
Biomaterials Designed to Modulate Reactive Oxygen Species for Enhanced Bone Regeneration in Diabetic Conditions

Abstract
:1. Introduction
2. The Role of ROS in Bone Metabolism under Diabetic Physiologic Conditions
2.1. ROS Production
2.1.1. Mitochondrial ROS Production
2.1.2. Non-Mitochondrial ROS Production
- Glyceraldehyde-3-phosphate in Glucose Autoxidation
- Polyol Pathway
- PKC Pathway
- Hexosamine Pathway
- Advanced Glycation End Products (AGEs)
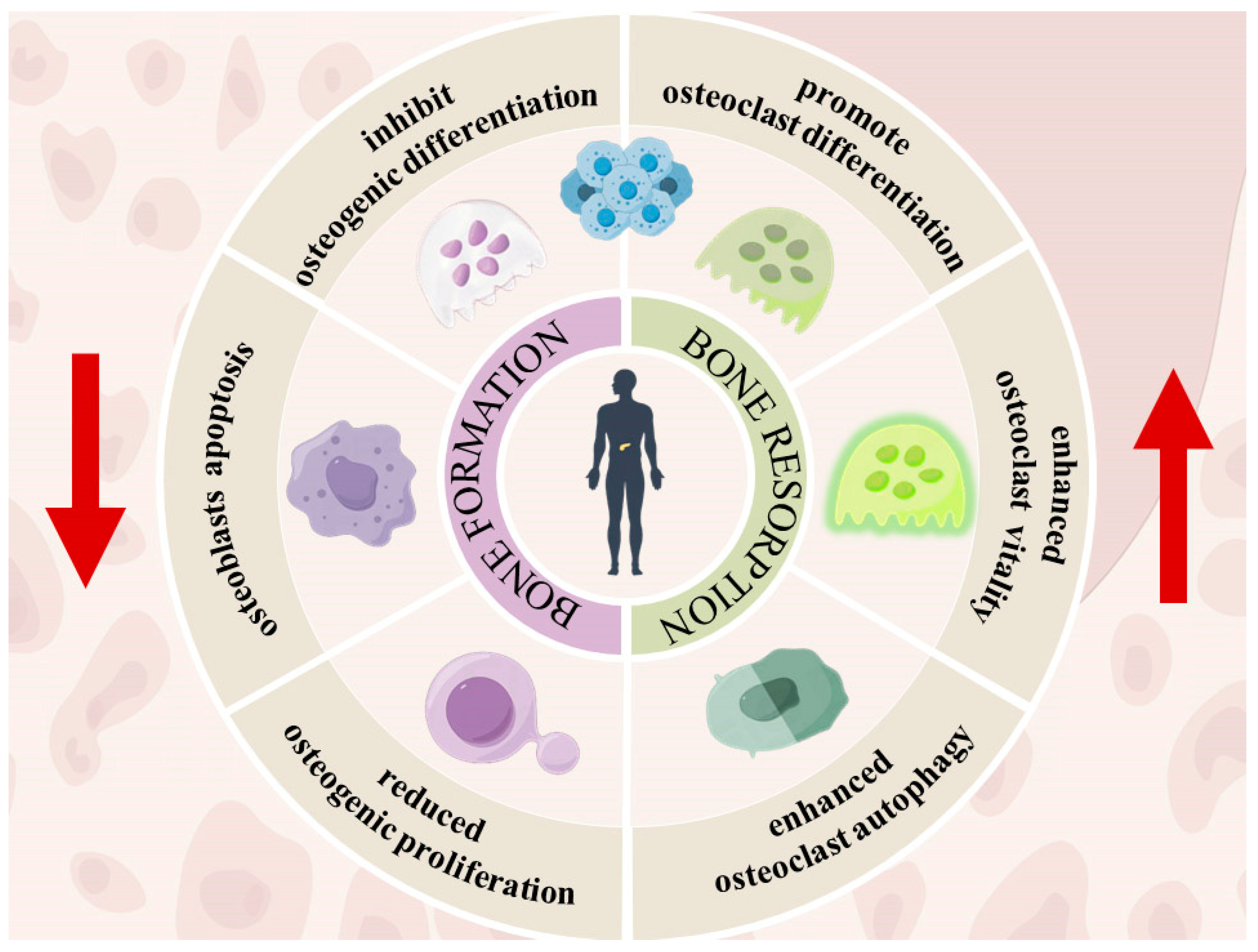
2.2. ROS on Bone Remodeling in Hyperglycemia
3. Chemical Compounds and Biomaterials as Antioxidant Agents
3.1. Natural
3.1.1. Endogenous
3.1.2. Exogenous
3.2. Synthetic
3.2.1. Small Molecules Chemical Compounds
3.2.2. Macromolecular Antioxidant Biomaterials
Scaffold

{kind=link}
| Antioxidant Biomaterial | Study Design | Targeted Model | ROS Scavenger Compounds | Ref. |
|---|---|---|---|---|
| ALA-loaded poly (lactic-co-glycolic acid) (PLGA) microspheres | In vitro/in vivo | BMSC/STZ-induced diabetic SD rats | α-lipoic acid | [90] |
| PGO-PHA-AG scaffold | In vitro/in vivo | BMSC/STZ-induced diabetic SD rats‘ mandibular periodontitis | Polydopamine | [92] |
| Three-dimensionally printed reduced glutathione grafted gelatine methacrylate (GelMA-g-GSH) | In vitro/in vivo | MC3T3-E1/Calvarium bone defects in bone defect SD rats | GSH | [99] |
| nCe-scaffold | In vitro/in vivo | MSC/calvarium bone defects in bone defect SD rats and STZ-induced diabetic SD rats | Nanoceria (Ce) | [100] |
| Thiol–methacrylate networks | In vitro | NA 1 | Thiol–methacrylate networks | [103] |
| Thiol–methacrylate interconnected porous scaffolds using emulsion templating | In vitro/in vivo | Bilateral calvarial defect model in Zucker diabetic fatty rats | Thiol–methacrylate networks | [104] |
Coating
- Material-Based Surface Modification
- Composite Coating Optimization
| Antioxidant Biomaterial | Study Design | Targeted Model | ROS Scavenger Compounds | Ref. |
|---|---|---|---|---|
| Ca-PA-modified SLA titanium surface | In vitro/In vivo | hBMSC/STZ-induced diabetic SD rats | Ca–phytic acid | [111] |
| Zn&Sr- Sulfonated PEEK | In vitro/In vivo | MC3TE-E1/STZ-induced diabetic SD rats | Zn and Sr ion | [120] |
| Three-dimensional coating of hydroxyapatite-functionalized nanoparticles of polydopamine (HA/nPDAs) | In vitro | MC3T3-E1 | nPDA | [124] |
| Silibinin-modified hydroxyapatite coating | In vitro/In vivo | MC3TE-E1/STZ-induced diabetic SD rats | Silibinin | [125] |
| Silk fibroin-based hydroxyapatite (SF/HA) hybrid coating | In vitro/In vivo | Primary rabbit osteoblasts/diabetic rabbits | SHT | [126] |
| HA/CS composite coating | In vitro/In vivo | Primary rat osteoblasts/STZ-induced diabetic sheep | Nano-HA/CS composite coating | [131] |
Nanoparticles
| Antioxidant Biomaterial | Study Design | Targeted Model | ROS Scavenger Compounds | Ref. |
|---|---|---|---|---|
| La-CNPs | In vitro/in vivo | MC3TE-E1/STZ-induced diabetic SD rats | Lanthanum | [139] |
| Chitosan gold nanoparticles conjugated with PPARγ cDNA | In vitro/in vivo | MC3TE-E1/STZ-induced diabetic SD rats | Chitosan gold nanoparticles conjugated with PPARγ cDNA | [141] |
| Curcumin-loaded tetrahedral framework nucleic acid (tFNA) particle | In vitro/in vivo | BMSC/diabetic osteoporosis mice using HFD + STZ | Curcumin | [146] |
Hydrogel
| Antioxidant Biomaterial | Study Design | Targeted Model | ROS Scavenger Compounds | Ref. |
|---|---|---|---|---|
| GelMA/POM nano-hydrogel | In vitro/In vivo | MC3TE-E1/STZ-induced diabetic SD rats | Mo-based polyoxometalate nanoclusters (POM) | [149] |
| Three-dimensionally printed reduced glutathione grafted gelatine methacrylate (GelMA-g-GSH) | In vitro/In vivo | MC3T3-E1/calvarium bone defects in bone defect SD rats | GSH | [99] |
| Metformin-loaded zeolitic imidazolate frameworks nanoparticle-modified hydrogel(GelMA/Met@ZIF-8) | In vitro/In vivo | BMDMs and MC3T3-E2/calvarium bone defects in bone defect SD rats | Metformin | [153] |
| a double-network hydrogel consisting of phenylboronic-acid-crosslinked poly(vinyl alcohol) and gelatin colloids | In vitro/In vivo | BMDMs and MC3T3-E1/calvarium bone defects in bone defect SD rats | BMP-2 (HIB) | [154]. |
| PDLLA-PEG-PDLLA-Met@MSN-SDF-1 hydrogel | In vitro/In vivo | rBMSCs/STZ-induced diabetic rats | Met@MSN (metformin) | [155] |
4. Future and Prospects
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Sarwar, N.; Gao, P.; Seshasai, S.R.; Gobin, R.; Kaptoge, S.; Di Angelantonio, E.; Ingelsson, E.; Lawlor, D.A.; Selvin, E.; Stampfer, M.; et al. Diabetes mellitus, fasting blood glucose concentration, and risk of vascular disease: A collaborative meta-analysis of 102 prospective studies. Lancet 2010, 375, 2215–2222. [Google Scholar] [CrossRef] [PubMed]
- Magliano, D.J.; Boyko, E.J. IDF Diabetes Atlas 10th Edition Scientific Committee. IDF Diabetes Atlas; International Diabetes Federation©: Brussels, Belgium, 2021. [Google Scholar]
- Li, Y.; Teng, D.; Shi, X.; Qin, G.; Qin, Y.; Quan, H.; Shi, B.; Sun, H.; Ba, J.; Chen, B.; et al. Prevalence of diabetes recorded in mainland China using 2018 diagnostic criteria from the American Diabetes Association: National cross sectional study. Bmj 2020, 369, m997. [Google Scholar] [CrossRef] [PubMed]
- Rains, J.L.; Jain, S.K. Oxidative stress, insulin signaling, and diabetes. Free Radic. Biol. Med. 2011, 50, 567–575. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Ba, Y.; Xing, Q.; Du, J.L. Diabetes mellitus and the risk of fractures at specific sites: A meta-analysis. BMJ Open 2019, 9, e024067. [Google Scholar] [CrossRef] [PubMed]
- Tanios, M.; Brickman, B.; Cage, E.; Abbas, K.; Smith, C.; Atallah, M.; Baroi, S.; Lecka-Czernik, B. Diabetes and Impaired Fracture Healing: A Narrative Review of Recent Literature. Curr. Osteoporos. Rep. 2022, 20, 229–239. [Google Scholar] [CrossRef] [PubMed]
- Giacco, F.; Brownlee, M. Oxidative stress and diabetic complications. Circ. Res. 2010, 107, 1058–1070. [Google Scholar] [CrossRef] [PubMed]
- Folli, F.; Corradi, D.; Fanti, P.; Davalli, A.; Paez, A.; Giaccari, A.; Perego, C.; Muscogiuri, G. The role of oxidative stress in the pathogenesis of type 2 diabetes mellitus micro- and macrovascular complications: Avenues for a mechanistic-based therapeutic approach. Curr. Diabetes Rev. 2011, 7, 313–324. [Google Scholar] [CrossRef] [PubMed]
- Sies, H. Chapter 13—Oxidative Stress: Eustress and Distress in Redox Homeostasis. In Stress: Physiology, Biochemistry, and Pathology; Fink, G., Ed.; Academic Press: Cambridge, MA, USA, 2019; pp. 153–163. [Google Scholar]
- Sies, H.; Jones, D.P. Reactive oxygen species (ROS) as pleiotropic physiological signalling agents. Nat. Rev. Mol. Cell Biol. 2020, 21, 363–383. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.D.; Amirthalingam, S.; Kim, S.L.; Lee, S.S.; Rangasamy, J.; Hwang, N.S. Biomimetic Materials and Fabrication Approaches for Bone Tissue Engineering. Adv. Healthc. Mater. 2017, 6, 1700612. [Google Scholar] [CrossRef]
- Bhatti, J.S.; Sehrawat, A.; Mishra, J.; Sidhu, I.S.; Navik, U.; Khullar, N.; Kumar, S.; Bhatti, G.K.; Reddy, P.H. Oxidative stress in the pathophysiology of type 2 diabetes and related complications: Current therapeutics strategies and future perspectives. Free Radic. Biol. Med. 2022, 184, 114–134. [Google Scholar] [CrossRef]
- Domazetovic, V.; Marcucci, G.; Iantomasi, T.; Brandi, M.L.; Vincenzini, M.T. Oxidative stress in bone remodeling: Role of antioxidants. Clin. Cases Min. Bone Metab. 2017, 14, 209–216. [Google Scholar] [CrossRef] [PubMed]
- Letts, J.A.; Sazanov, L.A. Clarifying the supercomplex: The higher-order organization of the mitochondrial electron transport chain. Nat. Struct. Mol. Biol. 2017, 24, 800–808. [Google Scholar] [CrossRef] [PubMed]
- Letts, J.A.; Fiedorczuk, K.; Sazanov, L.A. The architecture of respiratory supercomplexes. Nature 2016, 537, 644–648. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharjee, S.; Bhattacharjee, S. ROS and oxidative stress: Origin and implication. In Reactive Oxygen Species in Plant Biology; Springer: New Delhi, India, 2019; pp. 1–31. [Google Scholar]
- Skliarova, E.I.; Popova, T.N.; Shulgin, K.K. Effects of N-[Imino(1-Piperidinyl)Methyl] Guanidine on the Intensity of Free Radical Processes, Aconitase Activity, and Citrate Level in the Tissues of Rats with Experimental Type 2 Diabetes Mellitus. Bull. Exp. Biol. Med. 2016, 161, 261–265. [Google Scholar] [CrossRef]
- Smirnov, L.; Inchina, V.; IaV, K.; Kokoreva, E.; ZhV, B. Possible pharmacological correction of metabolic impairments experimental diabetes mellitus by antioxidant. Biomeditsinskaia Khimiia 2004, 50, 502–508. [Google Scholar] [PubMed]
- Hernansanz-Agustín, P.; Enríquez, J.A. Generation of reactive oxygen species by mitochondria. Antioxidants 2021, 10, 415. [Google Scholar] [CrossRef] [PubMed]
- Brownlee, M. The pathobiology of diabetic complications: A unifying mechanism. Diabetes 2005, 54, 1615–1625. [Google Scholar] [CrossRef] [PubMed]
- Yu, T.; Robotham, J.L.; Yoon, Y. Increased production of reactive oxygen species in hyperglycemic conditions requires dynamic change of mitochondrial morphology. Proc. Natl. Acad. Sci. USA 2006, 103, 2653–2658. [Google Scholar] [CrossRef] [PubMed]
- Yu, T.; Jhun, B.S.; Yoon, Y. High-glucose stimulation increases reactive oxygen species production through the calcium and mitogen-activated protein kinase-mediated activation of mitochondrial fission. Antioxid. Redox Signal. 2011, 14, 425–437. [Google Scholar] [CrossRef]
- Sakai, K.; Matsumoto, K.; Nishikawa, T.; Suefuji, M.; Nakamaru, K.; Hirashima, Y.; Kawashima, J.; Shirotani, T.; Ichinose, K.; Brownlee, M.; et al. Mitochondrial reactive oxygen species reduce insulin secretion by pancreatic β-cells. Biochem. Biophys. Res. Commun. 2003, 300, 216–222. [Google Scholar] [CrossRef]
- Wolff, S.P.; Dean, R.T. Glucose autoxidation and protein modification. The potential role of ‘autoxidative glycosylation’ in diabetes. Biochem. J. 1987, 245, 243–250. [Google Scholar] [CrossRef] [PubMed]
- Graier, W.F.; Posch, K.; Fleischhacker, E.; Wascher, T.C.; Kostner, G.M. Increased superoxide anion formation in endothelial cells during hyperglycemia: An adaptive response or initial step of vascular dysfunction? Diabetes Res. Clin. Pract. 1999, 45, 153–160. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Deng, J.; Sun, D.; Chen, S.; Yao, X.; Wang, N.; Zhang, J.; Gu, Q.; Zhang, S.; Wang, J.; et al. FBXW7 alleviates hyperglycemia-induced endothelial oxidative stress injury via ROS and PARP inhibition. Redox Biol. 2022, 58, 102530. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Wei, F. DNA Damage and Repair in Diabetes Mellitus and Its Complications. Med. Recapitul. 2021, 27, 2203–2208. [Google Scholar]
- Du, X.; Matsumura, T.; Edelstein, D.; Rossetti, L.; Zsengellér, Z.; Szabó, C.; Brownlee, M. Inhibition of GAPDH activity by poly(ADP-ribose) polymerase activates three major pathways of hyperglycemic damage in endothelial cells. J. Clin. Investig. 2003, 112, 1049–1057. [Google Scholar] [CrossRef]
- Ott, M.; Gogvadze, V.; Orrenius, S.; Zhivotovsky, B. Mitochondria, oxidative stress and cell death. Apoptosis 2007, 12, 913–922. [Google Scholar] [CrossRef]
- Srivastava, S.K.; Ramana, K.V.; Bhatnagar, A. Role of aldose reductase and oxidative damage in diabetes and the consequent potential for therapeutic options. Endocr. Rev. 2005, 26, 380–392. [Google Scholar] [CrossRef] [PubMed]
- Yan, L.j. Redox imbalance stress in diabetes mellitus: Role of the polyol pathway. Anim. Models Exp. Med. 2018, 1, 7–13. [Google Scholar] [CrossRef]
- Wu, J.; Jin, Z.; Zheng, H.; Yan, L.J. Sources and implications of NADH/NAD(+) redox imbalance in diabetes and its complications. Diabetes Metab. Syndr. Obes. 2016, 9, 145–153. [Google Scholar] [CrossRef]
- Song, J.; Yang, X.; Yan, L.J. Role of pseudohypoxia in the pathogenesis of type 2 diabetes. Hypoxia 2019, 7, 33–40. [Google Scholar] [CrossRef]
- Gallagher, E.J.; LeRoith, D.; Stasinopoulos, M.; Zelenko, Z.; Shiloach, J. Polyol accumulation in muscle and liver in a mouse model of type 2 diabetes. J. Diabetes Complicat. 2016, 30, 999–1007. [Google Scholar] [CrossRef] [PubMed]
- Geraldes, P.; King, G.L. Activation of protein kinase C isoforms and its impact on diabetic complications. Circ. Res. 2010, 106, 1319–1331. [Google Scholar] [CrossRef] [PubMed]
- Inoguchi, T.; Li, P.; Umeda, F.; Yu, H.Y.; Kakimoto, M.; Imamura, M.; Aoki, T.; Etoh, T.; Hashimoto, T.; Naruse, M.; et al. High glucose level and free fatty acid stimulate reactive oxygen species production through protein kinase C—dependent activation of NAD(P)H oxidase in cultured vascular cells. Diabetes 2000, 49, 1939–1945. [Google Scholar] [CrossRef] [PubMed]
- Cosentino-Gomes, D.; Rocco-Machado, N.; Meyer-Fernandes, J.R. Cell signaling through protein kinase C oxidation and activation. Int. J. Mol. Sci. 2012, 13, 10697–10721. [Google Scholar] [CrossRef] [PubMed]
- Ighodaro, O.M. Molecular pathways associated with oxidative stress in diabetes mellitus. Biomed. Pharmacother. 2018, 108, 656–662. [Google Scholar] [CrossRef] [PubMed]
- Du, X.L.; Edelstein, D.; Rossetti, L.; Fantus, I.G.; Goldberg, H.; Ziyadeh, F.; Wu, J.; Brownlee, M. Hyperglycemia-induced mitochondrial superoxide overproduction activates the hexosamine pathway and induces plasminogen activator inhibitor-1 expression by increasing Sp1 glycosylation. Proc. Natl. Acad. Sci. USA 2000, 97, 12222–12226. [Google Scholar] [CrossRef] [PubMed]
- Lund, J.; Ouwens, D.M.; Wettergreen, M.; Bakke, S.S.; Thoresen, G.H.; Aas, V. Increased Glycolysis and Higher Lactate Production in Hyperglycemic Myotubes. Cells 2019, 8, 1101. [Google Scholar] [CrossRef] [PubMed]
- Asadipooya, K.; Uy, E.M. Advanced Glycation End Products (AGEs), Receptor for AGEs, Diabetes, and Bone: Review of the Literature. J. Endocr. Soc. 2019, 3, 1799–1818. [Google Scholar] [CrossRef] [PubMed]
- González, I.; Morales, M.A.; Rojas, A. Polyphenols and AGEs/RAGE axis. Trends and challenges. Food Res. Int. 2020, 129, 108843. [Google Scholar] [CrossRef]
- Guimarães, E.L.; Empsen, C.; Geerts, A.; van Grunsven, L.A. Advanced glycation end products induce production of reactive oxygen species via the activation of NADPH oxidase in murine hepatic stellate cells. J. Hepatol. 2010, 52, 389–397. [Google Scholar] [CrossRef]
- Lin, N.; Zhang, H.; Su, Q. Advanced glycation end-products induce injury to pancreatic beta cells through oxidative stress. Diabetes Metab. 2012, 38, 250–257. [Google Scholar] [CrossRef]
- Yao, D.; Brownlee, M. Hyperglycemia-induced reactive oxygen species increase expression of the receptor for advanced glycation end products (RAGE) and RAGE ligands. Diabetes 2010, 59, 249–255. [Google Scholar] [CrossRef]
- Moldogazieva, N.T.; Mokhosoev, I.M.; Mel’nikova, T.I.; Porozov, Y.B.; Terentiev, A.A. Oxidative Stress and Advanced Lipoxidation and Glycation End Products (ALEs and AGEs) in Aging and Age-Related Diseases. Oxid. Med. Cell Longev. 2019, 2019, 3085756. [Google Scholar] [CrossRef]
- Tao, H.; Ge, G.; Liang, X.; Zhang, W.; Sun, H.; Li, M.; Geng, D. ROS signaling cascades: Dual regulations for osteoclast and osteoblast. Acta Biochim. Biophys. Sin. 2020, 52, 1055–1062. [Google Scholar] [CrossRef] [PubMed]
- Carnevale, V.; Romagnoli, E.; D’Erasmo, E. Skeletal involvement in patients with diabetes mellitus. Diabetes/Metab. Res. Rev. 2004, 20, 196–204. [Google Scholar] [CrossRef]
- Atashi, F.; Modarressi, A.; Pepper, M.S. The role of reactive oxygen species in mesenchymal stem cell adipogenic and osteogenic differentiation: A review. Stem Cells Dev. 2015, 24, 1150–1163. [Google Scholar] [CrossRef]
- Almeida, M.; Han, L.; Martin-Millan, M.; O’Brien, C.A.; Manolagas, S.C. Oxidative stress antagonizes Wnt signaling in osteoblast precursors by diverting beta-catenin from T cell factor- to forkhead box O-mediated transcription. J. Biol. Chem. 2007, 282, 27298–27305. [Google Scholar] [CrossRef] [PubMed]
- Ambrogini, E.; Almeida, M.; Martin-Millan, M.; Paik, J.H.; Depinho, R.A.; Han, L.; Goellner, J.; Weinstein, R.S.; Jilka, R.L.; O’Brien, C.A.; et al. FoxO-mediated defense against oxidative stress in osteoblasts is indispensable for skeletal homeostasis in mice. Cell Metab. 2010, 11, 136–146. [Google Scholar] [CrossRef] [PubMed]
- Rached, M.T.; Kode, A.; Xu, L.; Yoshikawa, Y.; Paik, J.H.; Depinho, R.A.; Kousteni, S. FoxO1 is a positive regulator of bone formation by favoring protein synthesis and resistance to oxidative stress in osteoblasts. Cell Metab. 2010, 11, 147–160. [Google Scholar] [CrossRef]
- Zhang, Y.; Xiong, Y.; Zhou, J.; Xin, N.; Zhu, Z.; Wu, Y. FoxO1 expression in osteoblasts modulates bone formation through resistance to oxidative stress in mice. Biochem. Biophys. Res. Commun. 2018, 503, 1401–1408. [Google Scholar] [CrossRef]
- Hussein, A.M.; Sina, M. p-Nonylphenol Impairment of Osteogenic Differentiation of Mesenchymal Stem Cells was Found to be Due to Oxidative Stress and Down-Regulation of RUNX2 and BMP. Endocr. Metab. Immune Disord. Drug Targets 2020, 20, 1336–1346. [Google Scholar] [CrossRef]
- Li, J.; He, W.; Liao, B.; Yang, J. FFA-ROS-P53-mediated mitochondrial apoptosis contributes to reduction of osteoblastogenesis and bone mass in type 2 diabetes mellitus. Sci. Rep. 2015, 5, 12724. [Google Scholar] [CrossRef]
- Park, B.G.; Yoo, C.I.; Kim, H.T.; Kwon, C.H.; Kim, Y.K. Role of mitogen-activated protein kinases in hydrogen peroxide-induced cell death in osteoblastic cells. Toxicology 2005, 215, 115–125. [Google Scholar] [CrossRef]
- Li, X.; Han, Y.; Guan, Y.; Zhang, L.; Bai, C.; Li, Y. Aluminum Induces Osteoblast Apoptosis Through the Oxidative Stress-Mediated JNK Signaling Pathway. Biol. Trace Elem. Res. 2012, 150, 502–508. [Google Scholar] [CrossRef] [PubMed]
- Newsholme, P.; Rebelato, E.; Abdulkader, F.; Krause, M.; Carpinelli, A.; Curi, R. Reactive oxygen and nitrogen species generation, antioxidant defenses, and β-cell function: A critical role for amino acids. J. Endocrinol. 2012, 214, 11–20. [Google Scholar] [CrossRef]
- Fu, J.; Cui, Q.; Yang, B.; Hou, Y.; Wang, H.; Xu, Y.; Wang, D.; Zhang, Q.; Pi, J. The impairment of glucose-stimulated insulin secretion in pancreatic β-cells caused by prolonged glucotoxicity and lipotoxicity is associated with elevated adaptive antioxidant response. Food Chem. Toxicol. 2017, 100, 161–167. [Google Scholar] [CrossRef] [PubMed]
- Rashidi, A.; Kirkwood, T.B.; Shanley, D.P. Metabolic evolution suggests an explanation for the weakness of antioxidant defences in beta-cells. Mech. Ageing Dev. 2009, 130, 216–221. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Wang, X.; Li, Y.-k.; Xue, P. The effects of IRS-1 on osteogenic differentiation and the mechanism. J. Hebei Med. Univ. 2020, 41, 1130–1135. [Google Scholar] [CrossRef]
- Roy, B. Biomolecular basis of the role of diabetes mellitus in osteoporosis and bone fractures. World J. Diabetes 2013, 4, 101–113. [Google Scholar] [CrossRef]
- Agidigbi, T.S.; Kim, C. Reactive Oxygen Species in Osteoclast Differentiation and Possible Pharmaceutical Targets of ROS-Mediated Osteoclast Diseases. Int. J. Mol. Sci. 2019, 20, 3576. [Google Scholar] [CrossRef]
- Ha, H.; Kwak, H.B.; Lee, S.W.; Jin, H.M.; Kim, H.M.; Kim, H.H.; Lee, Z.H. Reactive oxygen species mediate RANK signaling in osteoclasts. Exp. Cell Res. 2004, 301, 119–127. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Dong, G.; Jeon, H.H.; Elazizi, M.; La, L.B.; Hameedaldeen, A.; Xiao, E.; Tian, C.; Alsadun, S.; Choi, Y.; et al. FOXO1 mediates RANKL-induced osteoclast formation and activity. J. Immunol. 2015, 194, 2878–2887. [Google Scholar] [CrossRef]
- Wang, K.; Niu, J.; Kim, H.; Kolattukudy, P.E. Osteoclast precursor differentiation by MCPIP via oxidative stress, endoplasmic reticulum stress, and autophagy. J. Mol. Cell Biol. 2011, 3, 360–368. [Google Scholar] [CrossRef] [PubMed]
- Sul, O.J.; Li, G.; Kim, J.E.; Kim, E.S.; Choi, H.S. 7-ketocholesterol enhances autophagy via the ROS-TFEB signaling pathway in osteoclasts. J. Nutr. Biochem. 2021, 96, 108783. [Google Scholar] [CrossRef] [PubMed]
- Lee, N.K.; Choi, Y.G.; Baik, J.Y.; Han, S.Y.; Jeong, D.-w.; Bae, Y.S.; Kim, N.; Lee, S.Y. A crucial role for reactive oxygen species in RANKL-induced osteoclast differentiation. Blood 2005, 106, 852–859. [Google Scholar] [CrossRef] [PubMed]
- Ikebuchi, Y.; Aoki, S.; Honma, M.; Hayashi, M.; Sugamori, Y.; Khan, M.; Kariya, Y.; Kato, G.; Tabata, Y.; Penninger, J.M.; et al. Coupling of bone resorption and formation by RANKL reverse signalling. Nature 2018, 561, 195–200. [Google Scholar] [CrossRef] [PubMed]
- Flieger, J.; Flieger, W.; Baj, J.; Maciejewski, R. Antioxidants: Classification, Natural Sources, Activity/Capacity Measurements, and Usefulness for the Synthesis of Nanoparticles. Materials 2021, 14, 4135. [Google Scholar] [CrossRef] [PubMed]
- Abedi, N.; Sajadi-Javan, Z.S.; Kouhi, M.; Ansari, L.; Khademi, A.; Ramakrishna, S. Antioxidant Materials in Oral and Maxillofacial Tissue Regeneration: A Narrative Review of the Literature. Antioxidants 2023, 12, 594. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, P.; Kumar, S.; Dutta, D.; Gupta, V. Role of antioxidants in common health diseases. Res. J. Pharm. Technol. 2009, 2, 238–244. [Google Scholar]
- Wang, Y.; Branicky, R.; Noë, A.; Hekimi, S. Superoxide dismutases: Dual roles in controlling ROS damage and regulating ROS signaling. J. Cell Biol. 2018, 217, 1915–1928. [Google Scholar] [CrossRef]
- He, L.; He, T.; Farrar, S.; Ji, L.; Liu, T.; Ma, X. Antioxidants Maintain Cellular Redox Homeostasis by Elimination of Reactive Oxygen Species. Cell Physiol. Biochem. 2017, 44, 532–553. [Google Scholar] [CrossRef] [PubMed]
- Fourquet, S.; Huang, M.E.; D’Autreaux, B.; Toledano, M.B. The dual functions of thiol-based peroxidases in H2O2 scavenging and signaling. Antioxid. Redox Signal. 2008, 10, 1565–1576. [Google Scholar] [CrossRef] [PubMed]
- Uyar, I.S.; Onal, S.; Akpinar, M.B.; Gonen, I.; Sahin, V.; Uguz, A.C.; Burma, O. Alpha lipoic acid attenuates inflammatory response during extracorporeal circulation. Cardiovasc. J. Afr. 2013, 24, 322–326. [Google Scholar] [CrossRef] [PubMed]
- Becker, B.F.; Reinholz, N.; Leipert, B.; Raschke, P.; Permanetter, B.; Gerlach, E. Role of uric acid as an endogenous radical scavenger and antioxidant. Chest 1991, 100, 176s–181s. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.H.; Meza, C.A.; Clarke, H.; Kim, J.S.; Hickner, R.C. Vitamin D and Endothelial Function. Nutrients 2020, 12, 575. [Google Scholar] [CrossRef] [PubMed]
- Gęgotek, A.; Skrzydlewska, E. Ascorbic acid as antioxidant. Vitam. Horm. 2023, 121, 247–270. [Google Scholar] [CrossRef] [PubMed]
- Liebler, D.C. The role of metabolism in the antioxidant function of vitamin E. Crit. Rev. Toxicol. 1993, 23, 147–169. [Google Scholar] [CrossRef] [PubMed]
- Li, M.S.; Adesina, S.E.; Ellis, C.L.; Gooch, J.L.; Hoover, R.S.; Williams, C.R. NADPH oxidase-2 mediates zinc deficiency-induced oxidative stress and kidney damage. Am. J. Physiol. Cell Physiol. 2017, 312, C47–C55. [Google Scholar] [CrossRef] [PubMed]
- Menon, V.P.; Sudheer, A.R. Antioxidant and anti-inflammatory properties of curcumin. In The Molecular Targets and Therapeutic Uses of Curcumin in Health and Disease; Aggarwal, B.B., Surh, Y.-J., Shishodia, S., Eds.; Springer: Boston, MA, USA, 2007; pp. 105–125. [Google Scholar]
- Amalraj, A.; Pius, A.; Gopi, S.; Gopi, S. Biological activities of curcuminoids, other biomolecules from turmeric and their derivatives—A review. J. Tradit. Complement. Med. 2017, 7, 205–233. [Google Scholar] [CrossRef]
- Qiu, L.; Gao, C.; Wang, H.; Ren, Y.; Li, J.; Li, M.; Du, X.; Li, W.; Zhang, J. Effects of dietary polyphenol curcumin supplementation on metabolic, inflammatory, and oxidative stress indices in patients with metabolic syndrome: A systematic review and meta-analysis of randomized controlled trials. Front. Endocrinol. 2023, 14, 1216708. [Google Scholar] [CrossRef]
- Richard, D.; Kefi, K.; Barbe, U.; Bausero, P.; Visioli, F. Polyunsaturated fatty acids as antioxidants. Pharmacol. Res. 2008, 57, 451–455. [Google Scholar] [CrossRef] [PubMed]
- Stoia, M.; Oancea, S. Low-Molecular-Weight Synthetic Antioxidants: Classification, Pharmacological Profile, Effectiveness and Trends. Antioxidants 2022, 11, 638. [Google Scholar] [CrossRef] [PubMed]
- Karageorgiou, V.; Kaplan, D. Porosity of 3D biomaterial scaffolds and osteogenesis. Biomaterials 2005, 26, 5474–5491. [Google Scholar] [CrossRef]
- Huang, X.; Lou, Y.; Duan, Y.; Liu, H.; Tian, J.; Shen, Y.; Wei, X. Biomaterial scaffolds in maxillofacial bone tissue engineering: A review of recent advances. Bioact. Mater. 2024, 33, 129–156. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Wu, G.; Qiu, J. Interactions between cells and biomaterials in tissue engineering: A review. Sheng Wu Gong. Cheng Xue Bao 2021, 37, 2668–2677. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.Z.; Song, L.; Zhang, Z.Y.; Lv, M.M. Control-released Alpha-lipoic acid-loaded PLGA microspheres enhance bone formation in type 2 diabetic rat model. Int. J. Clin. Exp. Pathol. 2017, 10, 10019–10031. [Google Scholar] [PubMed]
- Wu, Y.; Ta, H.T. Different approaches to synthesising cerium oxide nanoparticles and their corresponding physical characteristics, and ROS scavenging and anti-inflammatory capabilities. J. Mater. Chem. B 2021, 9, 7291–7301. [Google Scholar] [CrossRef]
- Li, Y.; Yang, L.; Hou, Y.; Zhang, Z.; Chen, M.; Wang, M.; Liu, J.; Wang, J.; Zhao, Z.; Xie, C.; et al. Polydopamine-mediated graphene oxide and nanohydroxyapatite-incorporated conductive scaffold with an immunomodulatory ability accelerates periodontal bone regeneration in diabetes. Bioact. Mater. 2022, 18, 213–227. [Google Scholar] [CrossRef] [PubMed]
- Cifuentes-Mendiola, S.E.; Moreno-Fierros, L.; González-Alva, P.; García-Hernández, A.L. Docosahexaenoic acid improves altered mineralization proteins, the decreased quality of hydroxyapatite crystals and suppresses oxidative stress induced by high glucose. Exp. Ther. Med. 2022, 23, 235. [Google Scholar] [CrossRef]
- Lin, C.W.; Lee, C.Y.; Lin, S.Y.; Kang, L.; Fu, Y.C.; Chen, C.H.; Wang, C.K. Bone-Targeting Nanoparticles of a Dendritic (Aspartic acid)(3)-Functionalized PEG-PLGA Biopolymer Encapsulating Simvastatin for the Treatment of Osteoporosis in Rat Models. Int. J. Mol. Sci. 2022, 23, 530. [Google Scholar] [CrossRef]
- Hong, S.; Na, Y.S.; Choi, S.; Song, I.T.; Kim, W.Y.; Lee, H. Non-Covalent Self-Assembly and Covalent Polymerization Co-Contribute to Polydopamine Formation. Adv. Funct. Mater. 2012, 22, 4711–4717. [Google Scholar] [CrossRef]
- Li, H.; Xi, J.; Donaghue, A.G.; Keum, J.; Zhao, Y.; An, K.; McKenzie, E.R.; Ren, F. Synthesis and catalytic performance of polydopamine supported metal nanoparticles. Sci. Rep. 2020, 10, 10416. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Yang, L.; Yang, P.; Jiang, S.; Liu, X.; Li, Y. Polydopamine free radical scavengers. Biomater. Sci. 2020, 8, 4940–4950. [Google Scholar] [CrossRef] [PubMed]
- Gaucher, C.; Boudier, A.; Bonetti, J.; Clarot, I.; Leroy, P.; Parent, M. Glutathione: Antioxidant Properties Dedicated to Nanotechnologies. Antioxidants 2018, 7, 62. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Shen, M.; Hou, Q.; Wu, Z.; Xu, J.; Wang, L. 3D printing of reduced glutathione grafted gelatine methacrylate hydrogel scaffold promotes diabetic bone regeneration by activating PI3K/Akt signaling pathway. Int. J. Biol. Macromol. 2022, 222, 1175–1191. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.K.; Yoon, D.S.; Mandakhbayar, N.; Li, C.; Kurian, A.G.; Lee, N.-H.; Lee, J.-H.; Kim, H.-W. Diabetic bone regeneration with nanoceria-tailored scaffolds by recapitulating cellular microenvironment: Activating integrin/TGF-β co-signaling of MSCs while relieving oxidative stress. Biomaterials 2022, 288, 121732. [Google Scholar] [CrossRef] [PubMed]
- Hensarling, R.; Rahane, S.; Leblanc, A.; Sparks, B.; White, E.; Locklin, J.; Patton, D. Thiol-isocyanate “click” reactions: Rapid development of functional polymeric surfaces. Polym. Chem. 2011, 2, 88–90. [Google Scholar] [CrossRef]
- Hoyle, C.E.; Lowe, A.B.; Bowman, C.N. Thiol-click chemistry: A multifaceted toolbox for small molecule and polymer synthesis. Chem. Soc. Rev. 2010, 39, 1355–1387. [Google Scholar] [CrossRef] [PubMed]
- Touchet, T.; Briggs, S.; Graul, L.; Maitland, D.J. Development and Characterization of Oxidatively Responsive Thiol-Ene Networks for Bone Graft Applications. ACS Appl. Bio Mater. 2022, 5, 2633–2642. [Google Scholar] [CrossRef]
- Touchet, T.J.; Horelica, M.; Gruenbaum, R.; Lewy, K.; Hines, E.; Stranahan, L.; Saunders, W.B.; Maitland, D.J. Fabrication and In Vivo Assessment of Oxidatively Responsive PolyHIPE Scaffolds for Use in Diabetic Orthopedic Applications. Macromol. Biosci. 2024, 24, 2300393. [Google Scholar] [CrossRef]
- Boyan, B.D.; Olivares-Navarrete, R.; Berger, M.B.; Hyzy, S.L.; Schwartz, Z. Role of Wnt11 during Osteogenic Differentiation of Human Mesenchymal Stem Cells on Microstructured Titanium Surfaces. Sci. Rep. 2018, 8, 8588. [Google Scholar] [CrossRef]
- Jenkins, T.L.; Little, D. Synthetic scaffolds for musculoskeletal tissue engineering: Cellular responses to fiber parameters. npj Regen. Med. 2019, 4, 15. [Google Scholar] [CrossRef] [PubMed]
- Buser, D.; Schenk, R.K.; Steinemann, S.; Fiorellini, J.P.; Fox, C.H.; Stich, H. Influence of surface characteristics on bone integration of titanium implants. A histomorphometric study in miniature pigs. J. Biomed. Mater. Res. 1991, 25, 889–902. [Google Scholar] [CrossRef] [PubMed]
- Kieswetter, K.; Schwartz, Z.; Hummert, T.W.; Cochran, D.L.; Simpson, J.; Dean, D.D.; Boyan, B.D. Surface roughness modulates the local production of growth factors and cytokines by osteoblast-like MG-63 cells. J. Biomed. Mater. Res. 1996, 32, 55–63. [Google Scholar] [CrossRef]
- Jemat, A.; Ghazali, M.J.; Razali, M.; Otsuka, Y. Surface Modifications and Their Effects on Titanium Dental Implants. Biomed Res. Int. 2015, 2015, 791725. [Google Scholar] [CrossRef]
- Javier Gil, F.; Planell, J.A.; Padrós, A.; Aparicio, C. The effect of shot blasting and heat treatment on the fatigue behavior of titanium for dental implant applications. Dent. Mater. 2007, 23, 486–491. [Google Scholar] [CrossRef] [PubMed]
- Dong, S.; Zhao, T.; Wu, W.; Zhang, Z.; Wu, J.; Cai, K.; Li, G.; Lv, J.; Zhou, H.; Tang, C. Sandblasted/Acid-Etched Titanium Surface Modified with Calcium Phytate Enhances Bone Regeneration in a High-Glucose Microenvironment by Regulating Reactive Oxygen Species and Cell Senescence. ACS Biomater. Sci. Eng. 2023, 9, 4720–4734. [Google Scholar] [CrossRef]
- Awad, N.K.; Edwards, S.L.; Morsi, Y.S. A review of TiO2 NTs on Ti metal: Electrochemical synthesis, functionalization and potential use as bone implants. Mater. Sci. Eng. C 2017, 76, 1401–1412. [Google Scholar] [CrossRef]
- Huang, J.; Zhang, X.; Yan, W.; Chen, Z.; Shuai, X.; Wang, A.; Wang, Y. Nanotubular topography enhances the bioactivity of titanium implants. Nanomed. Nanotechnol. Biol. Med. 2017, 13, 1913–1923. [Google Scholar] [CrossRef]
- Li, Y.; Song, Y.; Ma, A.; Li, C. Surface Immobilization of TiO2 Nanotubes with Bone Morphogenetic Protein-2 Synergistically Enhances Initial Preosteoblast Adhesion and Osseointegration. Biomed Res. Int. 2019, 2019, 5697250. [Google Scholar] [CrossRef]
- Li, Y.; Li, B.; Song, Y.; Ma, A.; Li, C.; Zhang, X.; Li, H.; Zhang, Q.; Zhang, K. Improved osteoblast adhesion and osseointegration on TiO2 nanotubes surface with hydroxyapatite coating. Dent. Mater. J. 2019, 38, 278–286. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Zhang, H.; Chan, S.M.; Li, R.; Wu, Y.; Cai, M.; Wang, A.; Wang, Y. TiO2 Nanotubes Alleviate Diabetes-Induced Osteogenetic Inhibition. Int. J. Nanomed. 2020, 15, 3523–3537. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Li, R.; Yang, J.; Cai, M.; Lee, Y.; Wang, A.; Cheng, B.; Wang, Y. Bioadaptation of implants to In vitro and In vivo oxidative stress pathological conditions via nanotopography-induced FoxO1 signaling pathways to enhance Osteoimmunal regeneration. Bioact. Mater. 2021, 6, 3164–3176. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Huang, S.; Peng, Q. Metal Ion-Doped Hydroxyapatite-Based Materials for Bone Defect Restoration. Bioengineering 2023, 10, 1367. [Google Scholar] [CrossRef] [PubMed]
- Luo, J.; Mamat, B.; Yue, Z.; Zhang, N.; Xu, X.; Li, Y.; Su, Z.; Ma, C.; Zang, F.; Wang, Y. Multi-metal ions doped hydroxyapatite coatings via electrochemical methods for antibacterial and osteogenesis. Colloid. Interface Sci. Commun. 2021, 43, 100435. [Google Scholar] [CrossRef]
- Wang, H.; Fu, X.; Shi, J.; Li, L.; Sun, J.; Zhang, X.; Han, Q.; Deng, Y.; Gan, X. Nutrient Element Decorated Polyetheretherketone Implants Steer Mitochondrial Dynamics for Boosted Diabetic Osseointegration. Adv. Sci. 2021, 8, e2101778. [Google Scholar] [CrossRef] [PubMed]
- Qiu, X.-t.; Rao, C.-y.; Li, T.; Zhou, R.-h. Research Progress in Biomimetic Synthesis of Nano-Hydroxyapatite in Bone Tissue Engineering. J. Sichuan Univ. Med. Sci. 2021, 52, 740–746. [Google Scholar] [CrossRef]
- Li, G.; Zhou, T.; Lin, S.; Shi, S.; Lin, Y. Nanomaterials for Craniofacial and Dental Tissue Engineering. J. Dent. Res. 2017, 96, 725–732. [Google Scholar] [CrossRef] [PubMed]
- Ding, Q.; Cui, J.; Shen, H.; He, C.; Wang, X.; Shen, S.G.F.; Lin, K. Advances of nanomaterial applications in oral and maxillofacial tissue regeneration and disease treatment. WIREs Nanomed. Nanobiotechnol. 2021, 13, e1669. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Li, X.; Li, J.; Li, D.; Wang, Q.; Teng, W. Dopamine-assisted co-deposition of hydroxyapatite-functionalised nanoparticles of polydopamine on implant surfaces to promote osteogenesis in environments with high ROS levels. Mater. Sci. Eng. C Mater. Biol. Appl. 2021, 131, 112473. [Google Scholar] [CrossRef]
- Tao, Z.S.; Wang, H.S.; Li, T.L.; Wei, S. Silibinin-modified Hydroxyapatite coating promotes the osseointegration of titanium rods by activation SIRT1/SOD2 signaling pathway in diabetic rats. J. Mater. Sci. Mater. Med. 2022, 33, 62. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.Y.; Cui, D.; Wang, Z.; Liu, B.; Yu, H.L.; Yuan, H.; Xiang, L.B.; Zhou, D.P. Silk Fibroin/Hydroxyapatite Coating Improved Osseointegration of Porous Titanium Implants under Diabetic Conditions via Activation of the PI3K/Akt Signaling Pathway. ACS Biomater. Sci. Eng. 2022, 8, 2908–2919. [Google Scholar] [CrossRef] [PubMed]
- Di Martino, A.; Sittinger, M.; Risbud, M.V. Chitosan: A versatile biopolymer for orthopaedic tissue-engineering. Biomaterials 2005, 26, 5983–5990. [Google Scholar] [CrossRef] [PubMed]
- Xie, W.; Xu, P.; Liu, Q. Antioxidant activity of water-soluble chitosan derivatives. Bioorg. Med. Chem. Lett. 2001, 11, 1699–1701. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Xu, Q.; Chen, Y.; Wan, A. Effect of concentration and molecular weight of chitosan and its derivative on the free radical scavenging ability. J. Biomed. Mater. Res. A 2014, 102, 911–916. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Ma, X.-Y.; Feng, Y.-F.; Ma, Z.-S.; Wang, J.; Ma, T.-C.; Qi, W.; Lei, W.; Wang, L. Osseointegration of chitosan coated porous titanium alloy implant by reactive oxygen species-mediated activation of the PI3K/AKT pathway under diabetic conditions. Biomaterials 2015, 36, 44–54. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.Y.; Feng, Y.F.; Wang, T.S.; Lei, W.; Li, X.; Zhou, D.P.; Wen, X.X.; Yu, H.L.; Xiang, L.B.; Wang, L. Involvement of FAK-mediated BMP-2/Smad pathway in mediating osteoblast adhesion and differentiation on nano-HA/chitosan composite coated titanium implant under diabetic conditions. Biomater. Sci. 2017, 6, 225–238. [Google Scholar] [CrossRef] [PubMed]
- Khan, I.; Saeed, K.; Khan, I. Nanoparticles: Properties, applications and toxicities. Arab. J. Chem. 2019, 12, 908–931. [Google Scholar] [CrossRef]
- Radulescu, D.M.; Surdu, V.A.; Ficai, A.; Ficai, D.; Grumezescu, A.M.; Andronescu, E. Green Synthesis of Metal and Metal Oxide Nanoparticles: A Review of the Principles and Biomedical Applications. Int. J. Mol. Sci. 2023, 24, 5397. [Google Scholar] [CrossRef]
- Yusof, H.M.; Mohamad, R.; Zaidan, U.H. Microbial synthesis of zinc oxide nanoparticles and their potential application as an antimicrobial agent and a feed supplement in animal industry: A review. J. Anim. Sci. Biotechnol. 2019, 10, 57. [Google Scholar]
- Steinbrenner, H.; Sies, H. Protection against reactive oxygen species by selenoproteins. Biochim. Biophys. Acta 2009, 1790, 1478–1485. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Bian, W.; Liu, S.; Huang, K. Selenium protects bone marrow stromal cells against hydrogen peroxide-induced inhibition of osteoblastic differentiation by suppressing oxidative stress and ERK signaling pathway. Biol. Trace Elem. Res. 2012, 150, 441–450. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Liang, Z.; Su, Z.; Huang, J.; Pi, Y.; Ouyang, Y.; Luo, T.; Guo, L. Selenium-Doped Mesoporous Bioactive Glass Regulates Macrophage Metabolism and Polarization by Scavenging ROS and Promotes Bone Regeneration In Vivo. ACS Appl. Mater. Interfaces 2023, 15, 34378–34396. [Google Scholar] [CrossRef] [PubMed]
- Poleboina, S.; Sheth, V.G.; Sharma, N.; Sihota, P.; Kumar, N.; Tikoo, K. Selenium nanoparticles stimulate osteoblast differentiation via BMP-2/MAPKs/β-catenin pathway in diabetic osteoporosis. Nanomedicine 2022, 17, 607–625. [Google Scholar] [CrossRef] [PubMed]
- Hu, W.; Yie, K.H.R.; Liu, C.; Zhu, J.; Huang, Z.; Zhu, B.; Zheng, D.; Yang, B.; Huang, B.; Yao, L.; et al. Improving the valence self-reversible conversion of cerium nanoparticles on titanium implants by lanthanum doping to enhance ROS elimination and osteogenesis. Dent. Mater. 2022, 38, 1362–1375. [Google Scholar] [CrossRef] [PubMed]
- Thakur, N.; Manna, P.; Das, J. Synthesis and biomedical applications of nanoceria, a redox active nanoparticle. J. Nanobiotechnol. 2019, 17, 84. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.H.; Kim, J.S.; Kim, J.E.; Lee, M.H.; Jeon, J.G.; Park, I.S.; Yi, H.K. Nanoparticle mediated PPARγ gene delivery on dental implants improves osseointegration via mitochondrial biogenesis in diabetes mellitus rat model. Nanomedicine 2017, 13, 1821–1832. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Tian, T.; Zhou, R.; Li, S.; Ma, W.; Zhang, Y.; Liu, N.; Shi, S.; Li, Q.; Xie, X.; et al. Design, fabrication and applications of tetrahedral DNA nanostructure-based multifunctional complexes in drug delivery and biomedical treatment. Nat. Protoc. 2020, 15, 2728–2757. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Xie, X.; Ma, W.; Zhan, Y.; Mao, C.; Shao, X.; Lin, Y. Multi-targeted Antisense Oligonucleotide Delivery by a Framework Nucleic Acid for Inhibiting Biofilm Formation and Virulence. Nanomicro Lett. 2020, 12, 74. [Google Scholar] [CrossRef]
- Li, S.; Liu, Y.; Zhang, T.; Lin, S.; Shi, S.; He, J.; Xie, Y.; Cai, X.; Tian, T.; Lin, Y. A Tetrahedral Framework DNA-Based Bioswitchable miRNA Inhibitor Delivery System: Application to Skin Anti-Aging. Adv. Mater. 2022, 34, e2204287. [Google Scholar] [CrossRef]
- Li, Y.; Tang, Y.; Shi, S.; Gao, S.; Wang, Y.; Xiao, D.; Chen, T.; He, Q.; Zhang, J.; Lin, Y. Tetrahedral Framework Nucleic Acids Ameliorate Insulin Resistance in Type 2 Diabetes Mellitus via the PI3K/Akt Pathway. ACS Appl. Mater. Interfaces 2021, 13, 40354–40364. [Google Scholar] [CrossRef]
- Li, Y.; Cai, Z.; Ma, W.; Bai, L.; Luo, E.; Lin, Y. A DNA tetrahedron-based ferroptosis-suppressing nanoparticle: Superior delivery of curcumin and alleviation of diabetic osteoporosis. Bone Res. 2024, 12, 14. [Google Scholar] [CrossRef] [PubMed]
- Bai, X.; Gao, M.; Syed, S.; Zhuang, J.; Xu, X.; Zhang, X.-Q. Bioactive hydrogels for bone regeneration. Bioact. Mater. 2018, 3, 401–417. [Google Scholar] [CrossRef]
- Liu, T.; Weng, W.; Zhang, Y.; Sun, X.; Yang, H. Applications of Gelatin Methacryloyl (GelMA) Hydrogels in Microfluidic Technique-Assisted Tissue Engineering. Molecules 2020, 25, 5305. [Google Scholar] [CrossRef]
- Liao, X.; Shen, M.; Li, T.; Feng, L.; Lin, Z.; Shi, G.; Pei, G.; Cai, X. Combined Molybdenum Gelatine Methacrylate Injectable Nano-Hydrogel Effective Against Diabetic Bone Regeneration. Int. J. Nanomed. 2023, 18, 5925–5942. [Google Scholar] [CrossRef]
- Yoval-Sánchez, B.; Ansari, F.; Lange, D.; Galkin, A. Effect of metformin on intact mitochondria from liver and brain: Concept revisited. Eur. J. Pharmacol. 2022, 931, 175177. [Google Scholar] [CrossRef]
- Batandier, C.; Guigas, B.; Detaille, D.; El-Mir, M.Y.; Fontaine, E.; Rigoulet, M.; Leverve, X.M. The ROS production induced by a reverse-electron flux at respiratory-chain complex 1 is hampered by metformin. J. Bioenerg. Biomembr. 2006, 38, 33–42. [Google Scholar] [CrossRef] [PubMed]
- Jiating, L.; Buyun, J.; Yinchang, Z. Role of Metformin on Osteoblast Differentiation in Type 2 Diabetes. Biomed Res Int 2019, 2019, 9203934. [Google Scholar] [CrossRef] [PubMed]
- Lao, A.; Wu, J.; Li, D.; Shen, A.; Li, Y.; Zhuang, Y.; Lin, K.; Wu, J.; Liu, J. Functionalized Metal-Organic Framework-Modified Hydrogel That Breaks the Vicious Cycle of Inflammation and ROS for Repairing of Diabetic Bone Defects. Small 2023, 19, e2206919. [Google Scholar] [CrossRef]
- Li, D.; Chen, K.; Tang, H.; Hu, S.; Xin, L.; Jing, X.; He, Q.; Wang, S.; Song, J.; Mei, L.; et al. A Logic-Based Diagnostic and Therapeutic Hydrogel with Multistimuli Responsiveness to Orchestrate Diabetic Bone Regeneration. Adv. Mater. 2022, 34, 2108430. [Google Scholar] [CrossRef]
- Wang, H.; Chang, X.; Ma, Q.; Sun, B.; Li, H.; Zhou, J.; Hu, Y.; Yang, X.; Li, J.; Chen, X.; et al. Bioinspired drug-delivery system emulating the natural bone healing cascade for diabetic periodontal bone regeneration. Bioact. Mater. 2023, 21, 324–339. [Google Scholar] [CrossRef] [PubMed]
- Andreyev, A.Y.; Kushnareva, Y.E.; Starkova, N.N.; Starkov, A.A. Metabolic ROS Signaling: To Immunity and Beyond. Biochemistry (Moscow) 2020, 85, 1650–1667. [Google Scholar] [CrossRef] [PubMed]
- McKeegan, K.; Mason, S.A.; Trewin, A.J.; Keske, M.A.; Wadley, G.D.; Della Gatta, P.A.; Nikolaidis, M.G.; Parker, L. Reactive oxygen species in exercise and insulin resistance: Working towards personalized antioxidant treatment. Redox Biol. 2021, 44, 102005. [Google Scholar] [CrossRef] [PubMed]
- Zhong, O.; Hu, J.; Wang, J.; Tan, Y.; Hu, L.; Lei, X. Antioxidant for treatment of diabetic complications: A meta-analysis and systematic review. J. Biochem. Mol. Toxicol. 2022, 36, e23038. [Google Scholar] [CrossRef] [PubMed]
- Rahimi-Madiseh, M.; Malekpour-Tehrani, A.; Bahmani, M.; Rafieian-Kopaei, M. The research and development on the antioxidants in prevention of diabetic complications. Asian Pac. J. Trop. Med. 2016, 9, 825–831. [Google Scholar] [CrossRef] [PubMed]
- Duvvuri, L.S.; Katiyar, S.; Kumar, A.; Khan, W. Delivery aspects of antioxidants in diabetes management. Expert. Opin. Drug Deliv. 2015, 12, 827–844. [Google Scholar] [CrossRef] [PubMed]
- Mura, S.; Nicolas, J.; Couvreur, P. Stimuli-responsive nanocarriers for drug delivery. Nat. Mater. 2013, 12, 991–1003. [Google Scholar] [CrossRef]
- Yang, B.; Chen, Y.; Shi, J. Reactive Oxygen Species (ROS)-Based Nanomedicine. Chem. Rev. 2019, 119, 4881–4985. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, M.; Zhao, Z.; Yi, J. Biomaterials Designed to Modulate Reactive Oxygen Species for Enhanced Bone Regeneration in Diabetic Conditions. J. Funct. Biomater. 2024, 15, 220. https://doi.org/10.3390/jfb15080220
Li M, Zhao Z, Yi J. Biomaterials Designed to Modulate Reactive Oxygen Species for Enhanced Bone Regeneration in Diabetic Conditions. Journal of Functional Biomaterials. 2024; 15(8):220. https://doi.org/10.3390/jfb15080220
Chicago/Turabian StyleLi, Mingshan, Zhihe Zhao, and Jianru Yi. 2024. "Biomaterials Designed to Modulate Reactive Oxygen Species for Enhanced Bone Regeneration in Diabetic Conditions" Journal of Functional Biomaterials 15, no. 8: 220. https://doi.org/10.3390/jfb15080220