Detection of SARS-CoV-2 Neutralizing Antibodies in Vaccinated Pregnant Women and Neonates by Using a Lateral Flow Immunoassay Coupled with a Spectrum-Based Reader

 ,
,  ,
,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Patient Selection
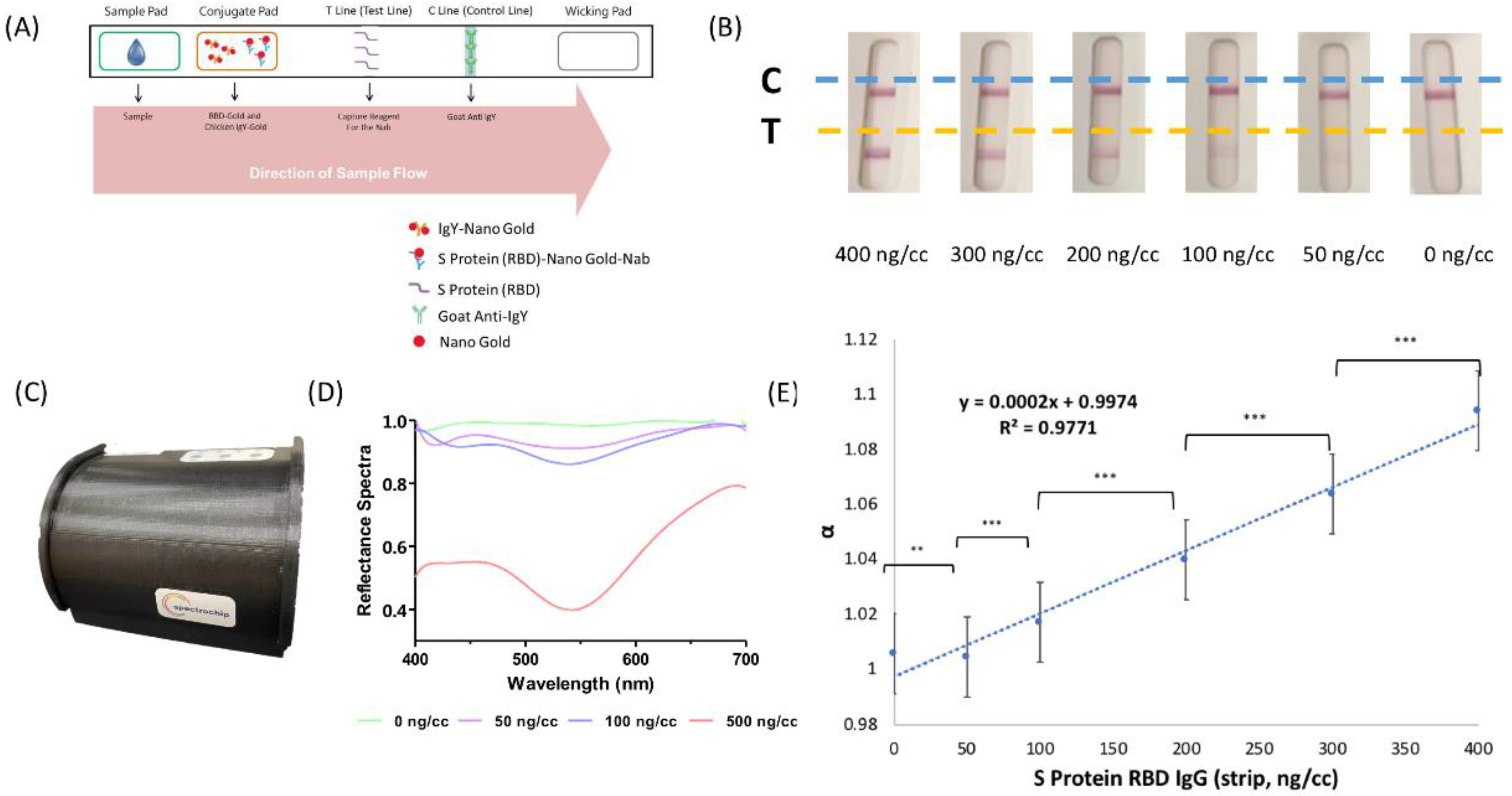
2.2. Rapid Test Strip by Lateral Flow Immunoassays
2.3. Reflectance Spectral Analysis
2.4. S1 Receptor Binding Domain IgG Antibody Detection by Enzyme-Linked Immunosorbent Assay (ELISA)
2.5. Neutralizing Antibody Inhibition Test of Wild-Type, Delta-Type, and Omicron-Type SARS-CoV-2
2.6. Statistics
3. Results
3.1. Participant Characteristics
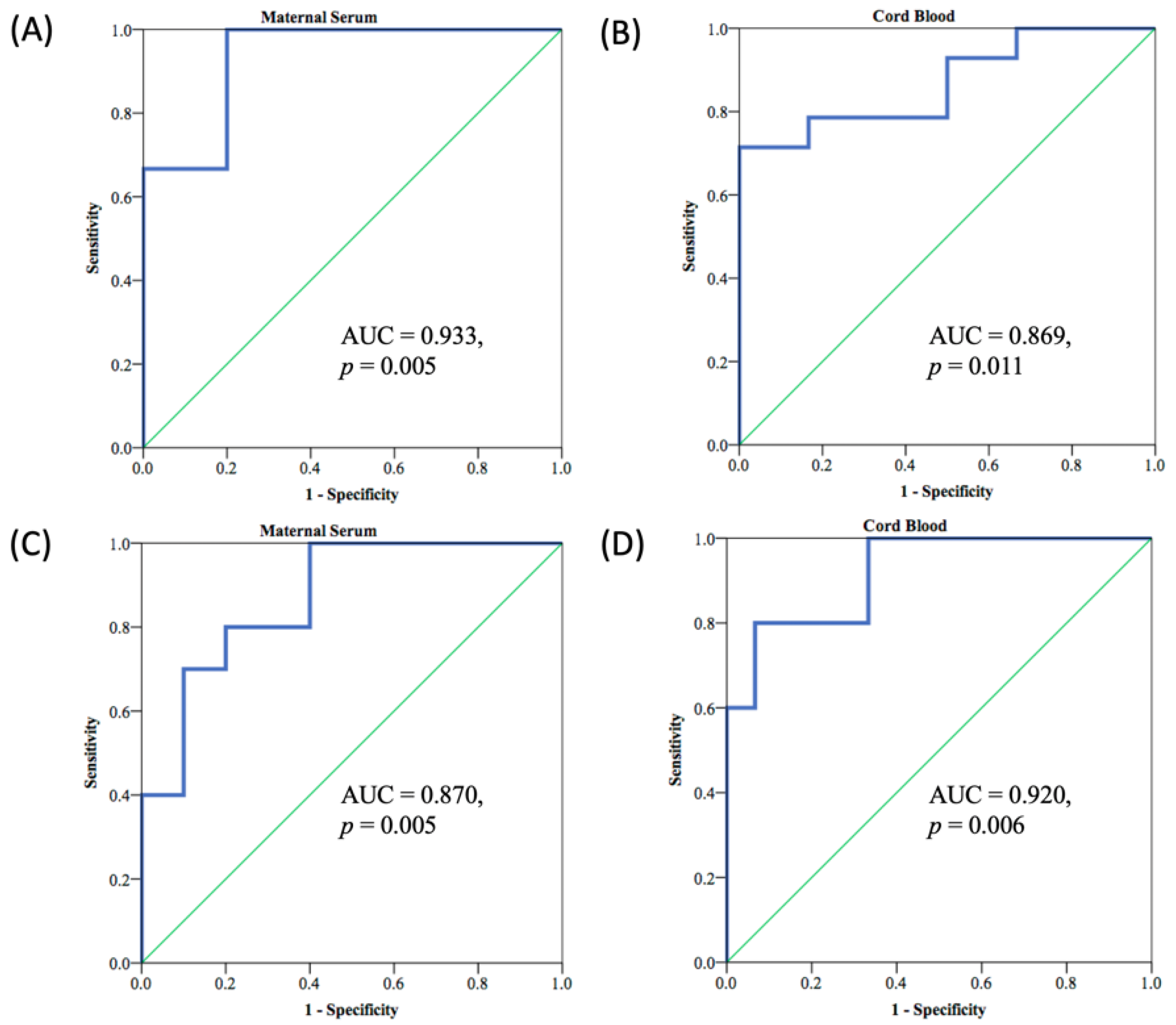
3.2. Neutralizing Antibodies in Maternal and Neonatal Serum
3.3. SRBD IgG Detection by Lateral Flow Immunoassay with Spectrum Analyzer
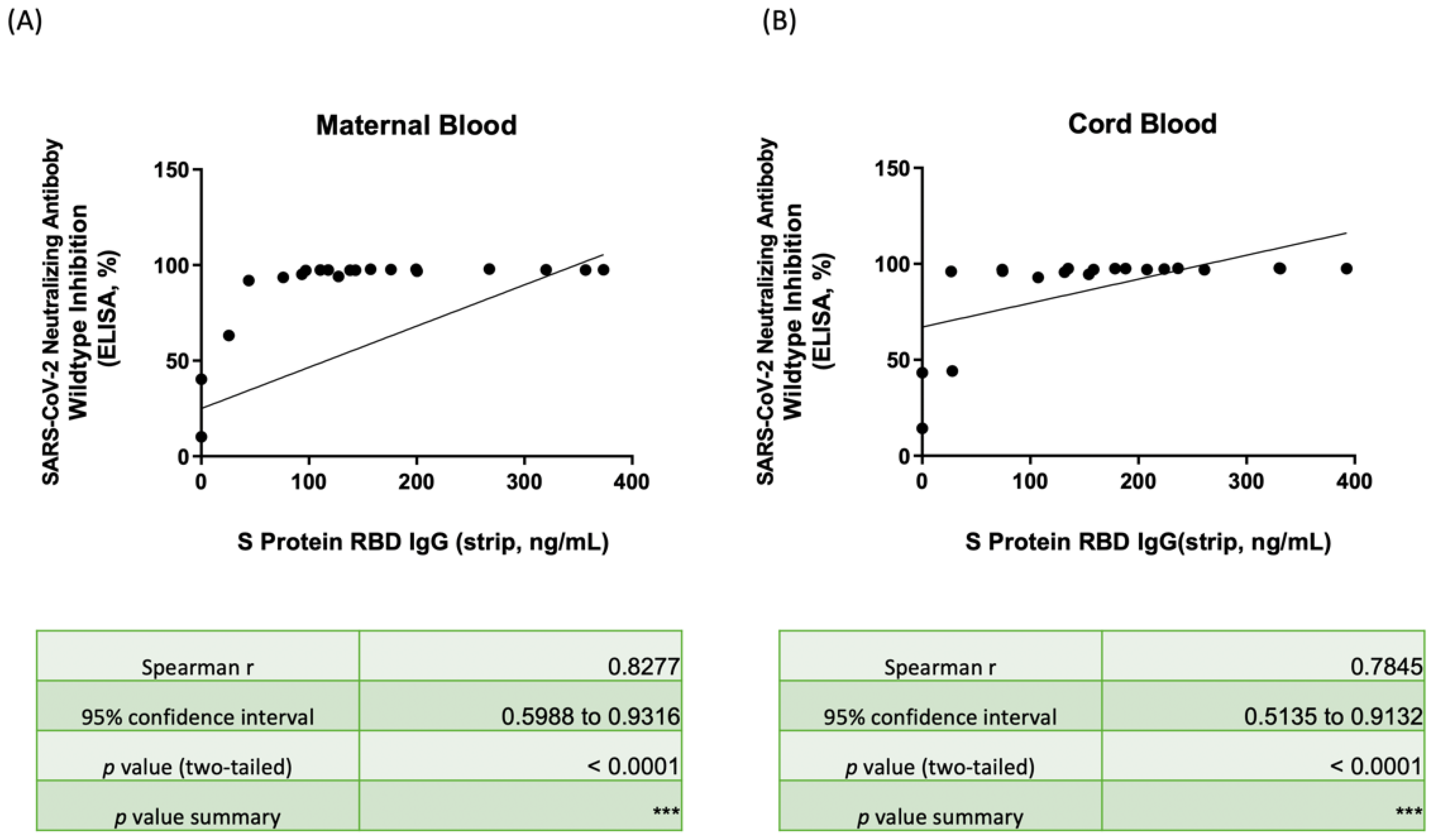
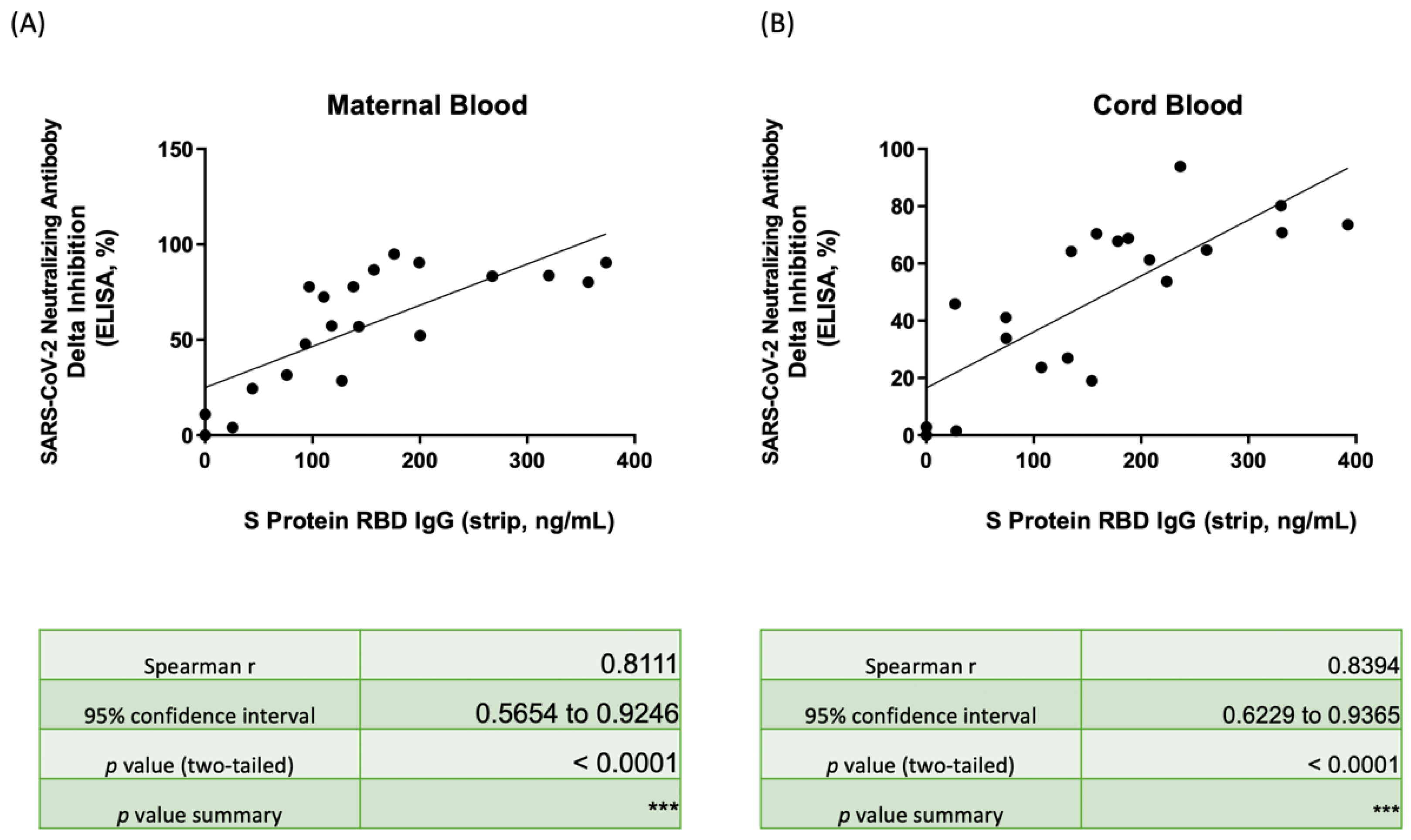
3.4. Nab Inhibition Rates of Wild-, Delta-, and Omicron-Type SARS-CoV-2 with SRBD IgG Level
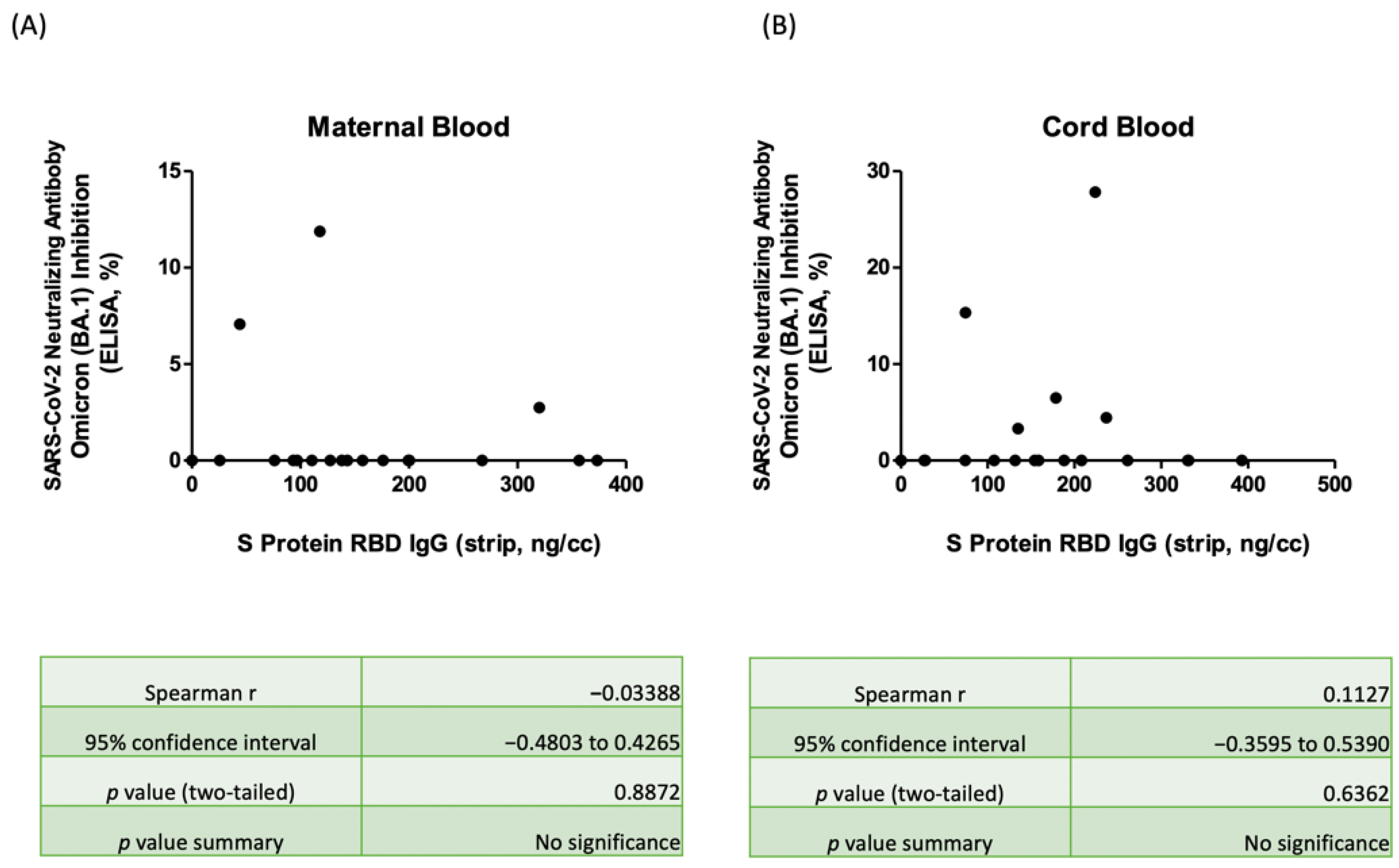
3.5. Correlation of Nab Concentration and SRBD IgG Level
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A


References
- Lu, R.; Zhao, X.; Li, J.; Niu, P.; Yang, B.; Wu, H.; Wang, W.; Song, H.; Huang, B.; Zhu, N.; et al. Genomic characterisation and epidemiology of 2019 novel coronavirus: Implications for virus origins and receptor binding. Lancet 2020, 395, 565–574. [Google Scholar] [CrossRef] [Green Version]
- Institute for Health Metrics and Evaluation. COVID-19 Results Briefing. Available online: https://www.healthdata.org/sites/default/files/files/1_briefing_Global_5.pdf (accessed on 21 January 2022).
- Polack, F.P.; Thomas, S.J.; Kitchin, N.; Absalon, J.; Gurtman, A.; Lockhart, S.; Perez, J.L.; Pérez Marc, G.; Moreira, E.D.; Zerbini, C.; et al. Safety and Efficacy of the BNT162b2 mRNA Covid-19 Vaccine. N. Engl. J. Med. 2020, 383, 2603–2615. [Google Scholar] [CrossRef] [PubMed]
- Baden, L.R.; El Sahly, H.M.; Essink, B.; Kotloff, K.; Frey, S.; Novak, R.; Diemert, D.; Spector, S.A.; Rouphael, N.; Creech, C.B.; et al. Efficacy and Safety of the mRNA-1273 SARS-CoV-2 Vaccine. N. Engl. J. Med. 2021, 384, 403–416. [Google Scholar] [CrossRef] [PubMed]
- Butt, A.A.; Omer, S.B.; Yan, P.; Shaikh, O.S.; Mayr, F.B. SARS-CoV-2 Vaccine Effectiveness in a High-Risk National Population in a Real-World Setting. Ann. Intern. Med. 2021, 174, 1404–1408. [Google Scholar] [CrossRef]
- Paris, C.; Perrin, S.; Hamonic, S.; Bourget, B.; Roué, C.; Brassard, O.; Tadié, E.; Gicquel, V.; Bénézit, F.; Thibault, V.; et al. Effectiveness of mRNA-BNT162b2, mRNA-1273, and ChAdOx1 nCoV-19 vaccines against COVID-19 in healthcare workers: An observational study using surveillance data. Clin. Microbiol. Infect. 2021, 27, 1699.e5–1699.e8. [Google Scholar] [CrossRef] [PubMed]
- Allotey, J.; Stallings, E.; Bonet, M.; Yap, M.; Chatterjee, S.; Kew, T.; Debenham, L.; Llavall, A.C.; Dixit, A.; Zhou, D.; et al. Clinical manifestations, risk factors, and maternal and perinatal outcomes of coronavirus disease 2019 in pregnancy: Living systematic review and meta-analysis. BMJ 2020, 370, m3320. [Google Scholar] [CrossRef] [PubMed]
- Villar, J.; Ariff, S.; Gunier, R.B.; Thiruvengadam, R.; Rauch, S.; Kholin, A.; Roggero, P.; Prefumo, F.; do Vale, M.S.; Cardona-Perez, J.A.; et al. Maternal and Neonatal Morbidity and Mortality Among Pregnant Women With and Without COVID-19 Infection: The INTERCOVID Multinational Cohort Study. JAMA Pediatr. 2021, 175, 817–826. [Google Scholar] [CrossRef] [PubMed]
- Pettirosso, E.; Giles, M.; Cole, S.; Rees, M. COVID-19 and pregnancy: A review of clinical characteristics, obstetric outcomes and vertical transmission. Aust. N. Z. J. Obstet. Gynaecol. 2020, 60, 640–659. [Google Scholar] [CrossRef]
- Gurol-Urganci, I.; Jardine, J.E.; Carroll, F.; Draycott, T.; Dunn, G.; Fremeaux, A.; Harris, T.; Hawdon, J.; Morris, E.; Muller, P.; et al. Maternal and perinatal outcomes of pregnant women with SARS-CoV-2 infection at the time of birth in England: National cohort study. Am. J. Obstet. Gynecol. 2021, 225, 522.e1–522.e11. [Google Scholar] [CrossRef]
- Zimmermann, P.; Curtis, N. COVID-19 in Children, Pregnancy and Neonates: A Review of Epidemiologic and Clinical Features. Pediatr. Infect. Dis. J. 2020, 39, 469–477. [Google Scholar] [CrossRef]
- Keller-Stanislawski, B.; Englund, J.A.; Kang, G.; Mangtani, P.; Neuzil, K.; Nohynek, H.; Pless, R.; Lambach, P.; Zuber, P. Safety of immunization during pregnancy: A review of the evidence of selected inactivated and live attenuated vaccines. Vaccine 2014, 32, 7057–7064. [Google Scholar] [CrossRef] [PubMed]
- Albrecht, M.; Arck, P.C. Vertically Transferred Immunity in Neonates: Mothers, Mechanisms and Mediators. Front. Immunol. 2020, 11, 555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, C.J.; Fu, Y.C.; Lin, Y.P.; Shen, C.F.; Sun, D.J.; Chen, H.Y.; Cheng, C.M. Evaluation of Transplacental Antibody Transfer in SARS-CoV-2-Immunized Pregnant Women. Vaccines 2022, 10, 101. [Google Scholar] [CrossRef] [PubMed]
- Pulliam, J.R.C.; van Schalkwyk, C.; Govender, N.; von Gottberg, A.; Cohen, C.; Groome, M.J.; Dushoff, J.; Mlisana, K.; Moultrie, H. Increased risk of SARS-CoV-2 reinfection associated with emergence of the Omicron variant in South Africa. medRxiv 2021. [Google Scholar] [CrossRef]
- Zhao, J.; Yuan, Q.; Wang, H.; Liu, W.; Liao, X.; Su, Y.; Wang, X.; Yuan, J.; Li, T.; Li, J.; et al. Antibody Responses to SARS-CoV-2 in Patients With Novel Coronavirus Disease 2019. Clin. Infect. Dis. 2020, 71, 2027–2034. [Google Scholar] [CrossRef]
- Hung, K.F.; Hung, C.H.; Hong, C.; Chen, S.C.; Sun, Y.C.; Wen, J.W.; Kuo, C.H.; Ko, C.H.; Cheng, C.M. Quantitative Spectrochip-Coupled Lateral Flow Immunoassay Demonstrates Clinical Potential for Overcoming Coronavirus Disease 2019 Pandemic Screening Challenges. Micromachines 2021, 12, 321. [Google Scholar] [CrossRef]
- O’Farrell, B. Evolution in Lateral Flow–Based Immunoassay Systems. In Lateral Flow Immunoassay; Wong, R., Tse, H., Eds.; Humana Press: Totowa, NJ, USA, 2009; pp. 1–33. [Google Scholar]
- Weihs, F.; Anderson, A.; Trowell, S.; Caron, K. Resonance Energy Transfer-Based Biosensors for Point-of-Need Diagnosis—Progress and Perspectives. Sensors 2021, 21, 660. [Google Scholar] [CrossRef]
- Li, J.; Macdonald, J. Multiplexed lateral flow biosensors: Technological advances for radically improving point-of-care diagnoses. Biosens. Bioelectron. 2016, 83, 177–192. [Google Scholar] [CrossRef]
- Deeks, J.J.; Dinnes, J.; Takwoingi, Y.; Davenport, C.; Spijker, R.; Taylor-Phillips, S.; Adriano, A.; Beese, S.; Dretzke, J.; Ferrante di Ruffano, L.; et al. Antibody tests for identification of current and past infection with SARS-CoV-2. Cochrane Database Syst. Rev. 2020, 6, Cd013652. [Google Scholar] [CrossRef]
- Antiochia, R. Paper-Based Biosensors: Frontiers in Point-of-Care Detection of COVID-19 Disease. Biosensors 2021, 11, 110. [Google Scholar] [CrossRef]
- Wang, Y.C.; Lee, Y.T.; Yang, T.; Sun, J.R.; Shen, C.F.; Cheng, C.M. Current diagnostic tools for coronaviruses-From laboratory diagnosis to POC diagnosis for COVID-19. Bioeng. Transl. Med. 2020, 5, e10177. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.W.; Shen, C.F.; Liu, C.C.; Cheng, C.M. A Paper-Based IL-6 Test Strip Coupled With a Spectrum-Based Optical Reader for Differentiating Influenza Severity in Children. Front. Bioeng. Biotechnol. 2021, 9, 752681. [Google Scholar] [CrossRef] [PubMed]
- GenScript USA Inc. cPass SARS-CoV-2 Neutralization Antibody Detection Kit. Available online: https://www.fda.gov/media/143583/download (accessed on 5 February 2022).
- World Health Organization. WHO Coronavirus (COVID-19) Dashboard. Available online: https://covid19.who.int (accessed on 8 April 2022).
- Shang, W.; Yang, Y.; Rao, Y.; Rao, X. The outbreak of SARS-CoV-2 pneumonia calls for viral vaccines. NPJ Vaccines 2020, 5, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hyams, C.; Marlow, R.; Maseko, Z.; King, J.; Ward, L.; Fox, K.; Heath, R.; Tuner, A.; Friedrich, Z.; Morrison, L.; et al. Effectiveness of BNT162b2 and ChAdOx1 nCoV-19 COVID-19 vaccination at preventing hospitalisations in people aged at least 80 years: A test-negative, case-control study. Lancet. Infect. Dis. 2021, 21, 1539–1548. [Google Scholar] [CrossRef]
- Lopez Bernal, J.; Andrews, N.; Gower, C.; Robertson, C.; Stowe, J.; Tessier, E.; Simmons, R.; Cottrell, S.; Roberts, R.; O’Doherty, M.; et al. Effectiveness of the Pfizer-BioNTech and Oxford-AstraZeneca vaccines on covid-19 related symptoms, hospital admissions, and mortality in older adults in England: Test negative case-control study. BMJ 2021, 373, n1088. [Google Scholar] [CrossRef] [PubMed]
- Khoury, D.S.; Cromer, D.; Reynaldi, A.; Schlub, T.E.; Wheatley, A.K.; Juno, J.A.; Subbarao, K.; Kent, S.J.; Triccas, J.A.; Davenport, M.P. Neutralizing antibody levels are highly predictive of immune protection from symptomatic SARS-CoV-2 infection. Nat. Med. 2021, 27, 1205–1211. [Google Scholar] [CrossRef]
- Letko, M.; Marzi, A.; Munster, V. Functional assessment of cell entry and receptor usage for SARS-CoV-2 and other lineage B betacoronaviruses. Nat. Microbiol. 2020, 5, 562–569. [Google Scholar] [CrossRef] [Green Version]
- Makoah, N.A.; Tipih, T.; Litabe, M.M.; Brink, M.; Sempa, J.B.; Goedhals, D.; Burt, F.J. A systematic review and meta-analysis of the sensitivity of antibody tests for the laboratory confirmation of COVID-19. Future Virol. 2021, 17, 119–139. [Google Scholar] [CrossRef]
- Huang, R.L.; Fu, Y.C.; Wang, Y.C.; Hong, C.; Yang, W.C.; Wang, I.J.; Sun, J.R.; Chen, Y.; Shen, C.F.; Cheng, C.M. A Lateral Flow Immunoassay Coupled with a Spectrum-Based Reader for SARS-CoV-2 Neutralizing Antibody Detection. Vaccines 2022, 10, 271. [Google Scholar] [CrossRef]
- Beharier, O.; Plitman Mayo, R.; Raz, T.; Nahum Sacks, K.; Schreiber, L.; Suissa-Cohen, Y.; Chen, R.; Gomez-Tolub, R.; Hadar, E.; Gabbay-Benziv, R.; et al. Efficient maternal to neonatal transfer of antibodies against SARS-CoV-2 and BNT162b2 mRNA COVID-19 vaccine. J. Clin. Investig. 2021, 131, e150319. [Google Scholar] [CrossRef]
- Matsui, Y.; Li, L.; Prahl, M.; Cassidy, A.G.; Ozarslan, N.; Golan, Y.; Gonzalez, V.J.; Lin, C.Y.; Jigmeddagva, U.; Chidboy, M.A.; et al. Neutralizing Antibody Activity Against SARS-CoV-2 Variants in Gestational Age-Matched Mother-Infant Dyads. medRxiv 2021. [Google Scholar] [CrossRef]
- Gray, K.J.; Bordt, E.A.; Atyeo, C.; Deriso, E.; Akinwunmi, B.; Young, N.; Baez, A.M.; Shook, L.L.; Cvrk, D.; James, K.; et al. COVID-19 vaccine response in pregnant and lactating women: A cohort study. medRxiv 2021. [Google Scholar] [CrossRef]
- Moustsen-Helms, I.R.; Emborg, H.-D.; Nielsen, J.; Nielsen, K.F.; Krause, T.G.; Mølbak, K.; Møller, K.L.; Berthelsen, A.-S.N.; Valentiner-Branth, P. Vaccine effectiveness after 1st and 2nd dose of the BNT162b2 mRNA Covid-19 Vaccine in long-term care facility residents and healthcare workers—A Danish cohort study. medRxiv 2021. [Google Scholar] [CrossRef]
- Thomas, S.J.; Moreira, E.D., Jr.; Kitchin, N.; Absalon, J.; Gurtman, A.; Lockhart, S.; Perez, J.L.; Pérez Marc, G.; Polack, F.P.; Zerbini, C.; et al. Safety and Efficacy of the BNT162b2 mRNA Covid-19 Vaccine through 6 Months. N. Engl. J. Med. 2021, 385, 1761–1773. [Google Scholar] [CrossRef] [PubMed]
- Lipsitch, M. Challenges of Vaccine Effectiveness and Waning Studies. Clin. Infect. Dis. 2019, 68, 1631–1633. [Google Scholar] [CrossRef] [PubMed]
- Wilhelm, A.; Widera, M.; Grikscheit, K.; Toptan, T.; Schenk, B.; Pallas, C.; Metzler, M.; Kohmer, N.; Hoehl, S.; Helfritz, F.A.; et al. Reduced Neutralization of SARS-CoV-2 Omicron Variant by Vaccine Sera and Monoclonal Antibodies. medRxiv 2021. [Google Scholar] [CrossRef]
- Andrews, N.; Stowe, J.; Kirsebom, F.; Toffa, S.; Sachdeva, R.; Gower, C.; Ramsay, M.; Bernal, J.L. Effectiveness of COVID-19 booster vaccines against covid-19 related symptoms, hospitalisation and death in England. Nat. Med. 2022, 28, 831–837. [Google Scholar] [CrossRef]
- Thompson, M.G.; Natarajan, K.; Irving, S.A.; Rowley, E.A.; Griggs, E.P.; Gaglani, M.; Klein, N.P.; Grannis, S.J.; DeSilva, M.B.; Stenehjem, E.; et al. Effectiveness of a Third Dose of mRNA Vaccines Against COVID-19-Associated Emergency Department and Urgent Care Encounters and Hospitalizations Among Adults During Periods of Delta and Omicron Variant Predominance-VISION Network, 10 States, August 2021–January 2022. MMWR Morb. Mortal. Wkly. Rep. 2022, 71, 139–145. [Google Scholar] [CrossRef]
- Abu-Raddad, L.J.; Chemaitelly, H.; Ayoub, H.H.; Yassine, H.M.; Benslimane, F.M.; Al Khatib, H.A.; Tang, P.; Hasan, M.R.; Coyle, P.; Al Kanaani, Z.; et al. Association of Prior SARS-CoV-2 Infection With Risk of Breakthrough Infection Following mRNA Vaccination in Qatar. JAMA 2021, 326, 1930–1939. [Google Scholar] [CrossRef]
- Mizrahi, B.; Lotan, R.; Kalkstein, N.; Peretz, A.; Perez, G.; Ben-Tov, A.; Chodick, G.; Gazit, S.; Patalon, T. Correlation of SARS-CoV-2-breakthrough infections to time-from-vaccine. Nat. Commun. 2021, 12, 6379. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | One-Dose Group (Median, IQR) | Two-Dose Group (Median, IQR) (Interval of Second Administration to Delivery) | 0–2 Weeks | 2–4 Weeks | 4–8 Weeks |
|---|---|---|---|---|---|
| Maternal SRBD IgG by Strip and spectrum analyzer | 0 ng/mL ** (0–12.87) | 143.27 ng/mL *** (110.61–200.35) | 146.91 ng/mL 1 (120.19–173.63) | 127.99 ng/mL 2 (102.28–184.02) | 157.14 ng/mL 3 (127.55–267.39) |
| Cord blood SRBD IgG by Strip and spectrum analyzer | 0 ng/mL ** (0–13.92) | 178.40 ng/mL *** (131.62–236.61) | 207.58 ng/mL 1 (180.85–234.31) | 146.75 ng/mL 2 (114.19–195.52) | 188.03 ng/mL 3 (131.62–330.32) |
| Maternal neutralizing antibody (Wild-type) | 40.32% ** (25.28–51.74) | 97.45% *** (96.82–97.61) | 95.99% 1 (95.58–96.41) | 97.39% 2 (97.26–97.54) | 97.48% 3 (97.35–97.74) |
| Cord blood neutralizing antibody (Wild-type) | 43.33% ** (28.80–43.74) | 97.11% *** (96.24–97.56) | 95.78% 1 (95.18–96.38) | 97.10% 2 (96.31–97.28) | 97.56% 3 (97.03–97.59) |
| Cord to maternal ratio (Wild-type) | 1.07 ** (0.89–1.23) | 0.999 *** (0.996–1.00) | 0.99 1 (0.99–1.01) | 1.00 2 (0.99–1.00) | 1.00 3 (1.00–1.00) |
| Maternal neutralizing antibody (Delta-type) | 4.01% ** (0.87–7.51) | 77.82% **** (52.13–83.63) | 49.96% 1 (48.87–51.06) | 79.27% 2 (62.42–82.90) | 80.24% 3 (59.98–86.71) |
| Cord blood neutralizing antibody (Delta-type) | 1.44% ** (2.16) | 64.21% **** (41.14–70.38) | 41.86% 1 (30.46–53.26) | 57.48% 2 (47.82–63.47) | 68.79% 3 (41.14–73.56) |
| Cord to maternal ratio (Delta-type) | 0.92 ** (0.86–1.40) | 0.87 **** (0.77–0.94) | 0.86 1 (0.61–1.11) | 0.78 2 (0.75–0.85) | 0.92 3 (0.85–0.94) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, W.-C.; Lin, Y.-P.; Cheng, C.-M.; Shen, C.-F.; Li, C.-W.; Wang, Y.-K.; Shih, T.-Y.; Hong, C.; Chang, T.-C.; Shen, C.-J. Detection of SARS-CoV-2 Neutralizing Antibodies in Vaccinated Pregnant Women and Neonates by Using a Lateral Flow Immunoassay Coupled with a Spectrum-Based Reader. Biosensors 2022, 12, 891. https://doi.org/10.3390/bios12100891
Chen W-C, Lin Y-P, Cheng C-M, Shen C-F, Li C-W, Wang Y-K, Shih T-Y, Hong C, Chang T-C, Shen C-J. Detection of SARS-CoV-2 Neutralizing Antibodies in Vaccinated Pregnant Women and Neonates by Using a Lateral Flow Immunoassay Coupled with a Spectrum-Based Reader. Biosensors. 2022; 12(10):891. https://doi.org/10.3390/bios12100891
Chicago/Turabian StyleChen, Wei-Chun, Yen-Pin Lin, Chao-Min Cheng, Ching-Fen Shen, Chang-Wei Li, Yu-Kuo Wang, Ting-Ying Shih, Chitsung Hong, Ting-Chang Chang, and Ching-Ju Shen. 2022. "Detection of SARS-CoV-2 Neutralizing Antibodies in Vaccinated Pregnant Women and Neonates by Using a Lateral Flow Immunoassay Coupled with a Spectrum-Based Reader" Biosensors 12, no. 10: 891. https://doi.org/10.3390/bios12100891
APA StyleChen, W.-C., Lin, Y.-P., Cheng, C.-M., Shen, C.-F., Li, C.-W., Wang, Y.-K., Shih, T.-Y., Hong, C., Chang, T.-C., & Shen, C.-J. (2022). Detection of SARS-CoV-2 Neutralizing Antibodies in Vaccinated Pregnant Women and Neonates by Using a Lateral Flow Immunoassay Coupled with a Spectrum-Based Reader. Biosensors, 12(10), 891. https://doi.org/10.3390/bios12100891