafyllou, M.S.; Triantafyllou, G.S.; Yue, D.K.P. Hydrodynamics of fishlike swimming. Ann. Rev. Fluid Mech. 2000, 32, 33–53. [Google Scholar] [CrossRef]
- Liu, N.; Peng, Y.L.; Liang, Y.; Lu, X. Flow over a traveling wavy foil with a passively flapping flat plate. Phys. Rev. E 2012, 85, 056316. [Google Scholar] [CrossRef]
- Wang, H.L.; Zheng, X.B.; Pröbsting, S.; Hu, C.H.; Wang, Q. An unsteady RANS simulation of the performance of an oscillating hydrofoil at a high Reynolds number. Ocean Eng. 2023, 274, 114097. [Google Scholar] [CrossRef]
- Ma, D.C.; Qiu, Z.; Li, G.H.; Wang, F.X. Numerical investigation on lag mechanism of drag-to-thrust transition for a pitching airfoil. Acta Aerodyn. Sin. 2021, 39, 91–103. [Google Scholar]
- Zhu, B.; Wang, F.L.; Xu, W.J.; Xiao, Q.S. Mode transition mechanisms in semi-passive flapping-wing propulsion or energy extraction. Phys. Fluids 2023, 35, 123604. [Google Scholar] [CrossRef]
- Zheng, Z.C.; Wei, Z. Study of mechanisms and factors that influence the formation of vortical wake of a heaving airfoil. Phys. Fluids 2012, 24, 103601. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Grid Notation | Grid Number | |||
|---|---|---|---|---|
| G1 | 250 × 100 | 0.006, 0.004 | 1.49 | 0.101 |
| G2 | 375 × 150 | 0.004, 0.00267 | 1.66 | 0.103 |
| G3 | 562 × 225 | 0.00267, 0.00178 | 1.68 | 0.103 |
| Test Condition | Diameter | Gap Distance | Strouhal Number |
|---|---|---|---|
| Case #1 | - | - | 0–1.2 |
| Case #2 | D/L = 0.2 | Ls/L = 0.3 | 0–1.2 |
| Case #3 | D/L = 0.2 | Ls/L = 1 | 0–1.2 |
| Case #4 | D/L = 0.2 | Ls/L = 2 | 0–1.2 |
| Case #5 | D/L = 0.2 | Ls/L = 4 | 0–1.2 |
| Case #6 | D/L = 0.4 | Ls/L = 2 | 0–1.2 |
| Case #7 | D/L = 0.8 | Ls/L = 2 | 0–1.2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chang, X.; Ma, B.; Xin, J. Numerical Studies on the Hydrodynamic Patterns and Energy-Saving Advantages of Fish Swimming in Vortical Flows of an Upstream Cylinder. J. Mar. Sci. Eng. 2024, 12, 1254. https://doi.org/10.3390/jmse12081254
Chang X, Ma B, Xin J. Numerical Studies on the Hydrodynamic Patterns and Energy-Saving Advantages of Fish Swimming in Vortical Flows of an Upstream Cylinder. Journal of Marine Science and Engineering. 2024; 12(8):1254. https://doi.org/10.3390/jmse12081254
Chicago/Turabian StyleChang, Xing, Bowen Ma, and Jianjian Xin. 2024. "Numerical Studies on the Hydrodynamic Patterns and Energy-Saving Advantages of Fish Swimming in Vortical Flows of an Upstream Cylinder" Journal of Marine Science and Engineering 12, no. 8: 1254. https://doi.org/10.3390/jmse12081254