In Vitro Findings of Titanium Functionalized with Estradiol via Polydopamine Adlayer




Abstract
:1. Introduction
2. Results
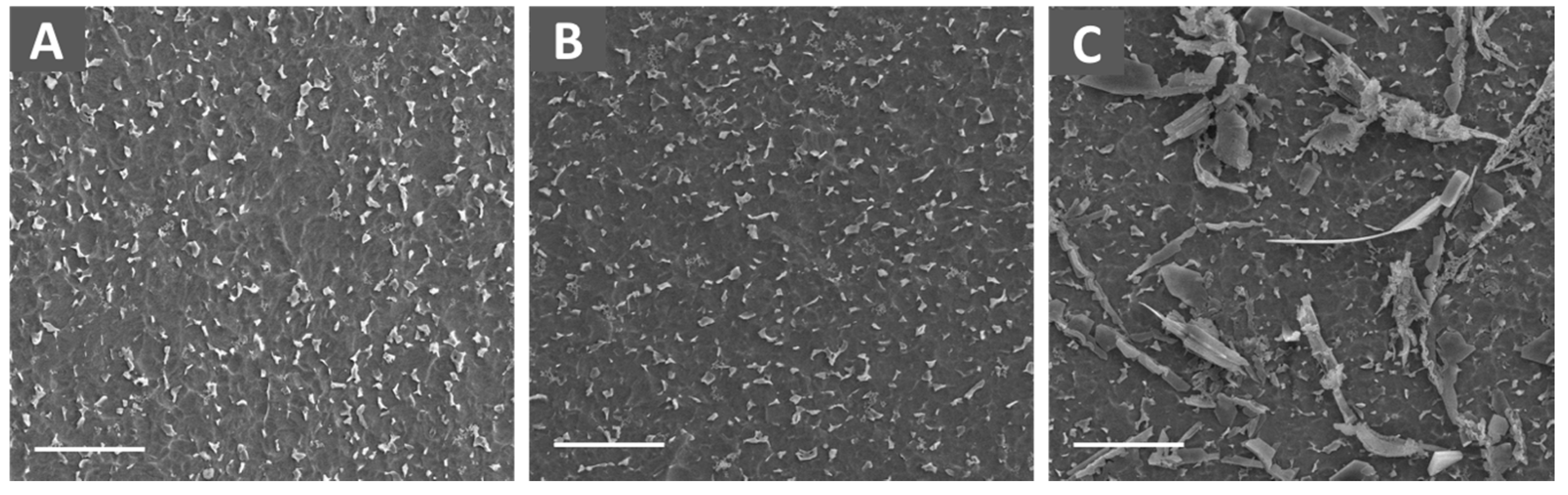
2.1. Surface Characterization
2.2. E2 Release
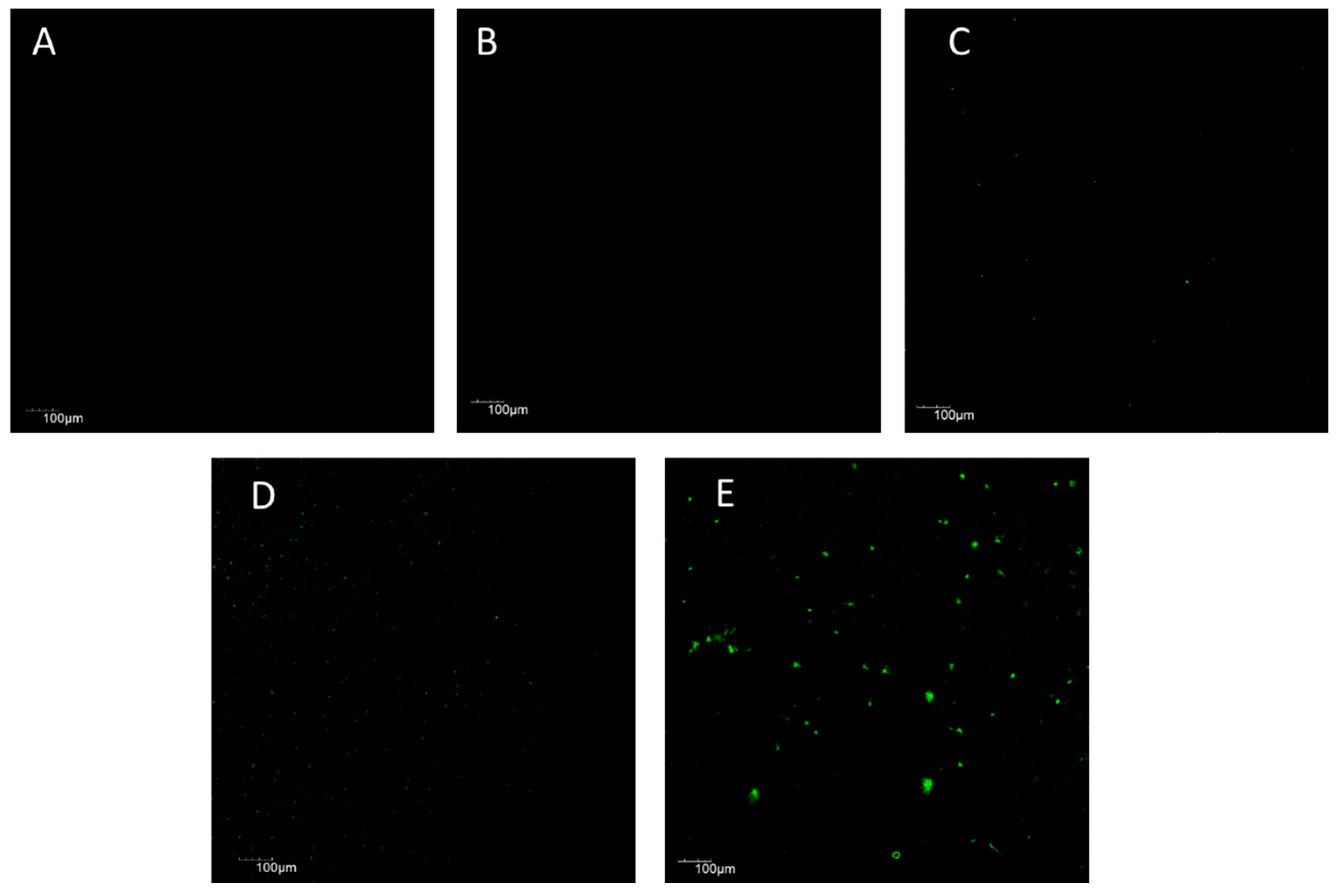
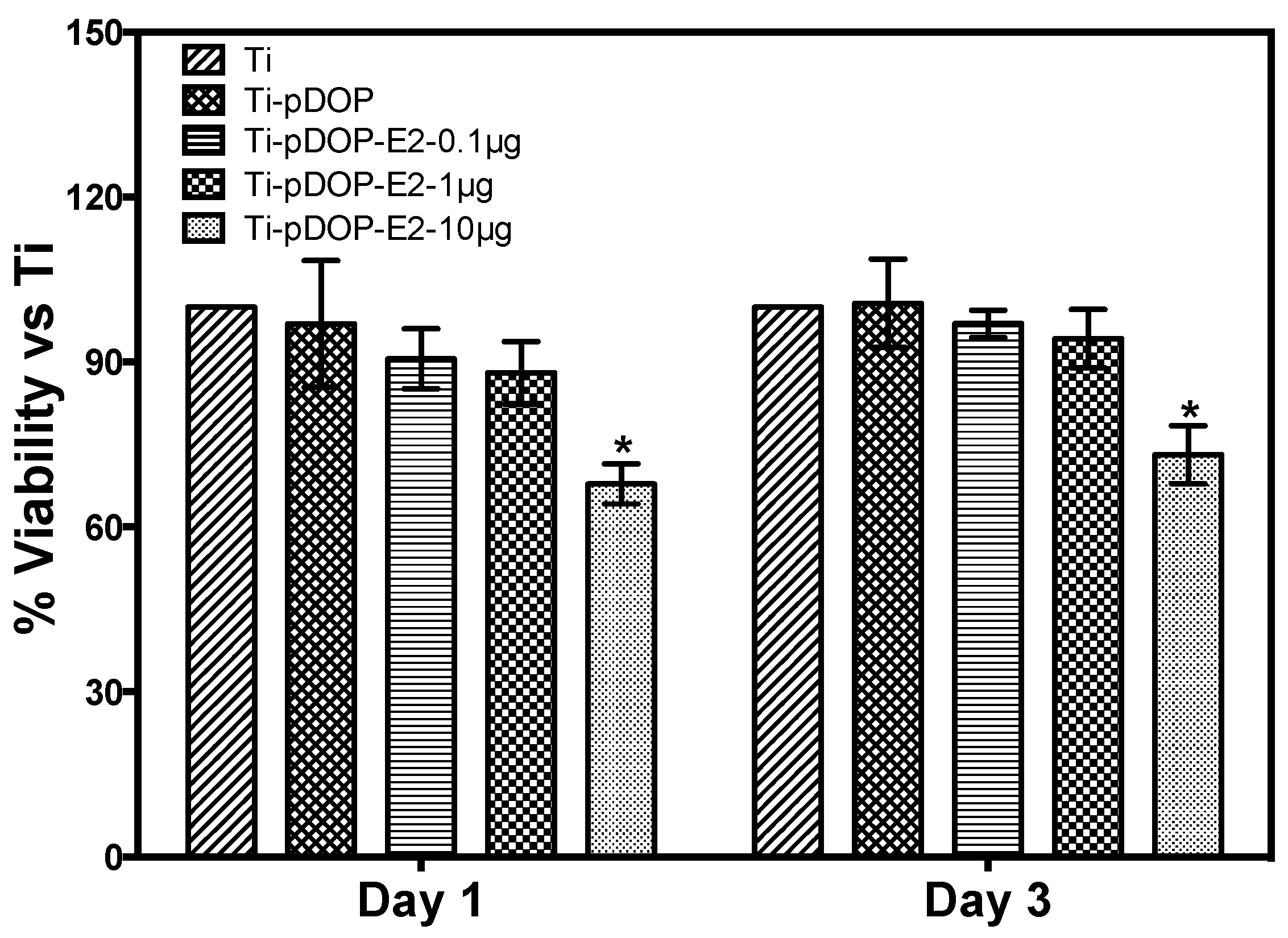
2.3. Cell Cytotoxicity Assay
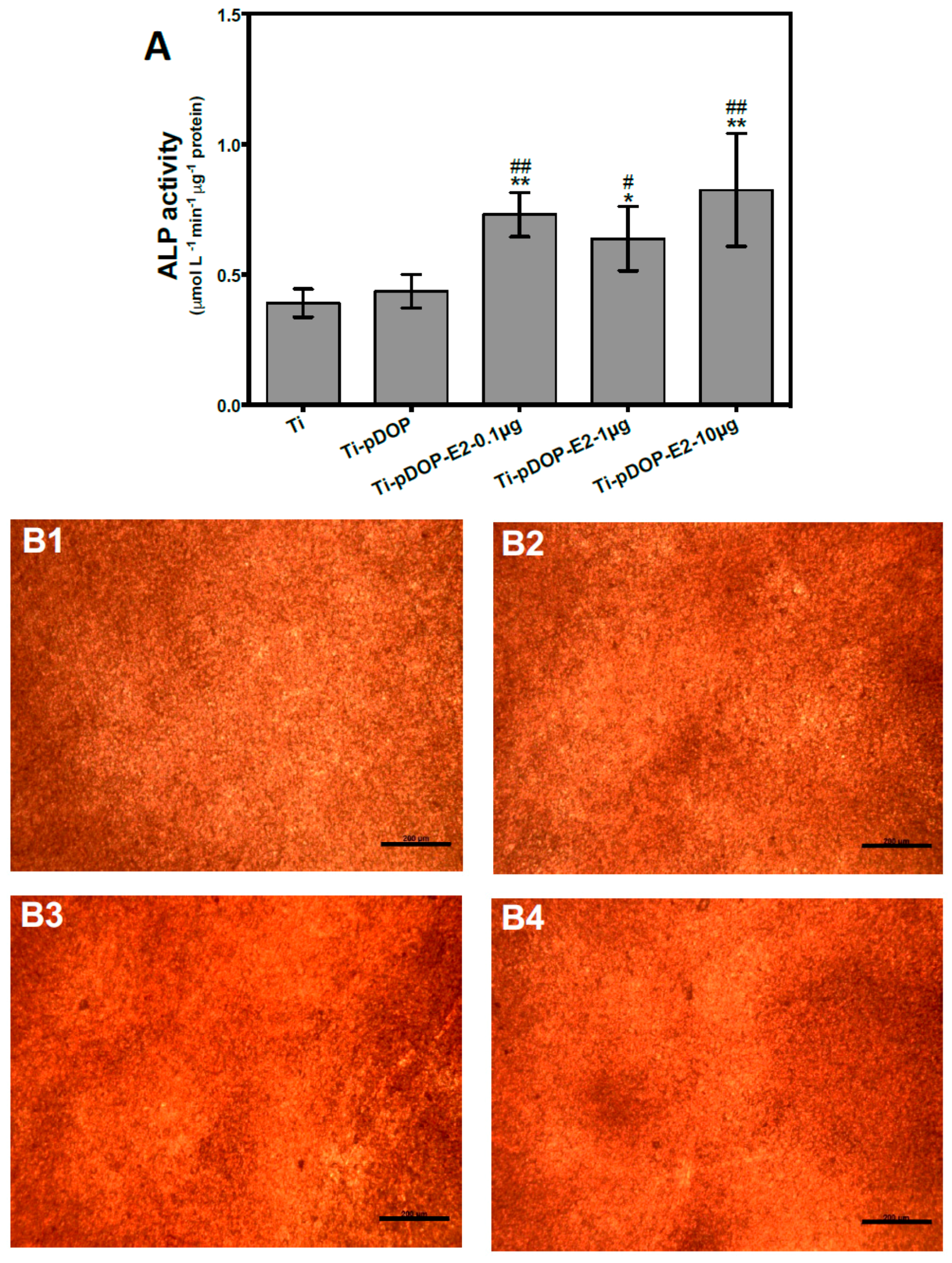
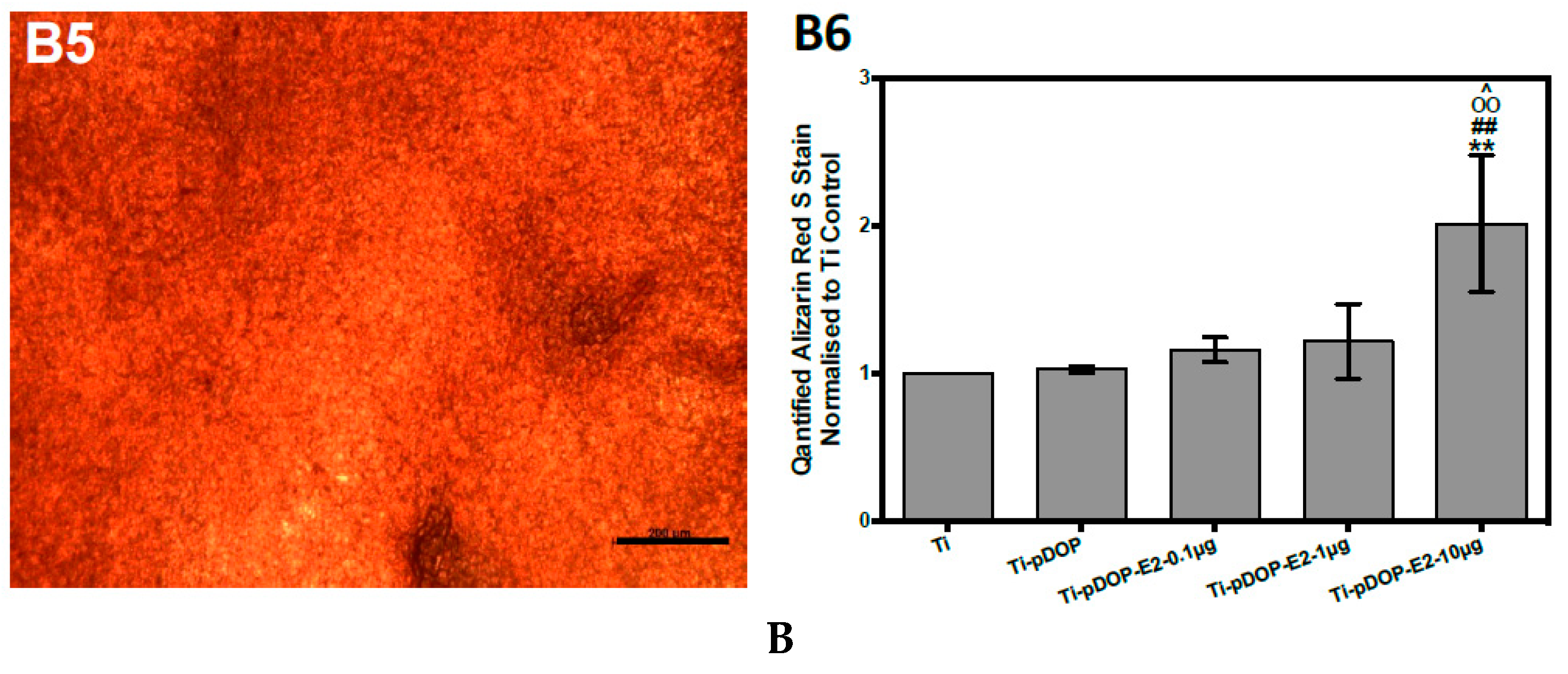
2.4. Osteoblast Differentiation and Calcium Deposition
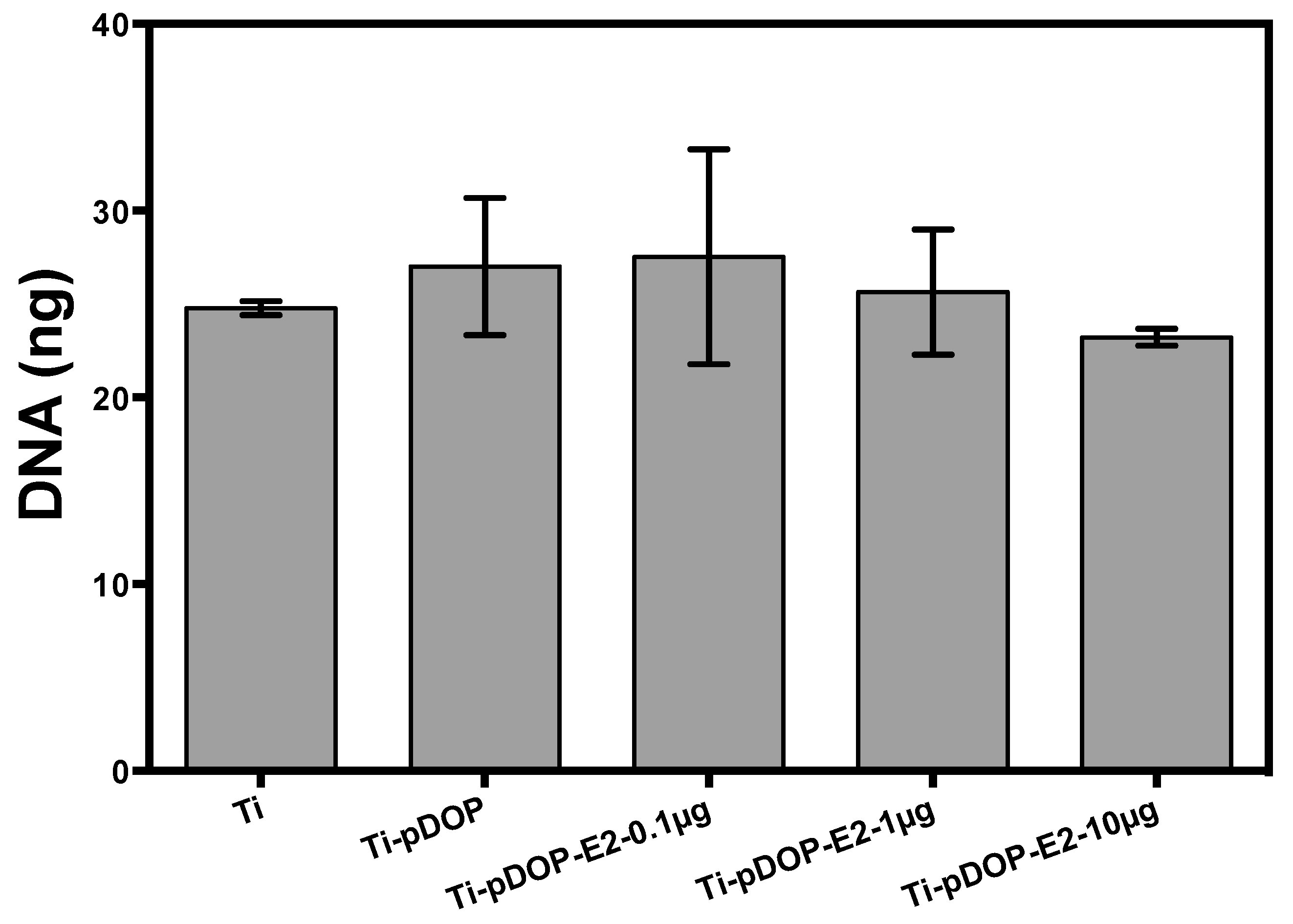
2.5. Total DNA of Osteoclast
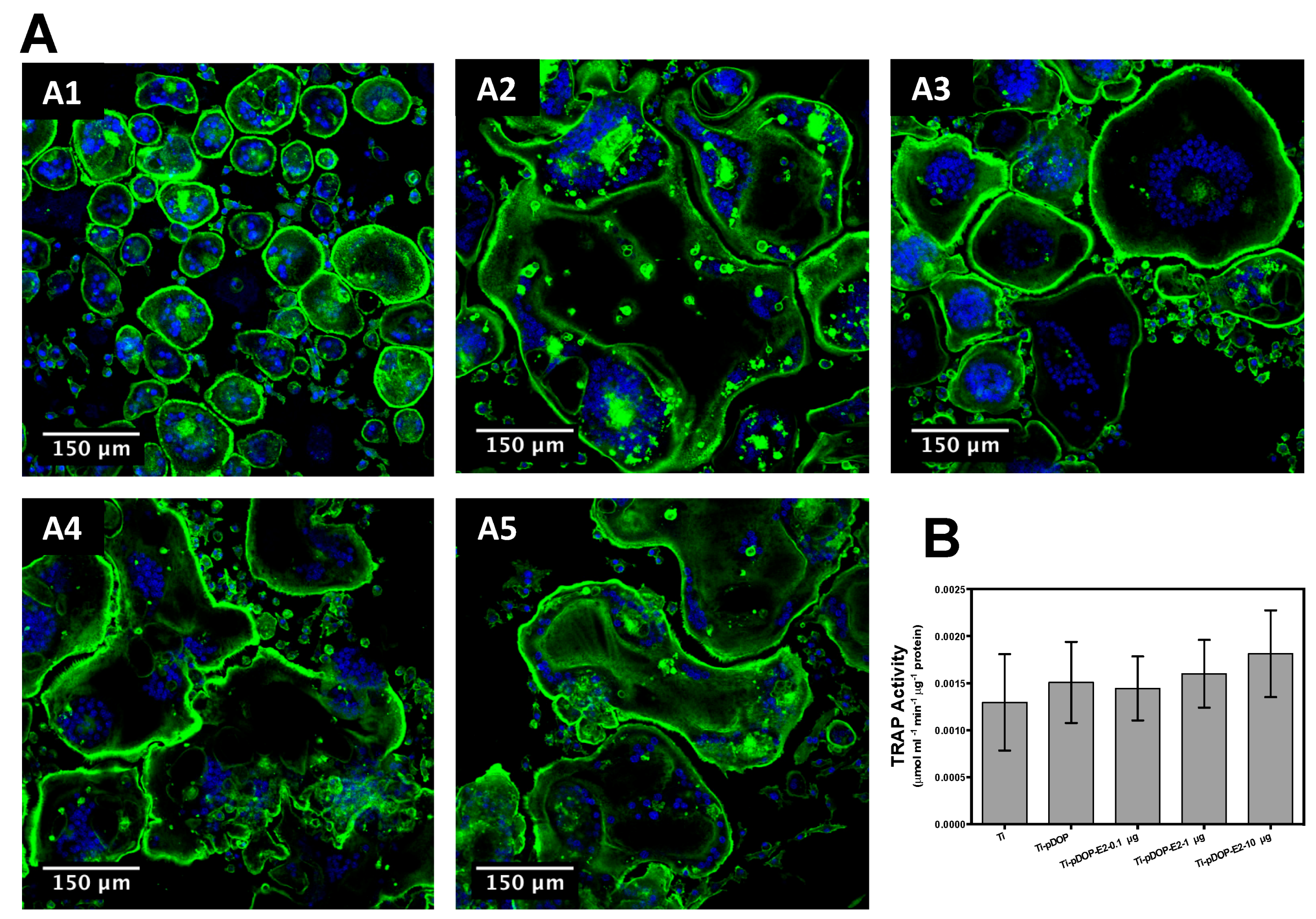
2.6. Osteoclast Actin Ring Formation and TRAP Activity
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Substrate Preparation
4.3. Characterization
4.4. E2 Release Assay
4.5. Cell Culture of MC3T3-E1 and RAW 264.7 Cells
4.6. Cytotoxicity Assay
4.7. ALP Activity Assay
4.8. Calcium Deposition
4.9. DNA Quantification
4.10. TRAP Activity of Osteoclasts
4.11. Actin Ring Staining
4.12. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Luisi, S.; Orlandini, C.; Biliotti, G.; Scolaro, V.; De Felice, G.; Regini, C.; Petraglia, F. Hormone replacement therapy in menopause and in premature ovarian insufficiency. Min. Ginecol. 2013, 65, 607–620. [Google Scholar]
- Cummings, S.R.; Browner, W.S.; Bauer, D.; Stone, K.; Ensrud, K.; Jamal, S.; Ettinger, B. Endogenous hormones and the risk of hip and vertebral fractures among older women. N. Engl. J. Med. 1998, 339, 733–738. [Google Scholar] [CrossRef] [PubMed]
- Manson, J.E.; Chlebowski, R.T.; Stefanick, M.L.; Aragaki, A.K.; Rossouw, J.E.; Prentice, R.L.; Anderson, G.; Howard, B.V.; Thomson, C.A.; LaCroix, A.Z. Menopausal hormone therapy and health outcomes during the intervention and extended poststopping phases of the women’s health initiative randomized trials. JAMA 2013, 310, 1353–1368. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, T.; Imai, Y.; Matsumoto, T.; Sato, S.; Takeuchi, K.; Igarashi, K.; Harada, Y.; Azuma, Y.; Krust, A.; Yamamoto, Y. Estrogen prevents bone loss via estrogen receptor α and induction of fas ligand in osteoclasts. Cell 2007, 130, 811–823. [Google Scholar] [CrossRef] [PubMed]
- Eghbali-Fatourechi, G.; Khosla, S.; Sanyal, A.; Boyle, W.J.; Lacey, D.L.; Riggs, B.L. Role of RANK ligand in mediating increased bone resorption in early postmenopausal women. J. Clin. Investig. 2003, 111, 1221–1230. [Google Scholar] [CrossRef] [PubMed]
- Hofbauer, L.C.; Khosla, S.; Dunstan, C.R.; Lacey, D.L.; Spelsberg, T.C.; Riggs, B.L. Estrogen stimulates gene expression and protein production of osteoprotegerin in human osteoblastic cells. Endocrinology 1999, 140, 4367–4370. [Google Scholar] [CrossRef] [PubMed]
- Chism, L.A. Overcoming resistance and barriers to the use of local estrogen therapy for the treatment of vaginal atrophy. Int. J. Womens Health 2012, 4, 551–557. [Google Scholar] [CrossRef] [PubMed]
- Moroni, A.; Faldini, C.; Marchetti, S.; Manca, M.; Consoli, V.; Giannini, S. Improvement of the bone-pin interface strength in osteoporotic bone with use of hydroxyapatite-coated tapered external-fixation pins. J. Bone Jt. Surg. 2001, 83, 717–721. [Google Scholar] [CrossRef]
- Tengvall, P.; Skoglund, B.; Askendal, A.; Aspenberg, P. Surface immobilized bisphosphonate improves stainless-steel screw fixation in rats. Biomaterials 2004, 25, 2133–2138. [Google Scholar] [CrossRef] [PubMed]
- Lenart, B.A.; Lorich, D.G.; Lane, J.M. Atypical Fractures of the Femoral Diaphysis in Postmenopausal Women Taking Alendronate. N. Engl. J. Med. 2008, 358, 1304–1306. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Cai, K.; Luo, Z.; Jandt, K.D. Layer-By-Layer Assembly of β-estradiol loaded mesoporous silica nanoparticles on titanium substrates and its implication for bone homeostasis. Adv. Mater. 2010, 22, 4146–4150. [Google Scholar] [CrossRef] [PubMed]
- Mamaeva, V.; Sahlgren, C.; Lindén, M. Mesoporous silica nanoparticles in medicine—Recent advances. Adv. Drug Deliv. Rev. 2013, 65, 689–702. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.; Dellatore, S.M.; Miller, W.M.; Messersmith, P.B. Mussel-Inspired surface chemistry for multifunctional coatings. Science 2007, 318, 426–430. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.; Na, Y.S.; Choi, S.; Song, I.T.; Kim, W.Y.; Lee, H. Non-covalent self-assembly and covalent polymerization co-contribute to polydopamine formation. Adv. Funct. Mater. 2012, 22, 4711–4717. [Google Scholar] [CrossRef]
- Poh, C.K.; Shi, Z.; Lim, T.Y.; Neoh, K.G.; Wang, W. The effect of VEGF functionalization of titanium on endothelial cells in vitro. Biomaterials 2010, 31, 1578–1585. [Google Scholar] [CrossRef] [PubMed]
- He, R.; Hu, X.; Tan, H.C.; Feng, J.; Steffi, C.; Wang, K.; Wang, W. Surface modification of titanium with curcumin: A promising strategy to combat fibrous encapsulation. J. Mater. Chem. B 2015, 3, 2137–2146. [Google Scholar] [CrossRef]
- Chien, C.Y.; Tsai, W.B. Poly(dopamine)-assisted immobilization of Arg-Gly-Asp peptides, hydroxyapatite, and bone morphogenic protein-2 on titanium to improve the osteogenesis of bone marrow stem cells. ACS Appl. Mater. Interfaces 2013, 5, 6975–6983. [Google Scholar] [CrossRef] [PubMed]
- Nanci, A.; Wuest, J.; Peru, L.; Brunet, P.; Sharma, V.; Zalzal, S.; McKee, M. Chemical modification of titanium surfaces for covalent attachment of biological molecules. J. Biomed. Mater. Res. A 1998, 40, 324–335. [Google Scholar] [CrossRef]
- Morishima, F.; Inokuchi, Y.; Ebata, T. Laser Spectroscopic Study of β-estradiol and its monohydrated clusters in a supersonic jet. J. Phys. Chem. A 2012, 116, 8201–8208. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Taw, J.; Chiang, C.-M. Control of drug crystallization in transdermal matrix system. Int. J. Pharm. 1996, 142, 115–119. [Google Scholar] [CrossRef]
- Needham, G.F.; Pfeiffer, R.R.; Engel, G.L.; Rutherford, B.S.; Allen, D.J. Effect of impurities on estradiol crystallization in a sustained-release implant. J. Pharm. Sci. 1992, 81, 1012–1014. [Google Scholar] [CrossRef] [PubMed]
- Lipp, R.; Müller-Fahrnow, A. Use of X-ray crystallography for the characterization of single crystals grown in steroid containing transdermal drug delivery systems. Eur. J. Pharm. Biopharm. 1999, 47, 133–138. [Google Scholar] [CrossRef]
- Czekanska, E.M.; Stoddart, M.J.; Richards, R.G.; Hayes, J.S. In search of an osteoblast cell model for in vitro research. Eur. Cell Mater. 2012, 24, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Quarles, L.D.; Yohay, D.A.; Lever, L.W.; Caton, R.; Wenstrup, R.J. Distinct proliferative and differentiated stages of murine MC3T3-E1 cells in culture: An in vitro model of osteoblast development. J. Bone Miner. Res. 1992, 7, 683–692. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Garner, S.C.; Quarles, L.D.; Anderson, J.J. Effects of genistein on expression of bone markers during MC3T3-E1 osteoblastic cell differentiation. J. Nutr. Biochem. 2003, 14, 342–349. [Google Scholar] [CrossRef]
- Ikegami, A.; Inoue, S.; Hosoi, T.; Mizuno, Y.; Nakamura, T.; Ouchi, Y.; Orimo, H. Immunohistochemical detection and northern blot analysis of estrogen receptor in osteoblastic cells. J. Bone Miner. Res. 1993, 8, 1103–1109. [Google Scholar] [CrossRef] [PubMed]
- Kanatani, M.; Sugimoto, T.; Kano, J.; Fukase, M.; Fujita, T. Effect of 17 β-estradiol on the proliferation of osteoblastic MC3T3-E1 cells via human monocytes. Biochem. Biophys. Res. Commun. 1991, 178, 866–870. [Google Scholar] [CrossRef]
- Shang, Z.-Z.; Li, X.; Sun, H.-Q.; Xiao, G.-N.; Wang, C.-W.; Gong, Q. Differentially expressed genes and signalling pathways are involved in mouse osteoblast-like MC3T3-E1 cells exposed to 17-[bgr] estradiol. Int. J. Oral Sci. 2014, 6, 142–149. [Google Scholar] [CrossRef] [PubMed]
- Granéli, C.; Thorfve, A.; Ruetschi, U.; Brisby, H.; Thomsen, P.; Lindahl, A.; Karlsson, C. Novel markers of osteogenic and adipogenic differentiation of human bone marrow stromal cells identified using a quantitative proteomics approach. Stem. Cell Res. 2014, 12, 153–165. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.-M.; Shih, L.-H.; Lee, J.-Y.; Shen, Y.-J.; Lee, H.-H. Estrogen Enhances activity of wnt signaling during osteogenesis by inducing fhl1 expression. J. Cell Biochem. 2015, 116, 1419–1430. [Google Scholar] [CrossRef] [PubMed]
- Kousteni, S.; Almeida, M.; Han, L.; Bellido, T.; Jilka, R.L.; Manolagas, S.C. Induction of osteoblast differentiation by selective activation of kinase-mediated actions of the estrogen receptor. Mol. Cell. Biol. 2007, 27, 1516–1530. [Google Scholar] [CrossRef] [PubMed]
- Gu, Y.X.; Du, J.; Si, M.S.; Mo, J.J.; Qiao, S.C.; Lai, H.C. The roles of PI3K/Akt signaling pathway in regulating MC3T3-E1 preosteoblast proliferation and differentiation on SLA and SLActive titanium surfaces. J. Biomed. Mater. Res. A 2013, 101, 748–754. [Google Scholar] [CrossRef] [PubMed]
- Puchtler, H.; Meloan, S.N.; Terry, M.S. On the history and mechanism of alizarin and alizarin red S stains for calcium. J. Histochem. Cytochem. 1969, 17, 110–124. [Google Scholar] [CrossRef] [PubMed]
- Vincent, C.; Kogawa, M.; Findlay, D.M.; Atkins, G.J. The generation of osteoclasts from RAW 264.7 precursors in defined, serum-free conditions. J. Bone Miner. Metab. 2009, 27, 114–119. [Google Scholar] [CrossRef] [PubMed]
- Collin-Osdoby, P.; Osdoby, P. RANKL-mediated osteoclast formation from murine RAW 264.7 cells. Methods Mol. Biol. 2012, 816, 187–202. [Google Scholar] [PubMed]
- Krum, S.A.; Miranda-Carboni, G.A.; Hauschka, P.V.; Carroll, J.S.; Lane, T.F.; Freedman, L.P.; Brown, M. Estrogen protects bone by inducing Fas ligand in osteoblasts to regulate osteoclast survival. EMBO J. 2008, 27, 535–545. [Google Scholar] [CrossRef] [PubMed]
- Jones, G.L.; Motta, A.; Marshall, M.J.; El Haj, A.J.; Cartmell, S.H. Osteoblast: Osteoclast co-cultures on silk fibroin, chitosan and PLLA films. Biomaterials 2009, 30, 5376–5384. [Google Scholar] [CrossRef] [PubMed]
- Rose, F.R.; Cyster, L.A.; Grant, D.M.; Scotchford, C.A.; Howdle, S.M.; Shakesheff, K.M. In vitro assessment of cell penetration into porous hydroxyapatite scaffolds with a central aligned channel. Biomaterials 2004, 25, 5507–5514. [Google Scholar] [CrossRef] [PubMed]
- Shevde, N.K.; Bendixen, A.C.; Dienger, K.M.; Pike, J.W. Estrogens suppress RANK ligand-induced osteoclast differentiation via a stromal cell independent mechanism involving c-Jun repression. Proc. Natl. Acad. Sci. USA 2000, 97, 7829–7834. [Google Scholar] [CrossRef] [PubMed]
- Robinson, L.J.; Yaroslavskiy, B.B.; Griswold, R.D.; Zadorozny, E.V.; Guo, L.; Tourkova, I.L.; Blair, H.C. Estrogen inhibits RANKL-stimulated osteoclastic differentiation of human monocytes through estrogen and RANKL-regulated interaction of estrogen receptor-α with BCAR1 and Traf6. Exp. Cell Res. 2009, 315, 1287–1301. [Google Scholar] [CrossRef] [PubMed]
- Lv, Y.; Wang, G.; Xu, W.; Tao, P.; Lv, X.; Wang, Y. Tartrate-resistant acid phosphatase 5b is a marker of osteoclast number and volume in RAW 264.7 cells treated with receptor-activated nuclear κB ligand. Exp. Ther. Med. 2015, 9, 143–146. [Google Scholar] [CrossRef] [PubMed]
- Ballanti, P.; Minisola, S.; Pacitti, M.T.; Scarnecchia, L.; Rosso, R.; Mazzuoli, G.F.; Bonucci, E. Tartrate-resistant acid phosphate activity as osteoclastic marker: Sensitivity of cytochemical assessment and serum assay in comparison with standardized osteoclast histomorphometry. Osteoporos. Int. 1997, 7, 39–43. [Google Scholar] [CrossRef] [PubMed]
- Halleen, J.M.; Alatalo, S.L.; Suominen, H.; Cheng, S.; Janckila, A.J.; Väänänen, H.K. Tartrate-resistant acid phosphatase 5b: A novel serum marker of bone resorption. J. Bone Miner. Res. 2000, 15, 1337–1345. [Google Scholar] [CrossRef] [PubMed]
- Miyamoto, T. Regulators of osteoclast differentiation and cell-cell fusion. Keio J. Med. 2011, 60, 101–105. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.P.; Wang, K.C.; Huang, J.D. Effect of estrogen on the activity and growth of human osteoclasts in vitro. Taiwan. J. Obstet. Gynecol. 2009, 48, 350–355. [Google Scholar] [CrossRef]
- Srivastava, S.; Toraldo, G.; Weitzmann, M.N.; Cenci, S.; Ross, F.P.; Pacifici, R. Estrogen decreases osteoclast formation by down-regulating receptor activator of NF-kappa B ligand (RANKL)-induced JNK activation. J. Biol. Chem. 2001, 276, 8836–8840. [Google Scholar] [CrossRef] [PubMed]
- Barber, T.A.; Golledge, S.L.; Castner, D.G.; Healy, K.E. Peptide-modified p (AAm-co-EG/AAc) IPNs grafted to bulk titanium modulate osteoblast behavior in vitro. J. Biomed. Mater. Res. A 2003, 64, 38–47. [Google Scholar] [CrossRef] [PubMed]
- Tan, H.C.; Poh, C.K.; Cai, Y.; Wang, W. Anti-fibrosis effect of BMP-7 peptide functionalization on cobalt chromium alloy. J. Orthop. Res. 2013, 31, 983–990. [Google Scholar] [CrossRef] [PubMed]
- Poh, C.K.; Shi, Z.; Tan, X.W.; Liang, Z.C.; Foo, X.M.; Tan, H.C.; Neoh, K.G.; Wang, W. Cobalt chromium alloy with immobilized BMP peptide for enhanced bone growth. J. Orthop. Res. 2011, 29, 1424–1430. [Google Scholar] [CrossRef] [PubMed]
- Shi, Z.; Neoh, K.G.; Zhong, S.P.; Yung, L.Y.L.; Kang, E.T.; Wang, W. In vitro antibacterial and cytotoxicity assay of multilayered polyelectrolyte-functionalized stainless steel. J. Biomed. Mater. Res. A 2006, 76A, 826–834. [Google Scholar] [CrossRef] [PubMed]
- Yu, F.; Dong, Y.; Yang, Y.W.; Lin, P.T.; Yu, H.H.; Sun, X.; Sun, X.F.; Zhou, H.; Huang, L.; Chen, J.H. Effect of an experimental direct pulp-capping material on the properties and osteogenic differentiation of human dental pulp stem cells. Sci. Rep. 2016, 6, 34713. [Google Scholar] [CrossRef] [PubMed]
- Lipp, R.; Laurent, H.; Gunther, C.; Reidl, J.; Esperling, P.; Tauber, U. Rational design of prodrugs for matrix type transdermal delivery systems: Gestodene esters. Proc. Int. Symp. Control Release Bioact. Mater. 1995, 22, 672–673. [Google Scholar]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Substrates | O% | Ti% | N% | C% |
|---|---|---|---|---|
| Ti | 47.89 | 38.29 | 1.44 | 38.29 |
| Ti-pDOP | 19.20 | 0.70 | 7.27 | 72.84 |
| Ti-pDOP-E2-0.1 µg | 20 | 0.66 | 6.06 | 73.22 |
| Ti-pDOP-E2-1 µg | 18.21 | 0.27 | 5.83 | 75.69 |
| Ti-pDOP-E2-10 µg | 18.86 | 0.42 | 4.65 | 76.07 |
| Substrates | Total Amount of E2 in Loading Solution (μg) | Amount of E2 in the Wash Solution (μg) | Surface Density of Loaded E2 (μg/cm2) |
|---|---|---|---|
| Ti-pDOP-E2-0.1 µg | 0.1 | 0.046 ± 0.1 | 0.054 ± 0.1 |
| Ti-pDOP-E2-1 µg | 1 | 0.28 ± 0.11 | 0.72 ± 0.11 |
| Ti-pDOP-E2-10 µg | 10 | 1.85 ± 0.49 | 8.15 ± 0.49 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Steffi, C.; Shi, Z.; Kong, C.H.; Wang, W. In Vitro Findings of Titanium Functionalized with Estradiol via Polydopamine Adlayer. J. Funct. Biomater. 2017, 8, 45. https://doi.org/10.3390/jfb8040045
Steffi C, Shi Z, Kong CH, Wang W. In Vitro Findings of Titanium Functionalized with Estradiol via Polydopamine Adlayer. Journal of Functional Biomaterials. 2017; 8(4):45. https://doi.org/10.3390/jfb8040045
Chicago/Turabian StyleSteffi, Chris, Zhilong Shi, Chee Hoe Kong, and Wilson Wang. 2017. "In Vitro Findings of Titanium Functionalized with Estradiol via Polydopamine Adlayer" Journal of Functional Biomaterials 8, no. 4: 45. https://doi.org/10.3390/jfb8040045
APA StyleSteffi, C., Shi, Z., Kong, C. H., & Wang, W. (2017). In Vitro Findings of Titanium Functionalized with Estradiol via Polydopamine Adlayer. Journal of Functional Biomaterials, 8(4), 45. https://doi.org/10.3390/jfb8040045