
Chaperonins: Nanocarriers with Biotechnological Applications

 , ,
, ,  , , and
, , and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
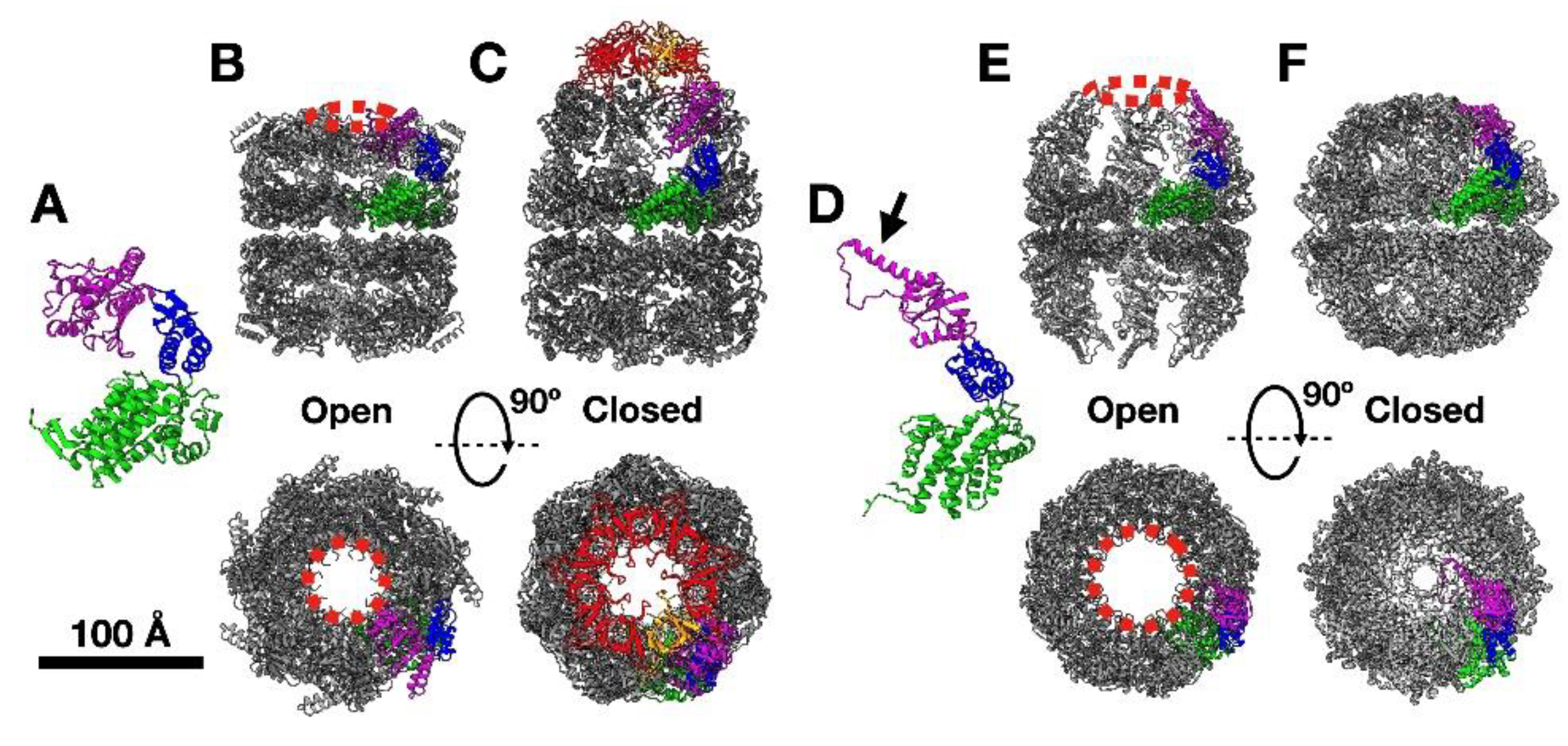
1. Chaperonins: Structure and Function
2. Chaperonins as Nanotechnological Devices
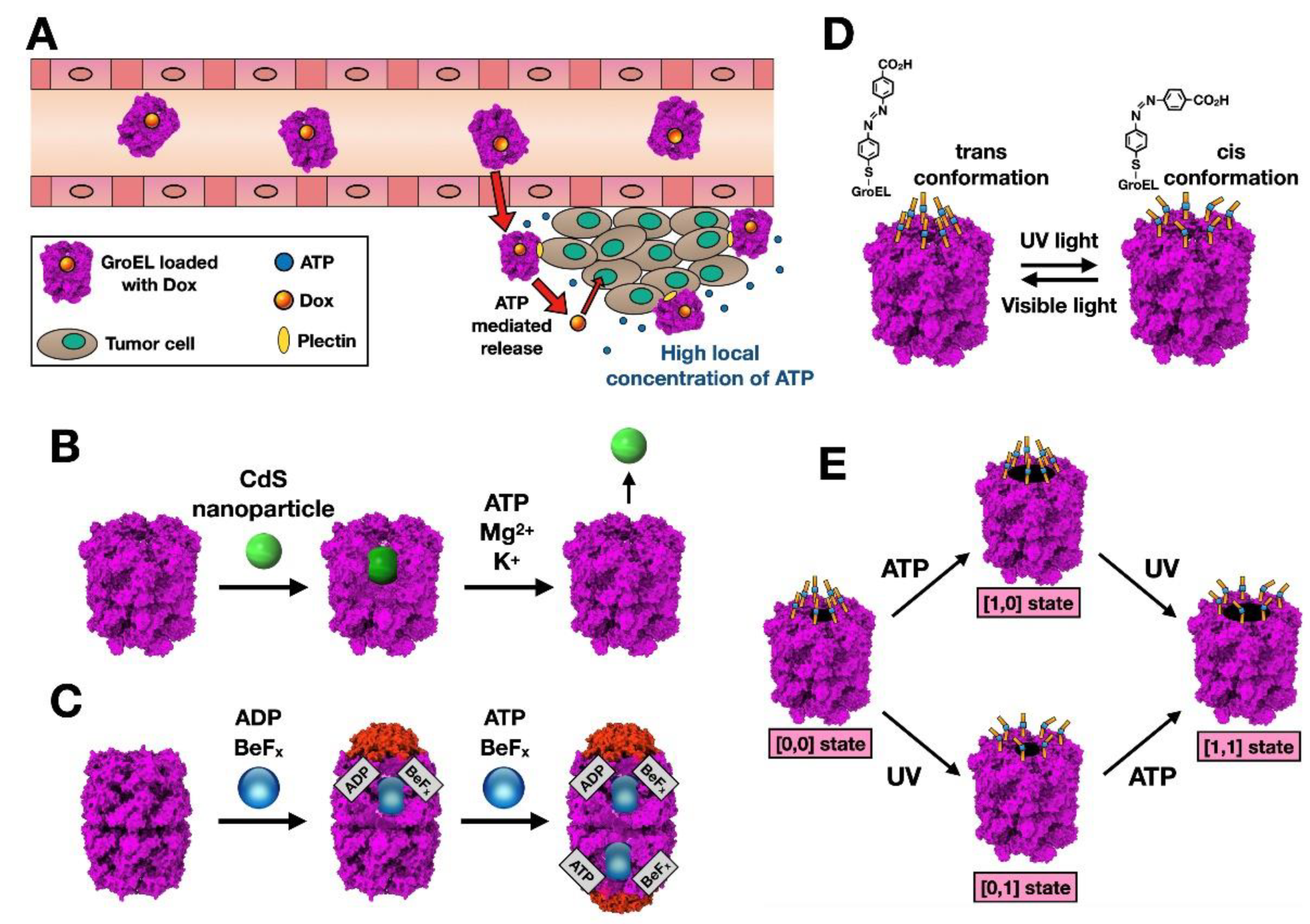
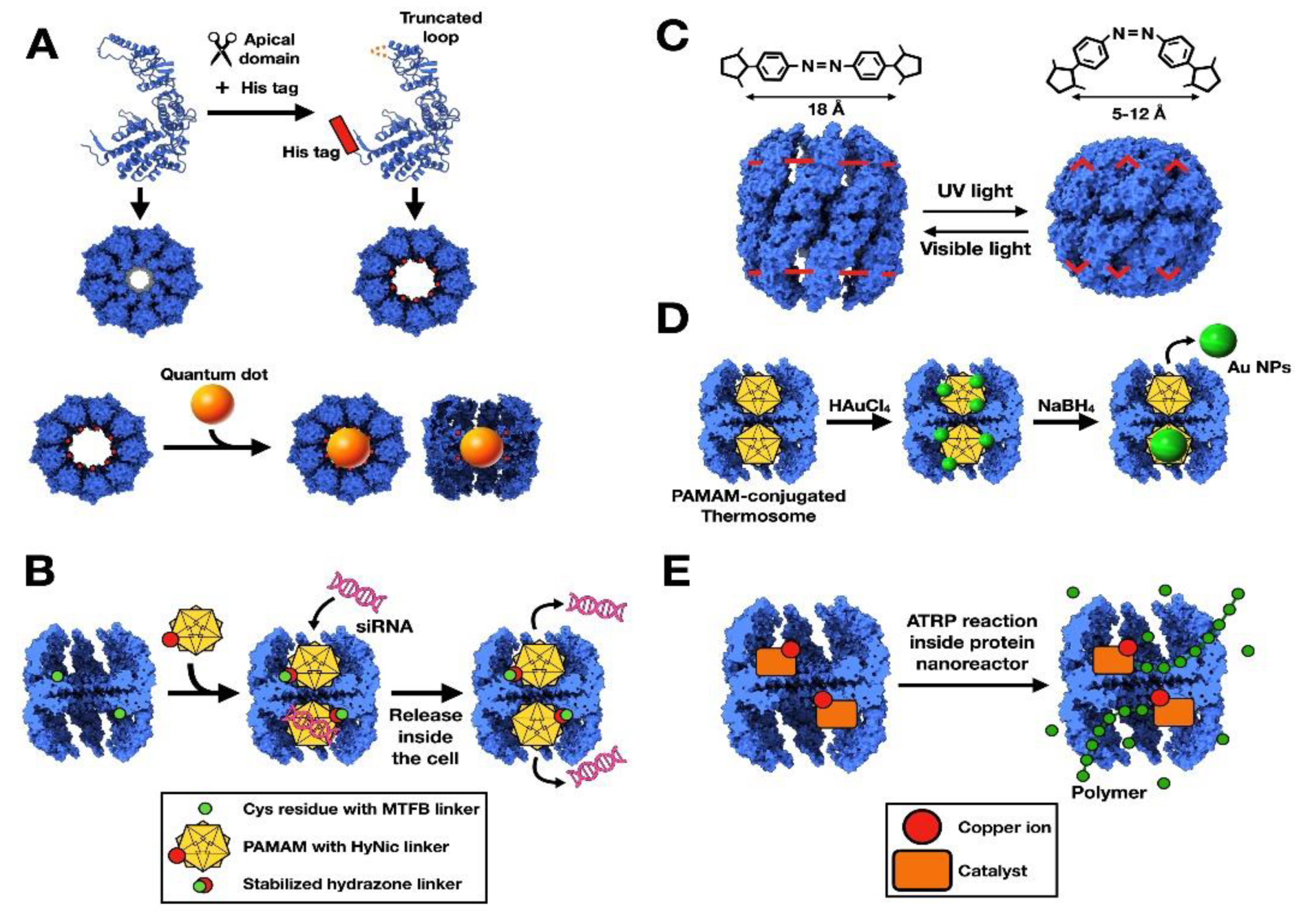
2.1. Chaperonins as Nanocarriers
2.2. Chaperonins as Nanoreactors
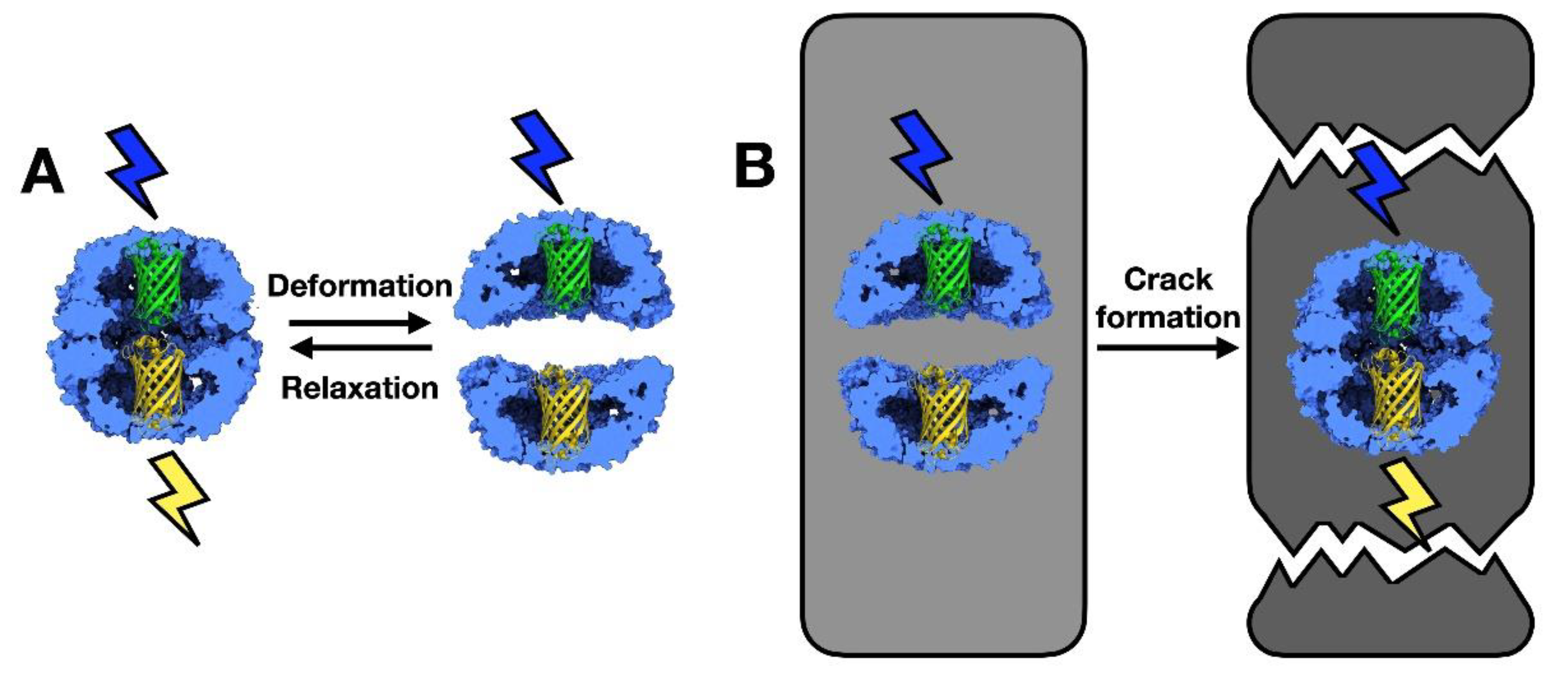
2.3. Chaperonins as Nanosensors
3. Chaperonins as High-Order Nanotechnological Devices
Author Contributions
Funding
Conflicts of Interest
References
- Hartl, F.U.; Bracher, A.; Hayer-Hartl, M. Molecular chaperones in protein folding and proteostasis. Nature 2011, 475, 324–332. [Google Scholar] [CrossRef]
- Bukau, B.; Weissman, J.; Horwich, A. Molecular chaperones and protein quality control. Cell 2006, 125, 443–451. [Google Scholar] [CrossRef] [PubMed]
- Labbadia, J.; Morimoto, R.I. The biology of proteostasis in aging and disease. Annu. Rev. Biochem. 2015, 84, 435–464. [Google Scholar] [CrossRef]
- Kinbara, K.; Aida, T. Toward intelligent molecular machines: Directed motions of biological and artificial molecules and assemblies. Chem. Rev. 2005, 105, 1377–1400. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Fernández, M.R.; Sot, B.; Valpuesta, J.M. Molecular chaperones: Functional mechanisms and nanotechnological applications. Nanotechnology 2016, 27, 324004. [Google Scholar] [CrossRef] [PubMed]
- Yebenes, H.; Mesa, P.; Munoz, I.G.; Montoya, G.; Valpuesta, J.M. Chaperonins: Two rings for folding. Trends Biochem. Sci. 2011, 36, 424–432. [Google Scholar] [CrossRef]
- Skjærven, L.; Cuellar, J.; Martinez, A.; Valpuesta, J.M. Dynamics, flexibility, and allostery in molecular chaperonins. FEBS Lett. 2015, 589, 2522–2532. [Google Scholar] [CrossRef] [PubMed]
- Braig, K.; Otwinowski, Z.; Hegde, R.; Boisvert, D.C.; Joachimiak, A.; Horwich, A.L.; Sigler, P.B. The crystal structure of the bacterial chaperonin GroEL at 2.8 A. Nature 1994, 371, 578–586. [Google Scholar] [CrossRef]
- Huo, Y.; Hu, Z.; Zhang, K.; Wang, L.; Zhai, Y.; Zhou, Q.; Lander, G.; Zhu, Y.; He, Y.; Pang, X.; et al. Crystal structure of group II chaperonin in the open state. Structure 2010, 18, 1270–1279. [Google Scholar] [CrossRef]
- Pereira, J.H.; Ralston, C.Y.; Douglas, N.R.; Meyer, D.; Knee, K.M.; Goulet, D.R.; King, J.A.; Frydman, J.; Adams, P.D. Crystal structures of a group II chaperonin reveal the open and closed states associated with the protein folding cycle. J. Biol. Chem. 2010, 285, 27958–27966. [Google Scholar] [CrossRef]
- Muñoz, I.G.; Yébenes, H.; Zhou, M.; Mesa, P.; Serna, M.; Park, A.Y.; Bragado-Nilsson, E.; Beloso, A.; de Cárcer, G.; Malumbres, M.; et al. Crystal structure of the open conformation of the mammalian chaperonin CCT in complex with tubulin. Nat. Struct. Mol. Biol. 2011, 18, 14–19. [Google Scholar] [CrossRef]
- Gruber, R.; Horovitz, A. Allosteric Mechanisms in Chaperonin Machines. Chem. Rev. 2016, 116, 6588–6606. [Google Scholar] [CrossRef]
- Xu, Z.; Horwich, A.L.; Sigler, P.B. The crystal structure of the asymmetric GroEL-GroES-(ADP)7 chaperonin complex. Nature 1997, 388, 741–750. [Google Scholar] [CrossRef] [PubMed]
- Ditzel, L.; Löwe, J.; Stock, D.; Stetter, K.O.; Huber, H.; Huber, R.; Steinbacher, S. Crystal structure of the thermosome, the archaeal chaperonin and homolog of CCT. Cell 1998, 93, 125–138. [Google Scholar] [CrossRef]
- Yuan, Y.; Du, C.; Zhu, J.; Wu, S.; Zhang, Y.; Ji, T.; Lei, J.; Yang, Y.; Gao, N.; Nie, G. Chaperonin-GroEL as a smart hydrophobic drug delivery and tumor targeting molecular machine for tumor therapy. Nano Lett. 2018, 18, 921–928. [Google Scholar] [CrossRef]
- Ishii, D.; Kinbara, K.; Ishida, Y.; Ishii, N.; Okochi, M.; Yohda, M.; Aida, T. Chaperonin-mediated stabilization and ATP-triggered release of semiconductor nanoparticles. Nature 2003, 423, 628–632. [Google Scholar] [CrossRef] [PubMed]
- Yoda, H.; Koike-Takeshita, A. TEM and STEM-EDS evaluation of metal nanoparticle encapsulation in GroEL/GroES complexes according to the reaction mechanism of chaperonin. Microscopy 2020, dfaa064. [Google Scholar] [CrossRef] [PubMed]
- Taguchi, H.; Tsukuda, K.; Motojima, F.; Koike-Takeshita, A.; Yoshida, M. BeFx Stops the Chaperonin Cycle of GroEL-GroES and Generates a Complex with Double Folding Chambers. J. Biol. Chem. 2004, 279, 45737–45743. [Google Scholar] [CrossRef]
- Muramatsu, S.; Kinbara, K.; Taguchi, H.; Ishii, N.; Aida, T. Semibiological molecular machine with an implemented ‘AND’ logic gate for regulation of protein folding. J. Am. Chem. Soc. 2006, 128, 3764–3769. [Google Scholar] [CrossRef]
- Nussbaumer, M.G.; Duskey, J.T.; Rother, M.; Renggli, K.; Chami, M.; Bruns, N. Chaperonin–Dendrimer Conjugates for siRNA Delivery. Adv. Sci. 2016, 3, 1600046. [Google Scholar] [CrossRef]
- Kagawa, H.K.; Osipiuk, J.; Maltsev, N.; Overbeek, R.; Quaite-Randall, E.; Joachimiak, A.; Trent, J.D. The 60 kDa heat shock proteins in the hyperthermophilic archaeon Sulfolobus shibatae. J. Mol. Biol. 1995, 253, 712–725. [Google Scholar] [CrossRef]
- Xie, H.; Li, Y.F.; Kagawa, H.K.; Trent, J.D.; Mudalige, K.; Cotlet, M.; Swanson, B.I. An Intrinsically fluorescent recognition ligand scaffold based on chaperonin protein and semiconductor quantum-dot conjugates. Small 2009, 5, 1036–1042. [Google Scholar] [CrossRef]
- Hoersch, D.; Roh, S.H.; Chiu, W.; Kortemme, T. Reprogramming an ATP-driven protein machine into a light-gated nanocage. Nat. Nanotechnol. 2013, 8, 928–932. [Google Scholar] [CrossRef] [PubMed]
- Nussbaumer, M.G.; Bisig, C.; Bruns, N. Using the dendritic polymer PAMAM to form gold nanoparticles in the protein cage thermosome. Chem. Commun. 2016, 52, 10537–10539. [Google Scholar] [CrossRef]
- Renggli, K.; Nussbaumer, M.G.; Urbani, R.; Pfohl, T.; Bruns, N. A chaperonin as protein nanoreactor for atom-transfer radical polymerization. Angew. Chem. Int. Ed. Engl. 2014, 53, 1443–1447. [Google Scholar] [CrossRef]
- Fawell, S.; Seery, J.; Daikh, Y.; Moore, C.; Chen, L.L.; Pepinsky, B.; Barsoum, J. Tat-mediated delivery of heterologous proteins into cells. Proc. Natl. Acad. Sci. USA 1994, 91, 664–668. [Google Scholar] [CrossRef] [PubMed]
- Jin, M.; Liu, C.; Han, W.; Cong, Y. TRiC/CCT Chaperonin: Structure and Function. Subcell Biochem. 2019, 93, 625–654. [Google Scholar] [PubMed]
- Sergeeva, O.A.; Chen, B.; Haase-Pettingell, C.; Ludtke, S.J.; Chiu, W.; King, J.A. Human CCT4 and CCT5 chaperonin subunits expressed in Escherichia coli form biologically active homo-oligomers. J. Biol. Chem. 2013, 288, 17734–17744. [Google Scholar] [CrossRef] [PubMed]
- Cotanda, P.; O’Reilly, R.K. Molecular recognition driven catalysis using polymeric nanoreactors. Chem. Commun. 2012, 48, 10280–10282. [Google Scholar] [CrossRef]
- Tanner, P.; Balasubramanian, V.; Palivan, C.G. Aiding nature’s organelles: Artificial peroxisomes play their role. Nano Lett. 2013, 13, 2875–2883. [Google Scholar] [CrossRef]
- Bruns, N.; Pustelny, K.; Bergeron, L.M.; Whitehead, T.A.; Clark, D.S. Mechanical nanosensor based on FRET within a thermosome: Damage-reporting polymeric materials. Angew. Chem. Int. Ed. Engl. 2009, 48, 5666–5669. [Google Scholar] [CrossRef] [PubMed]
- Rizzo, M.A.; Springer, G.; Segawa, K.; Zipfel, W.R.; Piston, D.W. Optimization of pairings and detection conditions for measurement of FRET between cyan and yellow fluorescent proteins. Microsc. Microanal. 2006, 12, 238–254. [Google Scholar] [CrossRef] [PubMed]
- Trent, J.D.; Kagawa, H.K.; Yaoi, T.; Olle, E.; Zaluzec, N.J. Chaperonin filaments: The archaeal cytoskeleton? Proc. Natl. Acad. Sci. USA 1997, 94, 5383–5388. [Google Scholar] [CrossRef] [PubMed]
- Furutani, M.; Iida, T.; Yoshida, T.; Maruyama, T. Group II chaperonin in a thermophilic methanogen, Methanococcus thermolithotrophicus. Chaperone activity and filament-forming ability. J. Biol. Chem. 1998, 273, 28399–28407. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Yagi, H.; Furutani, Y.; Nakamura, T.; Inaguma, A.; Guo, H.; Kong, Y.; Goto, Y. Self-assembly of the chaperonin GroEL nanocage induced at submicellar detergent. Sci. Rep. 2014, 4, 5614. [Google Scholar] [CrossRef]
- Biswas, S.; Kinbara, K.; Oya, N.; Ishii, N.; Taguchi, H.; Aida, T. A tubular biocontainer: Metal ion-induced 1D assembly of a molecularly engineered chaperonin. J. Am. Chem. Soc. 2009, 131, 7556–7557. [Google Scholar] [CrossRef]
- Sim, S.; Miyajima, D.; Niwa, T.; Taguchi, H.; Aida, T. Tailoring micrometer-long high-integrity 1D array of superparamagnetic nanoparticles in a nanotubular protein jacket and its lateral magnetic assembling behavior. J. Am. Chem. Soc. 2015, 137, 4658–4661. [Google Scholar] [CrossRef]
- Biswas, S.; Kinbara, K.; Niwa, T.; Taguchi, H.; Ishii, N.; Watanabe, S.; Miyata, K.; Kataoka, K.; Aida, T. Biomolecular robotics for chemomechanically driven guest delivery fuelled by intracellular ATP. Nat. Chem. 2013, 5, 613–620. [Google Scholar] [CrossRef]
- Sendai, T.; Biswas, B.; Aida, T. Photoreconfigurable Supramolecular Nanotube. J. Am. Chem. Soc. 2013, 135, 11509–11512. [Google Scholar] [CrossRef]
- Sim, S.; Niwa, T.; Taguchi, H.; Aida, T. Supramolecular Nanotube of Chaperonin GroEL: Length Control for Cellular Uptake Using Single-Ring GroEL Mutant as End-Capper. J. Am. Chem. Soc. 2016, 138, 11152–11155. [Google Scholar] [CrossRef]
- Weissman, J.S.; Hohl, C.M.; Kovalenko, O.; Kashi, Y.; Chen, S.; Braig, K.; Saibil, H.R.; Fenton, W.A.; Horwich, A.L. Mechanism of GroEL action: Productive release of polypeptide from a sequestered position under GroES. Cell 1995, 83, 577–587. [Google Scholar] [CrossRef]
- Keren, K.; Krueger, M.; Gilad, R.; Ben-Yoseph, G.; Sivan, U.; Braun, E. 2002 Sequence-specific molecular lithography on single DNA molecules. Science 2002, 297, 72–75. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.W.; Mao, C.; Flynn, C.E.; Belcher, A.M. Ordering of quantum dots using genetically engineered viruses. Science 2002, 296, 892–895. [Google Scholar] [CrossRef] [PubMed]
- Ellis, M.J.; Knapp, S.; Koeck, P.J.; Fakoor-Biniaz, Z.; Ladenstein, R.; Hebert, H. Two-dimensional crystallization of the chaperonin TF55 from the hyperthermophilic archaeon Sulfolobus solfataricus. J. Struct. Biol. 1998, 123, 30–36. [Google Scholar] [CrossRef] [PubMed]
- Koeck, P.J.; Kagawa, H.K.; Ellis, M.J.; Hebert, H.; Trent, J.D. Two-dimensional crystals of reconstituted beta-subunits of the chaperonin TF55 from Sulfolobus shibatae. Biochim. Biophys. Acta 1998, 1429, 40–44. [Google Scholar] [CrossRef]
- Yaoi, T.; Kagawa, H.K.; Trent, J.D. Chaperonin filaments: Their formation and an evaluation of methods for studying them. Arch. Biochem. Biophys. 1998, 356, 55–62. [Google Scholar] [CrossRef]
- McMillan, R.A.; Paavola, C.D.; Howard, J.; Chan, S.L.; Zaluzec, N.J.; Trent, J.D. Ordered nanoparticle arrays formed on engineered chaperonin protein templates. Nat. Mater. 2002, 1, 247–252. [Google Scholar] [CrossRef]
- McMillan, R.A.; Howard, J.; Zaluzec, N.J.; Kagawa, H.K.; Mogul, R.; Li, Y.F.; Paavola, C.D.; Trent, J.D. A self-assembling protein template for constrained synthesis and patterning of nanoparticle arrays. J. Am. Chem. Soc. 2005, 127, 2800–2801. [Google Scholar] [CrossRef]
- Li, Y.; Paavola, C.D.; Kawaga, H.; Chan, S.L.; Trent, J.D. Mutant chaperonin proteins: New tools for nanotechnology. Nanotechnology 2007, 18, 455101. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pipaón, S.; Gragera, M.; Bueno-Carrasco, M.T.; García-Bernalt Diego, J.; Cantero, M.; Cuéllar, J.; Fernández-Fernández, M.R.; Valpuesta, J.M. Chaperonins: Nanocarriers with Biotechnological Applications. Nanomaterials 2021, 11, 503. https://doi.org/10.3390/nano11020503
Pipaón S, Gragera M, Bueno-Carrasco MT, García-Bernalt Diego J, Cantero M, Cuéllar J, Fernández-Fernández MR, Valpuesta JM. Chaperonins: Nanocarriers with Biotechnological Applications. Nanomaterials. 2021; 11(2):503. https://doi.org/10.3390/nano11020503
Chicago/Turabian StylePipaón, Sergio, Marcos Gragera, M. Teresa Bueno-Carrasco, Juan García-Bernalt Diego, Miguel Cantero, Jorge Cuéllar, María Rosario Fernández-Fernández, and José María Valpuesta. 2021. "Chaperonins: Nanocarriers with Biotechnological Applications" Nanomaterials 11, no. 2: 503. https://doi.org/10.3390/nano11020503
APA StylePipaón, S., Gragera, M., Bueno-Carrasco, M. T., García-Bernalt Diego, J., Cantero, M., Cuéllar, J., Fernández-Fernández, M. R., & Valpuesta, J. M. (2021). Chaperonins: Nanocarriers with Biotechnological Applications. Nanomaterials, 11(2), 503. https://doi.org/10.3390/nano11020503