
Osteogenic Protection against Fine Dust with Erucic Acid-Induced Exosomes

Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Cell Viability Test
2.3. Evaluation of the Concentration of Induced Exosomes
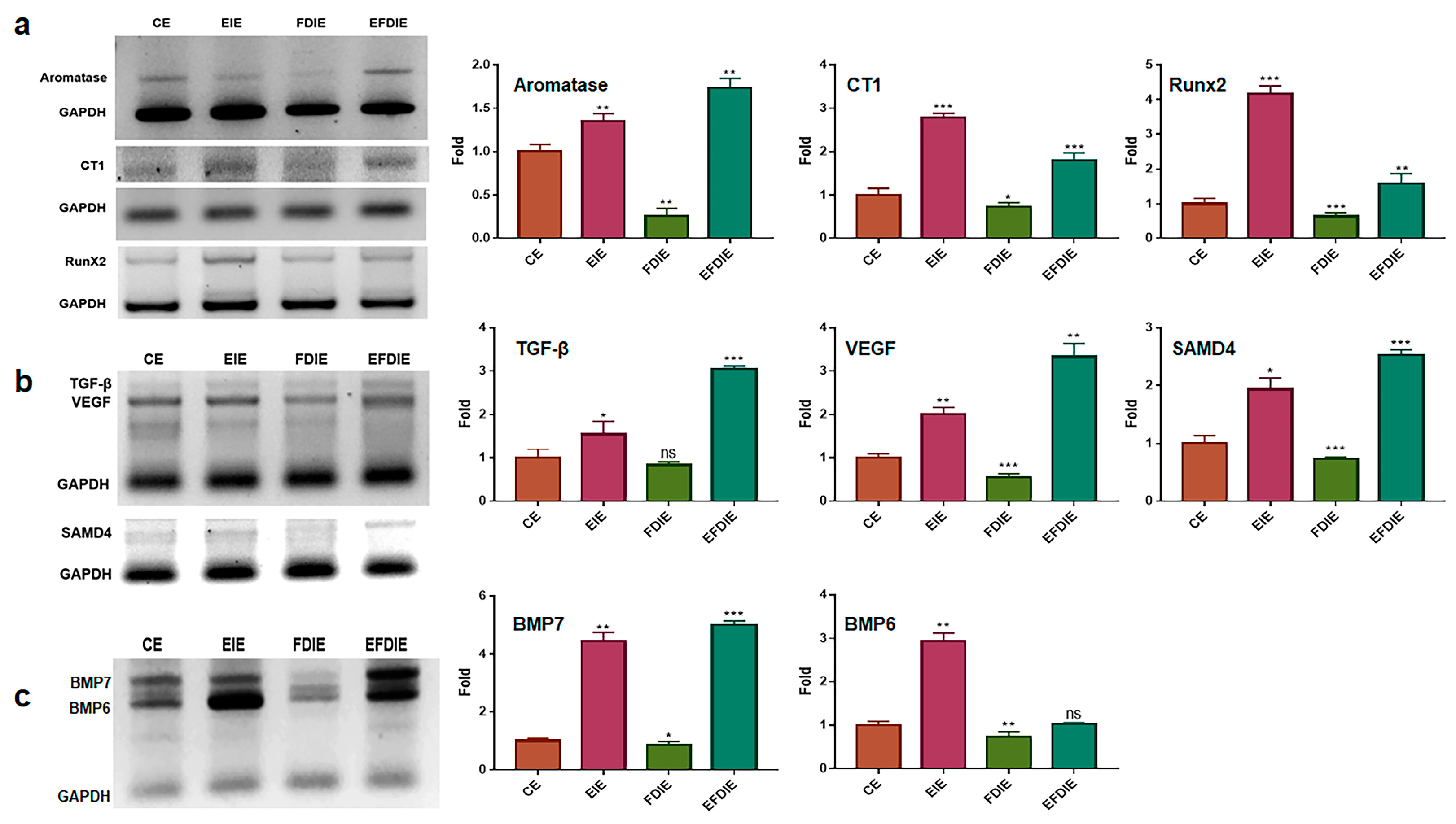
2.4. Quantitative PCR
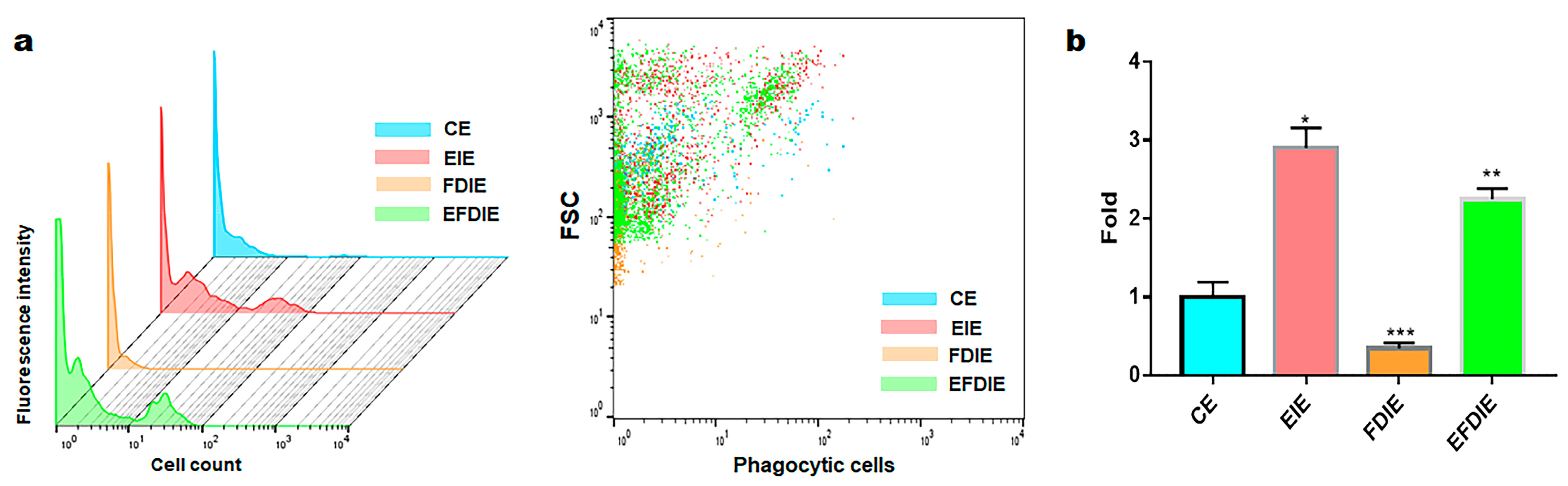
2.5. Phagocytic Activity Test
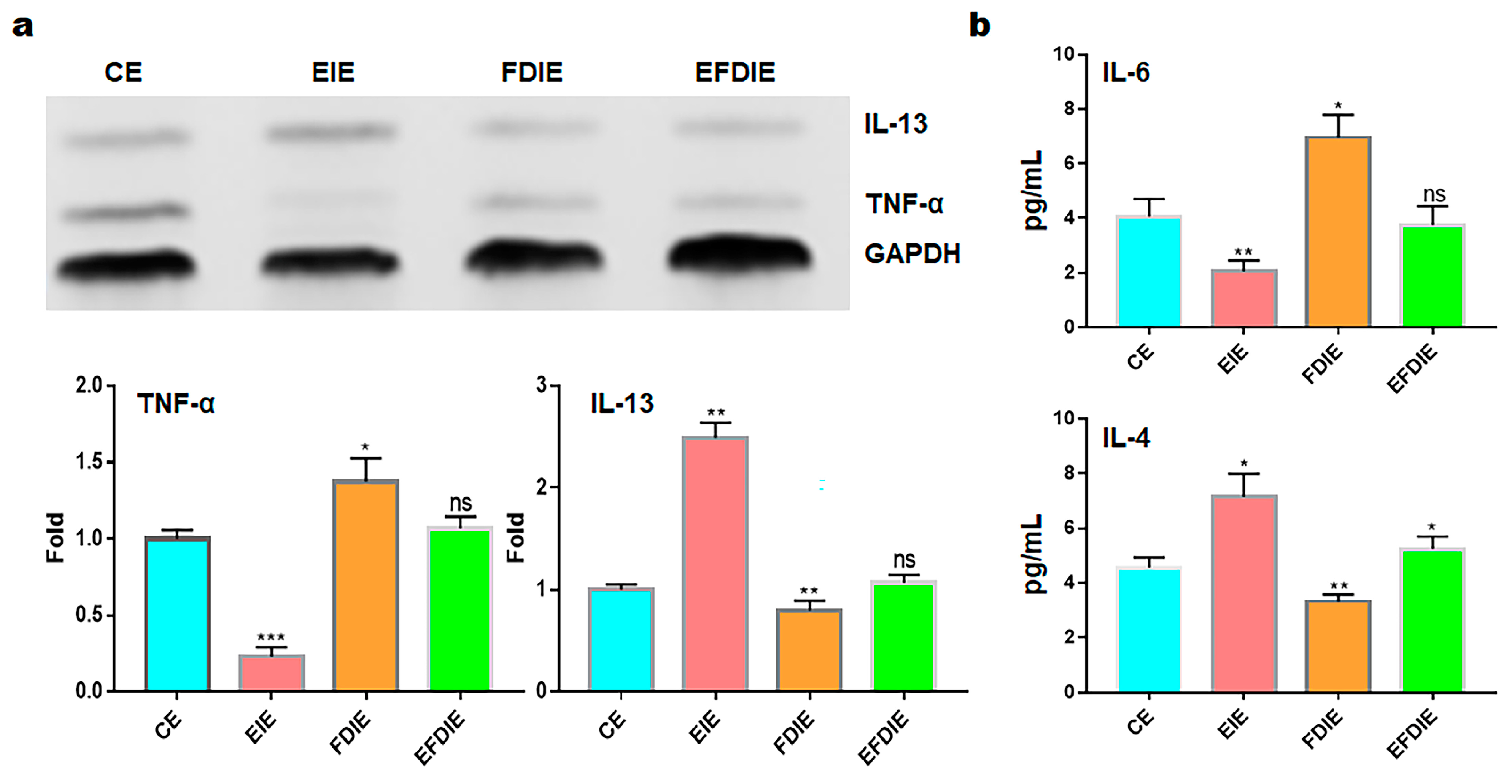
2.6. Evaluating of Cytokine Concentration
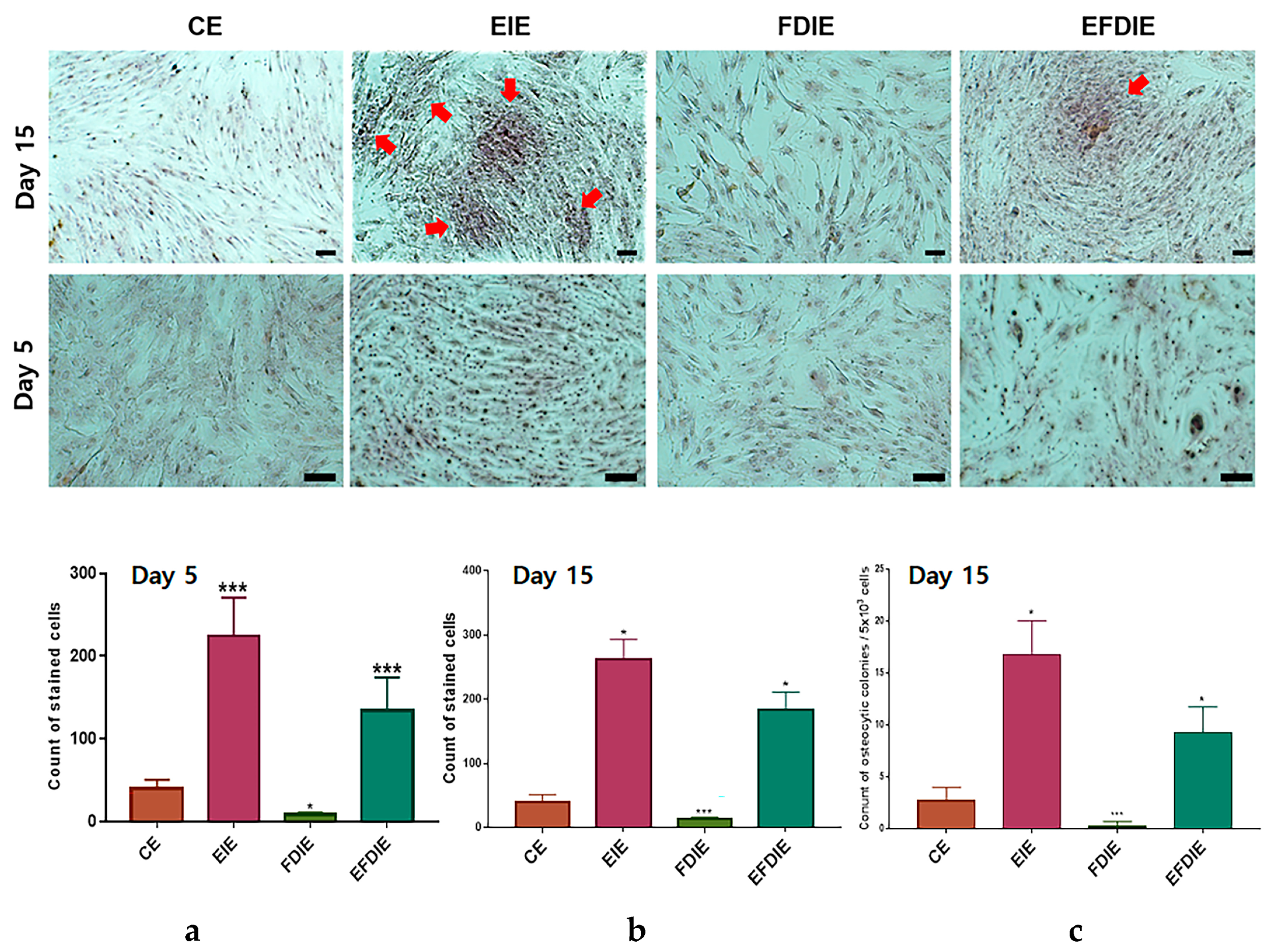
2.7. Alizarin O Staining
2.8. Profiling of microRNA in MSEIEs
2.9. Statistical Analysis
3. Results
3.1. Antiapoptotic Function of Erucic Acid in Fibroblasts Exposed to Fine Dust
3.2. Protecting and Enhancing of Osteocytic Differentiation by the Induced Exosomes
3.3. Activation of Immunity in Macrophages with Induced Exosomes
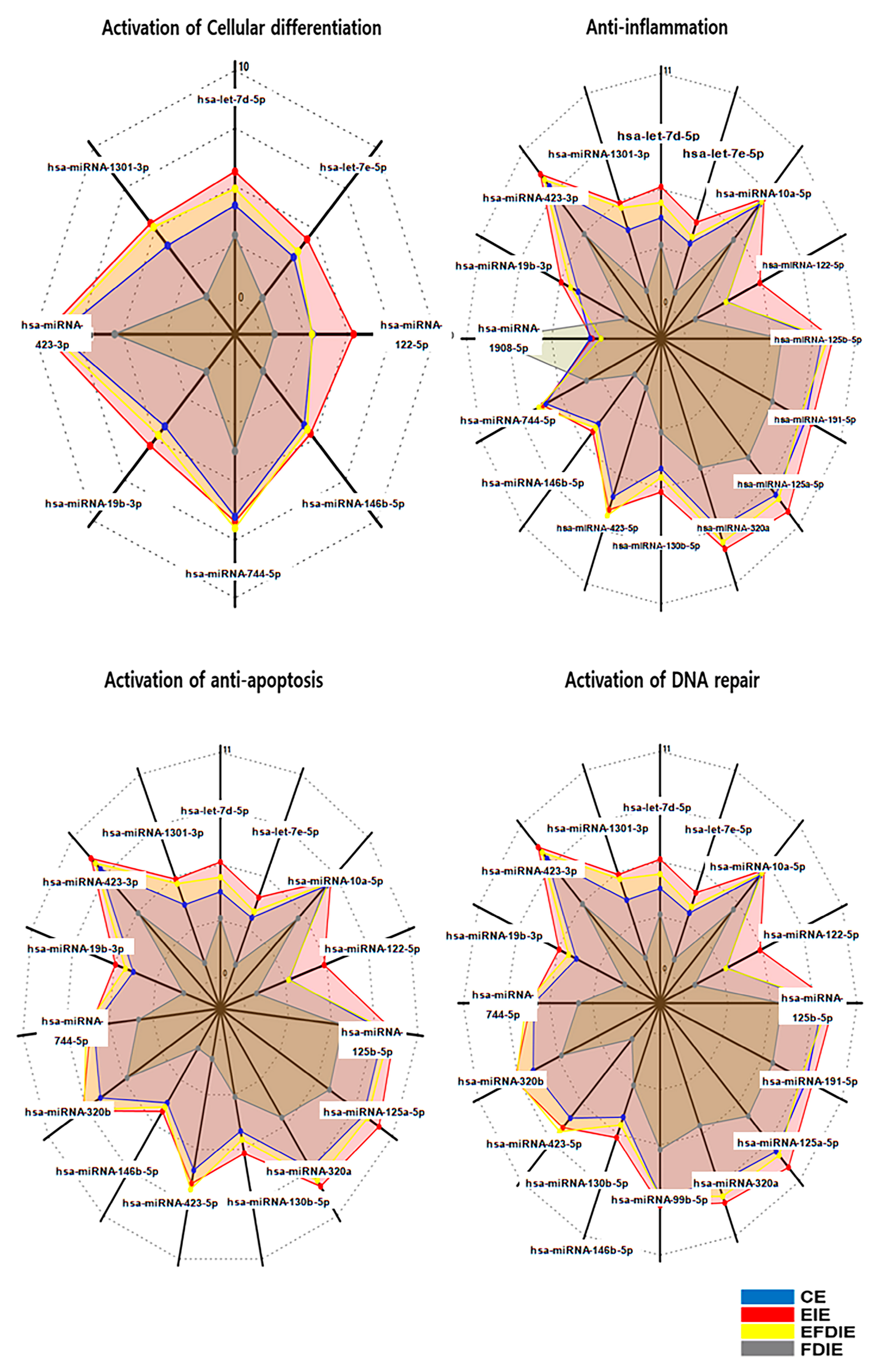
3.4. Profiling of miRNAs in Varios Exosomes
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhang, J.; Chen, Z.; Shan, D.; Wu, Y.; Zhao, Y.; Li, C.; Shu, Y.; Linghu, X.; Wang, B. Adverse effects of exposure to fine particles and ultrafine particles in the environment on different organs of organisms. J. Environ. Sci. 2024, 135, 449–473. [Google Scholar] [CrossRef]
- Combes, A.; Franchineau, G. Fine particle environmental pollution and cardiovascular diseases. Metabolism 2019, 100, 153944. [Google Scholar] [CrossRef]
- Zhang, Z.; Dong, B.; Li, S.; Chen, G.; Yang, Z.; Dong, Y.; Wang, Z.; Ma, J.; Guo, Y. Exposure to ambient particulate matter air pollution, blood pressure and hypertension in children and adolescents: A national cross-sectional study in China. Environ. Int. 2019, 128, 103–108. [Google Scholar] [CrossRef] [PubMed]
- Javadinejad, S.; Dara, R.; Jafary, F. Health impacts of extreme events. Saf. Extrem. Environ. 2020, 2, 171–181. [Google Scholar] [CrossRef]
- Xu, H.; Jia, Y.; Sun, Z.; Su, J.; Liu, Q.S.; Zhou, Q.; Jiang, G. Environmental pollution, a hidden culprit for health issues. Eco-Environ. Health 2022, 1, 31–45. [Google Scholar] [CrossRef]
- Park, Y.; Shin, G.H.; Jin, G.S.; Jin, S.; Kim, B.; Lee, S.G. Effects of black jade on osteogenic differentiation of adipose derived stem cells under benzopyrene. Appl. Sci. 2021, 11, 1346. [Google Scholar] [CrossRef]
- Fan, H.; Zhao, C.; Yang, Y.; Yang, X. Spatio-Temporal Variations of the PM2. 5/PM10 Ratios and Its Application to Air Pollution Type Classification in China. Front. Environ. Sci. 2021, 14, 692440. [Google Scholar]
- Wada, Y.; Ikemoto, T.; Morine, Y.; Imura, S.; Saito, Y.; Yamada, S.; Shimada, M. The differences in the characteristics of insulin-producing cells using human adipose-tissue derived mesenchymal stem cells from subcutaneous and visceral tissues. Sci. Rep. 2019, 9, 13204. [Google Scholar] [CrossRef]
- Bhattarai, G.; Shrestha, S.K.; Sim, H.-J.; Lee, J.-C.; Kook, S.-H. Effects of fine particulate matter on bone marrow-conserved hematopoietic and mesenchymal stem cells: A systematic review. Exp. Mol. Med. 2024, 56, 118–128. [Google Scholar] [CrossRef]
- Adami, G.; Cattani, G.; Rossini, M.; Viapiana, O.; Olivi, P.; Orsolini, G.; Bertoldo, E.; Fracassi, E.; Gatti, D.; Fassio, A. Association between exposure to fine particulate matter and osteoporosis: A population-based cohort study. Osteoporos. Int. 2022, 33, 169–176. [Google Scholar] [CrossRef]
- Korzh, M.; Vorontsov, P.; Ashukina, N.; Maltseva, V. Age-related features of bone regeneration (literature review). Orthop. Traumatol. Prosthet. 2021, 3, 92–100. [Google Scholar] [CrossRef]
- Velletri, T.; Xie, N.; Wang, Y.; Huang, Y.; Yang, Q.; Chen, X.; Chen, Q.; Shou, P.; Gan, Y.; Cao, G. P53 functional abnormality in mesenchymal stem cells promotes osteosarcoma development. Cell Death Dis. 2016, 7, e2015. [Google Scholar] [CrossRef] [PubMed]
- Holleville, N.; Matéos, S.; Bontoux, M.; Bollerot, K.; Monsoro-Burq, A.H. Dlx5 drives Runx2 expression and osteogenic differentiation in developing cranial suture mesenchyme. Dev. Biol. 2007, 304, 860–874. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Hilton, M.J.; Anolik, J.H.; Welle, S.L.; Zhao, C.; Yao, Z.; Li, X.; Wang, Z.; Boyce, B.F.; Xing, L. NOTCH inhibits osteoblast formation in inflammatory arthritis via noncanonical NF-κB. J. Clin. Investig. 2014, 124, 3200–3214. [Google Scholar] [CrossRef] [PubMed]
- Bonewald, L.F. Cell–cell and cell–matrix interactions in bone. In Intercellular Signaling in Development and Disease: Cell Signaling Collection; Academic Press: Cambridge, MA, USA, 2011; p. 109. [Google Scholar]
- Mandal, C.C. Osteolytic metastasis in breast cancer: Effective prevention strategies. Expert Rev. Anticancer. Ther. 2020, 20, 797–811. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Huang, J.; Chen, C.; Wang, Y.; Hao, Z.; Chen, T.; Wang, J.; Li, J. Strategies of macrophages to maintain bone homeostasis and promote bone repair: A narrative review. J. Funct. Biomater. 2022, 14, 18. [Google Scholar] [CrossRef] [PubMed]
- Sahasrabudhe, M. Crismer values and erucic acid contents of rapeseed oils. J. Am. Oil Chem. Soc. 1977, 54, 323–324. [Google Scholar] [CrossRef]
- Goyal, A.; Dubey, N.; Verma, A.; Agrawal, A. Erucic acid: A possible therapeutic agent for neurodegenerative diseases. Curr. Mol. Med. 2024, 24, 419–427. [Google Scholar] [CrossRef] [PubMed]
- Wani, I.A.; ul Ashraf, Z.; Muzzaffar, S. Erucic acid. In Handbook of Plant and Animal Toxins in Food; CRC Press: Boca Raton, FL, USA, 2022; pp. 169–176. [Google Scholar]
- Galanty, A.; Grudzińska, M.; Paździora, W.; Paśko, P. Erucic acid—Both sides of the story: A concise review on its beneficial and toxic properties. Molecules 2023, 28, 1924. [Google Scholar] [CrossRef]
- Chen, X.; Shang, L.; Deng, S.; Li, P.; Chen, K.; Gao, T.; Zhang, X.; Chen, Z.; Zeng, J. Peroxisomal oxidation of erucic acid suppresses mitochondrial fatty acid oxidation by stimulating malonyl-CoA formation in the rat liver. J. Biol. Chem. 2020, 295, 10168–10179. [Google Scholar] [CrossRef]
- Akhtar, M.J.; Khan, S.A. Chemistry and Biological Activity of Mustard Oil: Therapeutic Benefits and Risk to Healthcare. Rev. Bras. De Farmacogn. 2024, 34, 65–79. [Google Scholar] [CrossRef]
- Wendlinger, C.; Hammann, S.; Vetter, W. Various concentrations of erucic acid in mustard oil and mustard. Food Chem. 2014, 153, 393–397. [Google Scholar] [CrossRef] [PubMed]
- Tkach, M.; Théry, C. Communication by extracellular vesicles: Where we are and where we need to go. Cell 2016, 164, 1226–1232. [Google Scholar] [CrossRef]
- Shen, X.; Song, S.; Chen, N.; Liao, J.; Zeng, L. Stem cell-derived exosomes: A supernova in cosmetic dermatology. J. Cosmet. Dermatol. 2021, 20, 3812–3817. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Li, S.; Du, C.; Zhang, Y.; Li, Y.; Chu, L.; Han, X.; Galons, H.; Zhang, Y.; Sun, H. Exosomes from different cells: Characteristics, modifications, and therapeutic applications. Eur. J. Med. Chem. 2020, 207, 112784. [Google Scholar] [CrossRef] [PubMed]
- Park, Y.; Lee, K.; Kim, S.W.; Lee, M.W.; Kim, B.; Lee, S.G. Effects of induced exosomes from endometrial cancer cells on tumor activity in the presence of Aurea helianthus extract. Molecules 2021, 26, 2207. [Google Scholar] [CrossRef] [PubMed]
- Bouillon, R. Diabetic bone disease. Calcif. Tissue Int. 1991, 49, 155–160. [Google Scholar] [CrossRef]
- Harris, P.E.; Bouloux, P.-M.G. Metabolic bone disease. In Endocrinology in Clinical Practice, Harris, P.E., Bouloux, P.M., Eds.; CRC Press: Boca Raton, FL, USA, 2014; Volume 243. [Google Scholar]
- de Groot, A.C. Propolis: A review of properties, applications, chemical composition, contact allergy, and other adverse effects. Dermatitis 2013, 24, 263–282. [Google Scholar] [CrossRef]
- Ritz, B.; Hoffmann, B.; Peters, A. The effects of fine dust, ozone, and nitrogen dioxide on health. Dtsch. Ärzteblatt Int. 2019, 116, 881. [Google Scholar] [CrossRef]
- Kim, K.E.; Cho, D.; Park, H.J. Air pollution and skin diseases: Adverse effects of airborne particulate matter on various skin diseases. Life Sci. 2016, 152, 126–134. [Google Scholar] [CrossRef]
- Lee, C.-W.; Chi, M.-C.; Peng, K.-T.; Chiang, Y.-C.; Hsu, L.-F.; Yan, Y.-L.; Li, H.-Y.; Chen, M.-C.; Lee, I.-T.; Lai, C.-H. Water-soluble fullerenol C60 (OH) 36 toward effective anti-air pollution induced by urban particulate matter in HaCaT cell. Int. J. Mol. Sci. 2019, 20, 4259. [Google Scholar] [CrossRef] [PubMed]
- Kirindage, K.G.I.S.; Jayasinghe, A.M.K.; Cho, N.; Cho, S.H.; Yoo, H.M.; Fernando, I.P.S.; Ahn, G. Fine-dust-induced skin inflammation: Low-molecular-weight fucoidan protects keratinocytes and underlying fibroblasts in an integrated culture model. Mar. Drugs 2022, 21, 12. [Google Scholar] [CrossRef] [PubMed]
- Amarasekara, D.S.; Kim, S.; Rho, J. Regulation of osteoblast differentiation by cytokine networks. Int. J. Mol. Sci. 2021, 22, 2851. [Google Scholar] [CrossRef] [PubMed]
- Stein, G.S.; Lian, J.B. Molecular mechanisms mediating proliferation/differentiation interrelationships during progressive development of the osteoblast phenotype. Endocr. Rev. 1993, 14, 424–442. [Google Scholar] [CrossRef] [PubMed]
- Pitkänen, S. In Vitro and In Vivo Osteogenesis and Vasculogenesis in Synthetic Bone Grafts; Tampere University: Tampere, Finland, 2020. [Google Scholar]
- Komori, T. Regulation of proliferation, differentiation and functions of osteoblasts by Runx2. Int. J. Mol. Sci. 2019, 20, 1694. [Google Scholar] [CrossRef] [PubMed]
- Loi, F.; Córdova, L.A.; Zhang, R.; Pajarinen, J.; Lin, T.-h.; Goodman, S.B.; Yao, Z. The effects of immunomodulation by macrophage subsets on osteogenesis in vitro. Stem Cell Res. Ther. 2016, 7, 15. [Google Scholar] [CrossRef] [PubMed]
- Niu, Y.; Wang, Z.; Shi, Y.; Dong, L.; Wang, C. Modulating macrophage activities to promote endogenous bone regeneration: Biological mechanisms and engineering approaches. Bioact. Mater. 2021, 6, 244–261. [Google Scholar] [CrossRef]
- McPhillips, K.; Janssen, W.J.; Ghosh, M.; Byrne, A.; Gardai, S.; Remigio, L.; Bratton, D.L.; Kang, J.L.; Henson, P. TNF-α inhibits macrophage clearance of apoptotic cells via cytosolic phospholipase A2 and oxidant-dependent mechanisms. J. Immunol. 2007, 178, 8117–8126. [Google Scholar] [CrossRef] [PubMed]
- Yi, S.; Jiang, X.; Tang, X.; Li, Y.; Xiao, C.; Zhang, J.; Zhou, T. IL-4 and IL-10 promotes phagocytic activity of microglia by up-regulation of TREM2. Cytotechnology 2020, 72, 589–602. [Google Scholar] [CrossRef]
- Meo, S.A.; Rasheed, S.; Khan, M.M.; Shujauddin, S.; Al-Tuwaijri, A.S. Effect of cement dust exposure on phagocytic function of polymorphonuclear neutrophils in cement mill workers. Int. J. Occup. Med. Environ. Health 2008, 21, 133–139. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Band Size (bp) | F/R* | Seq (5’ → 3’) |
|---|---|---|---|
| AKT | 682 | F | GGCTGCCAAGTGTCAAATCC |
| R | AGTGCTCCCCCACTTACTTG | ||
| NFκB-P50 | 480 | F | CGGAGCCCTCTTTCACAGTT |
| R | TTCAGCTTAGGAGCGAAGGC | ||
| NFκB-P52 | 410 | F | AGGTGCTGTAGCGGGATTTC |
| R | AGAGGCACTGTATAGGGCAG | ||
| Bcl2 | 386 | F | CTGCTGACATGCTTGGAAAA |
| R | ATTGGGCTACCCCAGCAATG | ||
| IL13 | 361 | F | CCACGGTCATTGCTCTCACT |
| R | CCCGCCTACCCAAGACATTT | ||
| TNF-α | 262 | F | AAGAGGGAGAGAAGCAACTAC |
| R | AGGAGAAGAGGCTGAGGAAC | ||
| Aromatase | 287 | F | TCAGAGCAACCTTCTTAGGCTC |
| R | AGAAAAGTTACCTGAGAGGCCA | ||
| CT1 | 542 | F | GCTCGTGGAAATGATGGTGC |
| R | CCTCGCTTTCCTTCCTCTCC | ||
| RunX2 | 470 | F | TTGCAGCCATAAGAGGGTAG |
| R | GTCACTTTCTTGGAGCAGGA | ||
| BMP7 | 577 | F | CCCGGGTAGCGCGTAG |
| R | CGTTCCCGGATGTAGTCCTT | ||
| BMP6 | 520 | F | CTTCCCATCCTTTCTGCGAGC |
| R | GGGCCACCATGAAGTTTACC | ||
| TGF-β | 369 | F | CTGTCCAACATGATCGTGCG |
| R | AGTGCCCAAGGTGCTCAATA | ||
| VEGF | 316 | F | ACTGCCATCCAATCGAGACC |
| R | GAAGGCAAGACCCCACCATA | ||
| SAMD4 | 247 | F | TCTTGACAGTGTTCCACGGG |
| R | GCAAAGCCAAGGAAGCACAT | ||
| GAPDH | 210 | F | GTGGTCTCCTCTGACTTCAACA |
| R | CTCTTCCTCTTGTGCTCTTGCT |
| Categories | Activation of Differentiation | Anti-Inflammation | Anti-Apoptosis | Activation of DNA Repair |
|---|---|---|---|---|
| Upregulation Genes | hsa-let-7d-5p | hsa-let-7d-5p | hsa-let-7d-5p | hsa-let-7d-5p |
| hsa-let-7e-5p | hsa-let-7e-5p | hsa-let-7e-5p | hsa-let-7e-5p | |
| hsa-miRNA-1301-3p * | hsa-miRNA-1301-3p | hsa-miRNA-1301-3p | hsa-miRNA-1301-3p | |
| hsa-miRNA-423-3p | hsa-miRNA-423-3p | hsa-miRNA-423-3p | hsa-miRNA-423-3p | |
| hsa-miRNA-19b-3p | hsa-miRNA-19b-3p | hsa-miRNA-19b-3p | hsa-miRNA-19b-3p | |
| - | hsa-miRNA-1908-5p * | - | - | |
| hsa-miRNA-744-5p | hsa-miRNA-744-5p | hsa-miRNA-744-5p | hsa-miRNA-744-5p | |
| hsa-miRNA-146b-5p | hsa-miRNA-146b-5p | hsa-miRNA-146b-5p | - | |
| - | hsa-miRNA-423-5p * | hsa-miRNA-423-5p * | hsa-miRNA-423-5p * | |
| - | hsa-miRNA-130b-5p | hsa-miRNA-130b-5p | hsa-miRNA-130b-5p | |
| - | hsa-miRNA-320a | hsa-miRNA-320a | hsa-miRNA-320a | |
| - | - | hsa-miRNA-320b | hsa-miRNA-320b | |
| - | hsa-miRNA-125a-5p | - | hsa-miRNA-125a-5p | |
| - | hsa-miRNA-191-5p | - | hsa-miRNA-191-5p | |
| - | hsa-miRNA-125b-5p | hsa-miRNA-125b-5p | hsa-miRNA-125b-5p | |
| hsa-miRNA-122-5p | hsa-miRNA-122-5p | hsa-miRNA-122-5p * | hsa-miRNA-122-5p | |
| - | hsa-miRNA-10a-5p | hsa-miRNA-10a-5p | hsa-miRNA-10a-5p | |
| - | - | hsa-miRNA-99b-5p |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, H.; Kim, B. Osteogenic Protection against Fine Dust with Erucic Acid-Induced Exosomes. J. Funct. Biomater. 2024, 15, 215. https://doi.org/10.3390/jfb15080215
Kim H, Kim B. Osteogenic Protection against Fine Dust with Erucic Acid-Induced Exosomes. Journal of Functional Biomaterials. 2024; 15(8):215. https://doi.org/10.3390/jfb15080215
Chicago/Turabian StyleKim, Hyunjung, and Boyong Kim. 2024. "Osteogenic Protection against Fine Dust with Erucic Acid-Induced Exosomes" Journal of Functional Biomaterials 15, no. 8: 215. https://doi.org/10.3390/jfb15080215