Development of the Sensing Platform for Protein Tyrosine Kinase Activity

 , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Preparation of Tyrosinase-Based Electrode
2.3. Fabrication of the Miniature Protein Tyrosine Kinase Sensing Platform
2.4. Protein Tyrosine Kinase Reaction
2.5. Electrochemical Measurement
3. Results and Discussion
3.1. Characterization of Miniature Detection Platform
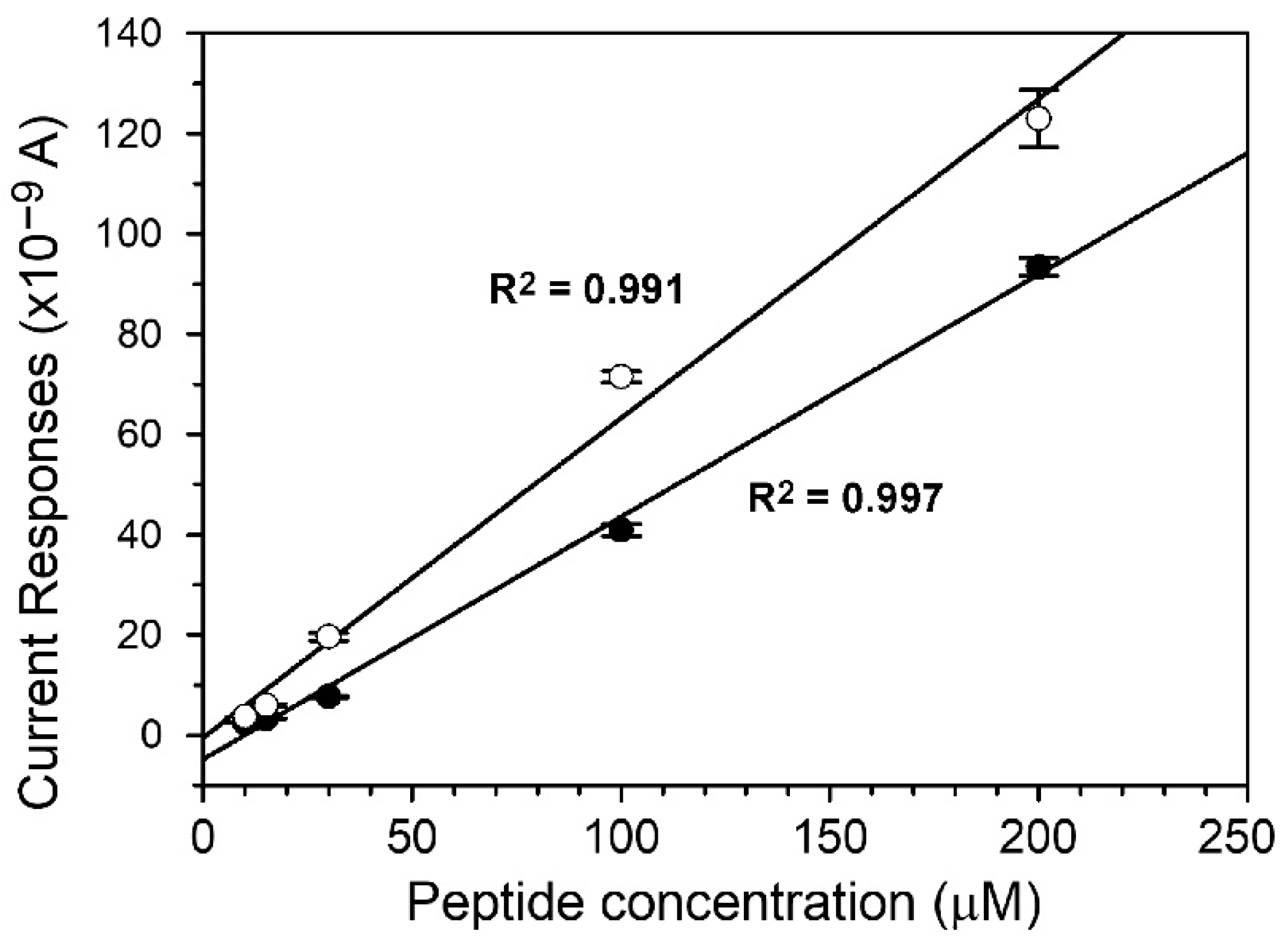
3.2. Detection of Her2 and Hck Activity on the Miniature Sensing Platform
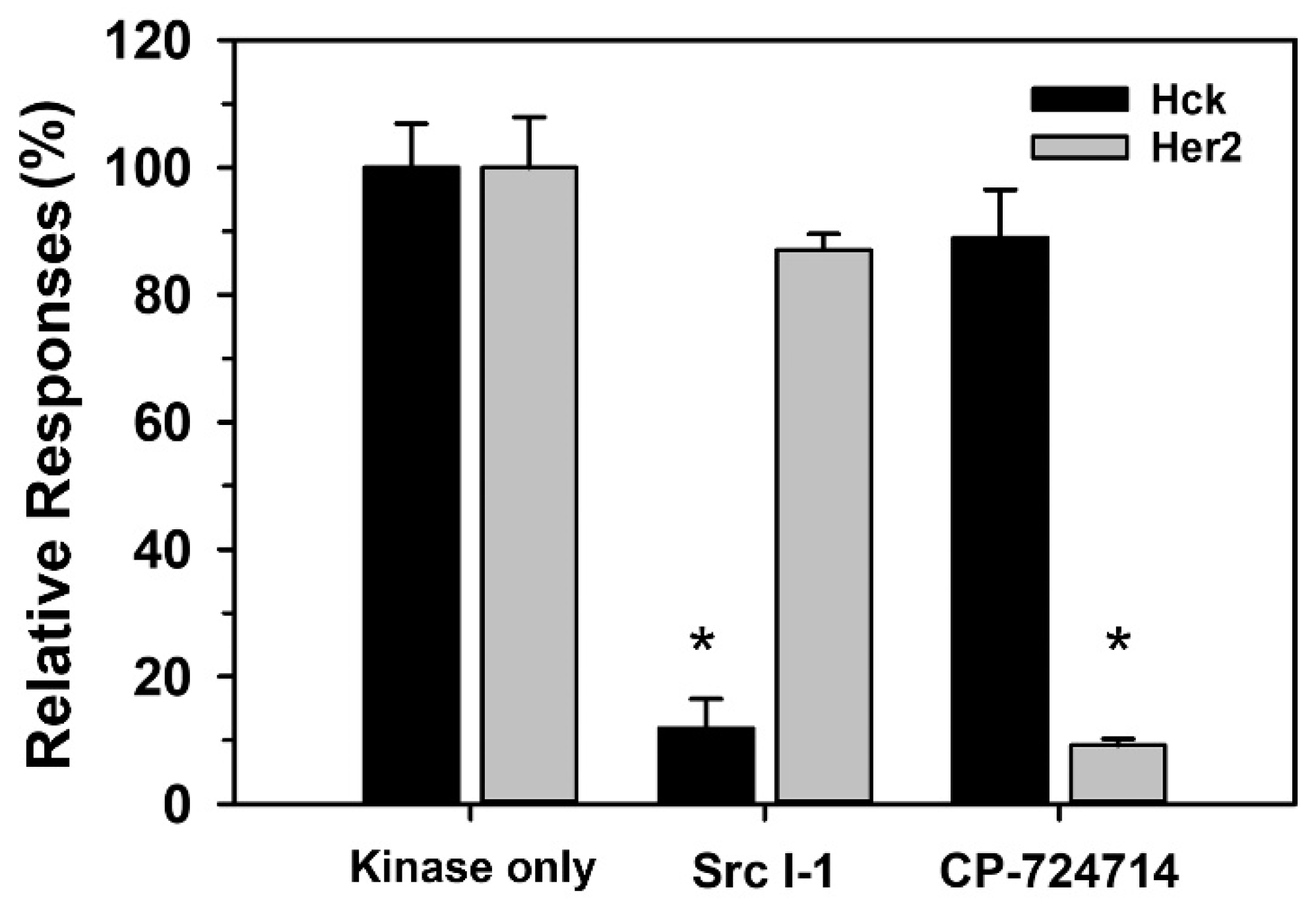
3.3. The Effect of Inhibitors to the Activity of Hck and Her2
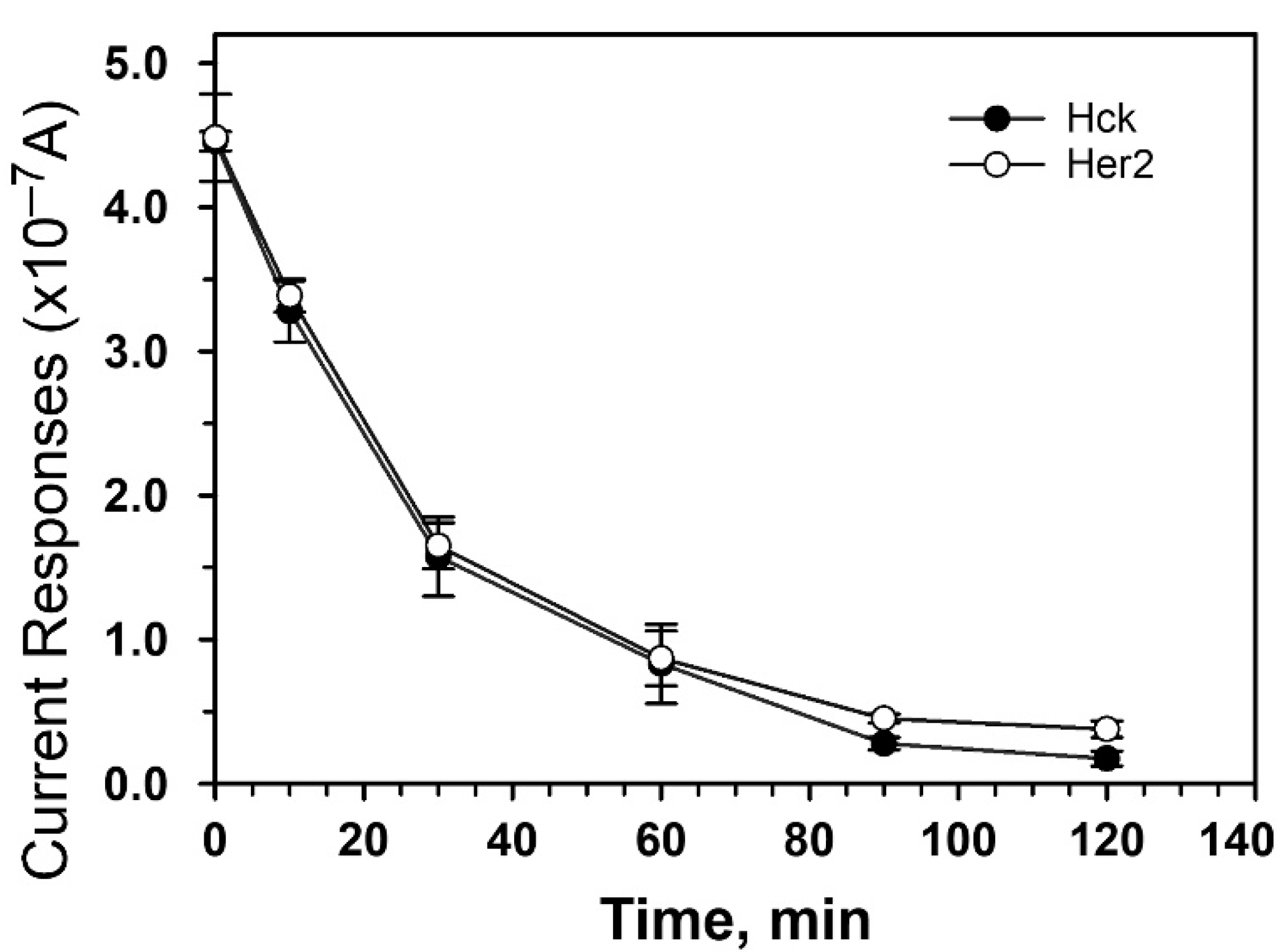
3.4. Interference Effect of Cultural Medium to the Protein Tyrosine Kinase Activity
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hunter, T. Protein kinases and phosphatases: The yin and yang of protein phosphorylation and signaling. Cell 1995, 80, 225–236. [Google Scholar] [CrossRef] [Green Version]
- Manning, G.; Whyte, D.B.; Martinez, R.; Hunter, T.; Sudarsanam, S. The Protein Kinase Complement of the Human Genome. Science 2002, 298, 1912–1934. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Graves, J.D.; Krebs, E.G. Protein phosphorylation and signal transduction. Pharmacol. Ther. 1999, 82, 111–121. [Google Scholar] [CrossRef]
- Hunter, T.; Plowman, G.D. The protein kinases of budding yeast: Six score and more. Trends Biochem. Sci. 1997, 1, 18–22. [Google Scholar] [CrossRef]
- Hubbard, S.R.; Till, J.H. Protein Tyrosine Kinase Structure and Function. Annu. Rev. Biochem. 2000, 69, 373–398. [Google Scholar] [CrossRef] [Green Version]
- Paul, M.K.; Mukhopadhyay, A.K. Tyrosine kinase—Role and significance in Cancer. Int. J. Med. Sci. 2004, 1, 101–115. [Google Scholar] [CrossRef] [Green Version]
- Vlahovic, G.; Crawford, J. Activation of tyrosine kinases in cancer. Oncologist 2003, 8, 531–538. [Google Scholar] [CrossRef]
- Anselmo, A.N.; Cobb, M.H. Protein kinase function and glutathionylation. Biochem. J. 2004, 381, 675–677. [Google Scholar] [CrossRef] [Green Version]
- Gould, K.I.; Nurse, P. Tyrosine phosphorylation of the fission yeast cdc2+ protein kinase regulates entry into mitosis. Nature 1989, 342, 39–45. [Google Scholar] [CrossRef]
- Giamas, G.; Stebbing, J.; Vorgias, C.E.; Knippschild, U. Protein kinases as targets for cancer treatment. Pharmacogenomics 2007, 8, 1005–1016. [Google Scholar] [CrossRef] [PubMed]
- Normanno, N.; De Luca, A.; Bianco, C.; Strizzi, L.; Mancino, M.; Maiello, M.R.; Carotenuto, A.; De Feo, G.; Caponigro, F.; Salomon, D.S. Epidermal growth factor receptor (EGFR) signaling in cancer. Gene 2006, 366, 2–16. [Google Scholar] [CrossRef]
- Riese, D.J., 2nd; Stern, D.F. Specificity within the EGF family/ErbB receptor family signaling network. Bioessays 1998, 2, 41–48. [Google Scholar] [CrossRef]
- Moasser, M.M. The oncogene HER2; Its signaling and transforming functions and its role in human cancer pathogenesis. Oncogene 2007, 26, 6469–6487. [Google Scholar] [CrossRef] [Green Version]
- Lohrisch, C.; Piccart, M. An overview of HER2. Semi. Oncol. 2001, 28, 3–11. [Google Scholar] [CrossRef]
- Omar, N.; Yan, B.; Salto-Tellez, M. HER2: An emerging biomarker in non-breast and non-gastric cancers. Pathogenesis 2015, 2, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Ziegler, S.F.; Marth, J.D.; Lewis, D.B.; Perlmutter, R.M. Novel protein-tyrosine kinase gene (hck) preferentially expressed in cells of hematopoietic origin. Mol. Cell Biol. 1987, 7, 2276–2285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poh, A.R.; O’Donoghue, R.J.; Ernst, M. Hematopoietic cell kinase (HCK) as a therapeutic target in immune and cancer cells. Oncotarget 2015, 6, 15752–15771. [Google Scholar] [CrossRef] [Green Version]
- Pearson, R.B.; Kemp, B.E. Protein kinase phosphorylation site sequences and consensus specificity motifs: Tabulations. Methods Enzymol. 1991, 200, 62–81. [Google Scholar] [PubMed]
- Rothman, D.M.; Shults, M.D.; Imperiali, B. Chemical approaches for investigating phosphorylation in signal transduction networks. Trends Cell Biol. 2005, 15, 502–510. [Google Scholar] [CrossRef]
- Beausoleil, S.A.; Villén, J.; Gerber, S.A.; Rush, J.; Gygi, S.P. A probability-based approach for high-throughput protein phosphorylation analysis and site localization. Nat. Biotechnol. 2006, 24, 1285–1292. [Google Scholar] [CrossRef] [PubMed]
- Glenney, J.R., Jr.; Zokas, L.; Kamps, M.P. Monoclonal antibodies to phosphotyrosine. J. Immunol. Method 1988, 109, 277–285. [Google Scholar] [CrossRef]
- Freeman, R.; Finder, T.; Gill, R.; Willner, I. Probing Protein Kinase (CK2) and Alkaline Phosphatase with CdSe/ZnS Quantum Dots. Nano Lett. 2010, 10, 2192–2196. [Google Scholar] [CrossRef] [PubMed]
- Kiew, L.V.; Chang, C.-Y.; Huang, S.-Y.; Wang, P.-W.; Heh, C.-H.; Liu, C.-T.; Cheng, C.-H.; Lu, Y.-X.; Chen, Y.-C.; Huang, Y.-X.; et al. Development of flexible electrochemical impedance spectroscopy-based biosensing platform for rapid screening of SARS-CoV-2 inhibitors. Biosens. Bioelectron. 2021, 183, 113213. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.-Y.; Kung, Y.-A.; Huang, P.-N.; Chang, S.-Y.; Gong, Y.-N.; Han, Y.-J.; Chiang, H.-J.; Liu, K.-T.; Lee, K.-M.; Chang, C.-Y.; et al. Stability of SARS-CoV-2 Spike G614 Variant Surpasses That of the D614 Variant after Cold Storage. mSphere 2021, 6, e00104-21. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.-Y.; Huang, Y.-T.; Chang, P.-C.; Su, C.-H.; Hsu, K.-C.; Li, X.; Wu, C.-H.; Chang, C.-C. Surface active flexible palladium nano-thin-film electrode development for biosensing. Inorg. Chem. Commun. 2019, 107, 107461. [Google Scholar] [CrossRef]
- Chang, C.-Y.; Chen, W.; Su, C.-H.; Chang, P.-C.; Huang, Y.-T.; Hsu, K.-C.; Yuan, C.-J.; Chang, C.-C. Enhanced Bioconjugation on Sputtered Palladium Nano-thin-film Electrode. Appl. Phys. Lett. 2019, 114, 093702. [Google Scholar] [CrossRef] [Green Version]
- Tan, D.; Li, F.; Zhou, B. Electrochemical assay methods for protein kinase activity. Int. J. Electrochem. Sci. 2019, 14, 5707–5725. [Google Scholar] [CrossRef]
- Song, H.; Kerman, K.; Kraatz, H.-B. Electrochemical detection of kinase-catalyzed phosphorylation using ferrocene-conjugated ATP. Chem. Commun. 2008, 4, 502–504. [Google Scholar] [CrossRef] [PubMed]
- Kerman, K.; Song, H.; Duncan, J.S.; Litchfield, D.W.; Kraatz, H.-B. Peptide Biosensors for the Electrochemical Measurement of Protein Kinase Activity. Anal. Chem. 2008, 80, 9395–9401. [Google Scholar] [CrossRef]
- Marti, S.; Labib, M.; Kraatz, H.-B. Enzymatically modified peptide surfaces: Towards general electrochemical sensor platform for protein kinase catalyzed phosphorylations. Analyst 2011, 136, 107–112. [Google Scholar] [CrossRef] [PubMed]
- Kerman, K.; Kraatz, H.-B. Electrochemical detection of protein tyrosine kinase-catalysed phosphorylation using gold nanoparticles. Biosens. Bioelectron. 2009, 24, 1484–1489. [Google Scholar] [CrossRef] [PubMed]
- Kerman, K.; Miyuki Chikae, M.; Yamamura, S.; Tamiya, E. Gold nanoparticle-based electrochemical detection of protein phosphorylation. Anal. Chim. Acta 2007, 588, 26–33. [Google Scholar] [CrossRef] [PubMed]
- Ji, J.; Yang, H.; Liu, Y.; Chen, H.; Kong, J.; Liu, B. TiO2-assisted silver enhanced biosensor for kinase activity profiling. Chem. Commun. 2009, 1508–1510. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Cao, Y.; Li, Y.; Liang, Z.; Li, G. Electrochemical strategy for detection of phosphorylation based on enzyme-linked Electrocatalysis. J. Electroanal. Chem. 2011, 656, 274–278. [Google Scholar] [CrossRef]
- Kerman, K.; Vestergaard, M.; Tamiya, E. Label-Free Electrical Sensing of Small-Molecule Inhibition on Tyrosine Phosphorylation. Anal. Chem. 2007, 79, 6881–6885. [Google Scholar] [CrossRef] [PubMed]
- Chikua, M.; Horisawab, K.; Doib, N.; Yanagawab, H.; Einaga, Y. Electrochemical detection of tyrosine derivatives and protein tyrosine kinase activity using boron-doped diamond electrodes. Biosens. Bioelectron. 2010, 26, 235–240. [Google Scholar] [CrossRef]
- Li, B.; Shi, X.; Gu, W.; Zhao, K.; Chen, N.; Xian, Y. Graphene based electrochemical biosensor for label-free measurement of the activity and inhibition of protein. tyrosine kinase. Analyst 2013, 138, 7212–7217. [Google Scholar] [CrossRef]
- Wang, C.-L.; Wei, L.-Y.; Yuan, C.-J.; Hwang, K.-C. Reusable amperometric biosensor for measuring protein tyrosine kinase activity. Anal. Chem. 2012, 84, 971–977. [Google Scholar] [CrossRef]
- Yokoyama, N.; deBakker, C.D.; Zappacosta, F.; Huddleston, M.J.; Annan, R.S.; Ravichandran, K.S.; Miller, W.T. Identification of tyrosine residues on ELMO1 that are phosphorylated by the Src-family kinase Hck. Biochemistry 2005, 44, 8841–8849. [Google Scholar] [CrossRef] [Green Version]
- Tian, G.; Cory, M.; Smith, A.A.; Knight, W.B. Structural Determinants for Potent, Selective Dual Site Inhibition of Human pp60c-src by 4-Anilinoquinazolines. Biochemistry 2001, 40, 7084–7091. [Google Scholar] [CrossRef]
- Arkin, M.; Moasser, M.M. HER2 directed small molecule antagonists. Curr. Opin. Investig. Drug 2008, 9, 1264–1276. [Google Scholar]
- Sanada, H.; Suzue, R.; Nakashima, Y.; Kawada, S. Effect of thiol compounds on melanin formation by tyrosinase. Biochim. Biophysic. Acta 1972, 261, 258–266. [Google Scholar] [CrossRef]
- Liu, L.; Cheng, C.; Chang, Y.; Ma, H.; Hao, Y. Two sensitive electrochemical strategies for the detection of protein kinase activity based on the 4-mercaptophenylboronic acid-induced in situ assembly of silver nanoparticles. Sens. Actuat. B Chem. 2017, 248, 178–186. [Google Scholar] [CrossRef]
- Yang, Y.; Guo, L.H.; Qu, N.; Wei, M.Y.; Zhao, L.X.; Wan, B. Label-free electrochemical measurement of protein tyrosine kinase activity and inhibition based on electro-catalyzed tyrosine signaling. Biosens. Bioelectron. 2011, 28, 284–290. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Electrode Types | Working Mechanism | Reusability (R.S.D.) | Sample Volume Required (μL) | Protein Tyrosine Kinases | Linear Range of Detection | Limitation of Detection | Ref. |
|---|---|---|---|---|---|---|---|
| Miniature Tyrosinase/CFP | Tyrosinase-based Tyr Oxidation | High (6.6%) | Src | N.D. | N.D. | ||
| 1–2 μL | Hck | 1–100 U/mL | 1 U/mL | This study | |||
| HerB | 1–100 U/mL | 1 U/mL | |||||
| Peptide-immobilized SPCE | AuNP-based redox response | Low (N.A.) | 25 μL | Src | N.A. | 5 U/mL | [32] |
| MWCNT -modified SPCE | Direct oxidation of Tyr | High (N.A.) | 20 μL | Src | N.A. | 5 U/mL | [35] |
| Graphene-modified glassy carbon electrode | Graphene-assisted direct oxidation of Tyr | High (N.A.) | 20 μL | Src | 0.26 to 33.79 nM | 0.087 nM | [37] |
| Tyrosinase/CFP | Tyrosinase-based Tyr Oxidation | High (2.87%) | 20 μL | Src | 1.9–237.6 U/mL | 0.23 U/mL | [38] |
| Peptide-immobilized Gold electrode | 4-mercaptophenylboronic acid (MPBA)MPBA-assisted AgNP aggregates-based redox response | Low (N.A.) | -- | Src | 10–80 ng/mL | 1.2 ng/mL | [43] |
| Peptide-immobilized ITO electrode | Os(bpy)3+2-mediate Tyr oxidation | Low (N.A.) | -- | EGFR | N.A. | 1 U/mL | [44] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wei, L.-Y.; Lin, W.; Leo, B.-F.; Kiew, L.-V.; Chang, C.-C.; Yuan, C.-J. Development of the Sensing Platform for Protein Tyrosine Kinase Activity. Biosensors 2021, 11, 240. https://doi.org/10.3390/bios11070240
Wei L-Y, Lin W, Leo B-F, Kiew L-V, Chang C-C, Yuan C-J. Development of the Sensing Platform for Protein Tyrosine Kinase Activity. Biosensors. 2021; 11(7):240. https://doi.org/10.3390/bios11070240
Chicago/Turabian StyleWei, Lan-Yi, Wei Lin, Bey-Fen Leo, Lik-Voon Kiew, Chia-Ching Chang, and Chiun-Jye Yuan. 2021. "Development of the Sensing Platform for Protein Tyrosine Kinase Activity" Biosensors 11, no. 7: 240. https://doi.org/10.3390/bios11070240
APA StyleWei, L.-Y., Lin, W., Leo, B.-F., Kiew, L.-V., Chang, C.-C., & Yuan, C.-J. (2021). Development of the Sensing Platform for Protein Tyrosine Kinase Activity. Biosensors, 11(7), 240. https://doi.org/10.3390/bios11070240