Influence of the luxR Regulatory Gene Dosage and Expression Level on the Sensitivity of the Whole-Cell Biosensor to Acyl-Homoserine Lactone

 , , , ,
, , , ,
Abstract
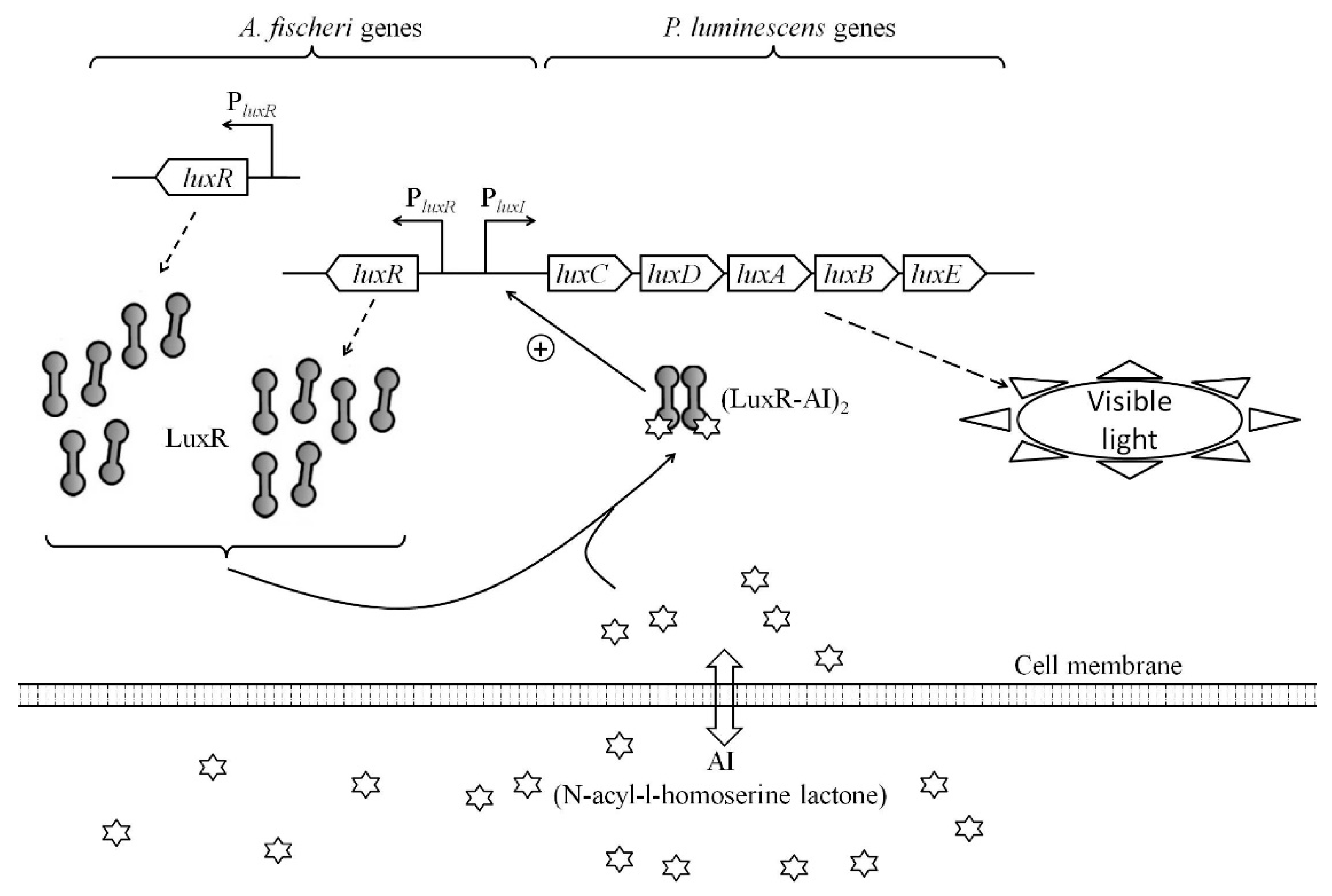
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Plasmids
2.2. Culture Medium and Growth Conditions
2.3. Measurement of Bioluminescence
2.4. Determination of the Minimum Detectable Concentration
2.5. Data Processing
2.6. DNA Manipulation
2.7. Chemicals
3. Results
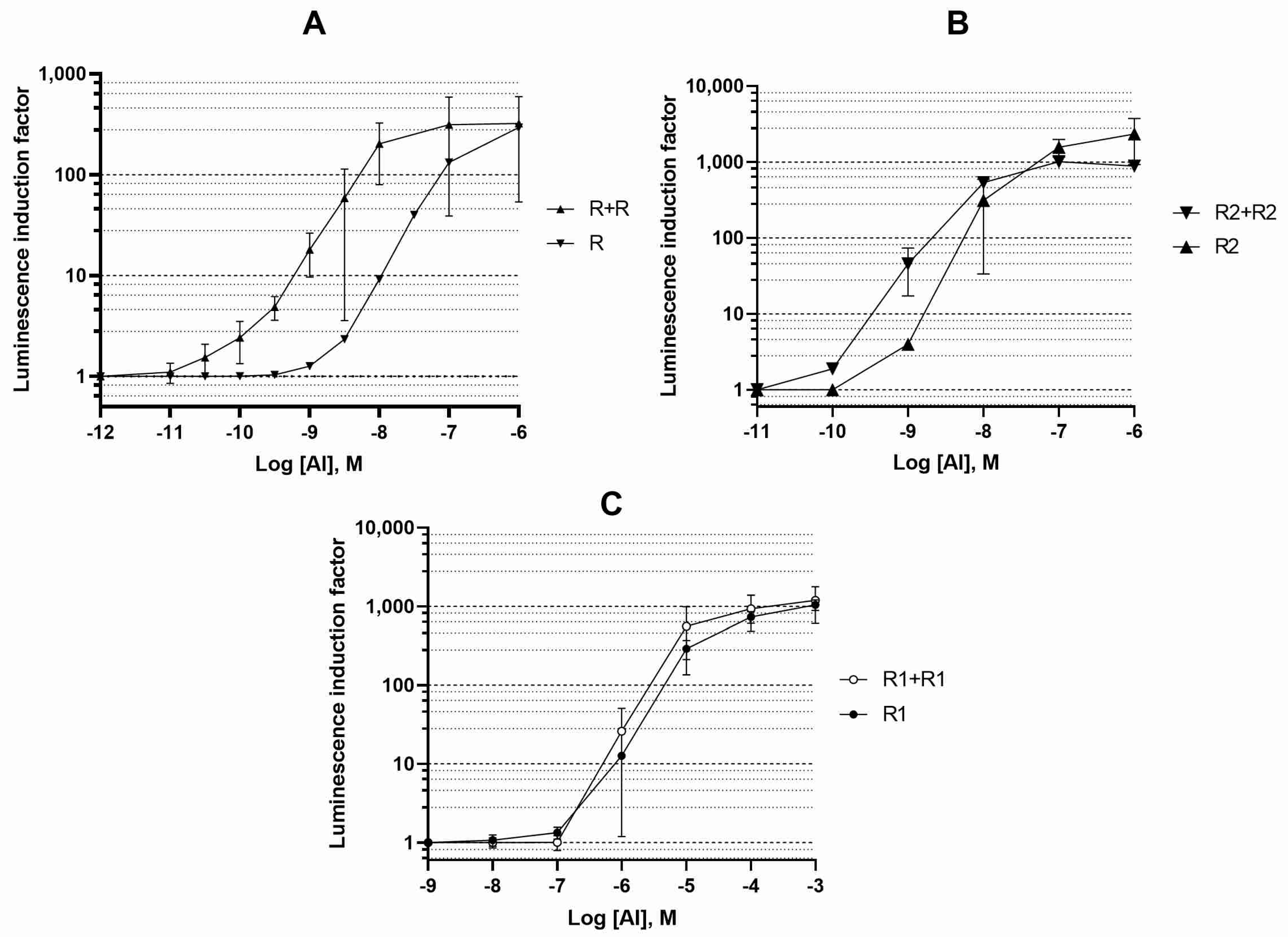
3.1. Influence of the Regulatory Gene Dosage on the Biosensor Sensitivity to AI
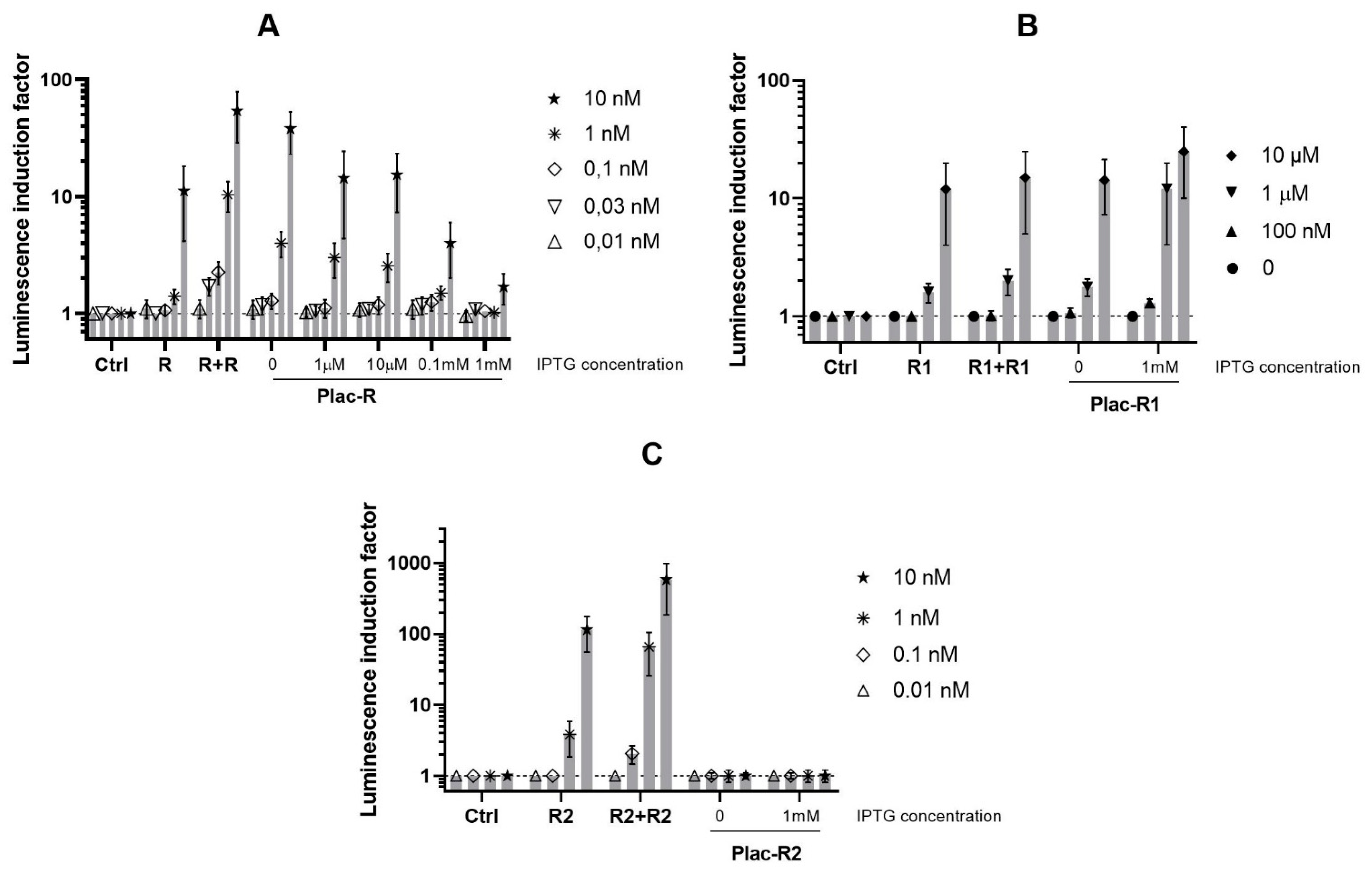
3.2. Influence of the luxR Genes Expression Level on the Sensitivity of the Biosensor
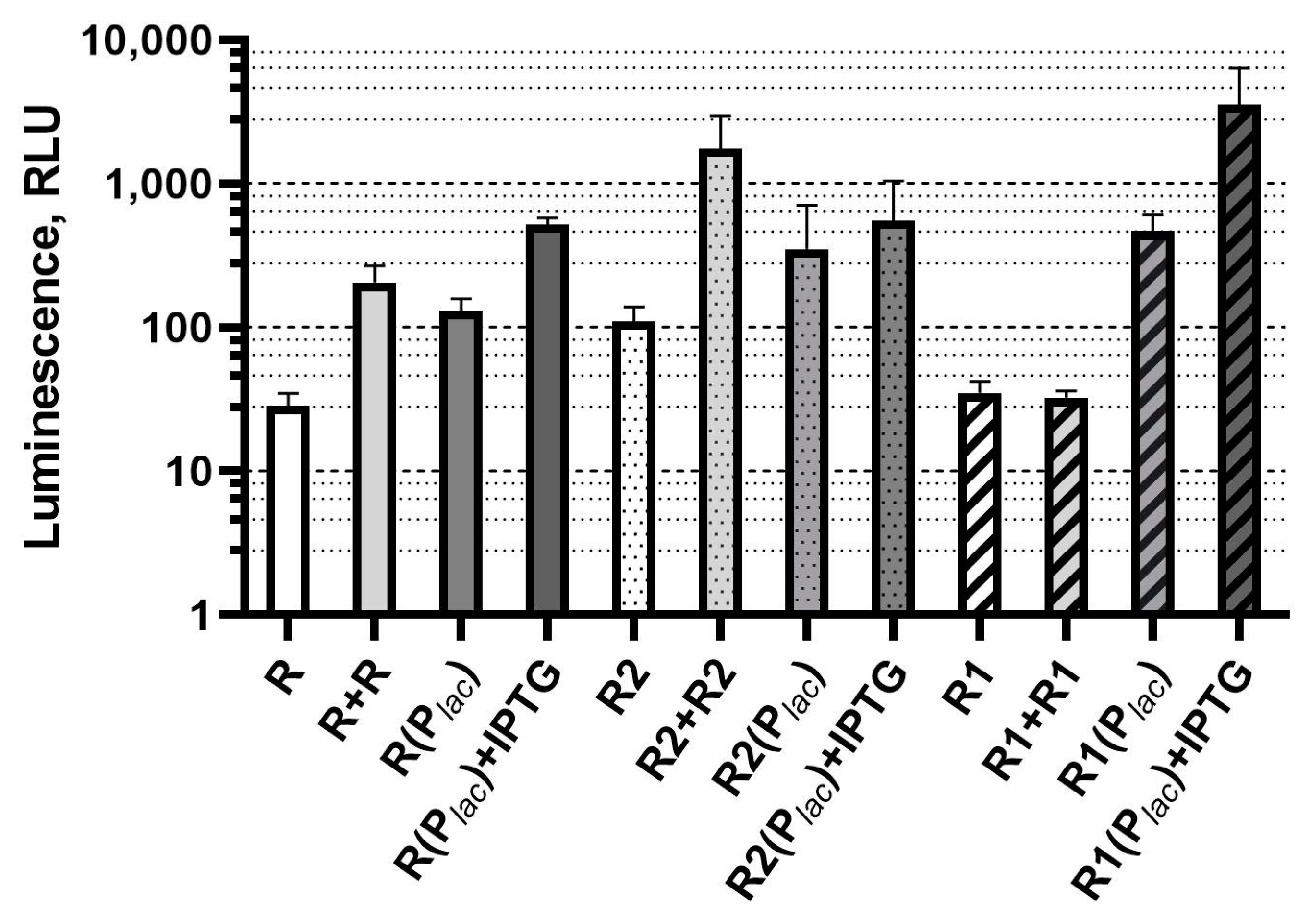
3.3. Influence of the luxR Dosage and Expression Level on the Base Luminescence of Biosensor Cells
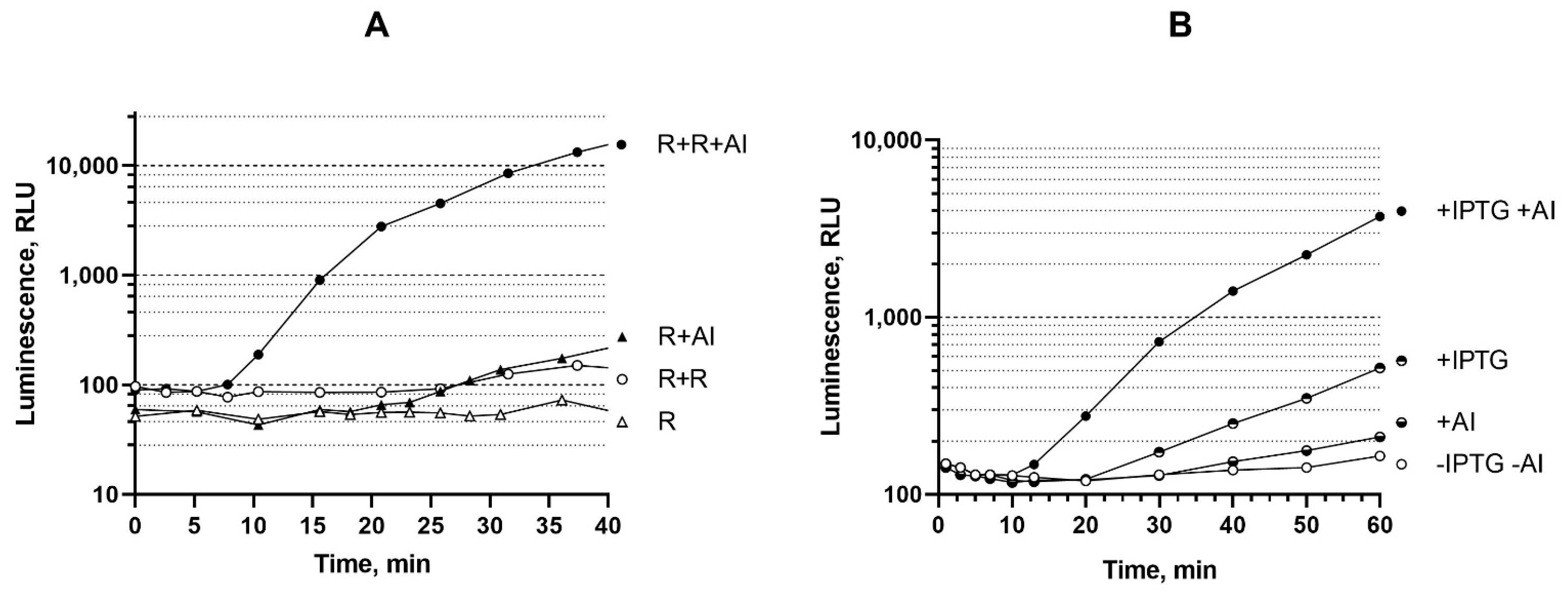
3.4. Influence of the Dosage and Expression Level of luxR Genes on the Biosensor Response Time
3.5. Testing the Lux-Biosensor E. coli MG1655 pVFR1 pSVRAF in Expedition Conditions
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nealson, K.H.; Platt, T.; Hastings, J.W. Cellular control of the synthesis and activity of the bacterial luminescent system. J. Bacteriol. 1970, 104, 313–322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eberhard, A.; Burlingame, A.L.; Eberhard, C.; Kenyon, G.L.; Nealson, K.H.; Oppenheimer, N.J. Structural identification of autoinducer of Photobacterium fischeri luciferase. Biochemistry 1981, 20, 2444–2449. [Google Scholar] [CrossRef]
- Choi, S.H.; Greenberg, E.P. Genetic evidence for multimerization of LuxR, the transcriptional activator of Vibrio fischeri luminescence. Mol. Mar. Biol. Biotechnol. 1992, 1, 408–413. [Google Scholar]
- Fidopiastis, P.M.; Sørum, H.; Ruby, E.G. Cryptic luminescence in the cold-water fish pathogen Vibrio salmonicida. Arch. Microbiol. 1999, 171, 205–209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manukhov, I.V.; Khrul’nova, S.A.; Baranova, A.; Zavilgelsky, G.B. Comparative analysis of the lux operons in Aliivibrio logei KCh1 (a Kamchatka Isolate) and Aliivibrio salmonicida. J. Bacteriol. 2011, 193, 3998–4001. [Google Scholar] [CrossRef] [Green Version]
- Konopleva, M.N.; Khrulnova, S.A.; Baranova, A.; Ekimov, L.V.; Bazhenov, S.V.; Goryanin, I.I.; Manukhov, I.V. A combination of luxR1 and luxR2 genes activates Pr-promoters of psychrophilic Aliivibrio logei lux-operon independently of chaperonin GroEL/ES and protease Lon at high concentrations of autoinducer. Biochem. Biophys. Res. Commun. 2016, 473, 1158–1162. [Google Scholar] [CrossRef] [PubMed]
- Ng, W.-L.; Bassler, B.L. Bacterial Quorum-Sensing Network Architectures. Annu. Rev. Genet. 2009, 43, 197–222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hansen, H.; Purohit, A.A.; Leiros, H.K.S.; Johansen, J.A.; Kellermann, S.J.; Bjelland, A.M.; Willassen, N.P. The autoinducer synthases LuxI and AinS are responsible for temperature-dependent AHL production in the fish pathogen Aliivibrio salmonicida. BMC Microbiol. 2015, 15, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Leipert, J.; Treitz, C.; Leippe, M.; Tholey, A. Identification and quantification of N-acyl homoserine lactones involved in bacterial communication by small-scale synthesis of internal dtandards and Matrix-Assisted Laser Desorption/Ionization Mass Spectrometry. J. Am. Soc. Mass Spectrom. 2017, 28, 2538–2547. [Google Scholar] [CrossRef]
- Chan, K.G.; Cheng, H.J.; Chen, J.W.; Yin, W.F.; Ngeow, Y.F. Tandem mass spectrometry detection of quorum sensing activity in multidrug resistant clinical isolate Acinetobacter baumannii. Sci. World J. 2014, 2014, 891041. [Google Scholar] [CrossRef]
- Miller, C.; Gilmore, J. Detection of Quorum-Sensing molecules for pathogenic molecules using cell-based and cell-free biosensors. Antibiotics 2020, 9, 259. [Google Scholar] [CrossRef]
- Andersen, J.B.; Heydorn, A.; Hentzer, M.; Eberl, L.; Geisenberger, O.; Christensen, B.B.; Molin, S.; Givskov, M. gfp-based N-acyl homoserine-lactone sensor systems for detection of bacterial communication. Appl. Environ. Microbiol. 2001, 67, 575–585. [Google Scholar] [CrossRef] [Green Version]
- Steindler, L.; Venturi, V. Detection of quorum-sensing N-acyl homoserine lactone signal molecules by bacterial biosensors. FEMS Microbiol. Lett. 2007, 266, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Asfour, H. Anti-quorum sensing natural compounds. J. Microsc. Ultrastruct. 2018, 6, 1. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Costantino, V.; Venturi, V.; Steindler, L. Quorum sensing inhibitors from the sea discovered using bacterial N-acyl-homoserine lactone-based biosensors. Mar. Drugs 2017, 15, 53. [Google Scholar] [CrossRef] [Green Version]
- Rai, N.; Rai, R.; Venkatesh, K.V. Quorum Sensing Biosensors. In Quorum Sensing vs Quorum Quenching: A battle with no end in sight; Springer India: New Delhi, India, 2015; pp. 171–183. [Google Scholar] [CrossRef]
- Pearson, J.P.; Gray, K.M.; Passador, L.; Tucker, K.D.; Eberhard, A.; Iglewski, B.H.; Greenberg, E.P. Structure of the autoinducer required for expression of Pseudomonas aeruginosa virulence genes. Proc. Natl. Acad. Sci. USA 1994, 91, 197–201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Winson, M.K.; Swift, S.; Fish, L.; Throup, J.P.; Jørgensen, F.; Chhabra, S.R.; Bycroft, B.W.; Williams, P.; Stewart, G.S.A. Construction and analysis of luxCDABE -based plasmid sensors for investigating N -acyl homoserine lactone-mediated quorum sensing. FEMS Microbiol. Lett. 1998, 163, 185–192. [Google Scholar] [CrossRef]
- Colton, D.M.; Stabb, E.V.; Hagen, S.J. Modeling analysis of signal sensitivity and specificity by Vibrio fischeri LuxR variants. PLoS ONE 2015, 10, e0126474. [Google Scholar] [CrossRef]
- Zhu, J.; Chai, Y.; Zhong, Z.; Li, S.; Winans, S.C. Agrobacterium bioassay strain for ultrasensitive detection of N-acylhomoserine lactone-type Quorum-Sensing molecules: Detection of autoinducers in Mesorhizobium huakuii. Appl. Environ. Microbiol. 2003, 69, 6949–6953. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fidopiastis, P.M.; Miyamoto, C.M.; Jobling, M.G.; Meighen, E.A.; Ruby, E.G. LitR, a new transcriptional activator in Vibrio fischeri, regulates luminescence and symbiotic light organ colonization. Mol. Microbiol. 2002, 45, 131–143. [Google Scholar] [CrossRef] [Green Version]
- Khrulnova, S.A.; Baranova, A.; Bazhenov, S.V.; Goryanin, I.I.; Konopleva, M.N.; Maryshev, I.V.; Salykhova, A.I.; Vasilyeva, A.V.; Manukhov, I.V.; Zavilgelsky, G.B. Lux-operon of the marine psychrophilic bacterium Aliivibrio logei: A comparative analysis of the LuxR1/LuxR2 regulatory activity in Escherichia coli cells. Microbiology 2016, 162, 717–724. [Google Scholar] [CrossRef] [PubMed]
- Melkina, O.E.; Goryanin, I.I.; Bazhenov, S.V.; Manukhov, I.V.; Zavilgelsky, G.B. Comparative analysis of Aliivibrio logei luxR1 and luxR2 genes regulation in Escherichia coli cells. Arch. Microbiol. 2019, 201, 1415–1425. [Google Scholar] [CrossRef]
- Zavil’gel’skiĭ, G.B.; Manukhov, I.V. Lon-protease participates in the regulation of transcription of the Lux-operon of Vibrio fischeri. Genetika 1994, 30, 337–341. [Google Scholar] [PubMed]
- Guyer, M.S.; Reed, R.R.; Steitz, J.A.; Low, K.B. Identification of a sex-factor-affinity site in E. coli as gamma delta. Cold Spring Harb. Symp. Quant. Biol. 1981, 45 Pt 1, 135–140. [Google Scholar] [CrossRef]
- Khrul’nova, S.A.; Manukhov, I.V.; Zavil’gel’skiĭ, G.B. "Quorum sensing" regulation of lux gene expression and the structure of lux operon in marine bacteria Alivibrio logei. Russ. J. Genet. 2011, 47, 1596–1603. [Google Scholar]
- Manukhov, I.V.; Kotova, V.I.; Zavil’gel’skiĭ, G.B. Host factors in the regulation of the Vibrio fischeri lux operon in Escherichia coli cells. Mikrobiologiia 2006, 75, 525–531. [Google Scholar] [CrossRef]
- Manukhov, I.V.; Melkina, O.E.; Goryanin, I.I.; Baranova, A.V.; Zavilgelsky, G.B. The N-terminal domain of Aliivibrio fischeri LuxR is a target of the GroEL chaperonin. J. Bacteriol. 2010, 192, 5549–5551. [Google Scholar] [CrossRef] [Green Version]
- Green, M.R.; Sambrook, J. Molecular Cloning: A Laboratory Manual, 4th ed.; Cold Spring Harbor Laboratory Press: New York, NY, USA, 2012. [Google Scholar]
- Chan, X.Y.; How, K.Y.; Yin, W.F.; Chan, K.G. N-Acyl homoserine lactone-mediated quorum sensing in Aeromonas veronii biovar sobria strain 159: Identification of LuxRI homologs. Front. Cell. Infect. Microbiol. 2016, 6, 7. [Google Scholar] [CrossRef] [Green Version]
- Kubitschek, H.E.; Friske, J.A. Determination of bacterial cell volume with the Coulter Counter. J. Bacteriol. 1986, 168, 1466–1467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, S.H.; Greenberg, E.P. The C-terminal region of the Vibrio fischeri LuxR protein contains an inducer-independent lux gene activating domain. Proc. Natl. Acad. Sci. USA 1991, 88, 11115–11119. [Google Scholar] [CrossRef] [Green Version]
- Mel’kina, O.E.; Manukhov, I.V.; Zavil’gel’skiǐ, G.B. The C-terminal domain of the Vibrio fischeri transcription activator LuxR is not essential for degradation by Lon protease. Mol. Biol. (Mosk.) 2010, 44, 515–519. [Google Scholar] [CrossRef] [PubMed]
- Bazhenov, S.V.; Khrulnova, S.A.; Konopleva, M.N.; Manukhov, I.V. Seasonal changes in luminescent intestinal microflora of the fish inhabiting the Bering and Okhotsk seas. FEMS Microbiol. Lett. 2019, 366. [Google Scholar] [CrossRef] [PubMed]
- Egidius, E.; Wiik, R.; Andersen, K. Vibrio salmonicida sp. nov., a new fish pathogen. Int. J. Syst. Bacteriol. 1986, 36, 518–520. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Genotype | Source |
|---|---|---|
| E. coli MG1655 | F-, λ-, ilvG-, rfb-50, rph-1 | [25] |
| Aeromonas spp. BCh1, BCh2, BCh3, and BCh4 | wild type | fish intestine, Sea of Azov, Taganrog Bay |
| Plasmid | Description | Source |
| pIVA | pDEW201 promoter-probe vector with luxR1 A. logei under control of PluxR1 A. logei and luxCDABE P. luminescens under control of PluxCDABEG A. logei, Apr | [22] |
| pSV16 | pDEW201, luxR2 A. logei under control of PluxR2 A. logei, luxCDABE P. luminescens under control of PluxI A. logei | [26] |
| pR2 | pDEW201, luxCDABE P. luminescens under control of PluxI A. logei | This study |
| pVFR1 | pDEW201, luxR A. fischeri under control of PluxR A. fischeri, luxCDABE P. luminescens under control of PluxICDABEG A. fischeri | [27] |
| pIV2 | pACYC184, luxR2 A. logei under control of PluxR2 inserted into BamHI site, Cmr | [22] |
| pIV3 | pACYC184, luxR1 A. logei under control of PluxR1 inserted into BamHI site, Cmr | [22] |
| pSVRAF | pACYC184, luxR2 A. logei under control of PluxR inserted into BamHI site, Cmr | [23] |
| pGEX-luxR | pGEX-KG vector containing the luxR A. fischeri gene under Ptac promoter; Apr | [27] |
| pOM | pACYC184 with a BamHI/NruI fragment of A. fischeri DNA from pF1 (luxICDABEG under the PluxICDABEG promoter and lux-regulatory DNA between luxR and luxI [without luxR]); Cmr | [28] |
| p15Tc-lac | Gene-expression vector obtained by the ligation of the pGex-KG plasmid fragment (lacI and Plac) with pACYC184 plasmid fragment (ori p15A and TcR), Tcr | This study |
| p15Tc-luxR1 | p15Tc-lac, luxR1 A. logei under control of Plac, Tcr luxR1 was isolated from pIVA and cloned into p15Tc-lac | This study |
| p15Tc-luxR2 | p15Tc-lac, luxR2 A. logei under control of Plac, Tcr luxR2 was isolated from pSV16 and cloned into p15Tc-lac | This study |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bazhenov, S.; Novoyatlova, U.; Scheglova, E.; Fomin, V.; Khrulnova, S.; Melkina, O.; Chistyakov, V.; Manukhov, I. Influence of the luxR Regulatory Gene Dosage and Expression Level on the Sensitivity of the Whole-Cell Biosensor to Acyl-Homoserine Lactone. Biosensors 2021, 11, 166. https://doi.org/10.3390/bios11060166
Bazhenov S, Novoyatlova U, Scheglova E, Fomin V, Khrulnova S, Melkina O, Chistyakov V, Manukhov I. Influence of the luxR Regulatory Gene Dosage and Expression Level on the Sensitivity of the Whole-Cell Biosensor to Acyl-Homoserine Lactone. Biosensors. 2021; 11(6):166. https://doi.org/10.3390/bios11060166
Chicago/Turabian StyleBazhenov, Sergey, Uliana Novoyatlova, Ekaterina Scheglova, Vadim Fomin, Svetlana Khrulnova, Olga Melkina, Vladimir Chistyakov, and Ilya Manukhov. 2021. "Influence of the luxR Regulatory Gene Dosage and Expression Level on the Sensitivity of the Whole-Cell Biosensor to Acyl-Homoserine Lactone" Biosensors 11, no. 6: 166. https://doi.org/10.3390/bios11060166
APA StyleBazhenov, S., Novoyatlova, U., Scheglova, E., Fomin, V., Khrulnova, S., Melkina, O., Chistyakov, V., & Manukhov, I. (2021). Influence of the luxR Regulatory Gene Dosage and Expression Level on the Sensitivity of the Whole-Cell Biosensor to Acyl-Homoserine Lactone. Biosensors, 11(6), 166. https://doi.org/10.3390/bios11060166