
Osteoclast-Driven Polydopamine-to-Dopamine Release: An Upgrade Patch for Polydopamine-Functionalized Tissue Engineering Scaffolds

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
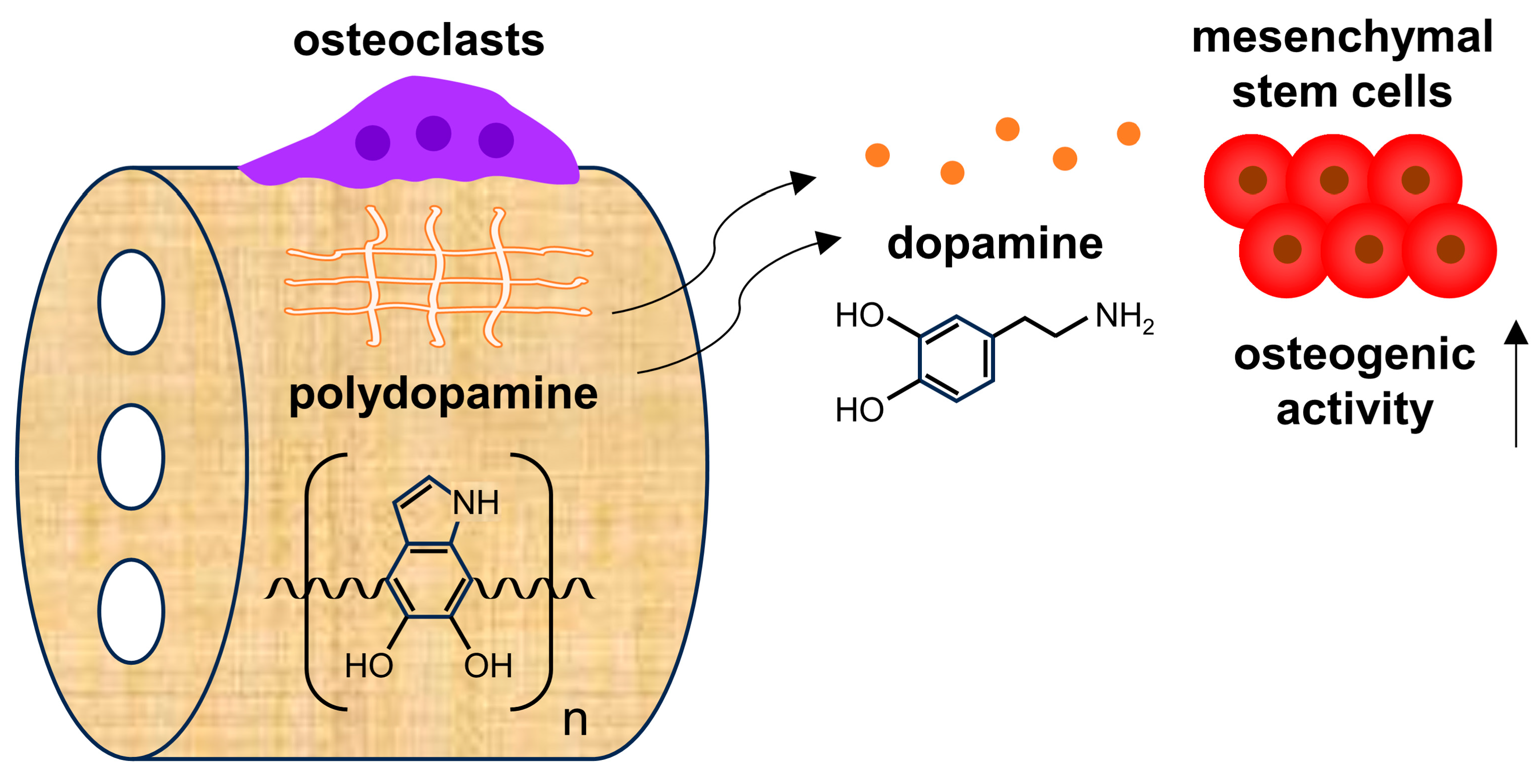
:1. Introduction
2. Materials and Methods
2.1. Fabrication of PDHC Material
2.2. OC Differentiation
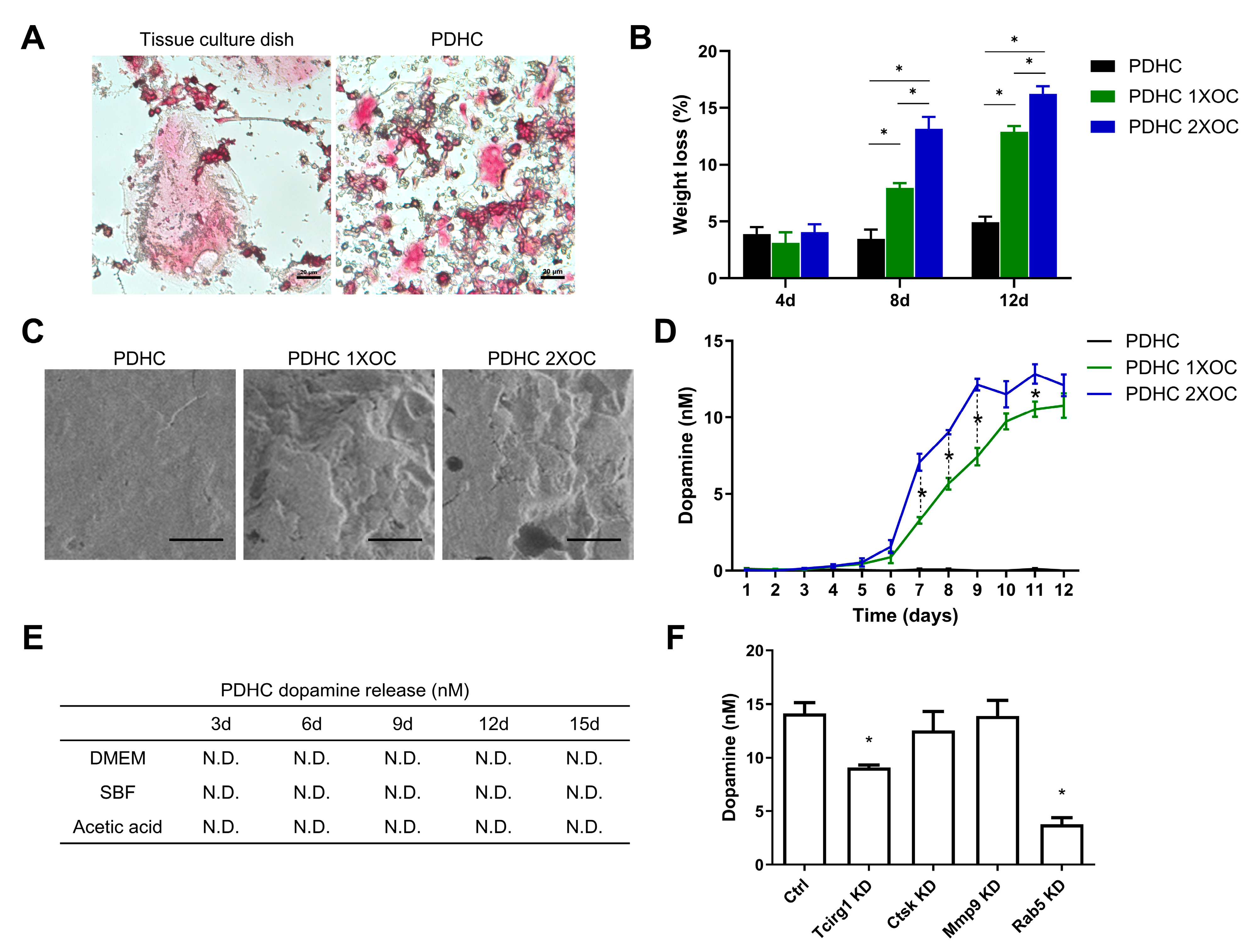
2.3. Assessment of Osteoclastic Degradation of PDHC Material
2.4. Dopamine Concentration Measurement
2.5. Gene Knockdown
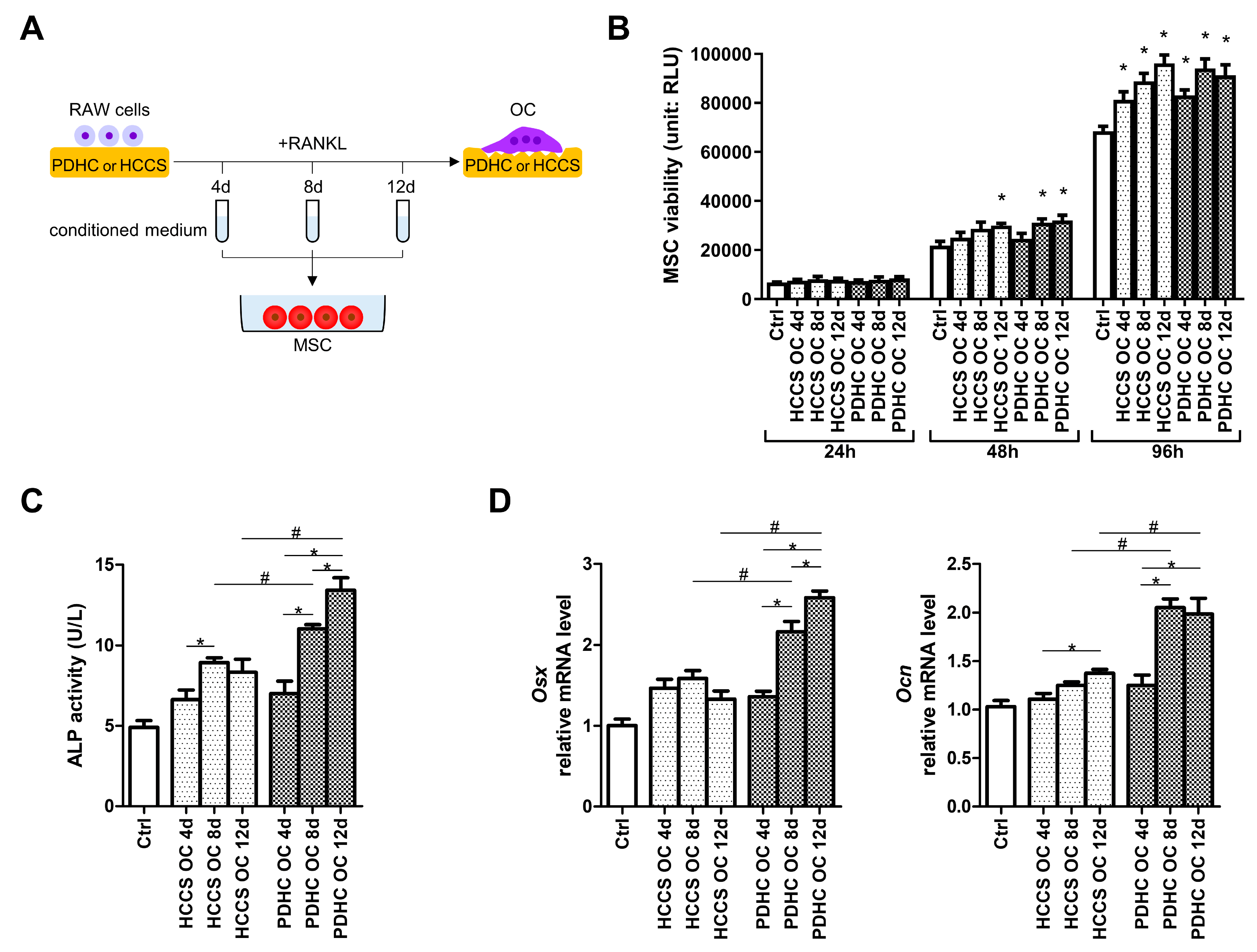
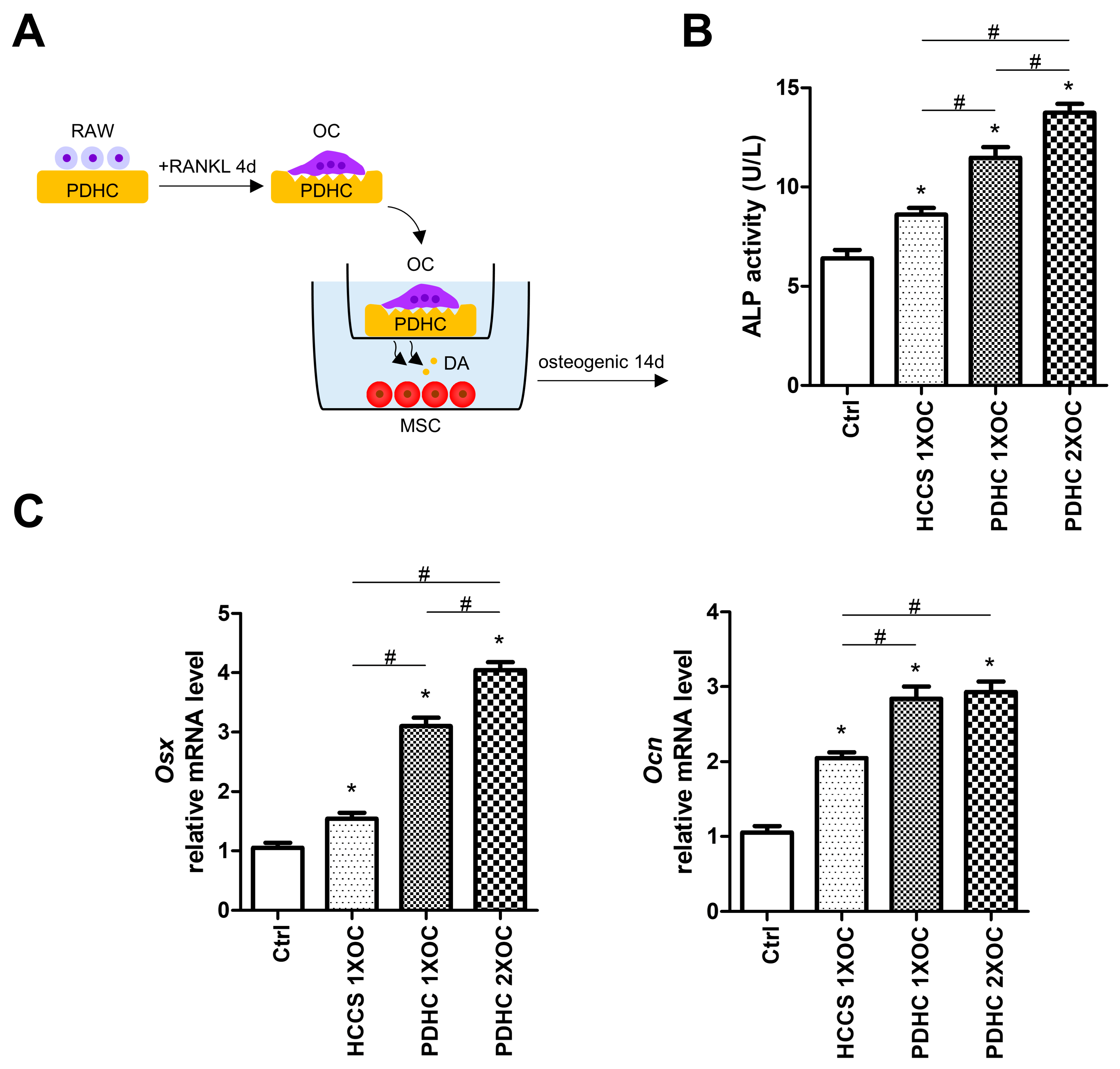
2.6. MSC Culture and Osteogenesis Evaluation
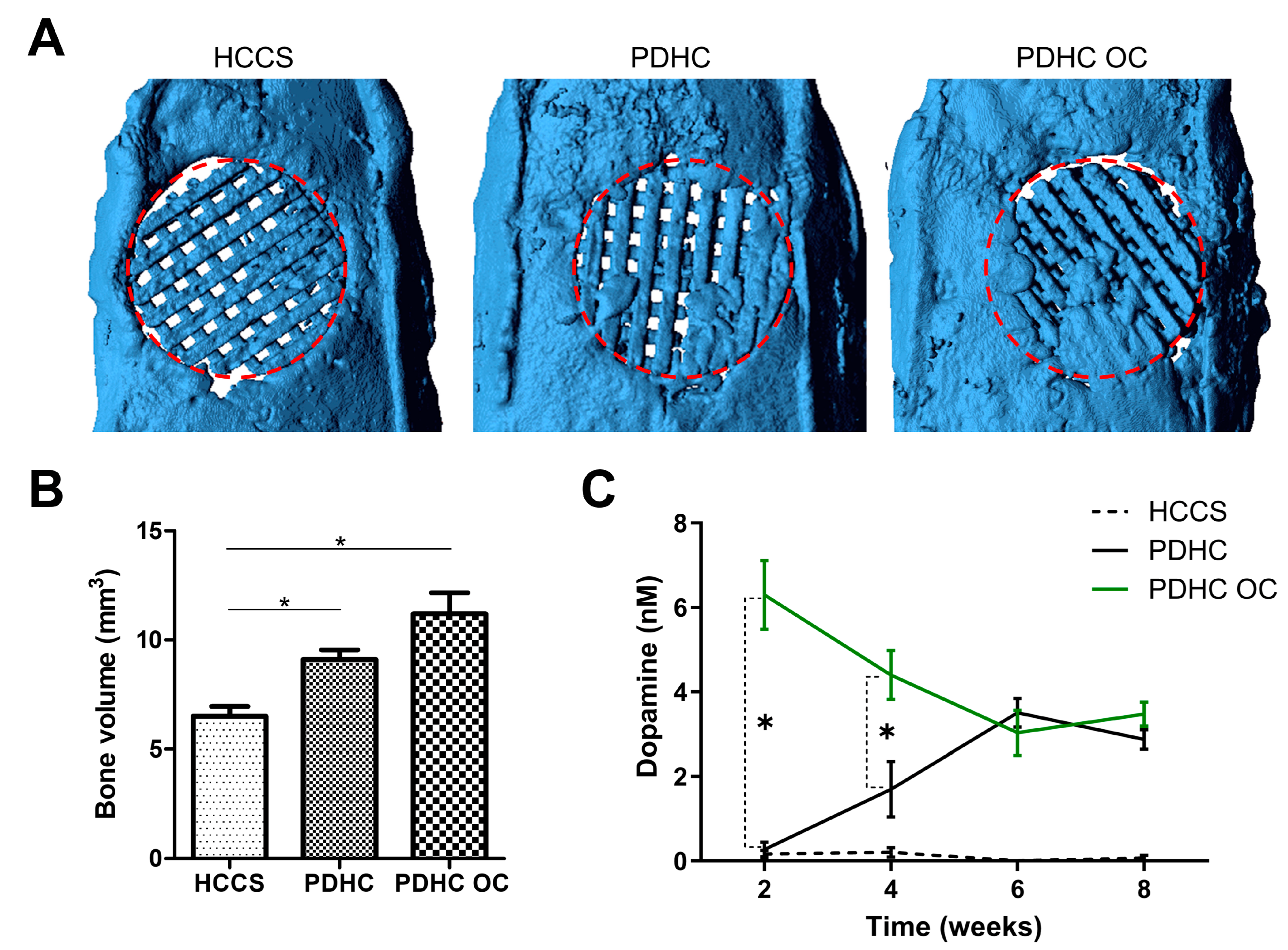
2.7. Calvarial Critical-Sized Defect Model
2.8. MicroCT Analysis
2.9. Statistical Analyses
3. Results and Discussion
3.1. PDHC Releases Dopamine Specifically upon Osteoclastic Resorption
3.2. PDHC-OC Interactions Release Dopamine That Shows Osteoinductive Activity
3.3. Manipulating Dopamine Release via Manipulating OC Number Influences PDHC’s Osteoinductive Effects
3.4. PDHC Provides a Sustained Release of Dopamine That Facilitates Calvaria Defect Repair
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Roddy, E.; DeBaun, M.R.; Daoud-Gray, A.; Yang, Y.P.; Gardner, M.J. Treatment of critical-sized bone defects: Clinical and tissue engineering perspectives. Eur. J. Orthop. Surg. Traumatol. 2018, 28, 351–362. [Google Scholar] [CrossRef] [PubMed]
- Bernstein, M.; Little, M.T.M.; Marecek, G. Current Management of Acute and Posttraumatic Critical Bone Defects. J. Orthop. Trauma 2024, 38, S1–S8. [Google Scholar] [CrossRef] [PubMed]
- Wong, S.K.; Yee, M.M.F.; Chin, K.Y.; Ima-Nirwana, S. A Review of the Application of Natural and Synthetic Scaffolds in Bone Regeneration. J. Funct. Biomater. 2023, 14, 286. [Google Scholar] [CrossRef] [PubMed]
- Ansari, M.A.A.; Golebiowska, A.A.; Dash, M.; Kumar, P.; Jain, P.K.; Nukavarapu, S.P.; Ramakrishna, S.; Nanda, H.S. Engineering biomaterials to 3D-print scaffolds for bone regeneration: Practical and theoretical consideration. Biomater. Sci. 2022, 10, 2789–2816. [Google Scholar] [CrossRef] [PubMed]
- Tolabi, H.; Bakhtiary, N.; Sayadi, S.; Tamaddon, M.; Ghorbani, F.; Boccaccini, A.R.; Liu, C. A critical review on polydopamine surface-modified scaffolds in musculoskeletal regeneration. Front. Bioeng. Biotechnol. 2022, 10, 1008360. [Google Scholar] [CrossRef] [PubMed]
- Barros, N.R.; Chen, Y.; Hosseini, V.; Wang, W.; Nasiri, R.; Mahmoodi, M.; Yalcintas, E.P.; Haghniaz, R.; Mecwan, M.M.; Karamikamkar, S.; et al. Recent developments in mussel-inspired materials for biomedical applications. Biomater. Sci. 2021, 9, 6653–6672. [Google Scholar] [CrossRef]
- Qi, J.; Wang, Y.; Chen, L.; Chen, L.; Wen, F.; Huang, L.; Rueben, P.; Zhang, C.; Li, H. 3D-printed porous functional composite scaffolds with polydopamine decoration for bone regeneration. Regen. Biomater. 2023, 10, rbad062. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Liu, A.; Fan, J.; Wang, M.; Dai, J.; Jin, X.; Deng, H.; Wang, X.; Liang, Y.; Li, H.; et al. A drug-loaded composite coating to improve osteogenic and antibacterial properties of Zn-1Mg porous scaffolds as biodegradable bone implants. Bioact. Mater. 2023, 27, 488–504. [Google Scholar] [CrossRef] [PubMed]
- Du, J.; Zhou, Y.; Bao, X.; Kang, Z.; Huang, J.; Xu, G.; Yi, C.; Li, D. Surface polydopamine modification of bone defect repair materials: Characteristics and applications. Front. Bioeng. Biotechnol. 2022, 10, 974533. [Google Scholar] [CrossRef]
- Shi, W.; Gao, Y.; Wu, Y.; Sun, J.; Xu, B.; Lu, X.; Wang, Q. A multifunctional polydopamine/genipin/alendronate nanoparticle licences fibrin hydrogels osteoinductive and immunomodulatory potencies for repairing bone defects. Int. J. Biol. Macromol. 2023, 249, 126072. [Google Scholar] [CrossRef]
- Yao, M.; Zou, Q.; Zou, W.; Xie, Z.; Li, Z.; Zhao, X.; Du, C. Bifunctional scaffolds of hydroxyapatite/poly(dopamine)/carboxymethyl chitosan with osteogenesis and anti-osteosarcoma effect. Biomater. Sci. 2021, 9, 3319–3333. [Google Scholar] [CrossRef]
- Sun, X.; Jiao, X.; Wang, Z.; Ma, J.; Wang, T.; Zhu, D.; Li, H.; Tang, L.; Li, H.; Wang, C.; et al. Polydopamine-coated 3D-printed beta-tricalcium phosphate scaffolds to promote the adhesion and osteogenesis of BMSCs for bone-defect repair: mRNA transcriptomic sequencing analysis. J. Mater. Chem. B 2023, 11, 1725–1738. [Google Scholar] [CrossRef] [PubMed]
- Chang, M.C.; Ko, C.C.; Douglas, W.H. Preparation of hydroxyapatite-gelatin nanocomposite. Biomaterials 2003, 24, 2853–2862. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.J.; Lee, Y.T.; Zou, R.; Daniel, R.; Ko, C.C. Polydopamine-Laced Biomimetic Material Stimulation of Bone Marrow Derived Mesenchymal Stem Cells to Promote Osteogenic Effects. Sci. Rep. 2017, 7, 12984. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Wu, T.H.; Hu, X.; Liu, J.; Wu, D.; Miguez, P.A.; Wright, J.T.; Zhang, S.; Chi, J.T.; Tseng, H.C.; et al. Biomimetic polydopamine-laced hydroxyapatite collagen material orients osteoclast behavior to an anti-resorptive pattern without compromising osteoclasts' coupling to osteoblasts. Biomater. Sci. 2021, 9, 7565–7574. [Google Scholar] [CrossRef] [PubMed]
- Detsch, R.; Boccaccini, A.R. The role of osteoclasts in bone tissue engineering. J. Tissue Eng. Regen. Med. 2015, 9, 1133–1149. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.; Sudhakara, P.; Singh, J.; Ilyas, R.A.; Asyraf, M.R.M.; Razman, M.R. Critical Review of Biodegradable and Bioactive Polymer Composites for Bone Tissue Engineering and Drug Delivery Applications. Polymers 2021, 13, 2623. [Google Scholar] [CrossRef] [PubMed]
- de Araujo, J.C.R.; Sobral Silva, L.A.; de Barros Lima, V.A.; Bastos Campos, T.M.; Lisboa Filho, P.N.; Okamoto, R.; de Vasconcellos, L.M.R. The Local Release of Teriparatide Incorporated in 45S5 Bioglass Promotes a Beneficial Effect on Osteogenic Cells and Bone Repair in Calvarial Defects in Ovariectomized Rats. J. Funct. Biomater. 2023, 14, 93. [Google Scholar] [CrossRef] [PubMed]
- Pu, Y.; Lin, X.; Zhi, Q.; Qiao, S.; Yu, C. Microporous Implants Modified by Bifunctional Hydrogel with Antibacterial and Osteogenic Properties Promote Bone Integration in Infected Bone Defects. J. Funct. Biomater. 2023, 14, 226. [Google Scholar] [CrossRef]
- Veis, D.J.; O'Brien, C.A. Osteoclasts, Master Sculptors of Bone. Annu. Rev. Pathol. 2023, 18, 257–281. [Google Scholar] [CrossRef]
- Lee, D.J.; Tseng, H.C.; Wong, S.W.; Wang, Z.; Deng, M.; Ko, C.C. Dopaminergic effects on in vitro osteogenesis. Bone Res. 2015, 3, 15020. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.X.; Ge, X.Y.; Wang, M.Y.; Ma, T.; Zhang, Y.; Lin, Y. Dopamine D1 receptor-mediated activation of the ERK signaling pathway is involved in the osteogenic differentiation of bone mesenchymal stem cells. Stem Cell Res. Ther. 2020, 11, 12. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Feng, C.; Zhang, W.; Wang, Z.; Zhong, M.; Tang, W.; Wang, Z.; Shi, H.; Yin, Z.; Shi, J.; et al. Activation of dopamine receptor D1 promotes osteogenic differentiation and reduces glucocorticoid-induced bone loss by upregulating the ERK1/2 signaling pathway. Mol. Med. 2022, 28, 23. [Google Scholar] [CrossRef]
- Sims, N.A.; Martin, T.J. Osteoclasts Provide Coupling Signals to Osteoblast Lineage Cells Through Multiple Mechanisms. Annu. Rev. Physiol. 2020, 82, 507–529. [Google Scholar] [CrossRef] [PubMed]
- Daponte, V.; Henke, K.; Drissi, H. Current perspectives on the multiple roles of osteoclasts: Mechanisms of osteoclast-osteoblast communication and potential clinical implications. Elife 2024, 13, e95083. [Google Scholar] [CrossRef]
- Zhang, Y.; Polman, M.; Mohammad, A.F.; Hermens, I.; Zhuang, Z.; Wang, H.; van den Beucken, J.J. Species-independent stimulation of osteogenic differentiation induced by osteoclasts. Biochem. Biophys. Res. Commun. 2022, 606, 149–155. [Google Scholar] [CrossRef]
- Einhorn, T.A.; Gerstenfeld, L.C. Fracture healing: Mechanisms and interventions. Nat. Rev. Rheumatol. 2015, 11, 45–54. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, L.; Hu, H.; Ko, C.-C. Osteoclast-Driven Polydopamine-to-Dopamine Release: An Upgrade Patch for Polydopamine-Functionalized Tissue Engineering Scaffolds. J. Funct. Biomater. 2024, 15, 211. https://doi.org/10.3390/jfb15080211
Wang L, Hu H, Ko C-C. Osteoclast-Driven Polydopamine-to-Dopamine Release: An Upgrade Patch for Polydopamine-Functionalized Tissue Engineering Scaffolds. Journal of Functional Biomaterials. 2024; 15(8):211. https://doi.org/10.3390/jfb15080211
Chicago/Turabian StyleWang, Lufei, Huamin Hu, and Ching-Chang Ko. 2024. "Osteoclast-Driven Polydopamine-to-Dopamine Release: An Upgrade Patch for Polydopamine-Functionalized Tissue Engineering Scaffolds" Journal of Functional Biomaterials 15, no. 8: 211. https://doi.org/10.3390/jfb15080211