
Fluorescent Nanoparticle-RNAi-Mediated Silencing of Sterol Carrier Protein-2 Gene Expression Suppresses the Growth, Development, and Reproduction of Helicoverpa armigera


 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Insects and Cells
2.2. Detection of FNPs and siRNA in H. armigera Cells and Larval Midgut
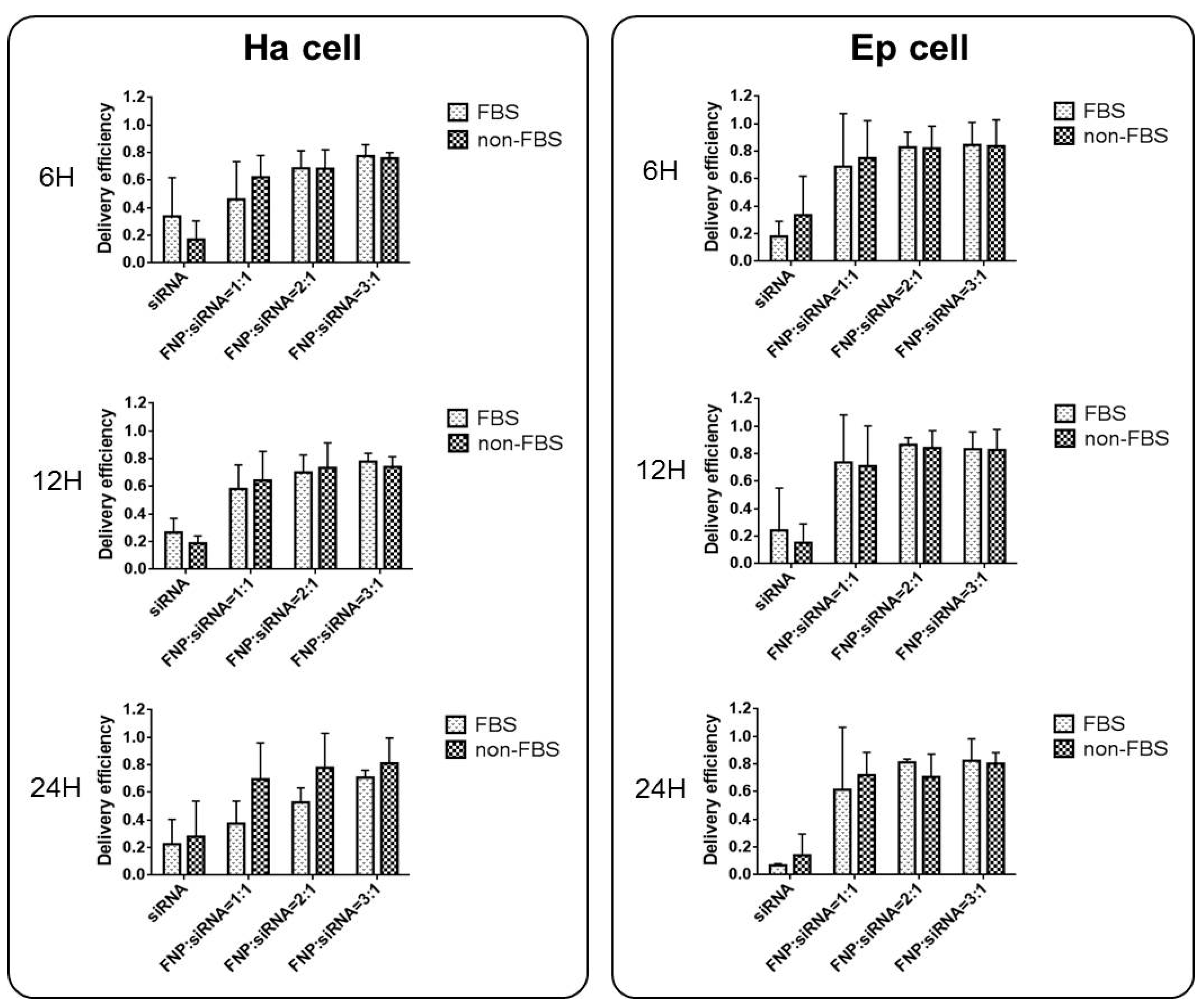
2.3. Analysis of the Effect of Serum Presence on Cell Transfection Efficiency of FNP-siRNA
2.4. HaSCP-2 Expression Silencing in Ha Cells by FNP-siRNA
2.5. FNP-HaSCP-2 siRNA Interference of the HaSCP-2 Gene by Oral Feeding in H. armigera
2.6. qRT-PCR Analysis
2.7. Cholesterol Level Assay
2.8. Biological Assays in H. armigera
2.9. Statistical Analyses
3. Results
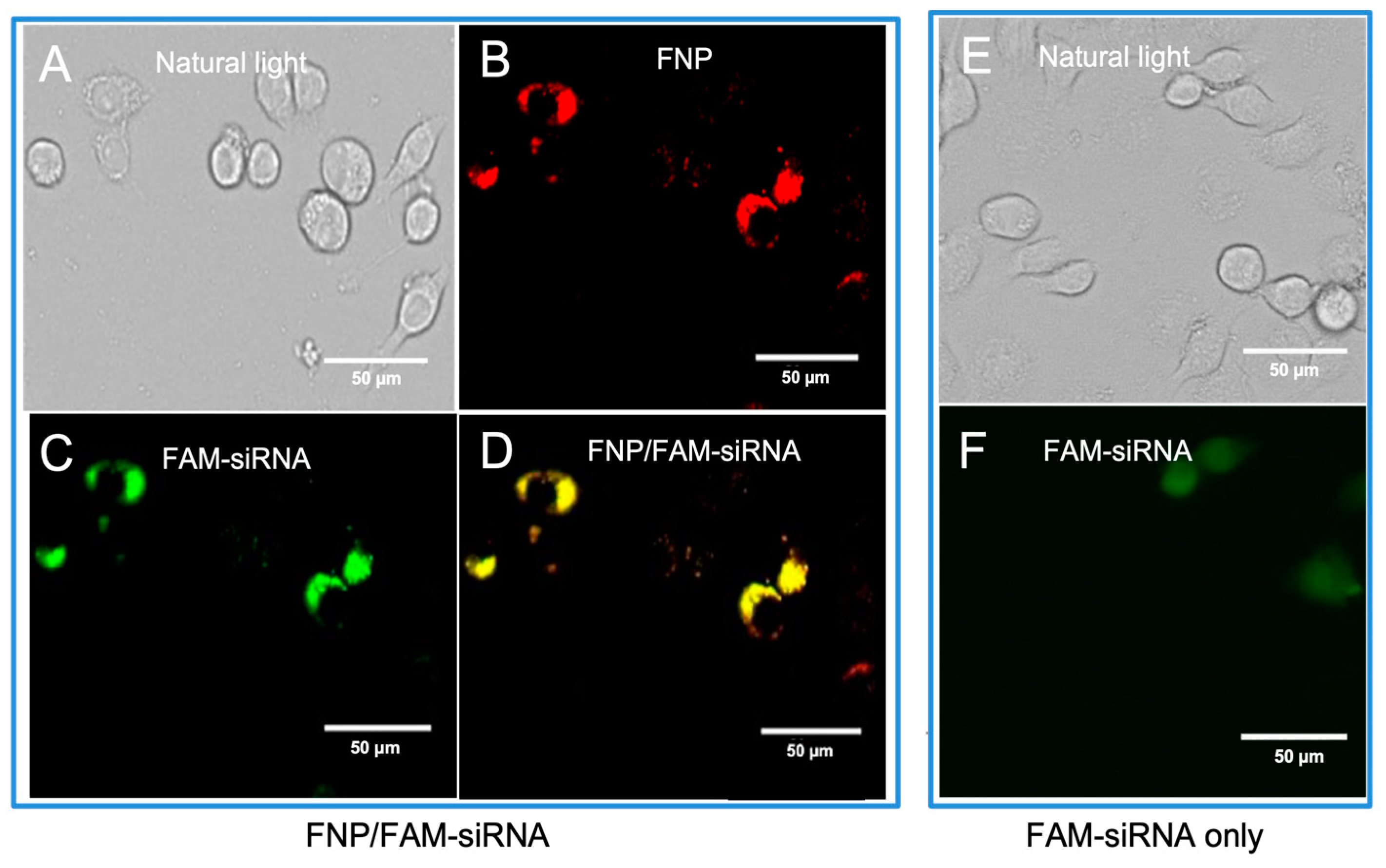
3.1. Fluorescence Images Visualizing the Distribution of FNP and siRNA in Ha Cells of H. armigera
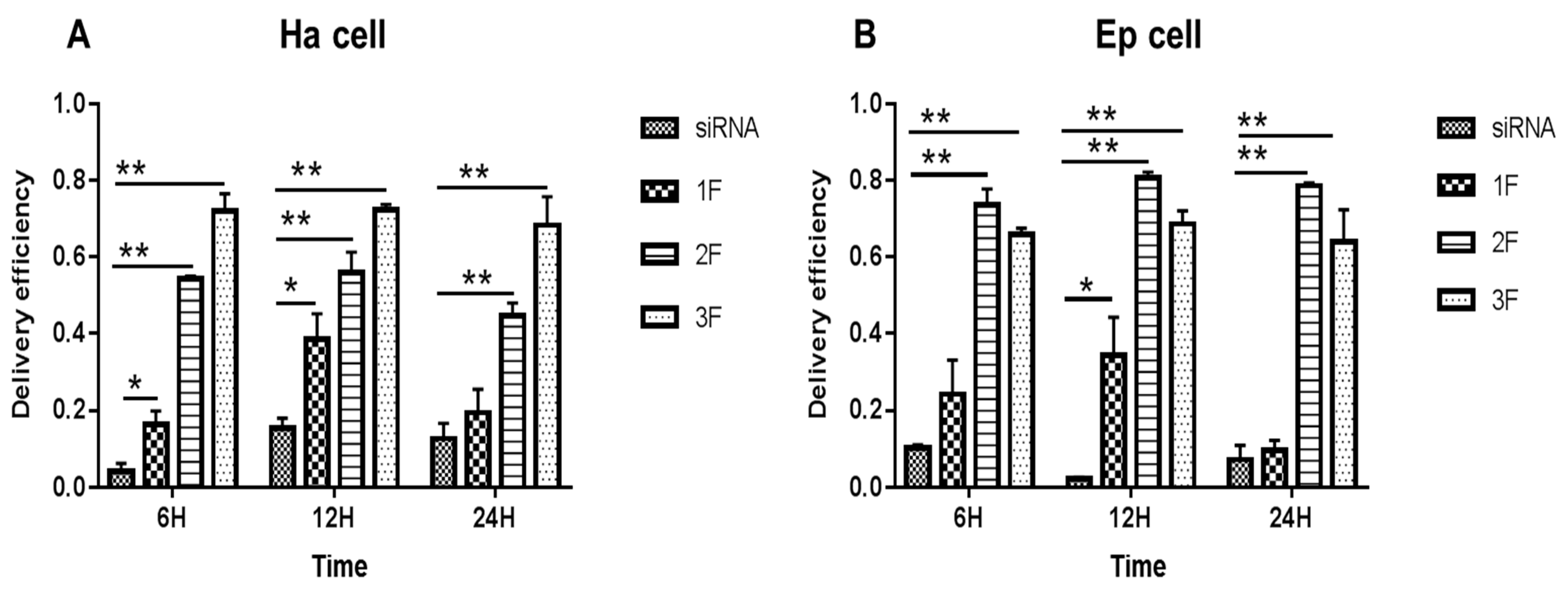
3.2. siRNA Delivery Efficiency of FNP In Vitro
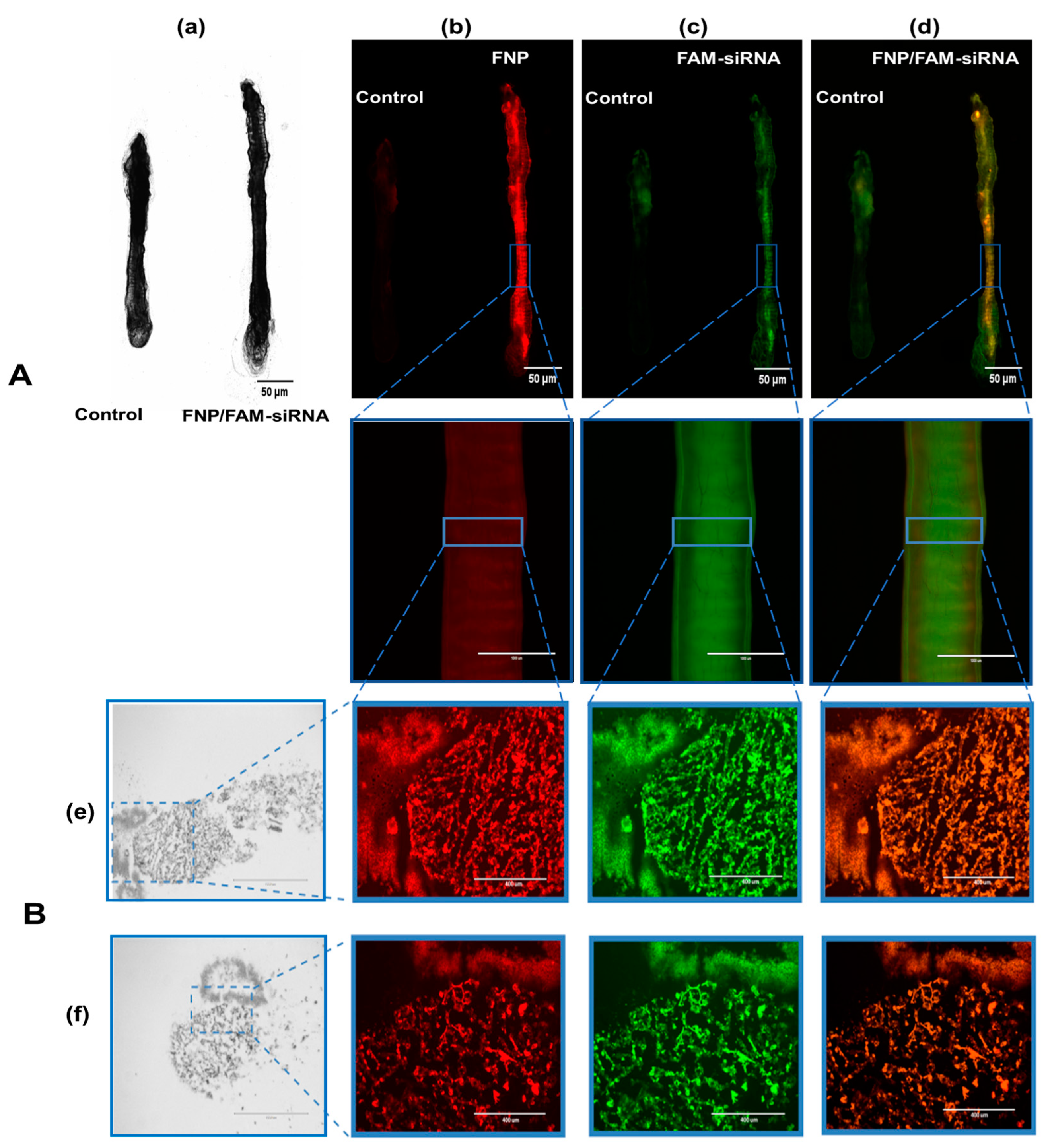
3.3. Fluorescence Images Visualizing the Distribution of FNP and siRNA in the Midgut of H. armigera
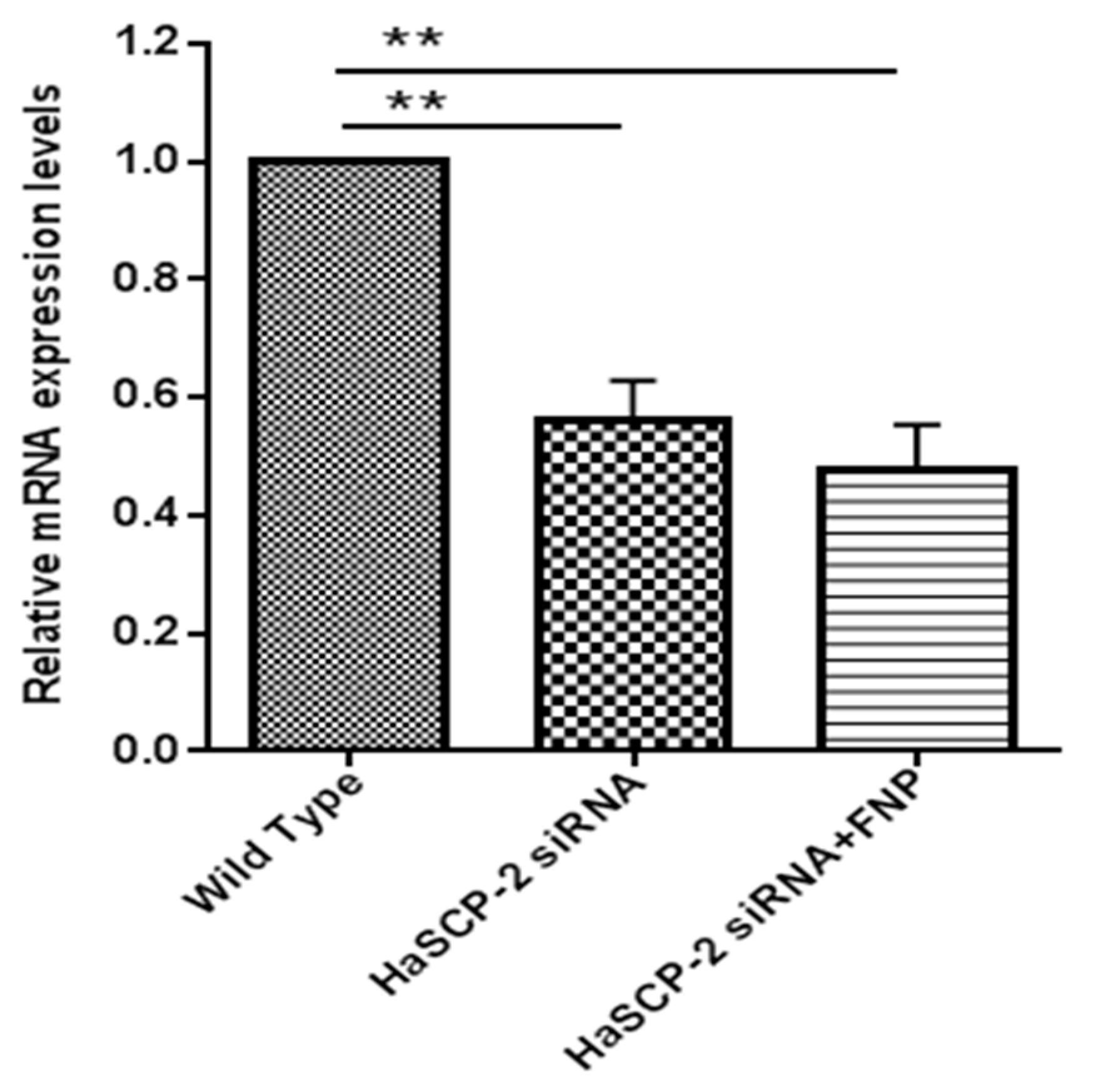
3.4. In Vitro HaSCP-2 Knockdown by FNP-siRNA in Ha Cells
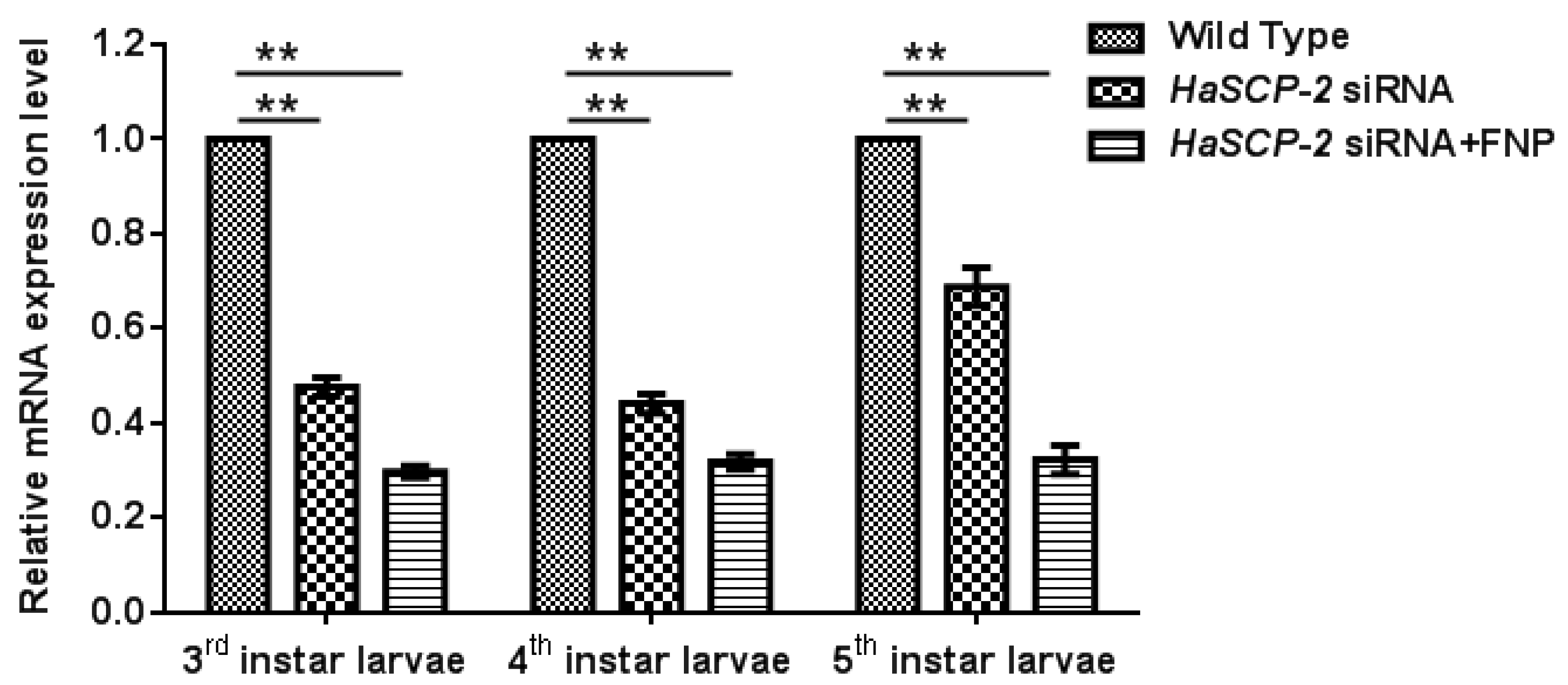
3.5. In Vivo HaSCP-2 Knockdown by FNP-siRNA in H. armigera
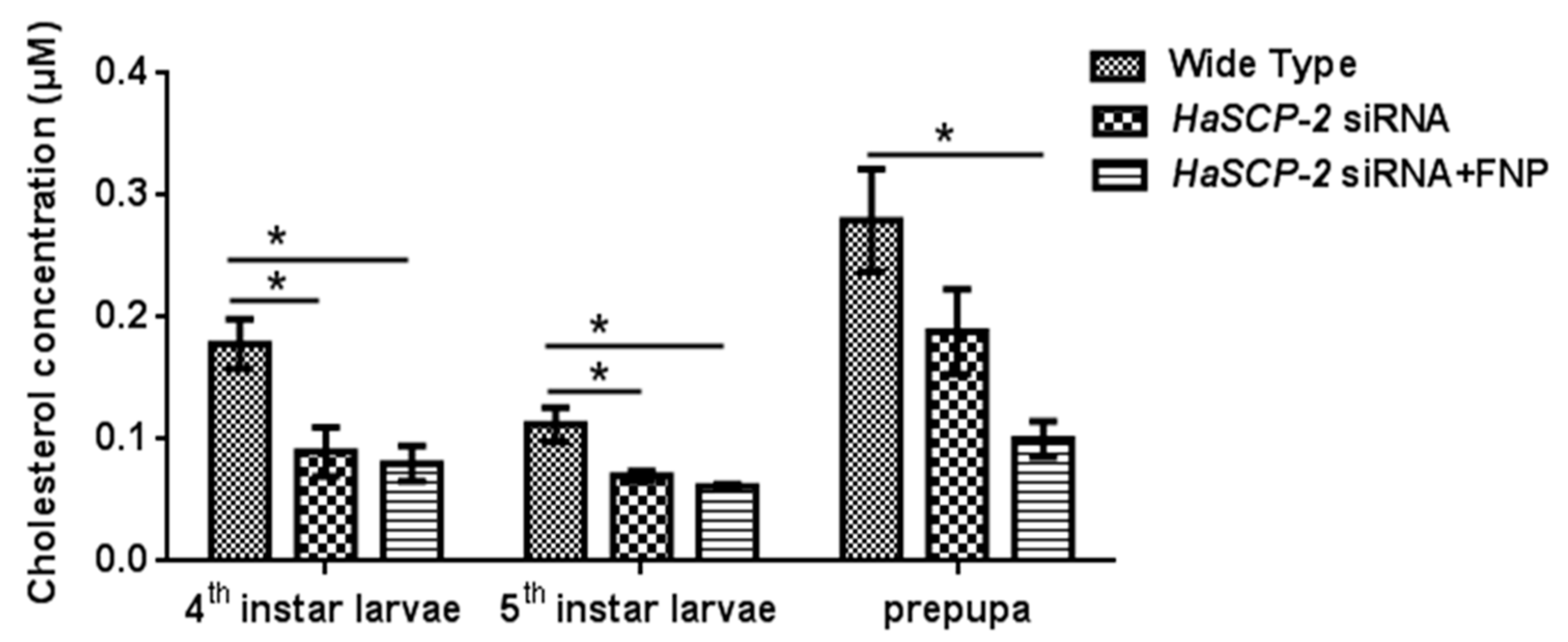
3.6. Effect of HaSCP-2 Knockdown on Cholesterol Accumulation by FNP-RNAi
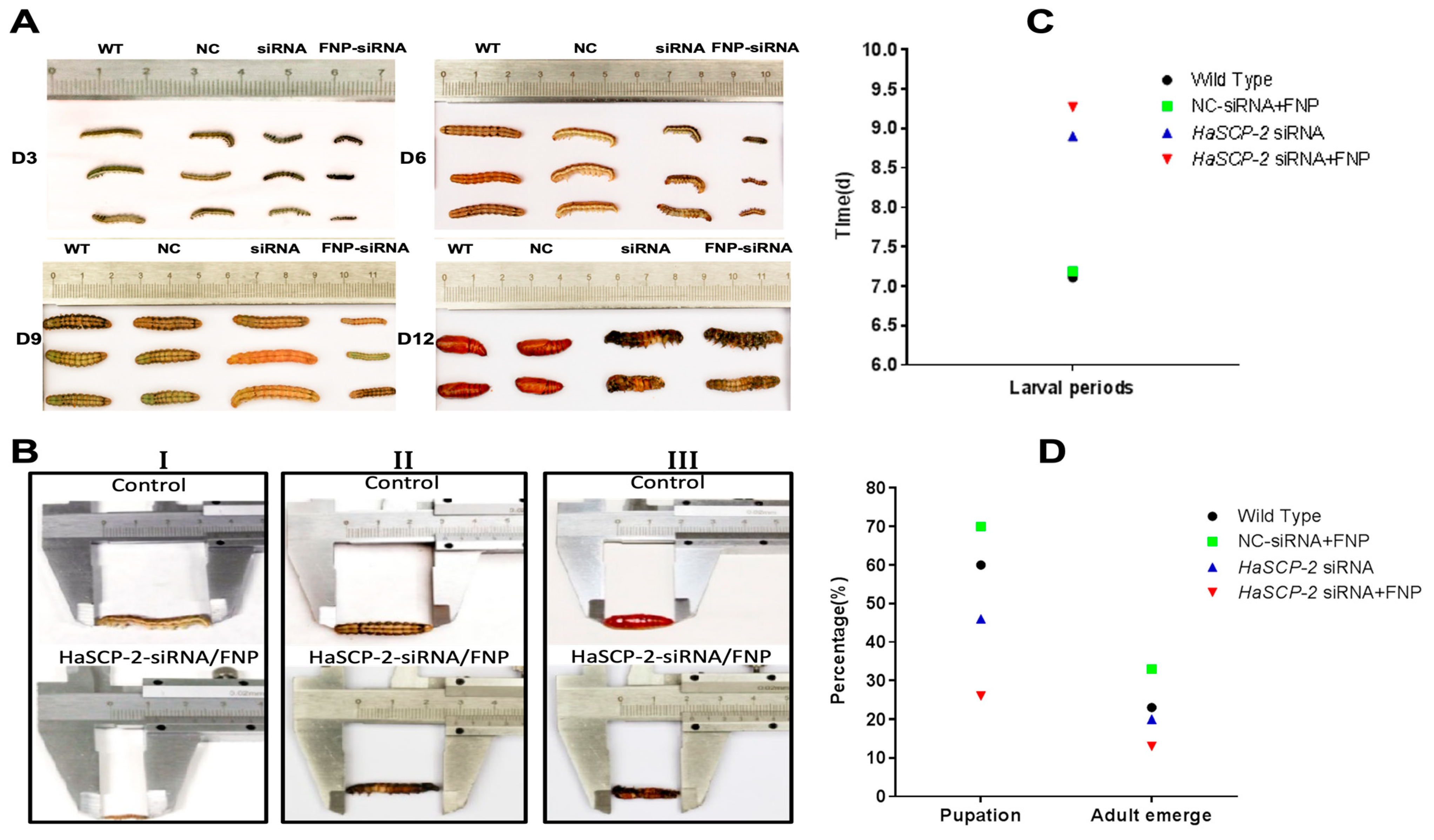
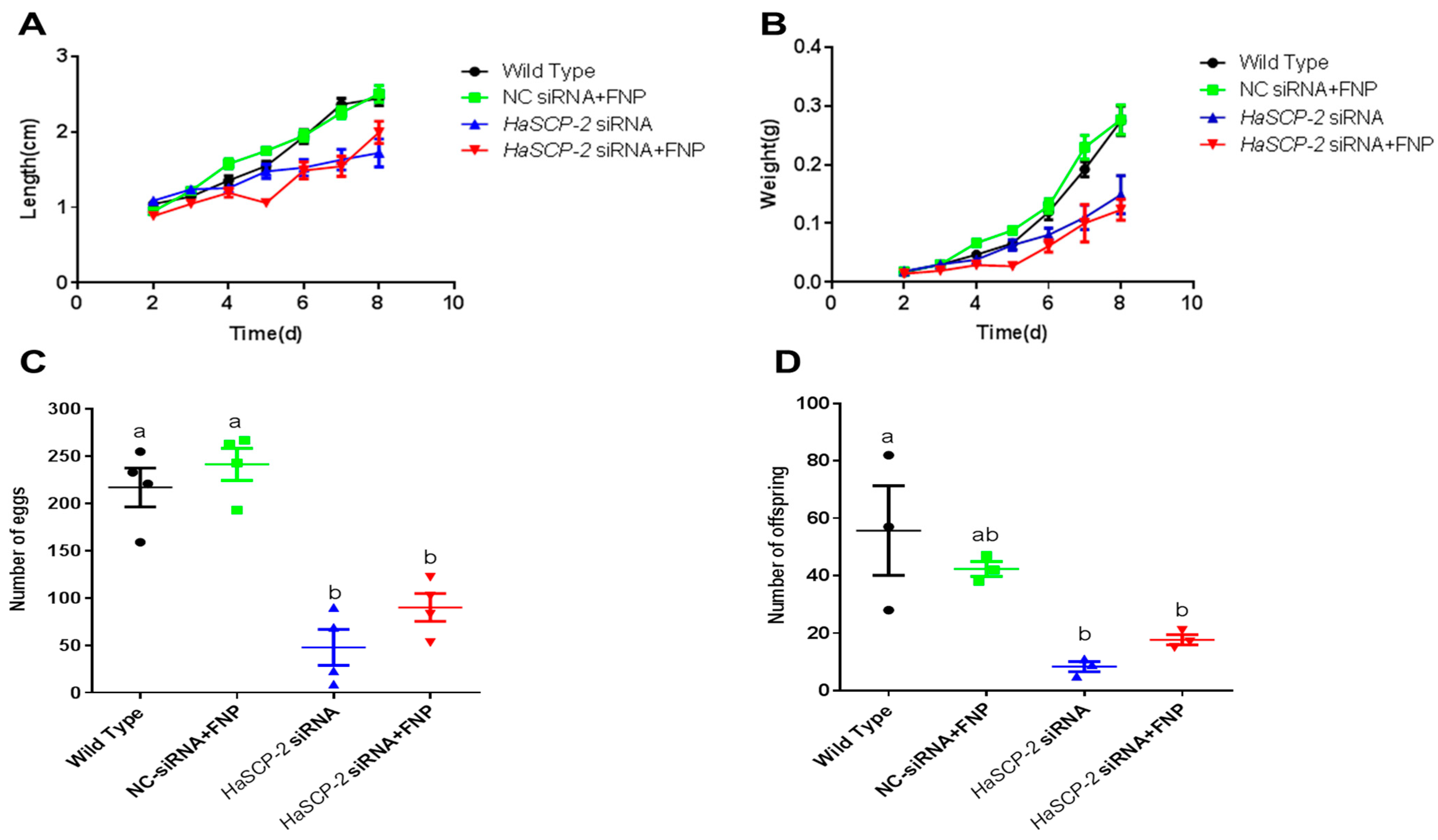
3.7. Effects of HaSCP-2 Silencing on the Growth, Development, and Reproduction of H. armigera by FNP-RNAi
4. Discussion
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cunningham, J.; Zalucki, M.; West, S. Learning in Helicoverpa armigera (Lepidoptera: Noctuidae): A new look at the behaviour and control of a polyphagous pest. Bull. Entomol. Res. 1999, 89, 201–207. [Google Scholar] [CrossRef] [Green Version]
- Brcbvault, T.; Oumarou, Y.; Achaleke, J.; Vaissayre, M.; Nibouche, S. Initial activity and persistence of insecticides for the control of bollworms (Lepidoptera: Noctuidae) in cotton crops. Crop Prot. 2009, 28, 401–406. [Google Scholar] [CrossRef]
- Joseph, A.; Thierry, B. Inheritance and stability of pyrethroid resistance in the cotton bollworm Helicoverpa armigera (Lepidoptera: Noctuidae) in Central Africa. Pest Manag Sci. 2010, 66, 137–141. [Google Scholar]
- Gassmann, A.; Carrière, Y.; Tabashnik, B. Fitness Costs of Insect Resistance to Bacillus thuringiensis. Annu. Rev. Entomol. 2009, 54, 147–163. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Wu, K.; Gould, F.; Wang, J.; Miao, J.; Gao, X.; Guo, Y. Increasing tolerance to Cry1Ac cotton from cotton bollworm, Helicoverpa armigera, was confirmed in Bt cotton farming area of China. Ecol. Entomol. 2007, 32, 366–375. [Google Scholar] [CrossRef]
- Jin, L.; Zhang, H.; Lu, Y.; Yang, Y.; Wu, K.; Tabashnik, B.; Wu, Y. Large-scale test of the natural refuge strategy for delaying insect resistance to transgenic Bt crops. Nat. Biotechnol. 2015, 33, 169–174. [Google Scholar] [CrossRef]
- Jin, L.; Wang, J.; Guan, F.; Zhang, J.; Yu, S.; Liu, S.; Xue, Y.; Li, L.; Wu, S.; Wang, X.; et al. Dominant point mutation in a tetraspanin gene associated with field-evolved resistance of cotton bollworm to transgenic Bt cotton. Proc. Natl. Acad. Sci. USA 2018, 115, 8402–8407. [Google Scholar] [CrossRef] [Green Version]
- Goldstein, J.; Brown, M. Regulation of the mevalonate pathway. Nature 1990, 343, 425–430. [Google Scholar] [CrossRef]
- Lasser, N.; Clayton, R. The intracellular distribution of sterols in Eurycotis floridana and its possible relation to subcellular membrane structures. J. Lipid Res. 1966, 7, 413–421. [Google Scholar] [CrossRef]
- Borovsky, D.; Whisenton, L.; Thomas, B.; Morton, S. Biosynthesis and distribution of ecdysone and 20-OH-ecdysone in Aedes aegypti. Arch Insect Biochem. 1986, 3, 19–30. [Google Scholar] [CrossRef]
- Jouni, Z.; Zamora, J.; Wells, M. Absorption and tissue distribution of cholesterol in Manduca sexta. Arch Insect Biochem. 2002, 49, 167–175. [Google Scholar] [CrossRef]
- Nes, W.; Lopez, M.; Zhou, W.; Guo, D.; Dowd, P.; Norton, R. Sterol utilization and metabolism by Heliothis zea. Lipids 1997, 32, 1317–1323. [Google Scholar] [CrossRef]
- Jing, X.; Behmer, S. Insect sterol nutrition: Physiological mechanisms, ecology, and applications. Annu. Rev. Entomol. 2020, 65, 251–271. [Google Scholar] [CrossRef]
- Vyazunova, I.; Lan, Q.; Liu, N. Insect sterol carrier protein-2 gene family: Structures and functions. Recent Adv. Insect Physiol. Toxicol. Mol. Biol. 2008, 37, 173–198. [Google Scholar]
- Lan, Q.; Massey, R. Subcellular localization of the mosquito sterol carrier protein-2 and sterol carrier protein-x. J. Lipid Res. 2004, 45, 1468–1474. [Google Scholar] [CrossRef] [Green Version]
- Kim, M.; Wessely, V.; Lan, Q. Identification of mosquito sterol carrier protein-2 inhibitors. J. Lipid Res. 2005, 46, 650–657. [Google Scholar] [CrossRef] [Green Version]
- Krebs, K.; Lan, Q. Isolation and expression of a sterol carrier protein-2 gene from the yellow fever mosquito, Aedes aegypti. Insect Mol. Biol. 2003, 12, 51–60. [Google Scholar] [CrossRef]
- Radek, J.; Dyer, D.; Lan, Q. Effects of mutations in Aedes aegypti sterol carrier protein-2 on the biological function of the protein. Biochemistry. 2010, 49, 7532–7541. [Google Scholar] [CrossRef] [Green Version]
- Peng, R.; Fu, Q.; Hong, H.; Tyler, S.; Lan, Q. THAP and ATF-2 regulated sterol carrier protein-2 promoter activities in the larval midgut of the yellow fever mosquito, Aedes aegypti. PLoS ONE 2017, 7, e46948. [Google Scholar] [CrossRef] [Green Version]
- Gong, J.; Hong, Y.; Zha, X.; Lu, C.; Zhu, Y.; Xia, Q. Molecular cloning and characterization of Bombyx mori sterol carrier protein x/sterol carrier protein 2 (SCPx/SCP2) gene. Mitochondrial DNA 2006, 17, 326–333. [Google Scholar]
- Guo, X.; Zheng, S.; Liu, L.; Feng, Q. The sterol carrier protein 2/3-oxoacyl-CoA thiolase (SCPx) is involved in cholesterol uptake in the midgut of Spodoptera litura: Gene cloning, expression, localization and functional analyses. BMC Mol. Biol. 2009, 10, 102. [Google Scholar] [CrossRef] [Green Version]
- Du, X.; Ma, H.; Zhang, X.; Liu, K.; Peng, J.; Lan, Q. Characterization of the sterol carrier protein-x/sterol carrier protein-2 gene in the cotton bollworm, Helicoverpa armigera. J. Insect Physiol. 2012, 58, 1413–1423. [Google Scholar] [CrossRef]
- Ma, H.; Ma, Y.; Liu, X.; Dyer, D.; Xu, P.; Liu, K. NMR structure and function of Helicoverpa armigera sterol carrier protein-2, an important insecticidal target from the cotton bollworm. Sci Rep. 2015, 5, 18186. [Google Scholar] [CrossRef]
- Fire, A.; Xu, S.; Montgomery, M.; Kostas, S.; Driver, S.; Mello, C. Potent and specific genetic interference by double-stranded RNA in Caenorhabditis elegans. Nature 1998, 391, 806–811. [Google Scholar] [CrossRef]
- Zamore, P. RNA interference: Listening to the sound of silence. Nat. Struct Biol. 2001, 8, 746–750. [Google Scholar] [CrossRef]
- Zhang, H.; Li, H.; Miao, X. Feasibility, limitation and possible solutions of RNAi-based technology for insect pest control. Insect Sci. 2013, 20, 15–30. [Google Scholar] [CrossRef]
- Huvenne, H.; Smagghe, G. Mechanisms of dsRNA uptake in insects and potential of RNAi for pest control: A review. J. Insect Physiol. 2009, 56, 227–235. [Google Scholar] [CrossRef]
- Bettencourt, R.; Terenius, O.; Faye, I. Hemolin gene silencing by ds-RNA injected into Cecropia pupae is lethal to next generation embryos. Insect Mol. Biol. 2002, 11, 267–271. [Google Scholar] [CrossRef]
- Mao, Y.; Cai, W.; Wang, J.; Hong, G.; Tao, X.; Wang, L. Silencing a cotton bollworm P450 monooxygenase gene by plant-mediated RNAi impairs larval tolerance of gossypol. Nat. Biotechnol. 2007, 25, 1307–1313. [Google Scholar] [CrossRef]
- Yu, N.; Christiaens, O.; Liu, J.; Niu, J.; Cappelle, K.; Caccia, S. Delivery of dsRNA for RNAi in insects: An overview and future directions. Insect Sci. 2013, 20, 4–14. [Google Scholar] [CrossRef]
- Clemens, J.; Worby, C.; Simonson, L.; Muda, M.; Maehama, T.; Hemmings, B. Use of double-stranded RNA interference in Drosophila cell lines to dissect signal transduction pathways. Proc. Natl. Acad. Sci. USA 2000, 97, 6499–6503. [Google Scholar] [CrossRef] [Green Version]
- Caplen, N.; Fleenor, J.; Fire, A.; Morgan, R. dsRNA-mediated gene silencing in cultured Drosophila cells: A tissue culture model for the analysis of RNA interference. Gene 2000, 252, 95–105. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Wu, M.; Wang, B.; Han, Z. Comparison of the RNA interference effects triggered by dsRNA and siRNA in Tribolium castaneum. Pest Manag. Sci. 2013, 69, 781–786. [Google Scholar] [CrossRef]
- Turner, C.; Davy, M.; MacDiarmid, R.; Plummer, K.; Birch, N.; Newcomb, R. RNA interference in the light brown apple moth, Epiphyas postvittana (Walker) induced by double-stranded RNA feeding. Insect Mol. Biol. 2006, 15, 383–391. [Google Scholar] [CrossRef]
- Surakasi, V.; Mohamed, A.; Kim, Y. RNA interference of β1 integrin subunit impairs development and immune responses of the beet armyworm, Spodoptera exigua. J. Insect Physiol. 2011, 57, 1537–1544. [Google Scholar] [CrossRef]
- Lin, Y.; Huang, J.; Liu, Y.; Belles, X.; Lee, H. Oral delivery of dsRNA lipoplexes to German cockroach protects dsRNA from degradation and induces RNAi response. Pest Manag. Sci. 2017, 73, 960–966. [Google Scholar] [CrossRef] [PubMed]
- Joga, M.; Zotti, M.; Guy, S. RNAi efficiency, systemic properties, and novel delivery methods for pest insect control: What We Know So Far. Front. Physiol. 2016, 7, e553. [Google Scholar] [CrossRef] [Green Version]
- Rooijen, N.; Nieuwmegen, R. Liposomes in immunology: Multilamellar phosphatidylcholine liposomes as a simple, biodegradable and harmless adjuvant without any immunogenic activity of its own. Immunol. Commun. 2009, 9, 234–256. [Google Scholar] [CrossRef]
- Taning, C.; Christiaens, O.; Berkvens, N.; Casteels, H.; Maes, M.; Smagghe, G. Oral RNAi to control Drosophila suzukii: Laboratory testing against larval and adult stages. J. Pest Sci. 2016, 89, 803–814. [Google Scholar] [CrossRef]
- Khan, A.; Ashfaq, M.; Kiss, Z.; Khan, A.; Mansoor, S.; Falk, B. Use of recombinant tobacco mosaic virus to achieve RNA interference in plants against the citrus mealybug, Planococcus citri (Hemiptera: Pseudococcidae). PLoS ONE 2017, 8, e7365. [Google Scholar] [CrossRef]
- He, B.; Chu, Y.; Yin, M.; Müllen, K.; An, C.; Shen, J. Fluorescent nanoparticle delivered dsRNA toward genetic control of insect pests. Adv. Mater. 2013, 25, 4580–4584. [Google Scholar] [CrossRef] [PubMed]
- Zheng, G.; Li, C.; Zhou, H.; Li, S.; Li, G. Establishment of two new cell lines from the embryonic tissue of Helicoverpa armigera (Lepidoptera: Noctuidae) and their responses to baculovirus infection. Acta Entomol. Sin. 2010, 53, 167–174. [Google Scholar]
- Shao, H.; Zheng, W.; Liu, P.; Wang, Q.; Wang, J.; Zhao, X. Establishment of a new cell line from lepidopteran epidermis and hormonal regulation on the genes. PLoS ONE 2008, 3, e3127. [Google Scholar] [CrossRef]
- Kuthiala, A.; Ritter, K. Esterification of cholesterol and cholestanol in the whole body, tissues, and frass of Heliothis zea. Arch Insect Biochem. Physiol. 1988, 7, 237–248. [Google Scholar] [CrossRef]
- Zhang, X.; Zhang, J.; Zhu, K. Chitosan/double-stranded RNA nanoparticle-mediated RNA interference to silence chitin synthase genes through larval feeding in the African malaria mosquito (Anopheles gambiae). Insect Mol. Biol. 2010, 19, 683–693. [Google Scholar] [CrossRef] [PubMed]
- Shen, D.; Zhou, F.; Xu, Z.; He, B.; Li, M.; Shen, J. Systemically interfering with immune response by a fluorescent cationic dendrimer delivered gene suppression. J. Mater. Chem. B 2014, 2, 4653–4659. [Google Scholar] [CrossRef]
- Yang, Y.; Jiang, Q.; Peng, M.; Zhou, Z.; Du, X.; Yin, M.; Shen, J.; Yan, S. A star polyamine-based nanocarrier delivery system for enhanced avermectin contact and stomach toxicity against green peach aphids. Nanomaterials 2022, 12, 1445. [Google Scholar] [CrossRef]
- Das, S.; Debnath, N.; Cui, Y.; Unrine, J.; Palli, S. Chitosan carbon quantum dot, and silica nanoparticle mediated dsRNA delivery for gene silencing in Aedes aegypti: A comparative analysis. ACS Appl. Mater. Interfaces 2015, 7, 19530–19535. [Google Scholar] [CrossRef]
- Chariou, P.; Ortega-Rivera, O.; Steinmetz, N. Nanocarriers for the delivery of medical, veterinary, and agricultural active ingredients. ACS Nano. 2020, 14, 2678–2701. [Google Scholar] [CrossRef]
- Yan, S.; Li, N.; Guo, Y.; Chen, Y.; Ji, C.; Yin, M.; Shen, J.; Zhang, J. Chronic exposure to the star polycation (SPc) nanocarrier in the larval stage adversely impairs life history traits in Drosophila melanogaster. J. Nanobiotechnol. 2022, 20, 515. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Sequence (5′→3′) | Length (Base) | Tm | Efficiency |
|---|---|---|---|---|
| Q HaSCP-2F | GAGAAAGTACGAGGCATCTATGG | 23 | 58 | 99% |
| Q HaSCP-2R | ATCGCTGATGGTGAAGGTG | 19 | 55 | |
| Q rpS3A-F | CCTCGTGAGGCCAACAT | 17 | 54 | 98% |
| Q rpS3A-R | TGATGGCACGGACCTGAGT | 19 | 57 | |
| HaSCP-2 siRNA | Sense:5′-GCAAAUGAGCUGAUCACAUTT-3′ Antisense: 5′-AUGUGAUCAGCUCAUUUGCTT-3′ | 21 | ||
| FAM-siRNA (Negative control) | Sense:5′-UUCUCCGAACGUGUCACGUTT-3′ Antisense: 5′-ACGUGACACGUUCGGAGAATT-3′ | 21 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Geng, K.; Zhang, Y.; Zhao, X.; Zhang, W.; Guo, X.; He, L.; Liu, K.; Yang, H.; Hong, H.; Peng, J.; et al. Fluorescent Nanoparticle-RNAi-Mediated Silencing of Sterol Carrier Protein-2 Gene Expression Suppresses the Growth, Development, and Reproduction of Helicoverpa armigera. Nanomaterials 2023, 13, 245. https://doi.org/10.3390/nano13020245
Geng K, Zhang Y, Zhao X, Zhang W, Guo X, He L, Liu K, Yang H, Hong H, Peng J, et al. Fluorescent Nanoparticle-RNAi-Mediated Silencing of Sterol Carrier Protein-2 Gene Expression Suppresses the Growth, Development, and Reproduction of Helicoverpa armigera. Nanomaterials. 2023; 13(2):245. https://doi.org/10.3390/nano13020245
Chicago/Turabian StyleGeng, Kexin, Ying Zhang, Xi Zhao, Wanlin Zhang, Xinhan Guo, Lu He, Kaiyu Liu, Hong Yang, Huazhu Hong, Jianxin Peng, and et al. 2023. "Fluorescent Nanoparticle-RNAi-Mediated Silencing of Sterol Carrier Protein-2 Gene Expression Suppresses the Growth, Development, and Reproduction of Helicoverpa armigera" Nanomaterials 13, no. 2: 245. https://doi.org/10.3390/nano13020245
APA StyleGeng, K., Zhang, Y., Zhao, X., Zhang, W., Guo, X., He, L., Liu, K., Yang, H., Hong, H., Peng, J., & Peng, R. (2023). Fluorescent Nanoparticle-RNAi-Mediated Silencing of Sterol Carrier Protein-2 Gene Expression Suppresses the Growth, Development, and Reproduction of Helicoverpa armigera. Nanomaterials, 13(2), 245. https://doi.org/10.3390/nano13020245