Review of Label-Free Monitoring of Bacteria: From Challenging Practical Applications to Basic Research Perspectives

 , , and
, , and
Abstract
:1. Introduction
2. Applications of Biosensors in Bacteria Detection
2.1. Recognition of Bacteria Using Biosensors
2.1.1. Optical Biosensors
2.1.2. Mechanical Biosensors
2.1.3. Electrochemical Biosensors
2.2. Resistance to Antibacterials and to Other Antimicrobial Agents
- -
- occupation and blockage of the active site of a lectin that mediates the adhesion of bacterial glycocalyx fibers to the fibers of host cells,
- -
- blockage of the “receptor” sites on host cells, the glycoprotein fibers to which bacterial fibers adhere directly, or
- -
- disruption of the production of glycocalyx fibers (mimics its normal substrate and therefore occupies the enzyme’s active site) [18].
2.3. Detection of Resistance Using Biosensors
Bacteriophage in Biosensors
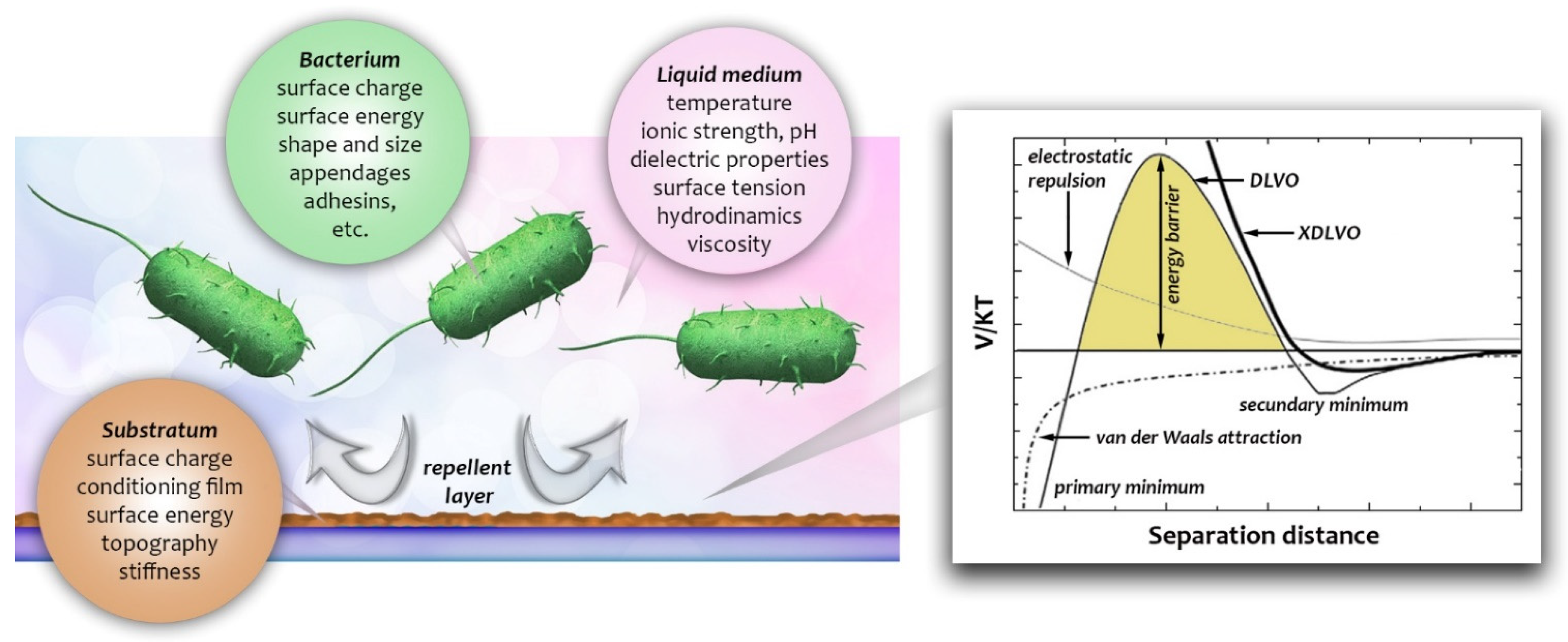
2.4. Strategies to Reduce and Control Bacterial Adhesion on Sensing Surfaces
2.5. Biosensor Applications to Elucidate Molecular Modes of Action—Research Challenges
3. Biological Mechanisms in Bacterial Adhesion as Potential Subjects of Label-Free Monitoring
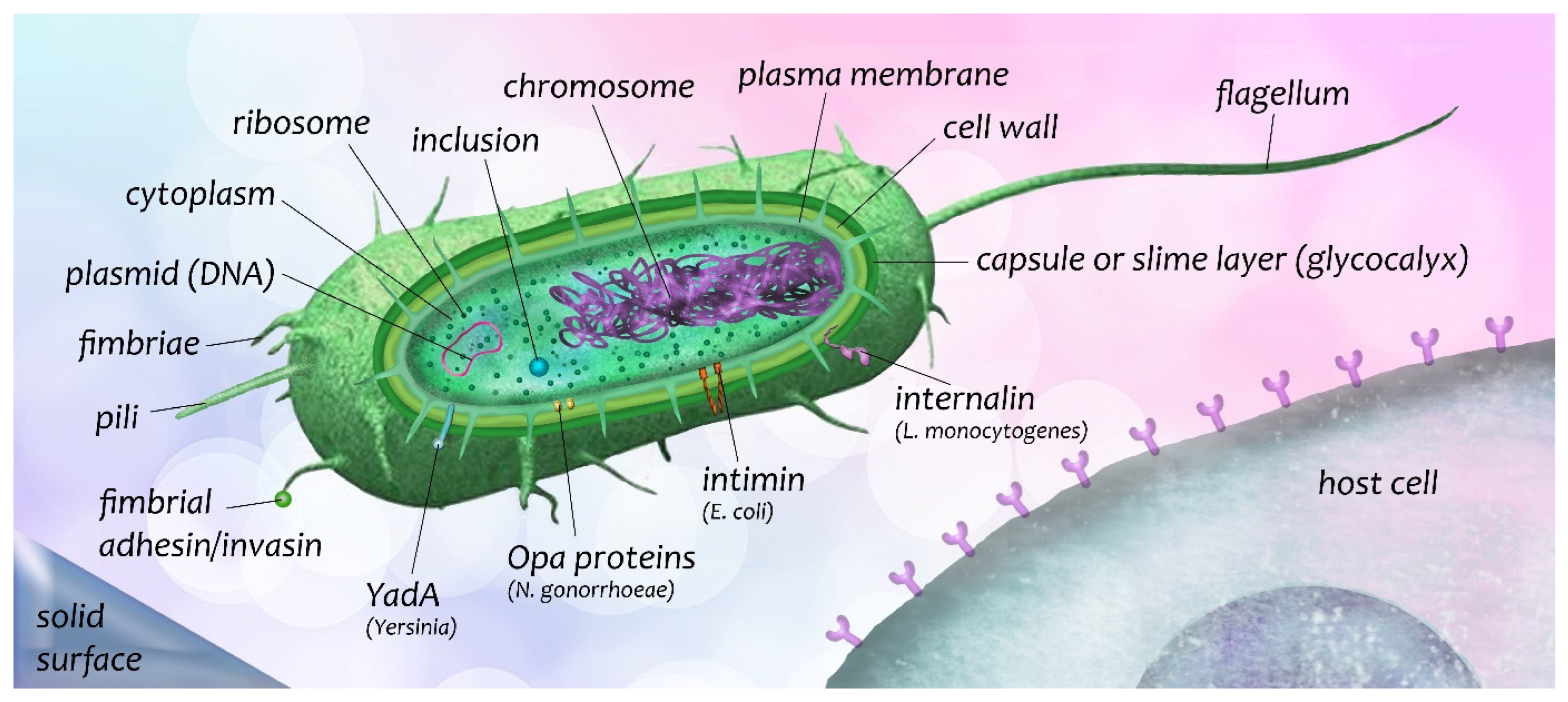
3.1. Adhesive Molecules of Bacteria and Bacterial Entry into Host Cells
3.1.1. Type III Secretion-Associated Invasion Mechanisms
3.1.2. Receptors on the Cell Surface
3.1.3. Adherence to ECM Proteins
3.1.4. Specific Cell Receptors Involved in Cell–Cell Interaction
3.1.5. Bacterial Entry into Host Cells
3.2. Role of Bacteria’s Glycocalyx in Their Adhesion
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kovacs, B.; Patko, D.; Klein, A.; Kakasi, B.; Saftics, A.; Kurunczi, S.; Vonderviszt, F.; Horvath, R. Bacteria repellent layer made of flagellin. Sens. Actuators B Chem. 2018, 257, 839–845. [Google Scholar] [CrossRef]
- Ramsden, J.J. Review of new experimental techniques for investigating random sequential adsorption. J. Stat. Phys. 1993, 73, 853–877. [Google Scholar] [CrossRef]
- Ramsden, J.J.; Máté, M. Kinetics of monolayer particle deposition. J. Chem. Soc. Faraday Trans. 1998, 94, 783–788. [Google Scholar] [CrossRef]
- Ramsden, J.J. Calcium-dependence of laminin binding to phospholipid membranes. Biopolymers 1993, 33, 475–477. [Google Scholar] [CrossRef]
- Guemouri, L.; Ogier, J.; Ramsden, J.J. Optical properties of protein monolayers during assembly. J. Chem. Phys. 1998, 109, 3265–3268. [Google Scholar] [CrossRef]
- Guemouri, L.; Ogier, J.; Zekhnini, Z.; Ramsden, J.J. The architecture of fibronectin at surfaces. J. Chem. Phys. 2000, 113, 8183–8186. [Google Scholar] [CrossRef]
- Horvath, R.; Ramsden, J.J. Quasi-isotropic analysis of anisotropic thin films on optical waveguides. Langmuir 2007, 23, 9330–9334. [Google Scholar] [CrossRef]
- Ramsden, J.J.; Li, S.-Y.; Prenosil, J.E.; Heinzle, E. Kinetics of adhesion and spreading of animal cells. Biotechnol. Bioeng. 1994, 43, 939–945. [Google Scholar] [CrossRef]
- Li, S.-Y.; Ramsden, J.J.; Prenosil, J.E.; Heinzle, E. Measurement of Adhesion and Spreading Kinetics of Baby Hamster Kidney and Hybridoma Cells Using an Integrated Optical Method. Biotechnol. Prog. 1994, 10, 520–524. [Google Scholar] [CrossRef]
- Horvath, R.; Cottier, K.; Pedersen, H.C.; Ramsden, J.J. Multidepth screening of living cells using optical waveguides. Biosens. Bioelectron. 2008, 24, 799–804. [Google Scholar] [CrossRef]
- Bai, W.; Zhao, K.S.; Asami, K. Dielectric properties of E. coli cell as simulated by the three-shell spheroidal model. Biophys. Chem. 2006, 122, 136–142. [Google Scholar] [CrossRef] [PubMed]
- Hölzel, R. Non-invasive determination of bacterial single cell properties by electrorotation. Biochim. Biophys. Acta Mol. Cell Res. 1999, 1450, 53–60. [Google Scholar] [CrossRef] [Green Version]
- Alderwick, L.J.; Harrison, J.; Lloyd, G.S.; Birch, H.L. The mycobacterial cell wall—Peptidoglycan and arabinogalactan. Cold Spring Harb. Perspect. Med. 2015, 5, a021113. [Google Scholar] [CrossRef] [Green Version]
- Maitra, A.; Munshi, T.; Healy, J.; Martin, L.T.; Vollmer, W.; Keep, N.H.; Bhakta, S. Cell wall peptidoglycan in Mycobacterium tuberculosis: An Achilles’ heel for the TB-causing pathogen. FEMS Microbiol. Rev. 2019, 43, 548–575. [Google Scholar] [CrossRef] [Green Version]
- Silva, M.T. Classical labeling of bacterial pathogens according to their lifestyle in the host: Inconsistencies and alternatives. Front. Microbiol. 2012, 3, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Hori, K.; Matsumoto, S. Bacterial adhesion: From mechanism to control. Biochem. Eng. J. 2010, 48, 424–434. [Google Scholar] [CrossRef]
- Dersch, P. Molecular and cellular mechanisms of bacterial entry into host cells. Contrib. Microbiol. 2003, 10, 183–209. [Google Scholar]
- Costerton, J.W.; Geesey, G.G.; Cheng, K.J. How bacteria stick. Sci. Am. 1978, 238, 86–95. [Google Scholar] [CrossRef]
- Costerton, J.W.; Irvin, R.T.; Cheng, K.J. The bacterial glycocalyx in nature and disease. Annu. Rev. Microbiol. 1981, 35, 299–324. [Google Scholar] [CrossRef]
- Ramsden, J.J. Photocatalytic antimicrobial coatings. Nanotechnol. Percept. 2015, 11, 146–168. [Google Scholar] [CrossRef]
- Reder-Christ, K.; Bendas, G. Biosensor applications in the field of antibiotic research—A review of recent developments. Sensors 2011, 11, 9450–9466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramsden, J.J. Can bacteria develop resistance to photocatalytically generated reactive oxygen species? J. Biol. Phys. Chem. 2017, 17, 47–51. [Google Scholar] [CrossRef]
- Ahmed, A.; Rushworth, J.V.; Hirst, N.A.; Millner, P.A. Biosensors for whole-cell bacterial detection. Clin. Microbiol. Rev. 2014, 27, 631–646. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scerbo, M.H.; Kaplan, H.B.; Dua, A.; Litwin, D.B.; Ambrose, C.G.; Moore, L.J.; Murray, C.C.K.; Wade, C.E.; Holcomb, J.B. Beyond Blood Culture and Gram Stain Analysis: A Review of Molecular Techniques for the Early Detection of Bacteremia in Surgical Patients. Surg. Infect. 2016, 17, 294–302. [Google Scholar] [CrossRef]
- Amjad, M. An Overview of the Molecular Methods in the Diagnosis of Gastrointestinal Infectious Diseases. Int. J. Microbiol. 2020, 2020, 8135724. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.F.; Chen, Y.C.; Wang, W.C.; Fang, Y.C.; Fukuoka, S.; Pride, D.T.; Pak, O.S. A Rapid and Low-Cost Pathogen Detection Platform by Using a Molecular Agglutination Assay. ACS Cent. Sci. 2018, 4, 1485–1494. [Google Scholar] [CrossRef] [Green Version]
- Elder, B.L.; Boraker, D.K.; Fives-Taylor, P.M. Whole-bacterial cell enzyme-linked immunosorbent assay for Streptococcus sanguis fimbrial antigens. J. Clin. Microbiol. 1982, 16, 141–144. [Google Scholar] [CrossRef] [Green Version]
- Burnham, C.D.; Carroll, C. Diagnosis of Clostridium difficile Infection: An Ongoing Conundrum for Clinicians and for Clinical Laboratories. ASM J. 2013, 26, 604–630. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, Y. PCR in diagnosis of infection: Detection of bacteria in cerebrospinal fluids. Clin. Diagn. Lab. Immunol. 2002, 9, 508–514. [Google Scholar] [CrossRef] [Green Version]
- Espy, M.J.; Uhl, J.R.; Sloan, L.M.; Buckwalter, S.P.; Jones, M.F.; Vetter, E.A.; Yao, J.D.C.; Wengenack, N.L.; Rosenblatt, J.E.; Iii, F.R.C.; et al. Real-Time PCR in Clinical Microbiology: Applications for Routine Laboratory Testing. Clin Microbiol Rev. 2006, 19, 165–256. [Google Scholar] [CrossRef] [Green Version]
- Sauer, S.; Kliem, M. Mass spectrometry tools for the classification and identification of bacteria. Nat. Rev. Microbiol. 2010, 8, 74–82. [Google Scholar] [CrossRef] [PubMed]
- Gfeller, K.Y.; Nugaeva, N.; Hegner, M. Rapid biosensor for detection of antibiotic-selective growth of Escherichia coli. Appl. Environ. Microbiol. 2005, 71, 2626–2631. [Google Scholar] [CrossRef] [Green Version]
- Obaje, E.A.; Cummins, G.; Schulze, H.; Mahmood, S.; Desmulliez, M.P.Y.; Bachmann, T.T. Carbon screen-printed electrodes on ceramic substrates for label-free molecular detection of antibiotic resistance. J. Interdiscip. Nanomed. 2016, 1, 93–109. [Google Scholar] [CrossRef]
- Székács, I.; Horváth, R.; Székács, A. Label-free optical biosensors for monitoring cellular processes and cytotoxic agents at interfaces using guided modes and advanced phase-contrast imaging techniques. In Biosensors for Security and Bioterrorism Applications, Advanced Sciences and Technologies for Security Applications; Nikolelis, D.P., Nikoleli, G.-P., Eds.; Springer: Cham, Switzerland, 2016. [Google Scholar] [CrossRef]
- Suaifan, G.A.R.Y.; Alhogail, S.; Zourob, M. Rapid and low-cost biosensor for the detection of Staphylococcus aureus. Biosens. Bioelectron. 2017, 90, 230–237. [Google Scholar] [CrossRef] [PubMed]
- Zourob, M.; Elwary, S.; Turner, A.P.F. Principles of Bacterial Detection: Biosensors, Recognition Receptors and Microsystems; Springer: New York, NY, USA, 2008. [Google Scholar]
- Paniel, N.; Baudart, J. Talanta Colorimetric and electrochemical genosensors for the detection of Escherichia coli DNA without ampli fi cation in seawater. Talanta 2013, 115, 133–142. [Google Scholar] [CrossRef]
- Anderson, M.J.; Miller, H.R.; Alocilja, E.C. PCR-less DNA co-polymerization detection of Shiga like toxin 1 (stx 1) in Escherichia coli O157: H7. Biosens. Bioelectron. 2013, 42, 581–585. [Google Scholar] [CrossRef]
- Foudeh, A.M.; Daoud, J.T.; Faucher, S.P.; Veres, T.; Tabrizian, M. Sub-femtomole detection of 16s rRNA from Legionella pneumophila using surface plasmon resonance imaging. Biosens. Bioelectron. 2014, 52, 129–135. [Google Scholar] [CrossRef]
- Gerasimova, Y.V.; Kolpashchikov, D.M. Detection of bacterial 16S rRNA using a molecular beacon-based X sensor. Biosens. Bioelectron. 2013, 41, 386–390. [Google Scholar] [CrossRef] [Green Version]
- Dastider, S.G.; Abdullah, A.; Jasim, I.; Yuksek, N.S.; Dweik, M.; Almasri, M. Low concentration E. coli O157:H7 bacteria sensing using microfluidic MEMS biosensor. Rev. Sci. Instrum. 2018, 89, 125009. [Google Scholar] [CrossRef]
- Kwizera, E.A.; Sun, M.; White, A.M.; Li, J.; He, X. Methods of generating dielectrophoretic force for microfluidic manipulation of bioparticles. ACS Biomater. Sci. Eng. 2021, 7, 2043–2063. [Google Scholar] [CrossRef]
- Zhang, H.; Chang, H.; Neuzil, P. DEP-on-a-chip: Dielectrophoresis applied to microfluidic platforms. Micromachines 2019, 10, 423. [Google Scholar] [CrossRef] [Green Version]
- Çetin, B.; Li, D. Dielectrophoresis in microfluidics technology. Electrophoresis 2011, 32, 2410–2427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, S.; Byrne, H.; O’Kennedy, R.J. Antibodies and antibody-derived analytical biosensors. Essays Biochem. 2016, 60, 9–18. [Google Scholar] [CrossRef] [PubMed]
- Belický, Š.; Katrlík, J.; Tkáč, J. Glycan and lectin biosensors. Essays Biochem. 2016, 60, 37–47. [Google Scholar] [CrossRef]
- Sorokulova, I.; Olsen, E.; Vodyanoy, V. Bacteriophage biosensors for antibiotic-resistant bacteria. Expert Rev. Med. Devices 2014, 11, 175–186. [Google Scholar] [CrossRef]
- Ramsden, J.J. Optical biosensors. J. Mol. Recogn. 1997, 10, 109–120. [Google Scholar] [CrossRef]
- Yeh, Y.; Ramsden, J.J. Quantification of the number of adsorbed bacteria on an optical waveguide. J. Biol. Phys. Chem. 2010, 10, 53–54. [Google Scholar] [CrossRef]
- Zhou, Z.; Zhang, Y.; Guo, M.; Huang, K.; Xu, W. Ultrasensitive magnetic DNAzyme-copper nanoclusters fluorescent biosensor with triple amplification for the visual detection of E. coli O157:H7. Biosens. Bioelectron. 2020, 167, 112475. [Google Scholar] [CrossRef]
- Yu, T.; Xu, H.; Zhao, Y.; Han, Y.; Zhang, Y.; Zhang, J.; Xu, C.; Wang, W.; Guo, Q.; Ge, J. Aptamer based high throughput colorimetric biosensor for detection of Staphylococcus aureus. Sci. Rep. 2020, 10, 9190. [Google Scholar] [CrossRef]
- Xue, L.; Zheng, L.; Zhang, H.; Jin, X.; Lin, J. An ultrasensitive fluorescent biosensor using high gradient magnetic separation and quantum dots for fast detection of foodborne pathogenic bacteria. Sens. Actuators B Chem. 2018, 265, 318–325. [Google Scholar] [CrossRef]
- Xu, L.; Lu, Z.; Cao, L.; Pang, H.; Zhang, Q.; Fu, Y.; Xiong, Y.; Li, Y.; Wang, X.; Wang, J.; et al. In-field detection of multiple pathogenic bacteria in food products using a portable fluorescent biosensing system. Food Control 2017, 75, 21–28. [Google Scholar] [CrossRef]
- Song, M.S.; Sekhon, S.S.; Shin, W.R.; Kim, H.C.; Min, J.; Ahn, J.Y.; Kim, Y.H. Detecting and discriminating shigella sonnei using an aptamer-based fluorescent biosensor platform. Molecules 2017, 22, 825. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, M.; Zhang, Q.; Brennan, J.D.; Li, Y. Graphene-DNAzyme-based fluorescent biosensor for Escherichia coli detection. MRS Commun. 2018, 8, 687–694. [Google Scholar] [CrossRef] [Green Version]
- Jeong, W.J.; Choi, S.H.; Lee, H.S.; Lim, Y. beom A fluorescent supramolecular biosensor for bacterial detection via binding-induced changes in coiled-coil molecular assembly. Sens. Actuators B Chem. 2019, 290, 93–99. [Google Scholar] [CrossRef]
- Mouffouk, F.; Rosa, A.M.; Martins, J.; Zourob, M.; Abu-salah, K.M.; Alrokayan, S.A. Development of a highly sensitive bacteria detection assay using fluorescent pH-responsive polymeric micelles. Biosens. Bioelectron. 2011, 26, 3517–3523. [Google Scholar] [CrossRef]
- Raji, M.A.; Chinnappan, R.; Shibl, A.; Suaifan, G.; Weber, K.; Cialla-May, D.; Popp, J.; El Shorbagy, E.; Al-Kattan, K.; Zourob, M. Low-cost colorimetric diagnostic screening assay for methicillin resistant Staphylococcus aureus. Talanta 2021, 225, 121946. [Google Scholar] [CrossRef]
- Alhogail, S.; Suaifan, G.A.R.Y.; Bikker, F.J.; Kaman, W.E.; Weber, K.; Cialla-May, D.; Popp, J.; Zourob, M.M. Rapid colorimetric detection of Pseudomonas aeruginosa in clinical isolates using a magnetic nanoparticle biosensor. ACS Omega 2019, 4, 21684–21688. [Google Scholar] [CrossRef] [Green Version]
- Suaifan, G.A.R.Y.; Alhogail, S.; Zourob, M. Paper-based magnetic nanoparticle-peptide probe for rapid and quantitative colorimetric detection of Escherichia coli O157:H7. Biosens. Bioelectron. 2017, 92, 702–708. [Google Scholar] [CrossRef]
- Alhogail, S.; Suaifan, G.A.R.Y.; Zourob, M. Rapid colorimetric sensing platform for the detection of Listeria monocytogenes foodborne pathogen. Biosens. Bioelectron. 2016, 86, 1061–1066. [Google Scholar] [CrossRef]
- Alamer, S.; Chinnappan, R.; Zourob, M. Development of rapid iImmuno-based nanosensors for the detection of pathogenic bacteria in poultry processing plants. Procedia Technol. 2017, 27, 23–26. [Google Scholar] [CrossRef]
- Peter, B.; Nador, J.; Juhasz, K.; Dobos, A.; Korosi, L.; Székács, I.; Patko, D.; Horvath, R. Incubator proof miniaturized Holomonitor to in situ monitor cancer cells exposed to green tea polyphenol and preosteoblast cells adhering on nanostructured titanate surfaces: Validity of the measured parameters and their corrections. J. Biomed. Opt. 2015, 20, 067002. [Google Scholar] [CrossRef] [PubMed]
- Homola, J. Surface plasmon resonance sensors for detection of chemical and biological species. Chem. Rev. 2008, 108, 462–493. [Google Scholar] [CrossRef] [PubMed]
- Ramsden, J.J. Experimental methods for investigating protein adsorption kinetics at surfaces. Q. Rev. Biophys. 1993, 27, 41–105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raghu, H.V.; Kumar, N. Rapid detection of Listeria monocytogenes in milk by surface plasmon resonance using wheat germ agglutinin. Food Anal. Methods 2020, 13, 982–991. [Google Scholar] [CrossRef]
- Saylan, Y.; Erdem, Ö.; Cihangir, N.; Denizli, A. Detecting fingerprints of waterborne bacteria on a sensor. Chemosensors 2019, 7, 33. [Google Scholar] [CrossRef] [Green Version]
- Zhou, C.; Zou, H.; Li, M.; Sun, C.; Ren, D.; Li, Y. Fiber optic surface plasmon resonance sensor for detection of E. coli O157:H7 based on antimicrobial peptides and AgNPs-rGO. Biosens. Bioelectron. 2018, 117, 347–353. [Google Scholar] [CrossRef]
- Padra, J.T.; Pagneux, Q.; Bouckaert, J.; Jijie, R.; Sundh, H.; Boukherroub, R.; Szunerits, S.; Lindén, S.K. Mucin modified SPR interfaces for studying the effect of flow on pathogen binding to Atlantic salmon mucins. Biosens. Bioelectron. 2019, 146, 111736. [Google Scholar] [CrossRef] [Green Version]
- Bhandari, D.; Chen, F.C.; Bridgman, R.C. Detection of Salmonella Typhimurium in romaine lettuce using a surface plasmon resonance biosensor. Biosensors 2019, 9, 94. [Google Scholar] [CrossRef] [Green Version]
- Taheri, R.A.; Rezayan, A.H.; Rahimi, F.; Mohammadnejad, J.; Kamali, M. Evaluating the potential of an antibody against recombinant OmpW antigen in detection of Vibrio cholerae by surface plasmon resonance (SPR) biosensor. Plasmonics 2017, 12, 1493–1504. [Google Scholar] [CrossRef]
- Zhang, X.; Tsuji, S.; Kitaoka, H.; Kobayashi, H.; Tamai, M.; Honjoh, K.I.; Miyamoto, T. Simultaneous detection of Escherichia coli O157:H7, Salmonella enteritidis, and Listeria monocytogenes at a very low level using simultaneous enrichment broth and multichannel SPR biosensor. J. Food Sci. 2017, 82, 2357–2363. [Google Scholar] [CrossRef]
- Makhneva, E.; Farka, Z.; Skládal, P.; Zajíčková, L. Cyclopropylamine plasma polymer surfaces for label-free SPR and QCM immunosensing of Salmonella. Sens. Actuators B Chem. 2018, 276, 447–455. [Google Scholar] [CrossRef]
- Trzaskowski, M.; Napiórkowska, A.; Augustynowicz-Kopeć, E.; Ciach, T. Detection of tuberculosis in patients with the use of portable SPR device. Sens. Actuators B Chem. 2018, 260, 786–792. [Google Scholar] [CrossRef]
- Vaisocherová-Lísalová, H.; Víšová, I.; Ermini, M.L.; Špringer, T.; Song, X.C.; Mrázek, J.; Lamačová, J.; Scott Lynn, N.; Šedivák, P.; Homola, J. Low-fouling surface plasmon resonance biosensor for multi-step detection of foodborne bacterial pathogens in complex food samples. Biosens. Bioelectron. 2016, 80, 84–90. [Google Scholar] [CrossRef]
- Hyeon, S.H.; Lim, W.K.; Shin, H.J. Novel surface plasmon resonance biosensor that uses full-length Det7 phage tail protein for rapid and selective detection of Salmonella enterica serovar Typhimurium. Biotechnol. Appl. Biochem. 2021, 68, 5–12. [Google Scholar] [CrossRef] [PubMed]
- Shin, H.J.; Lim, W.K. Rapid label-free detection of E. coli using a novel SPR biosensor containing a fragment of tail protein from phage lambda. Prep. Biochem. Biotechnol. 2018, 48, 498–505. [Google Scholar] [CrossRef] [PubMed]
- Torun, Ö.; Boyacı, I.H.; Temür, E.; Tamer, U. Comparison of sensing strategies in SPR biosensor for rapid and sensitive enumeration of bacteria. Biosens. Bioelectron. 2012, 37, 53–60. [Google Scholar] [CrossRef] [PubMed]
- Maurya, J.B.; Prajapati, Y.K.; Tripathi, R. Effect of molybdenum disulfide layer on surface plasmon resonance biosensor for the detection of bacteria. Silicon 2018, 10, 245–256. [Google Scholar] [CrossRef]
- Mudgal, N.; Yupapin, P.; Ali, J.; Singh, G. BaTiO3-Graphene-affinity layer–based surface plasmon resonance (SPR) biosensor for Pseudomonas bacterial detection. Plasmonics 2020, 15, 1221–1229. [Google Scholar] [CrossRef]
- Xu, Y.; Luo, Z.; Chen, J.; Huang, Z.; Wang, X.; An, H.; Duan, Y. ω-Shaped fiber-optic probe-based localized surface plasmon resonance biosensor for real-time detection of Salmonella typhimurium. Anal. Chem. 2018, 90, 13640–13646. [Google Scholar] [CrossRef]
- Kaushal, S.; Priyadarshi, N.; Pinnaka, A.K.; Soni, S.; Deep, A.; Singhal, N.K. Glycoconjugates coated gold nanorods based novel biosensor for optical detection and photothermal ablation of food borne bacteria. Sens. Actuators B Chem. 2019, 289, 207–215. [Google Scholar] [CrossRef]
- Ray, P.C.; Khan, S.A.; Singh, A.K.; Senapati, D.; Ray, P.C. Nanomaterials for targeted detection and photothermal killing of bacteria. Chem. Soc. Rev. 2012, 3193–3209. [Google Scholar] [CrossRef] [PubMed]
- Fu, J.; Park, B.; Zhao, Y. Limitation of a localized surface plasmon resonance sensor for Salmonella detection. Sens. Actuators B Chem. 2009, 141, 276–283. [Google Scholar] [CrossRef]
- Liu, T.Y.; Tsai, K.T.; Wang, H.H.; Chen, Y.; Chen, Y.H.; Chao, Y.C.; Chang, H.H.; Lin, C.H.; Wang, J.K.; Wang, Y.L. Functionalized arrays of Raman-enhancing nanoparticles for capture and culture-free analysis of bacteria in human blood. Nat. Commun. 2011, 2, 538. [Google Scholar] [CrossRef] [PubMed]
- Zhou, S.; Lu, C.; Li, Y.; Xue, L.; Zhao, C.; Tian, G.; Bao, Y.; Tang, L.; Lin, J.; Zheng, J. Gold nanobones enhanced ultrasensitive surface-enhanced Raman scattering aptasensor for detecting Escherichia coli O157:H7. ACS Sens. 2020, 5, 588–596. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Xiao, M.; Lai, W.; Alam, M.F.; Zhang, W.; Pei, H.; Wan, Y.; Li, L. A self-calibrating surface-enhanced Raman scattering-active system for bacterial phenotype detection. Anal. Chem. 2020, 92, 4491–4497. [Google Scholar] [CrossRef]
- Lee, T.; Lim, J.; Park, K.; Lim, E.K.; Lee, J.J. Peptidoglycan-binding protein metamaterials mediated enhanced and selective capturing of Gram-positive bacteria and their specific, ultra-sensitive, and reproducible detection via surface-enhanced Raman scattering. ACS Sens. 2020, 5, 3099–3108. [Google Scholar] [CrossRef]
- Adányi, N.; Bori, Z.; Szendro, I.; Erdélyi, K.; Wang, X.; Schröder, H.C.; Müller, W.E.G. Biosilica-based immobilization strategy for label-free OWLS sensors. Sens. Actuators B Chem. 2013, 177, 1–7. [Google Scholar] [CrossRef]
- Horváth, R.; Pedersen, H.C.; Skivesen, N.; Selmeczi, D.; Larsen, N.B. Optical waveguide sensor for on-line monitoring of bacteria. Opt. Lett. 2003, 28, 1233. [Google Scholar] [CrossRef] [Green Version]
- Horváth, R.; Lindvold, L.R.; Larsen, N.B. Reverse-symmetry waveguides: Theory and fabrication. Appl. Phys. B Lasers Opt. 2002, 74, 383–393. [Google Scholar] [CrossRef]
- Horvath, R.; Pedersen, H.C.; Skivesen, N.; Selmeczi, D.; Larsen, N.B. Monitoring of living cell attachment and spreading using reverse symmetry waveguide sensing. Appl. Phys. Lett. 2005, 86, 1–3. [Google Scholar] [CrossRef] [Green Version]
- Skivesen, N.; Horvath, R.; Pedersen, H.C. Optimization of metal-clad waveguide sensors. Sens. Actuators B Chem. 2005, 106, 668–676. [Google Scholar] [CrossRef]
- Skivesen, N.; Horvath, R.; Thinggaard, S.; Larsen, N.B.; Pedersen, H.C. Deep-probe metal-clad waveguide biosensors. Biosens. Bioelectron. 2007, 22, 1282–1288. [Google Scholar] [CrossRef] [PubMed]
- Zourob, M.; Mohr, S.; Brown, B.J.T.; Fielden, P.R.; McDonnell, M.; Goddard, N.J. The development of a metal clad leaky waveguide sensor for the detection of particles. Sens. Actuators B Chem. 2003, 90, 296–307. [Google Scholar] [CrossRef]
- Zourob, M.; Mohr, S.; Treves Brown, B.J.; Fielden, P.R.; McDonnell, M.B.; Goddard, N.J. An integrated optical leaky waveguide sensor with electrically induced concentration system for the detection of bacteria. Lab Chip 2005, 5, 1360–1365. [Google Scholar] [CrossRef]
- Zourob, M.; Mohr, S.; Goddard, N.J. Integrated deep-probe optical waveguides for label free bacterial detection. Conf. Proc. Int. Symp. Signals, Syst. Electron. 2007, 1, 49–52. [Google Scholar] [CrossRef]
- Zourob, M.; Hawkes, J.J.; Coakley, W.T.; Brown, B.J.T.; Fielden, P.R.; McDonnell, M.B.; Goddard, N.J. Optical leaky waveguide sensor for detection of bacteria with ultrasound attractor force. Anal. Chem. 2005, 77, 6163–6168. [Google Scholar] [CrossRef]
- Zourob, M.; Mohr, S.; Brown, B.J.T.; Fielden, P.R.; Mcdonnell, M.B.; Goddard, J. An Integrated Metal Clad Leaky Waveguide Sensor for Detection of Bacteria. Anal. Chem. 2005, 77, 232–242. [Google Scholar] [CrossRef]
- Yaghoubi, M.; Rahimi, F.; Negahdari, B.; Rezayan, A.H.; Shafiekhani, A. A lectin-coupled porous silicon-based biosensor: Label-free optical detection of bacteria in a real-time mode. Sci. Rep. 2020, 10, 16017. [Google Scholar] [CrossRef]
- Maldonado, J.; Estévez, M.C.; Fernández-Gavela, A.; González-López, J.J.; González-Guerrero, A.B.; Lechuga, L.M. Label-free detection of nosocomial bacteria using a nanophotonic interferometric biosensor. Analyst 2020, 145, 497–506. [Google Scholar] [CrossRef]
- Ramsden, J.J.; Kurrat, R.; Roush, D.J.; Gill, D.S.; Willson, R.C. Protein adsorption kinetics drastically altered by repositioning a single charge. J. Am. Chem. Soc. 1995, 117, 8511–8516. [Google Scholar] [CrossRef]
- Hussain, Y.; Krimb, J.; Granta, C. OTS adsorption: A dynamic QCM study. Colloids Surf. A Physicochem. Eng. Asp. 2005, 262, 81–86. [Google Scholar] [CrossRef]
- Yu, X.; Chen, F.; Wang, R.; Li, Y. Whole-bacterium SELEX of DNA aptamers for rapid detection of E.coli O157:H7 using a QCM sensor. J. Biotechnol. 2018, 266, 39–49. [Google Scholar] [CrossRef]
- Wang, L.; Wang, R.; Chen, F.; Jiang, T.; Wang, H.; Slavik, M.; Wei, H.; Li, Y. QCM-based aptamer selection and detection of Salmonella typhimurium. Food Chem. 2017, 221, 776–782. [Google Scholar] [CrossRef]
- Bayramoglu, G.; Ozalp, V.C.; Oztekin, M.; Arica, M.Y. Rapid and label-free detection of Brucella melitensis in milk and milk products using an aptasensor. Talanta 2019, 200, 263–271. [Google Scholar] [CrossRef] [PubMed]
- Bunroddith, K.; Viseshakul, N.; Chansiri, K.; Lieberzeit, P. QCM-based rapid detection of PCR amplification products of Ehrlichia canis. Anal. Chim. Acta 2018, 1001, 106–111. [Google Scholar] [CrossRef] [PubMed]
- Masdor, N.A.; Altintas, Z.; Tothill, I.E. Sensitive detection of Campylobacter jejuni using nanoparticles enhanced QCM sensor. Biosens. Bioelectron. 2016, 78, 328–336. [Google Scholar] [CrossRef]
- Fulgione, A.; Cimafonte, M.; Della Ventura, B.; Iannaccone, M.; Ambrosino, C.; Capuano, F.; Proroga, Y.T.R.; Velotta, R.; Capparelli, R. QCM-based immunosensor for rapid detection of Salmonella Typhimurium in food. Sci. Rep. 2018, 8, 16137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hong, S.R.; Kim, M.S.; Jeong, H.D.; Hong, S. Development of real-time and quantitative QCM immunosensor for the rapid diagnosis of Aeromonas hydrophila infection. Aquac. Res. 2017, 48, 2055–2063. [Google Scholar] [CrossRef]
- Latif, U.; Can, S.; Sussitz, H.F.; Dickert, F.L. Molecular imprinted based quartz crystal microbalance sensors for bacteria and spores. Chemosensors 2020, 8, 64. [Google Scholar] [CrossRef]
- Spieker, E.; Lieberzeit, P.A. Molecular imprinting studies for developing QCM-sensors for Bacillus Cereus. Procedia Eng. 2016, 168, 561–564. [Google Scholar] [CrossRef]
- Salam, F.; Uludag, Y.; Tothill, I.E. Talanta Real-time and sensitive detection of Salmonella Typhimurium using an automated quartz crystal microbalance (QCM) instrument with nanoparticles ampli fi cation. Talanta 2013, 115, 761–767. [Google Scholar] [CrossRef] [PubMed]
- Zheng, F.; Wang, P.; Du, Q.; Chen, Y.; Liu, N. Simultaneous and ultrasensitive detection of foodborne bacteria by gold nanoparticles-amplified microcantilever array biosensor. Front. Chem. 2019, 7, 232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X.; Wang, L.; Zhao, J.; Zhu, Y.; Yang, J.; Yang, F. Enhanced binding efficiency of microcantilever biosensor for the detection of yersinia. Sensors 2019, 19, 3326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gil-Santos, E.; Ruz, J.J.; Malvar, O.; Favero, I.; Lemaître, A.; Kosaka, P.M.; García-López, S.; Calleja, M.; Tamayo, J. Optomechanical detection of vibration modes of a single bacterium. Nat. Nanotechnol. 2020, 15, 469–474. [Google Scholar] [CrossRef] [PubMed]
- Sharma, H.; Mutharasan, R. Biosensors and Bioelectronics Rapid and sensitive immunodetection of Listeria monocytogenes in milk using a novel piezoelectric cantilever sensor. Biosens. Bioelectron. 2013, 45, 158–162. [Google Scholar] [CrossRef]
- Campbell, G.A.; Mutharasan, R.A.J. A Method of Measuring Escherichia Coli O157: H7 at 1 Cell/mL in 1 Liter Sample Using Antibody Functionalized Millimeter-Sized Cantilever Sensor. Environ. Sci. Technol. 2007, 41, 1668–1674. [Google Scholar] [CrossRef]
- Jasim, I.; Shen, Z.; Mlaji, Z.; Yuksek, N.S.; Abdullah, A.; Liu, J.; Dastider, S.G.; El-Dweik, M.; Zhang, S.; Almasri, M. An impedance biosensor for simultaneous detection of low concentration of Salmonella serogroups in poultry and fresh produce samples. Biosens. Bioelectron. 2019, 126, 292–300. [Google Scholar] [CrossRef]
- Xue, L.; Guo, R.; Huang, F.; Qi, W.; Liu, Y.; Cai, G.; Lin, J. An impedance biosensor based on magnetic nanobead net and MnO2 nanoflowers for rapid and sensitive detection of foodborne bacteria. Biosens. Bioelectron. 2021, 173, 112800. [Google Scholar] [CrossRef]
- Wang, S.; Peng, T.; Meng, Q.; Zhu, X.; Guo, L.; Yao, K.; Wang, Z.; Zheng, P.; Ren, Z.; He, Z.; et al. Rapid and ultrasensitive detection of Salmonella typhimurium using a novel impedance biosensor based on SiO2@MnO2 nanocomposites and interdigitated array microelectrodes. Sens. Actuators B Chem. 2020, 324, 128654. [Google Scholar] [CrossRef]
- Wang, L.; Huo, X.; Qi, W.; Xia, Z.; Li, Y.; Lin, J. Rapid and sensitive detection of Salmonella typhimurium using nickel nanowire bridge for electrochemical impedance amplification. Talanta 2020, 211, 120715. [Google Scholar] [CrossRef]
- Abdelrasoul, G.N.; Anwar, A.; MacKay, S.; Tamura, M.; Shah, M.A.; Khasa, D.P.; Montgomery, R.R.; Ko, A.I.; Chen, J. DNA aptamer-based non-faradaic impedance biosensor for detecting E. coli. Anal. Chim. Acta 2020, 1107, 135–144. [Google Scholar] [CrossRef] [PubMed]
- Golabi, M.; Kuralay, F.; Jager, E.W.H.; Beni, V.; Turner, A.P.F. Electrochemical bacterial detection using poly(3-aminophenylboronic acid)-based imprinted polymer. Biosens. Bioelectron. 2017, 93, 87–93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, L.; Wang, L.; Zhang, X.; Li, T.; Wang, Y.; Riaz, M.A.; Sui, X.; Yuan, Z.; Chen, Y. Interfacial engineering of graphenic carbon electrodes by antimicrobial polyhexamethylene guanidine hydrochloride for ultrasensitive bacterial detection. Carbon N. Y. 2020, 159, 185–194. [Google Scholar] [CrossRef]
- Rica, R.D.; Baldi, A. Selective Detection of Live Pathogens via surface-confined electric field perturbation on interdigitated silicon transducers. Anal. Chem. 2009, 81, 3830–3835. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mathelié-Guinlet, M.; Cohen-Bouhacina, T.; Gammoudi, I.; Martin, A.; Béven, L.; Delville, M.H.; Grauby-Heywang, C. Silica nanoparticles-assisted electrochemical biosensor for the rapid, sensitive and specific detection of Escherichia coli. Sens. Actuators B Chem. 2019, 292, 314–320. [Google Scholar] [CrossRef]
- Nguyen, T.H.P.; Tonezzer, M.; Dang, T.T.L.; Vu, Q.K.; Tran, Q.H.; Nguyen, D.H.; Nguyen, V.H. Stable electrochemical measurements of platinum screen-printed electrodes modified with vertical ZnO nanorods for bacterial detection. J. Nanomater. 2019, 2341268. [Google Scholar] [CrossRef] [Green Version]
- Farooq, U.; Ullah, M.W.; Yang, Q.; Aziz, A.; Xu, J.; Zhou, L.; Wang, S. High-density phage particles immobilization in surface-modified bacterial cellulose for ultra-sensitive and selective electrochemical detection of Staphylococcus aureus. Biosens. Bioelectron. 2020, 157, 112163. [Google Scholar] [CrossRef]
- Li, L.; Yuan, Y.; Chen, Y.; Zhang, P.; Bai, Y.; Bai, L. Aptamer based voltammetric biosensor for Mycobacterium tuberculosis antigen ESAT-6 using a nanohybrid material composed of reduced graphene oxide and a metal-organic framework. Microchim. Acta 2018, 185, 2–10. [Google Scholar] [CrossRef]
- Zuser, K.; Ettenauer, J.; Kellner, K.; Posnicek, T.; Mazza, G.; Brandl, M. A Sensitive Voltammetric Biosensor for Escherichia Coli Detection Using an Electroactive Substrate for β-d-Glucuronidase. IEEE Sens. J. 2019, 19, 7789–7802. [Google Scholar] [CrossRef]
- Eissa, S.; Zourob, M. Ultrasensitive peptide-based multiplexed electrochemical biosensor for the simultaneous detection of Listeria monocytogenes and Staphylococcus aureus. Microchim. Acta 2020, 187, 486. [Google Scholar] [CrossRef]
- Lee, C.S.; Kyu Kim, S.; Kim, M. Ion-sensitive field-effect transistor for biological sensing. Sensors 2009, 9, 7111–7131. [Google Scholar] [CrossRef] [PubMed]
- Silva, N.F.D.; Magalhães, J.M.C.S.; Barroso, M.F.; Oliva-Teles, T.; Freire, C.; Delerue-Matos, C. In situ formation of gold nanoparticles in polymer inclusion membrane: Application as platform in a label-free potentiometric immunosensor for Salmonella typhimurium detection. Talanta 2019, 194, 134–142. [Google Scholar] [CrossRef]
- Dhull, N.; Kaur, G.; Jain, P.; Mishra, P.; Singh, D.; Ganju, L.; Gupta, V.; Tomar, M. Label-free amperometric biosensor for Escherichia coli O157:H7 detection. Appl. Surf. Sci. 2019, 495, 143548. [Google Scholar] [CrossRef]
- Vásquez, G.; Rey, A.; Rivera, C.; Iregui, C.; Orozco, J. Amperometric biosensor based on a single antibody of dual function for rapid detection of Streptococcus agalactiae. Biosens. Bioelectron. 2017, 87, 453–458. [Google Scholar] [CrossRef]
- Boulade, M.; Morlay, A.; Piat, F.; Roupioz, Y.; Livache, T.; Charette, P.G.; Canva, M.; Leroy, L. Early detection of bacteria using SPR imaging and event counting: Experiments with: Listeria monocytogenes and Listeria innocua. RSC Adv. 2019, 9, 15554–15560. [Google Scholar] [CrossRef] [Green Version]
- Faridfar, G.; Zeinoddini, M.; Akbarzedehkolahi, S.; Faridfar, S.; Nemati, A.S. Immunodiagnostic of Vibrio cholerae O1 using localized surface plasmon resonance (LSPR) biosensor. Int. Microbiol. 2021, 24, 115–122. [Google Scholar] [CrossRef]
- Yaghubi, F.; Zeinoddini, M.; Saeedinia, A.R.; Azizi, A.; Samimi Nemati, A. Design of localized surface plasmon resonance (LSPR) biosensor for immunodiagnostic of E. coli O157:H7 using gold nanoparticles conjugated to the chicken antibody. Plasmonics 2020, 15, 1481–1487. [Google Scholar] [CrossRef]
- Kushwaha, A.S.; Kumar, A.; Kumar, R.; Srivastava, M.; Srivastava, S.K. Zinc oxide, gold and graphene-based surface plasmon resonance (SPR) biosensor for detection of Pseudomonas like bacteria: A comparative study. Optik 2018, 172, 697–707. [Google Scholar] [CrossRef]
- Galvan, D.D.; Parekh, V.; Liu, E.; Liu, E.L.; Yu, Q. Sensitive bacterial detection via dielectrophoretic-enhanced mass transport using surface-plasmon-resonance biosensors. Anal. Chem. 2018, 90, 14635–14642. [Google Scholar] [CrossRef]
- Prabowo, B.A.; Chang, Y.F.; Lai, H.C.; Alom, A.; Pal, P.; Lee, Y.Y.; Chiu, N.F.; Hatanaka, K.; Su, L.C.; Liu, K.C. Rapid screening of Mycobacterium tuberculosis complex (MTBC) in clinical samples by a modular portable biosensor. Sens. Actuators B Chem. 2018, 254, 742–748. [Google Scholar] [CrossRef]
- Prakash, O.; Sil, S.; Verma, T.; Umapathy, S. Direct detection of bacteria using positively charged Ag/Au bimetallic nanoparticles: A label-free surface-enhanced Raman scattering study coupled with multivariate analysis. J. Phys. Chem. C 2020, 124, 861–869. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Tang, M.; Liu, Y.; Huang, J.; Liu, Z.; Tian, H.; Zheng, Y.; de la Chapelle, M.L.; Zhang, Y.; Fu, W. Surface-enhanced Raman scattering method for the identification of methicillin-resistant Staphylococcus aureus using positively charged silver nanoparticles. Microchim. Acta 2019, 186, 102. [Google Scholar] [CrossRef] [PubMed]
- Yeh, Y.P.; Cacace, M.G.; Morgan, S.L.; Ramsden, J.J. Differential adhesion of bacteria to different uniform metal oxide surfaces. (submitted research article).
- Adányi, N.; Bori, Z.; Szendro, I.; Erdélyi, K.; Wang, X.; Schröder, H.C.; Müller, W.E.G. Bacterial sensors based on biosilica immobilization for label-free OWLS detection. N. Biotechnol. 2013, 30, 493–499. [Google Scholar] [CrossRef]
- Edina, N.; Hal, A. Application of electrochemical optical waveguide lightmode spectroscopy for studying the effect of different stress factors on lactic acid bacteria. Anal. Chim. Acta 2006, 574, 41–47. [Google Scholar] [CrossRef]
- Pohanka, M. QCM immunosensor for the determination of Staphylococcus aureus antigen. Chem. Pap. 2020, 74, 451–458. [Google Scholar] [CrossRef]
- Mertens, J.; Cuervo, A.; Carrascosa, J.L. Nanomechanical detection of Escherichia coli infection by bacteriophage T7 using cantilever sensors. Nanoscale 2019, 11, 17689–17698. [Google Scholar] [CrossRef]
- Xu, Y.; Li, C.; Jiang, Y.; Guo, M.; Yang, Y.; Yang, Y.; Yu, H. Electrochemical Impedance Spectroscopic Detection of E. coli with Machine Learning. J. Electrochem. Soc. 2020, 167, 047508. [Google Scholar] [CrossRef]
- Rengaraj, S.; Cruz-Izquierdo, Á.; Scott, J.L.; Di Lorenzo, M. Impedimetric paper-based biosensor for the detection of bacterial contamination in water. Sens. Actuators B Chem. 2018, 265, 50–58. [Google Scholar] [CrossRef]
- Wang, L.; Xue, L.; Guo, R.; Zheng, L.; Wang, S.; Yao, L.; Huo, X.; Liu, N.; Liao, M.; Li, Y.; et al. Combining impedance biosensor with immunomagnetic separation for rapid screening of Salmonella in poultry supply chains. Poult. Sci. 2020, 99, 1606–1614. [Google Scholar] [CrossRef]
- Abdullah, A.; Dastider, S.G.; Jasim, I.; Shen, Z.; Yuksek, N.; Zhang, S.; Dweik, M.; Almasri, M. Microfluidic based impedance biosensor for pathogens detection in food products. Electrophoresis 2019, 40, 508–520. [Google Scholar] [CrossRef]
- Silva, N.F.D.; Almeida, C.M.R.; Magalhães, J.M.C.S.; Gonçalves, M.P.; Freire, C.; Delerue-Matos, C. Development of a disposable paper-based potentiometric immunosensor for real-time detection of a foodborne pathogen. Biosens. Bioelectron. 2019, 141, 111317. [Google Scholar] [CrossRef] [PubMed]
- Özcan, M.; Sesal, N.C.; Şener, M.K.; Koca, A. An alternative strategy to detect bacterial contamination in milk and water: A newly designed electrochemical biosensor. Eur. Food Res. Technol. 2020, 246, 1317–1324. [Google Scholar] [CrossRef]
- Feng, Y.; Zhou, D.; Gao, L.; He, F. Electrochemical biosensor for rapid detection of bacteria based on facile synthesis of silver wire across electrodes. Biosens. Bioelectron. 2020, 168, 112527. [Google Scholar] [CrossRef]
- Hicks, J.M.; Halkerston, R.; Silman, N.; Jackson, S.K.; Aylott, J.W.; Rawson, F.J. Real-time bacterial detection with an intracellular ROS sensing platform. Biosens. Bioelectron. 2019, 141, 111430. [Google Scholar] [CrossRef]
- McLeod, J.; Park, C.; Cunningham, A.; O’Donnell, L.; Brown, R.S.; Kelly, F.; She, Z. Developing a toll-like receptor biosensor for Gram-positive bacterial detection and its storage strategies. Analyst 2020, 145, 6024–6031. [Google Scholar] [CrossRef]
- Bu, S.; Wang, K.; Li, Z.; Wang, C.; Hao, Z.; Liu, W.; Wan, J. An electrochemical biosensor based on methylene blue-loaded nanocomposites as signal-amplifying tags to detect pathogenic bacteria. Analyst 2020, 145, 4328–4334. [Google Scholar] [CrossRef]
- Nikkhoo, N.; Cumby, N.; Gulak, P.G.; Maxwell, K.L. Rapid bacterial detection via an all-electronic CMOS biosensor. PLoS ONE 2016, 11, e0162438. [Google Scholar] [CrossRef]
- Maldonado, J.; González-Guerrero, A.B.; Fernández-Gavela, A.; González-López, J.J.; Lechuga, L.M. Ultrasensitive label-free detection of unamplified multidrug-resistance bacteria genes with a bimodal waveguide interferometric biosensor. Diagnostics 2020, 10, 845. [Google Scholar] [CrossRef]
- Chen, Q.; Huang, F.; Cai, G.; Wang, M.; Lin, J. An optical biosensor using immunomagnetic separation, urease catalysis and pH indication for rapid and sensitive detection of Listeria monocytogenes. Sens. Actuators B Chem. 2018, 258, 447–453. [Google Scholar] [CrossRef]
- Hu, J.; Tang, F.; Jiang, Y.Z.; Liu, C. Rapid screening and quantitative detection of Salmonella using a quantum dot nanobead-based biosensor. Analyst 2020, 145, 2184–2190. [Google Scholar] [CrossRef]
- Kumar, N.; Wang, W.; Ortiz-Marquez, J.C.; Catalano, M.; Gray, M.; Biglari, N.; Hikari, K.; Ling, X.; Gao, J.; van Opijnen, T.; et al. Dielectrophoresis assisted rapid, selective and single cell detection of antibiotic resistant bacteria with G-FETs. Biosens. Bioelectron. 2020, 156, 112123. [Google Scholar] [CrossRef] [PubMed]
- Peter, B.; Lagzi, I.; Teraji, S.; Nakanishi, H.; Cervenak, L.; Zámbó, D.; Deák, A.; Molnár, K.; Truszka, M.; Szekacs, I.; et al. Interaction of positively charged gold nanoparticles with cancer cells monitored by an in situ label-free optical biosensor and transmission electron microscopy. ACS Appl. Mater. Interfaces 2018, 10, 26841–26850. [Google Scholar] [CrossRef] [PubMed]
- Peter, B.; Ungai-Salanki, R.; Szabó, B.; Nagy, A.G.; Szekacs, I.; Bösze, S.; Horvath, R. High-resolution adhesion kinetics of EGCG-exposed tumor cells on biomimetic interfaces: Comparative monitoring of cell viability using label-free biosensor and classic end-point assays. ACS Omega 2018, 3, 3882–3891. [Google Scholar] [CrossRef] [PubMed]
- Kanyo, N.; Kovacs, K.D.; Saftics, A.; Szekacs, I.; Peter, B.; Santa-Maria, A.R.; Walter, F.R.; Dér, A.; Deli, M.A.; Horvath, R. Glycocalyx regulates the strength and kinetics of cancer cell adhesion revealed by biophysical models based on high resolution label-free optical data. Sci. Rep. 2020, 10, 22422. [Google Scholar] [CrossRef] [PubMed]
- Kurucz, I.; Peter, B.; Prosz, A.; Szekacs, I.; Horvath, R.; Erdei, A. Sensors and Actuators B: Chemical Label-free optical biosensor for on-line monitoring the integrated response of human B cells upon the engagement of stimulatory and inhibitory immune receptors. Sens. Actuators B Chem. 2017, 240, 528–535. [Google Scholar] [CrossRef]
- Kliment, K.; Szekacs, I.; Peter, B.; Erdei, A.; Kurucz, I.; Horvath, R. Label-free real-time monitoring of the BCR-triggered activation of primary human B cells modulated by the simultaneous engagement of inhibitory receptors. Biosens. Bioelectron. 2021, 191, 113469. [Google Scholar] [CrossRef]
- Ramsden, J.J.; Horvath, R. Optical biosensors for cell adhesion. J. Recept. Signal Transduct. Res. 2009, 29, 211–223. [Google Scholar] [CrossRef]
- Farkas, E.; Tarr, R.; Gerecsei, T.; Saftics, A.; Kovács, K.D.; Stercz, B.; Domokos, J.; Peter, B.; Kurunczi, S.; Szekacs, I.; et al. Development and in-depth characterization of bacteria repellent and bacteria adhesive antibody-coated surfaces using optical waveguide biosensing. Biosensors 2022, 12, 56. [Google Scholar] [CrossRef]
- Baker, S.J.; Payne, D.J.; Rappuoli, R.; De Gregorio, E. Technologies to address antimicrobial resistance. Proc. Natl. Acad. Sci. USA 2018, 115, 12887–12895. [Google Scholar] [CrossRef] [Green Version]
- Buchy, P.; Ascioglu, S.; Buisson, Y.; Datta, S.; Nissen, M.; Tambyah, P.A.; Vong, S. Impact of vaccines on antimicrobial resistance. Int. J. Infect. Dis. 2020, 90, 188–196. [Google Scholar] [CrossRef] [Green Version]
- Teuber, M. Veterinary use and antibiotic resistance. Curr. Opin. Microbiol. 2001, 4, 493–499. [Google Scholar] [CrossRef]
- Wright, G.D. The antibiotic resistome: The nexus of chemical and genetic diversity. Nat. Rev. Microbiol. 2007, 5, 175–186. [Google Scholar] [CrossRef]
- Sauermann, R.; Graninger, W. Daptomycin: A review 4 years after first approval. Pharmacology 2008, 81, 79–91. [Google Scholar] [CrossRef] [PubMed]
- Wenzel, R.; Bate, G.; Kirkpatrick, P. Tigecycline. Nat. Rev. Drug Discov. 2005, 4, 809–810. [Google Scholar] [CrossRef]
- Conly, J.M.; Johnston, B.L. Where are all the new antibiotics? The new antibiotic paradox. Can. J. Infect. Dis. Med. Microbiol. 2005, 16, 159–160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, M.Z.H. Recent Biosensors for detection of antibiotics in animal derived food. Crit. Rev. Anal. Chem. 2020, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Cristea, C.; Tertis, M.; Galatus, R. Magnetic nanoparticles for antibiotics detection. Nanomaterials 2017, 7, 119. [Google Scholar] [CrossRef]
- Kling, A.; Chatelle, C.; Armbrecht, L.; Qelibari, E.; Kieninger, J.; Dincer, C.; Weber, W.; Urban, G. Multianalyte antibiotic detection on an electrochemical microfluidic platform. Anal. Chem. 2016, 88, 10036–10043. [Google Scholar] [CrossRef]
- Puchades, R.; Cha, C. Fast screening methods to detect antibiotic residues in food samples. TrAC Trends Anal. Chem. 2010, 29, 1038–1049. [Google Scholar] [CrossRef]
- Murray, E.M.; Allen, C.F.; Handy, T.E.; Hu, C.A.; Craig, W.R.; Seaton, S.C.; Wolfe, A.L. Development of a robust and quantitative high-throughput screening method for antibiotic production in bacterial libraries. ACS Omega 2019, 4, 15414–15420. [Google Scholar] [CrossRef]
- Lautenschläger, N.; Popp, P.F.; Mascher, T. Development of a novel heterologous β-lactam-specific whole-cell biosensor in Bacillus subtilis. J. Biol. Eng. 2020, 14, 21. [Google Scholar] [CrossRef] [PubMed]
- Yin, J.; Cheng, D.; Zhu, Y.; Liang, Y.; Yu, Z. Development of a whole-cell biosensor for detection of antibiotics targeting bacterial cell envelope in Bacillus subtilis. Appl. Microbiol. Biotechnol. 2022, 106, 789–798. [Google Scholar] [CrossRef]
- Ates, H.C.; Mohsenin, H.; Wenzel, C.; Glatz, R.T.; Wagner, H.J.; Bruch, R.; Hoefflin, N.; Spassov, S.; Streicher, L.; Lozano-Zahonero, S.; et al. Biosensor-enabled multiplexed on-site therapeutic drug monitoring of antibiotics. Adv. Mater. 2022, 34, 2104555. [Google Scholar] [CrossRef]
- Rawson, T.M.; Gowers, S.A.N.; Freeman, D.M.E.; Wilson, R.C.; Sharma, S.; Gilchrist, M.; MacGowan, A.; Lovering, A.; Bayliss, M.; Kyriakides, M.; et al. Microneedle biosensors for real-time, minimally invasive drug monitoring of phenoxymethylpenicillin: A first-in-human evaluation in healthy volunteers. Lancet Digit. Health 2019, 1, e335–e343. [Google Scholar] [CrossRef] [Green Version]
- Gupta, A.; Akin, D.; Bashir, R. Single virus particle mass detection using microresonators with nanoscale thickness. Appl. Phys. Lett. 2004, 1976, 9–12. [Google Scholar] [CrossRef]
- Ilic, B.; Czaplewski, D.; Zalalutdinov, M.; Craighead, H.G.; Neuzil, P.; Campagnolo, C.; Batt, C.; Ilic, B.; Czaplewski, D.; Zalalutdinov, M.; et al. Single cell detection with micromechanical oscillators Single cell detection with micromechanical oscillators. J. Vac. Sci. Technol. B Microelectron. Nanometer Struct. Process. Meas. Phenom. 2014, 19, 2825–2828. [Google Scholar] [CrossRef]
- Reyes, P.I.; Yang, K.; Zheng, A.; Li, R.; Li, G.; Lu, Y.; Tsang, C.K.; Zheng, S.X.F. Dynamic monitoring of antimicrobial resistance using magnesium zinc oxide nanostructure-modified quartz crystal microbalance. Biosens. Bioelectron. 2017, 93, 189–197. [Google Scholar] [CrossRef] [Green Version]
- Leonard, H.; Halachmi, S.; Ben-Dov, N.; Nativ, O.; Segal, E. Unraveling antimicrobial susceptibility of bacterial networks on micropillar architectures using intrinsic phase-shift spectroscopy. ACS Nano 2017, 11, 6167–6177. [Google Scholar] [CrossRef]
- Kumarasamy, K.K.; Toleman, M.A.; Walsh, T.R.; Bagaria, J.; Butt, F.; Balakrishnan, R.; Chaudhary, U.; Paterson, D.L.; Pearson, A.; Perry, C.; et al. Emergence of a new antibiotic resistance mechanism in India, Pakistan, and the UK: A molecular, biological, and epidemiological study. Lancet Infect. Dis. 2010, 10, 597–602. [Google Scholar] [CrossRef]
- Xu, L.; Liang, W.; Wen, Y.; Wang, L.; Yang, X.; Ren, S.; Jia, N.; Zuo, X.; Liu, G. An ultrasensitive electrochemical biosensor for the detection of mecA gene in methicillin-resistant Staphylococcus aureus. Biosens. Bioelectron. 2018, 99, 424–430. [Google Scholar] [CrossRef]
- Saucedo, N.M.; Srinives, S.; Mulchandani, A. Electrochemical biosensor for rapid detection of viable bacteria and antibiotic screening. J. Anal. Test. 2019, 3, 117–122. [Google Scholar] [CrossRef]
- Hu, C.; Kalsi, S.; Zeimpekis, I.; Sun, K.; Ashburn, P.; Turner, C.; Sutton, J.M.; Morgan, H. Ultra-fast electronic detection of antimicrobial resistance genes using isothermal amplification and Thin Film Transistor sensors. Biosens. Bioelectron. 2017, 96, 281–287. [Google Scholar] [CrossRef] [PubMed]
- Nag, P.; Sadani, K.; Mukherji, S.; Mukherji, S. Beta-lactam antibiotics induced bacteriolysis on LSPR sensors for assessment of antimicrobial resistance and quantification of antibiotics. Sens. Actuators B Chem. 2020, 311, 127945. [Google Scholar] [CrossRef]
- Sabhachandani, P.; Sarkar, S.; Zucchi, P.C.; Whitfield, B.A.; Kirby, J.E.; Hirsch, E.B.; Konry, T. Integrated microfluidic platform for rapid antimicrobial susceptibility testing and bacterial growth analysis using bead-based biosensor via fluorescence imaging. Microchim. Acta 2017, 184, 4619–4628. [Google Scholar] [CrossRef]
- Rohrbach, A. Stiffness of optical traps: Quantitative agreement between experiment and electromagnetic theory. Phys. Rev. Lett. 2005, 95, 168102. [Google Scholar] [CrossRef] [Green Version]
- Tardif, M.; Jager, J.B.; Marcoux, P.R.; Uchiyamada, K.; Picard, E.; Hadji, E.; Peyrade, D. Single-cell bacterium identification with a SOI optical microcavity. Appl. Phys. Lett. 2016, 109, 133510. [Google Scholar] [CrossRef]
- van Leest, T.; Caro, J. Cavity-enhanced optical trapping of bacteria using a silicon photonic crystal. Lab Chip 2013, 13, 4358–4365. [Google Scholar] [CrossRef] [Green Version]
- Conteduca, D.; Brunetti, G.; Dell’Olio, F.; Armenise, M.N.; Krauss, T.F.; Ciminelli, C. Monitoring of individual bacteria using electro-photonic traps. Biomed. Opt. Express 2019, 10, 3463. [Google Scholar] [CrossRef]
- Brunetti, G.; Conteduca, D.; Armenise, M.N.; Ciminelli, C. Novel micro-nano optoelectronic biosensor for label-free real-time biofilm monitoring. Biosensors 2021, 11, 361. [Google Scholar] [CrossRef]
- Subramanian, S.; Tolstaya, E.I.; Winkler, T.E.; Bentley, W.E.; Ghodssi, R. An integrated microsystem for real-time detection and threshold-activated treatment of bacterial biofilms. ACS Appl. Mater. Interfaces 2017, 9, 31362–31371. [Google Scholar] [CrossRef]
- Antadze, I.; Dadunashvili, M.; Gunia, S.; Balarjishvili, N.; Tevdoradze, E.; Pataridze, T.; Rj, O.; Hagius, S.; Elzer, P. Diversity of phage-host specificity in Brucella phage. J Bacteriol Mycol. 2017, 4, 1049. [Google Scholar]
- Singh, A.; Poshtiban, S.; Evoy, S. Recent advances in bacteriophage based biosensors for food-borne pathogen detection. Sensors 2013, 1763–1786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Albay, A.; Kisa, O.; Baylan, O.; Doganci, L. The evaluation of FAST Plaque TB TM test for the rapid diagnosis of tuberculosis. Diagn. Microbiol. Infect. Dis. 2003, 46, 211–215. [Google Scholar] [CrossRef]
- Albert, H.; Trollip, A.; Seaman, T.; Mole, R.J. Simple, phage-based (FASTPlaque) technology to determine rifampicin resistance of Mycobacterium tuberculosis directly from sputum. Int. J. Tuberc. Lung Dis. 2004, 8, 1114–1119. [Google Scholar] [PubMed]
- Guntupalli, R.; Sorokulova, I.; Olsen, E.; Globa, L.; Pustovyy, O.; Vodyanoy, V. Biosensor for detection of antibiotic resistant Staphylococcus bacteria. J. Vis. Exp. 2013, 75, 50474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guntupalli, R.; Sorokulova, I.; Krumnow, A.; Pustovyy, O.; Olsen, E.; Vodyanoy, V. Real-time optical detection of methicillin-resistant Staphylococcus aureus using lytic phage probes. Biosens. Bioelectron. 2008, 24, 151–154. [Google Scholar] [CrossRef] [PubMed]
- Guntupalli, R.; Sorokulova, I.; Olsen, E.; Globa, L.; Pustovyy, O.; Moore, T.; Chin, B.; Barbaree, J.; Vodyanoy, V. Detection and identification of methicillin resistant and sensitive strains of Staphylococcus aureus using tandem measurements. J. Microbiol. Methods 2012, 90, 182–191. [Google Scholar] [CrossRef]
- Zheng, S.; Bawazir, M.; Dhall, A.; Kim, H.E.; He, L.; Heo, J.; Hwang, G. Implication of surface properties, bacterial motility, and hydrodynamic conditions on bacterial surface sensing and their initial adhesion. Front. Bioeng. Biotechnol. 2021, 9, 1–22. [Google Scholar] [CrossRef]
- Kandemir, N.; Vollmer, W.; Jakubovics, N.S.; Chen, J. Mechanical interactions between bacteria and hydrogels. Sci. Rep. 2018, 8, 10893. [Google Scholar] [CrossRef]
- Epstein, A.K.; Wong, T.S.; Belisle, R.A.; Boggs, E.M.; Aizenberg, J. Liquid-infused structured surfaces with exceptional anti-biofouling performance. Proc. Natl. Acad. Sci. USA 2012, 109, 13182–13187. [Google Scholar] [CrossRef] [Green Version]
- Epa, V.C.; Hook, A.L.; Chang, C.; Yang, J.; Langer, R.; Anderson, D.G.; Williams, P.; Davies, M.C.; Alexander, M.R.; Winkler, D.A. Modelling and prediction of bacterial attachment to polymers. Adv. Funct. Mater. 2014, 24, 2085–2093. [Google Scholar] [CrossRef]
- van Loosdrecht, M.C.M.; Lyklema, J.; Norde, W.; Zehnder, A.J.B. Bacterial adhesion: A physicochemical approach. Microb. Ecol. 1989, 17, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Hotze, E.M.; Phenrat, T.; Lowry, G.V. Nanoparticle aggregation: Challenges to understanding transport and reactivity in the environment. J. Environ. Qual. 2010, 39, 1909–1924. [Google Scholar] [CrossRef] [Green Version]
- Cacace, M.G.; Landau, E.M.; Ramsden, J.J. The Hofmeister series: Salt and solvent effects on interfacial phenomena. Q. Rev. Biophys. 1997, 30, 241–277. [Google Scholar] [CrossRef]
- Van Oss, C.J. Hydrophobicity of biosurfaces—Origin, quantitative determination and interaction energies. Colloids Surf. B Biointerfaces 1995, 5, 91–110. [Google Scholar] [CrossRef]
- Chia, T.W.R.; Nguyen, V.T.; McMeekin, T.; Fegan, N.; Dykes, G.A. Stochasticity of bacterial attachment and its predictability by the extended derjaguin-landau-verwey-overbeek theory. Appl. Environ. Microbiol. 2011, 77, 3757–3764. [Google Scholar] [CrossRef] [Green Version]
- Salerno, M.B.; Logan, B.E.; Velegol, D. Importance of molecular details in predicting bacterial adhesion to hydrophobie surfaces. Langmuir 2004, 20, 10625–10629. [Google Scholar] [CrossRef]
- Calonder, C.; Talbot, J.; Ramsden, J.J. The distribution of electron donor-acceptor potential on protein surfaces. J. Phys. Chem. B 2001, 105, 725–729. [Google Scholar] [CrossRef]
- Cheng, Y.; Feng, G.; Moraru, C.I. Micro-and nanotopography sensitive bacterial attachment mechanisms: A review. Front. Microbiol. 2019, 10, 191. [Google Scholar] [CrossRef] [Green Version]
- Cheng, Y.; Moraru, C.I. Long-range interactions keep bacterial cells from liquid-solid interfaces: Evidence of a bacteria exclusion zone near Nafion surfaces and possible implications for bacterial attachment. Colloids Surf. B Biointerfaces 2018, 162, 16–24. [Google Scholar] [CrossRef]
- Lerner, R.N.; Lu, Q.; Zeng, H.; Liu, Y. The effects of biofilm on the transport of stabilized zerovalent iron nanoparticles in saturated porous media. Water Res. 2012, 46, 975–985. [Google Scholar] [CrossRef] [PubMed]
- Ostuni, E.; Chapman, R.G.; Liang, M.N.; Meluleni, G.; Pier, G.; Ingber, D.E.; Whitesides, G.M. Self-assembled monolayers that resist the adsorption of cells. Langmuir 2001, 17, 6336–6343. [Google Scholar] [CrossRef]
- Rodney, M.D. Biofilms: Microbial life on surfaces. Emerg. Infect. Dis. 2002, 8, 881–890. [Google Scholar]
- Harris, J.M.; Zalipsky, S. Poly(ethylene glycol) chemistry and biological applications. In ACS Symposium Series 680; American Chemical Society: Washington, DC, USA, 1997. [Google Scholar]
- Dong Park, K.; Soo Kim, Y.; Keun Han, D.; Ha Kim, Y.; Hee Bae Lee, E.; Suh, H.; Suk Choi, K. Bacterial adhesion on PEG modified polyurethane surfaces. Biomaterials 1998, 19, 851–859. [Google Scholar] [CrossRef]
- Roosjen, A.; Kaper, H.J.; van der Mei, H.C.; Norde, W.; Busscher, H.J. Inhibition of adhesion of yeasts and bacteria by poly(ethylene oxide)-brushes on glass in a parallel plate flow chamber. Microbiology 2003, 149, 3239–3246. [Google Scholar] [CrossRef] [Green Version]
- Tiller, J.C.; Liao, C.J.; Lewis, K.; Klibanov, A.M. Designing surfaces that kill bacteria on contact. Proc. Natl. Acad. Sci. USA 2001, 98, 5981–5985. [Google Scholar] [CrossRef] [Green Version]
- Hook, A.L.; Chang, C.Y.; Yang, J.; Luckett, J.; Cockayne, A.; Atkinson, S.; Mei, Y.; Bayston, R.; Irvine, D.J.; Langer, R.; et al. Combinatorial discovery of polymers resistant to bacterial attachment. Nat. Biotechnol. 2012, 30, 868–875. [Google Scholar] [CrossRef]
- Peter, B.; Farkas, E.; Forgacs, E.; Saftics, A.; Kovacs, B.; Kurunczi, S.; Szekacs, I.; Csampai, A.; Bosze, S.; Horvath, R. Green tea polyphenol tailors cell adhesivity of RGD displaying surfaces: Multicomponent models monitored optically. Sci. Rep. 2017, 7, 42220. [Google Scholar] [CrossRef] [Green Version]
- Ventrici De Souza, J.; Liu, Y.; Wang, S.; Dörig, P.; Kuhl, T.L.; Frommer, J.; Liu, G.Y. Three-dimensional nanoprinting via direct delivery. J. Phys. Chem. B 2018, 122, 956–962. [Google Scholar] [CrossRef] [Green Version]
- Cairns-Smith, A.G. 4. Organisms. Life Puzzle 2017, 415, 51–64. [Google Scholar] [CrossRef]
- Nguyen, L.T.; Haney, E.F.; Vogel, H.J. The expanding scope of antimicrobial peptide structures and their modes of action. Trends Biotechnol. 2011, 29, 464–472. [Google Scholar] [CrossRef] [PubMed]
- Engler, A.C.; Wiradharma, N.; Ong, Z.Y.; Coady, D.J.; Hedrick, J.L.; Yang, Y.Y. Emerging trends in macromolecular antimicrobials to fight multi-drug-resistant infections. Nano Today 2012, 7, 201–222. [Google Scholar] [CrossRef]
- Lienkamp, K.; Tew, G.N. Synthetic mimics of antimicrobial peptides-a versatile ring-opening metathesis polymerization based platform for the synthesis of selective antibacterial and cell-penetrating polymers. Chem. A Eur. J. 2009, 15, 11784–11800. [Google Scholar] [CrossRef] [PubMed]
- Page, K.; Wilson, M.; Parkin, I.P. Antimicrobial surfaces and their potential in reducing the role of the inanimate environment in the incidence of hospital-acquired infections. J. Mater. Chem. 2009, 19, 3818–3831. [Google Scholar] [CrossRef]
- Foster, T.J.; Geoghegan, J.A.; Ganesh, V.K.; Höök, M. Adhesion, invasion and evasion: The many functions of the surface proteins of Staphylococcus aureus. Nat. Rev. Microbiol. 2014, 12, 49–62. [Google Scholar] [CrossRef] [Green Version]
- Niemann, H.H.; Schubert, W.D.; Heinz, D.W. Adhesins and invasins of pathogenic bacteria: A structural view. Microbes Infect. 2004, 6, 101–112. [Google Scholar] [CrossRef]
- Pizarro-Cerdá, J.; Cossart, P. Bacterial adhesion and entry into host cells. Cell 2006, 124, 715–727. [Google Scholar] [CrossRef] [Green Version]
- Tiwari, S.; Jamal, S.B.; Hassan, S.S.; Carvalho, P.V.S.D.; Almeida, S.; Barh, D.; Ghosh, P.; Silva, A.; Castro, T.L.P.; Azevedo, V. Two-component signal transduction systems of pathogenic bacteria as targets for antimicrobial therapy: An overview. Front. Microbiol. 2017, 8, 1878. [Google Scholar] [CrossRef]
- Dufrêne, Y.F.; Persat, A. Mechanomicrobiology: How bacteria sense and respond to forces. Nat. Rev. Microbiol. 2020, 18, 227–240. [Google Scholar] [CrossRef]
- Miller, M.B.; Bassler, B.L. Quorum sensing in bacteria. Annu. Rev. Microbiol. 2001, 55, 165–199. [Google Scholar] [CrossRef] [Green Version]
- Berne, C.; Ellison, C.K.; Ducret, A.; Brun, Y.V. Bacterial adhesion at the single-cell level. Nat. Rev. Microbiol. 2018, 16, 616–627. [Google Scholar] [CrossRef] [PubMed]
- Wolska, K.I.; Grudniak, A.M.; Rudnicka, Z.; Markowska, K. Genetic control of bacterial biofilms. J. Appl. Genet. 2016, 57, 225–238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rossi, E.; Paroni, M.; Landini, P. Biofilm and motility in response to environmental and host-related signals in Gram negative opportunistic pathogens. J. Appl. Microbiol. 2018, 125, 1587–1602. [Google Scholar] [CrossRef] [Green Version]
- Leo, J.C.; Oberhettinger, P.; Schütz, M.; Linke, D. The inverse autotransporter family: Intimin, invasin and related proteins. Int. J. Med. Microbiol. 2015, 305, 276–282. [Google Scholar] [CrossRef]
- Anderson, S.; Felici, M. Safety, reliability and security of industrial computer systems. Reliab. Eng. Syst. Saf. 2005, 89, 1–5. [Google Scholar] [CrossRef]
- Sauer, F.G.; Mulvey, M.A.; Schilling, J.D.; Martinez, J.J.; Hultgren, S.J. Bacterial pili: Molecular mechanisms of pathogenesis. Curr. Opin. Microbiol. 2000, 3, 65–72. [Google Scholar] [CrossRef]
- Mandlik, A.; Swierczynski, A.; Das, A.; Ton-That, H. Pili in Gram-positive bacteria: Assembly, involvement in colonization and biofilm development. Trends Microbiol. 2008, 16, 33–40. [Google Scholar] [CrossRef] [Green Version]
- Proft, T.; Baker, E.N. Pili in Gram-negative and Gram-positive bacteria—Structure, assembly and their role in disease. Cell. Mol. Life Sci. 2009, 66, 613–635. [Google Scholar] [CrossRef]
- Kolenda, R.; Ugorski, M.; Grzymajlo, K. Everything you always wanted to know about Salmonella type 1 fimbriae, but were afraid to ask. Front. Microbiol. 2019, 10, 1017. [Google Scholar] [CrossRef]
- Chen, A.; Seifert, H.S. Interactions with host cells causes Neisseria meningitidis pili to become unglued. Front. Microbiol. 2011, 2, 66. [Google Scholar] [CrossRef] [Green Version]
- Oelschlaeger, T.A. Mini-review: Adhesins as invasins. Int. J. Med. Microbiol. 2001, 291, 7–14. [Google Scholar] [CrossRef] [PubMed]
- Wilson, J.W.; Schurr, M.J.; LeBlanc, C.L.; Ramamurthy, R.; Buchanan, K.L.; Nickerson, C.A. Mechanisms of bacterial pathogenicity. Postgrad. Med. J. 2002, 78, 216–224. [Google Scholar] [CrossRef] [PubMed]
- Finlay, B.B.; Falkow, S. Common themes in microbial pathogenicity. Microbiol. Rev. 1989, 53, 210–230. [Google Scholar] [CrossRef] [PubMed]
- Naumann, M.; Rudel, T.; Meyer, T.F. Host cell interactions and signalling with Neisseria gonorrhoeae. Curr. Opin. Microbiol. 1999, 2, 62–70. [Google Scholar] [CrossRef]
- Nassif, X. Interaction mechanisms of encapsulated meningococci with eucaryotic cells: What does this tell us about the crossing of the blood-brain barrier by Neisseria meningitidis? Curr. Opin. Microbiol. 1999, 2, 71–77. [Google Scholar] [CrossRef]
- Sadarangani, M.; Pollard, A.J.; Gray-Owen, S.D. Opa proteins and CEACAMs: Pathways of immune engagement for pathogenic Neisseria. FEMS Microbiol. Rev. 2011, 35, 498–514. [Google Scholar] [CrossRef] [Green Version]
- Dehio, C.; Gray-Owen, S.D.; Meyer, T.F. The role of neisserial Opa proteins in interactions with host cells. Trends Microbiol. 1998, 6, 489–495. [Google Scholar] [CrossRef]
- Hauck, C.R.; Meyer, T.F. “Small” talk: Opa proteins as mediators of Neisseria-host-cell communication. Curr. Opin. Microbiol. 2003, 6, 43–49. [Google Scholar] [CrossRef] [Green Version]
- Virji, M.; Makepeace, K.; David, J.P.; Achtman, M.; Moxon, E.R. Meningococcal Opa and Ope proteins: Their role in colonization and invasion of human epithelial and endothelial cells. Mol. Microbiol. 1993, 10, 499–510. [Google Scholar] [CrossRef]
- Roggenkamp, A.; Ackermann, N.; Jacobi, C.A.; Truelzsch, K.; Hoffmann, H.; Heesemann, J. Molecular analysis of transport and oligomerization of the Yersinia enterocolitica adhesin YadA. J. Bacteriol. 2003, 185, 3735–3744. [Google Scholar] [CrossRef] [Green Version]
- El Tahir, Y.; Skurnik, M. YadA, the multifaceted Yersinia adhesin. Int. J. Med. Microbiol. 2001, 291, 209–218. [Google Scholar] [CrossRef] [PubMed]
- Heise, T.; Dersch, P. Identification of a domain in Yersinia virulence factor YadA that is crucial for extracellular matrix-specific cell adhesion and uptake. Proc. Natl. Acad. Sci. USA 2006, 103, 3375–3380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lecuit, M.; Ohayon, H.; Braun, L.; Mengaud, J.; Cossart, P. Internalin of Listeria monocytogenes with an intact leucine-rich repeat region is sufficient to promote internalization. Infect. Immun. 1997, 65, 5309–5319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mengaud, J.; Ohayon, H.; Gounon, P.; Mege, R.M.; Cossart, P. E-cadherin is the receptor for internalin, a surface protein required for entry of L. monocytogenes into epithelial cells. Cell 1996, 84, 923–932. [Google Scholar] [CrossRef] [Green Version]
- Schubert, W.D.; Urbanke, C.; Ziehm, T.; Beier, V.; Machner, M.P.; Domann, E.; Wehland, J.; Chakraborty, T.; Heinz, D.W. Structure of internalin, a major invasion protein of Listeria monocytogenes, in complex with its human receptor E-cadherin. Cell 2002, 111, 825–836. [Google Scholar] [CrossRef] [Green Version]
- Cossart, P.; Sansonetti, P.J. Bacterial invasion: The paradigms of enteroinvasive pathogens. Science 2004, 304, 242–248. [Google Scholar] [CrossRef] [Green Version]
- Killackey, S.A.; Sorbara, M.T.; Girardin, S.E. Cellular aspects of Shigella pathogenesis: Focus on the manipulation of host cell processes. Front. Cell. Infect. Microbiol. 2016, 6, 38. [Google Scholar] [CrossRef] [Green Version]
- Goosney, D.L.; Knoechel, D.G.; Finlay, B.B. Enteropathogenic E. coli, Salmonella, and Shigella: Masters of host cell cytoskeletal exploitation. Emerg. Infect. Dis. 1999, 5, 216–223. [Google Scholar] [CrossRef]
- Hueck, C.J. Type III Protein secretion systems in bacterial pathogens of animals and plants. Microbiol. Mol. Biol. Rev. 1998, 62, 379–433. [Google Scholar] [CrossRef] [Green Version]
- He, S.Y. Type Iii protein secretion systems in plant and animal pathogenic bacteria. Annu. Rev. Phytopathol. 1998, 36, 363–392. [Google Scholar] [CrossRef]
- Deng, W.; Marshall, N.C.; Rowland, J.L.; McCoy, J.M.; Worrall, L.J.; Santos, A.S.; Strynadka, N.C.J.; Finlay, B.B. Assembly, structure, function and regulation of type III secretion systems. Nat. Rev. Microbiol. 2017, 15, 323–337. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.C.; Hung, C.F.; Aljuffali, I.A.; Fang, J.Y. The roles of the virulence factor IpaB in Shigella spp. in the escape from immune cells and invasion of epithelial cells. Microbiol. Res. 2015, 181, 43–51. [Google Scholar] [CrossRef] [PubMed]
- Collazo, C.M.; Galán, J.E. The invasion-associated type III system of Salmonella typhimurium directs the translocation of Sip proteins into the host cell. Mol. Microbiol. 1997, 24, 747–756. [Google Scholar] [CrossRef] [PubMed]
- Cowin, P.; Burke, B. Cytoskeleton-membrane interactions. Curr. Opin. Cell Biol. 1996, 8, 56–65. [Google Scholar] [CrossRef]
- Pizarro-Cerdá, J.; Charbit, A.; Enninga, J.; Lafont, F.; Cossart, P. Manipulation of host membranes by the bacterial pathogens Listeria, Francisella, Shigella and Yersinia. Semin. Cell Dev. Biol. 2016, 60, 155–167. [Google Scholar] [CrossRef]
- Lafont, F.; Van Nhieu, G.T.; Hanada, K.; Sansonetti, P.; Van der Goot, F.G. Initial steps of Shigella infection depend on the cholesterol/sphingolipid raft-mediated CD44-IpaB interaction. EMBO J. 2002, 21, 4449–4457. [Google Scholar] [CrossRef] [Green Version]
- Baumler, A.; Fang, F.C. Host specificity of bacterial pathogens. Cold Spring Harb. Perspect. Med. 2013, 3, a010041. [Google Scholar] [CrossRef] [Green Version]
- Pan, X.; Yang, Y.; Zhang, J.R. Molecular basis of host specificity in human pathogenic bacteria. Emerg. Microbes Infect. 2014, 3, 1–10. [Google Scholar] [CrossRef]
- Ljungh, Å.; Moran, A.P.; Wadström, T. Interactions of bacterial adhesins with extracellular matrix and plasma proteins: Pathogenic implications and therapeutic possibilities. FEMS Immunol. Med. Microbiol. 1996, 16, 117–126. [Google Scholar] [CrossRef]
- Paulsson, M.; Riesbeck, K. How bacteria hack the matrix and dodge the bullets of immunity. Eur. Respir. Rev. 2018, 27, 180018. [Google Scholar] [CrossRef]
- Vaca, D.J.; Thibau, A.; Schütz, M.; Kraiczy, P.; Happonen, L.; Malmström, J.; Kempf, V.A.J. Interaction with the host: The role of fibronectin and extracellular matrix proteins in the adhesion of Gram-negative bacteria. Med. Microbiol. Immunol. 2020, 209, 277–299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Speziale, P.; Pietrocola, G. The multivalent role of fibronectin-binding proteins A and B (FnBPA and FnBPB) of Staphylococcus aureus in host infections. Front. Microbiol. 2020, 11, 1–13. [Google Scholar] [CrossRef]
- Heino, J.; Kapyla, J. Cellular receptors of extracellular matrix molecules. Curr. Pharm. Des. 2009, 15, 1309–1317. [Google Scholar] [CrossRef] [PubMed]
- Isberg, R.R.; Tran Van Nhieu, G. Binding and internalization of microorganisms by integrin receptors. Trends Microbiol. 1994, 2, 10–14. [Google Scholar] [CrossRef]
- Boyle, E.C.; Finlay, B.B. Bacterial pathogenesis: Exploiting cellular adherence. Curr. Opin. Cell Biol. 2003, 15, 633–639. [Google Scholar] [CrossRef]
- Juliano, R.L. Signal transduction by cell adhesion receptors and the cytoskeleton: Functions of integrins, cadherins, selectins, and immunoglobulin-superfamily members. Annu. Rev. Pharmacol. Toxicol. 2002, 42, 283–323. [Google Scholar] [CrossRef] [Green Version]
- Hauck, C.R.; Agerer, F.; Muenzner, P.; Schmitter, T. Cellular adhesion molecules as targets for bacterial infection. Eur. J. Cell Biol. 2006, 85, 235–242. [Google Scholar] [CrossRef] [Green Version]
- Ruoslahti, E.; Pierschbacher, M.D. New perspectives in cell adhesion: RGD and integrins. Science 1987, 238, 491–497. [Google Scholar] [CrossRef]
- Takada, Y.; Ye, X.; Simon, S. The integrins. Genome Biol. 2007, 8, 215. [Google Scholar] [CrossRef] [Green Version]
- Hauck, C.R.; Borisova, M.; Muenzner, P. Exploitation of integrin function by pathogenic microbes. Curr. Opin. Cell Biol. 2012, 24, 637–644. [Google Scholar] [CrossRef]
- Grassl, G.A.; Bohn, E.; Müller, Y.; Bühler, O.T.; Autenrieth, I.B. Interaction of Yersinia enterocolitica with epithelial cells: Invasin beyond invasion. Int. J. Med. Microbiol. 2003, 293, 41–54. [Google Scholar] [CrossRef] [PubMed]
- Krukonis, E.S.; Isberg, R.R. Integrin β1-chain residues involved in substrate recognition and specificity of binding to invasin. Cell. Microbiol. 2000, 2, 219–230. [Google Scholar] [CrossRef] [PubMed]
- Isberg, R.R.; Hamburger, Z.; Dersch, P. Signaling and invasin-promoted uptake via integrin receptors. Microbes Infect. 2000, 2, 793–801. [Google Scholar] [CrossRef]
- Miyamoto, S.; Teramoto, H.; Gutkind, J.S.; Yamada, K.M. Integrins can collaborate with growth factors for phosphorylation of receptor tyrosine kinases and MAP kinase activation: Roles of integrin aggregation and occupancy of receptors. J. Cell Biol. 1996, 135, 1633–1642. [Google Scholar] [CrossRef]
- Bohn, E.; Sonnabend, M.; Klein, K.; Autenrieth, I.B. Bacterial adhesion and host cell factors leading to effector protein injection by type III secretion system. Int. J. Med. Microbiol. 2019, 309, 344–350. [Google Scholar] [CrossRef]
- Velasco-Velázquez, M.A.; Barrera, D.; González-Arenas, A.; Rosales, C.; Agramonte-Hevia, J. Macrophage—Mycobacterium tuberculosis interactions: Role of complement receptor 3. Microb. Pathog. 2003, 35, 125–131. [Google Scholar] [CrossRef]
- Payne, N.R.; Horwitz, M.A. Phagocytosis of Legionella pneumophila is mediated by human monocyte complement receptors. J. Exp. Med. 1987, 166, 1377–1389. [Google Scholar] [CrossRef] [Green Version]
- Saukkonen, K.; Cabellos, C.; Burroughs, M.; Prasad, S.; Tuomanen, E. Integrin-mediated localization of Bordetella pertussis within macrophages: Role in pulmonary colonization. J. Exp. Med. 1991, 173, 1143–1149. [Google Scholar] [CrossRef] [Green Version]
- Cossart, P. Interactions of the bacterial pathogen Listeria monocytogenes with mammalian cells: Bacterial factors, cellular ligands, and signaling. Folia Microbiol. 1998, 43, 291–303. [Google Scholar] [CrossRef]
- Hauck, C.R. Cell adhesion receptors—Signaling capacity and exploitation by bacterial pathogens. Med. Microbiol. Immunol. 2002, 191, 55–62. [Google Scholar] [CrossRef]
- Case, E.D.R.; Samuel, J.E. Contrasting lifestyles within the host cell. Microbiol. Spectr. 2016, 4, 1–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ribet, D.; Cossart, P. How bacterial pathogens colonize their hosts and invade deeper tissues. Microbes Infect. 2015, 17, 173–183. [Google Scholar] [CrossRef] [PubMed]
- Ralph, R.; Isberg, R.R.; Portnoy, D.A. The interaction of bacteria with mammalian cells. Annu. Rev. Cell Biol. 1992, 8, 333–363. [Google Scholar]
- Ray, K.; Marteyn, B.; Sansonetti, P.J.; Tang, C.M. Life on the inside: The intracellular lifestyle of cytosolic bacteria. Nat. Rev. Microbiol. 2009, 7, 333–340. [Google Scholar] [CrossRef]
- Thakur, A.; Mikkelsen, H.; Jungersen, G. Intracellular pathogens: Host immunity and microbial persistence strategies. J. Immunol. Res. 2019, 2019, 1356540. [Google Scholar] [CrossRef]
- Haas, A. Reprogramming the phagocytic pathway—Intracellular pathogens and their vacuoles. Mol. Membr. Biol. 1998, 15, 103–121. [Google Scholar] [CrossRef]
- Uribe-Quero, E.; Rosales, C. Control of phagocytosis by microbial pathogens. Front. Immunol. 2017, 8, 1368. [Google Scholar] [CrossRef] [Green Version]
- Francis, C.L.; Ryan, T.A.; Jones, B.D.; Smith, S.J.; Falkow, S. Ruffles induced by Salmonella and other stimuli direct macropinocytosis of bacteria. Nature 1993, 364, 639–642. [Google Scholar] [CrossRef]
- Ashida, H.; Mimuro, H.; Sasakawa, C. Shigella manipulates host immune responses by delivering effector proteins with specific roles. Front. Immunol. 2015, 6, 219. [Google Scholar] [CrossRef] [Green Version]
- Fredlund, J.; Santos, J.C.; Stévenin, V.; Weiner, A.; Latour-Lambert, P.; Rechav, K.; Mallet, A.; Krijnse-Locker, J.; Elbaum, M.; Enninga, J. The entry of Salmonella in a distinct tight compartment revealed at high temporal and ultrastructural resolution. Cell. Microbiol. 2018, 20, 3958–3973. [Google Scholar] [CrossRef]
- McGhie, E.J.; Hayward, R.D.; Koronakis, V. Cooperation between actin-binding proteins of invasive Salmonella: SipA potentiates SipC nucleation and bundling of actin. EMBO J. 2001, 20, 2131–2139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marra, A.; Isberg, R.R. Bacterial pathogenesis: Common entry mechanisms. Curr. Biol. 1996, 6, 1084–1086. [Google Scholar] [CrossRef] [Green Version]
- Costerton, A.J.W.; Stewart, P.S.; Greenberg, E.P. Bacterial biofilms: A common cause of persistent infections. Science 1999, 284, 1318–1322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fassel, T.A.; Edmiston, C.E. Ruthenium red and the bacterial glycocalyx. Biotech Histochem. 1999, 74, 194–212. [Google Scholar] [CrossRef]
- Nwodo, U.U.; Green, E.; Okoh, A.I. Bacterial exopolysaccharides: Functionality and prospects. Int. J. Mol. Sci. 2012, 13, 14002–14015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cramton, S.E.; Ulrich, M.; Götz, F.; Döring, G. Anaerobic conditions induce expression of polysaccharide intercellular adhesin in Staphylococcus aureus and Staphylococcus epidermidis. Infect. Immun. 2001, 69, 4079–4085. [Google Scholar] [CrossRef] [Green Version]
- Ye, F.; Ye, Y.; Li, Y. Effect of C/N ratio on extracellular polymeric substances (EPS) and physicochemical properties of activated sludge flocs. J. Hazard. Mater. 2011, 188, 37–43. [Google Scholar] [CrossRef]
- Lukacova, M.; Barák, I.; Kazár, J. Role of structural variations of polysaccharide antigens in the pathogenicity of Gram-negative bacteria. Clin. Microbiol. Infect. 2008, 14, 200–206. [Google Scholar] [CrossRef] [Green Version]
- Mostowy, R.J.; Holt, K.E. Diversity-Generating Machines: Genetics of Bacterial Sugar-Coating. Trends Microbiol. 2018, 26, 1008–1021. [Google Scholar] [CrossRef] [Green Version]
- Cress, B.F.; Englaender, J.A.; He, W.; Kasper, D.; Linhardt, R.J.; Koffas, M.A.G. Masquerading microbial pathogens: Capsular polysaccharides mimic host-tissue molecules. FEMS Microbiol. Rev. 2014, 38, 660–697. [Google Scholar] [CrossRef] [Green Version]
- Busscher, H.J.; van der Mei, H.C. How do bacteria know they are on a surface and regulate their response to an adhering state? PLoS Pathog. 2012, 8, e1002440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bell, G.I. Models for the Specific Adhesion of Cells to Cells. Science 1978, 200, 618–627. [Google Scholar] [CrossRef] [PubMed]
- Hall-Stoodley, L.; Costerton, J.W.; Stoodley, P. Bacterial biofilms: From the natural environment to infectious diseases. Nat. Rev. Microbiol. 2004, 2, 95–108. [Google Scholar] [CrossRef] [PubMed]
- Shi, X.; Zhu, X. Biofilm formation and food safety in food industries. Trends Food Sci. Technol. 2009, 20, 407–413. [Google Scholar] [CrossRef]
- Lam, J.; Chan, R.; Lam, K.; Costerton, J.W. Production of Mucoid Microcolonies by Pseudomonas aeruginosa within Infected Lungs in Cystic Fibrosis. Infect. Immun. 1980, 28, 546–556. [Google Scholar] [CrossRef] [PubMed]
- Hiemstra, P.S.; Bloemberg, G.V.; Grote, J.J. Demonstration of bacterial cells and glycocalyx in biofilms on human tonsils. Arch. Otolaryngol. Head Neck Surg. 2007, 133, 115–121. [Google Scholar]
- Fux, C.A.; Costerton, J.W.; Stewart, P.S.; Stoodley, P. Survival strategies of infectious biofilms. Trends Microbiol. 2005, 13, 34–40. [Google Scholar] [CrossRef]
- Parsek, M.R. Bacterial biofilms: An emerging link to disease pathogenesis. Annu. Rev. Microbiol. 2003, 57, 677–701. [Google Scholar] [CrossRef]
- Potera, C. Forging a link between biofilms and disease. Science 1999, 283, 1837–1839. [Google Scholar] [CrossRef] [PubMed]
- Kostakioti, M.; Hadjifrangiskou, M.; Hultgren, S.J. Bacterial biofilms: Development, dispersal, and therapeutic strategies in the dawn of the postantibiotic era. Cold Spring Harb. Perspect. Med. 2013, 3, a010306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fey, P.D. Modality of bacterial growth presents unique targets: How do we treat biofilm-mediated infections? Curr. Opin. Microbiol. 2010, 13, 610–615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flemming, H.-C. EPS—Then and now. Microorganisms 2016, 4, 41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramsden, J.J. Partition coefficients of drugs in bilayer lipid membranes. Experientia 1993, 49, 688–692. [Google Scholar] [CrossRef] [PubMed]
- Ramsden, J.J. OWLS: A versatile technique for sensing with bioarrays. Chimia 1999, 53, 67–71. [Google Scholar]
- Ramsden, J.J. Can smart sensor systems save the NHS? Nanotechnol. Percept. 2017, 13, 69–80. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Biosensor Type | Detected Bacteria | Limit of Detection | Assay Time | Ref. |
|---|---|---|---|---|
| SPR (with nonspecific adsorption of antibody) | E. coli | 4.8 × 105 CFU/mL | No data | Torun et al. [78] |
| SPR (with specific adsorption via avidin-biotin interaction) | E. coli | 6.2 × 103 CFU/mL | No data | Torun et al. [78] |
| SPR (with SAM formation of antibody) | E. coli | 35 CFU/mL | No data | Torun et al. [78] |
| SPR (with gold coated magnetic nanoparticles) * | E. coli | 3 CFU/mL | <70 min | Torun et al. [78] |
| SPR (with one- and two step sandwich assay) | S. typhimurium | 4.7 × 105 CFU/mL (one-step sandwich assay) 1.6–1.9 × 106 CFU/mL (two-step sandwich assay) | No data | Bhandari et al. [70] |
| SPR (full-length Det7 bacteriophage tail protein (Det7T)) | S. typhimurium | 5 × 104–5 × 107 CFU/mL | ∼20 min | Hyeon et al. [76] |
| BaTiO3-graphene-affinity layer–based SPR | Pseudomonas | No data | No data | Mudgal et al. [80] |
| Resolution-optimized prism- based SPR imaging (RO-SPRI) | L. monocytogenes and L. innocua | 2 × 102 CFU/mL | 7 h | Boulade et al. [137] |
| Localized surface plasmon resonance (LSPR) | V. cholerae O1 | 10 CFU/mL | 1 h | Faridfar et al. [138] |
| Localized surface plasmon resonance (LSPR) | E. coli O157:H7 | 10 CFU/mL | 2 h | Yaghubi et al. [139] |
| Fiber-optic SPR | E. coli O157:H7 | 5 × 102 CFU/mL | No data | Zhou et al. [68] |
| Fiber-optic LSPR | S. typhimurium | 128 CFU/mL | 100 min | Xu et al. [81] |
| SPR | E. coli, Salmonella spp. | 17–57 CFU/mL for E. coli and 7.4 × 103–11.7 × 103 CFU/mL for Salmonella sp. | <80 min | Vaisocherová-Lísalová et al. [75] |
| SPR | E. faecalis | 3.4 × 104 CFU/mL | No data | Saylan et al. [67] |
| SPR | M. tuberculosis | 104 CFU/mL | 15 min | Trzaskowski et al. [74] |
| SPR | Pseudomonas and Pseudomonas-like bacteria | no data | No data | Kushwaha et al. [140] |
| SPR | Pseudomonas-like bacteria | no data | No data | Maurya et al. [79] |
| SPR | E. coli | 3.0 × 102 CFU/mL | No data | Galvan et al. [141] |
| SPR | M. tuberculosis | 63 pg/mL | 2–4 h | Prabowo et al. [142] |
| SPR | V. cholerae | 50 CFU/mL | 60 min | Taheri et al. [71] |
| SPR | E.coli O157:H7, S. enteritidis, and L. monocytogenes | 14, 6, and 28 CFU/25 g (mL) | No data | Zhang et al. [72] |
| SPR | E. coli K-12 | 2 × 104 CFU/mL | 20 min | Shin et al. [77] |
| SPR | A. salmonicida, A. hydrophila, V. harveyi | no data | No data | Padra et al. [69] |
| SPR | S. typhimurium | 105 CFU/mL | 10 min | Makhneva et al. [73] |
| SPR | L. monocytogenes | 3.25 log CFU/100 μl | 7.5 min | Raghu et al. [66] |
| SERS | Various bacteria species | ~single bacterium | No data | Yu et al. [87] |
| SERS | E. coli, S. typhimurium, and B. subtilis | no data | No data | Prakash et al. [143] |
| SERS | E. coli O157:H7 | 3 CFU/mL | No data | Zhou et al. [86] |
| SERS | S. aureus | 1–1× 106 CFU/mL | 30 min | Lee et al. [88] |
| SERS | Methicillin-resistant S. aureus (MRSA), methicillin-sensitive S. aureus (MSSA), S. aureus 29213, S. aureus 25923, C. albicans, B. cereus, E. coli, P. aeruginosa | 1.0 × 108 cells/mL (used concentrations) | 45 min | Chen et al. [144] |
| OWLS | E. coli L. plantarum | No data, purpose of work was to determine kinetics | 2 min | Yeh et al. [145] |
| OWLS | E. coli BL21AI E. coli B200 | 2 × 109 CFU/mL was applied | ~150 min | Adányi et al. [146] |
| EC-OWLS | L. plantarum 2142 | 102–103 CFU/mL | ~115 min | Adányi et al. [147] |
| OWLS (reverse symmetry waveguide design using nanoporous substrate) | E. coli K12 | 60 cells/mm2 | Minutes | Horvath et al. [90] |
| MCLW | B. subtilis var. niger bacterial spore | 8 × 104 spores/mL | 60 min | Zourob et al. [99] |
| MCLW with ultrasound standing waves | B. subtilis var. niger | 1 × 103 spores/mL | 3 min | Zourob et al. [98] |
| MCLW with an electric field | B. subtilis var. niger | 1 × 103 spores/mL | 3 min | Zourob et al. [96] |
| QCM (with direct binding assay) | S. typhimurium | ~2 × 102 CFU/mL | 5 min | Salam et al. [113] |
| QCM (with sandwich assay) | S. typhimurium | ~1.01 × 102 CFU/mL | 9 min | Salam et al. [113] |
| QCM (with nanoparticle amplification) | S. typhimurium | 10–20 CFU/mL | 12 min | Salam et al. [113] |
| QCM | S. aureus | 5.18 × 108 CFU/mL | No data | Pohanka [148] |
| QCM | B. subtilis | no data | No data | Latif et al. [111] |
| QCM | C. jejuni | 150 CFU/ mL | No data | Masdor et al. [108] |
| QCM | E. coli O157:H7 | 1.46 × 103 CFU/mL | 50 min | Yu et al. [104] |
| QCM | S. typhimurium | 103 CFU/mL | 1 h | Wang et al. [105] |
| QCM | S. typhimurium | 100 CFU/mL | No data | Fulgione et al. [109] |
| QCM | S. typhimurium | 105 CFU/mL | 10 min | Makhneva et al. [73] |
| QCM | E. canis | 4.1 × 109 molecules/μL of 289 bp E. canis | No data | Bunroddith et al. [107] |
| QCM | B. cereus | no data | 10 min | Spieker et al. [112] |
| QCM | A. hydrophila | 1.25 × 107 CFU/mL | 5 min | Hong et al. [110] |
| QCM | B. melitensis | 1.02–1.07 CFU/mL | No data | Bayramoglu et al. [106] |
| Asymmetrically anchored PEMC (aPEMC) | L. monocytogenes | 102 cells/mL | ~30 min | Sharma et al. [117] |
| PEMC | E. coli (O157:H7) | 10 cells/mL | ~50 min | Campbell and Mutharasan [118] |
| Microcantilever array biosensor | E. coli O157:H7, V. parahaemolyticus, Salmonella, S. aureus, L. monocytogenes, Shigella | 1–9 CFU/mL | <1 h | Zheng et al. [114] |
| Microcantilever sensor | Yersinia | no data | No data | Liu et al. [115] |
| Nanomechanical sensor | E. coli | ~103 CFU/mL | ~45 min | Mertens et al. [149] |
| Ultrahigh frequency mechanical resonators | S. epidermidis | 1 CFU/mL | No data | Gil-Santos et al. [116] |
| Impedimetric sensor (conductive polycrystalline silicon interdigitated electrodes) | E. coli | 3 × 102 CFU/mL | <1 h | de la Rica et al. [126] |
| DNA-aptamer based impedance biosensor | E. coli | 9 CFU/ mL | No data | Abdelrasoul et al. [123] |
| Impedance biosensor | S. typhimurium | 7 CFU/mL | 40 min | Jasim et al. [119] |
| Impedance biosensor | S. typhimurium | 19 CFU/mL | 1.5 h | Xue et al. [120] |
| Impedance biosensor | S. typhimurium | 21 CFU/mL | 50 min | Wang et al. [121] |
| Impedance biosensor | E. coli, S. aureus | 2 CFU/ mL | 30 min | Zhu et al. [125] |
| Machine learning-based electrochemical impedance spectroscopy (EIS) | E. coli | 2 × 106 and 2 × 107 CFU/mL | No data | Xu et al. [150] |
| Cyclic voltammetry and electrochemical impedance spectroscopy (EIS) | S. enteritidis | 103 CFU/mL | No data | Nguyen et al. [128] |
| Electrochemical impedance spectroscopy (EIS) | S. epidermidis | 103–107 CFU/mL | No data | Golabi et al. [124] |
| Impedimetric paper-based biosensor | Bacterial cultures from sewage sludge | 1.9 × 103 CFU/mL | 45 min | Rengaraj et al. [151] |
| Impedance biosensor with immunomagnetic separation * | S. typhimurium | 102 CFU/mL | 2 h | Wang et al. [152] |
| MEMS-based impedance biosensor | E. coli O157:H7, S. typhimurium | 10 CFU/mL | 1 h | Abdullah et al. [153] |
| Electrochemical biosensor, cyclic voltammetry | E. coli | 1 CFU/mL | No data | Zuser et al. [131] |
| Voltammetric biosensor | L. monocytogenes and S. aureus | 9 CFU/mL for L. monocytogenes and 3 CFU/ mL for S. aureus | 1 min | Eissa and Zaurob [132] |
| Voltammetric biosensor | S. aureus | 3–5 CFU/mL | 0.5 h | Farooq et al. [129] |
| Potentiometric immunosensor | S. typhimurium | 5 CFU/mL | <1 h | Silva et al. [154] |
| Amperometric biosensor | E. coli O157:H7 | 1 CFU/mL | 1 h | Dhull et al. [135] |
| Electrochemical biosensor | P. aeruginosa | 1.8 × 10−6 mol dm−3 | 5 s | Özcan et al. [155] |
| Electrochemical biosensor | E. coli, S. enteritidis, L. innocua, P. aeruginosa and S. pneumoniae | 50 CFU/mL | 100 min | Feng et al. [156] |
| Electrochemical biosensor | Gram-negative bacteria with Toll-like receptor 4 | 368 nM | 1 s | Hicks et al. [157] |
| Electrochemical aptasensor | S. typhimurium | 80 CFU/mL | 2 h | Wang et al. [122] |
| Electrochemical TLR2/6 biosensors | B. licheniformis, E. hirae | 102 CFU/mL (B. licheniformis), 104 CFU/mL (E. hirae) | No data | McLeod et al. [158] |
| Electrochemical immuno-biosensor | E. coli | 103 CFU/mL | 30 min | Mathelié-Guinlet et al. [127] |
| Sandwich-type electrochemical biosensor * | E. coli O157:H7 | 32 CFU/mL | 120 min | Bu et al. [159] |
| All-electronic complementary metal oxide semiconductor (CMOS) biosensor | Gram-positive and Gram-negative bacteria | 107 CFU/mL | 25 min | Nikkhoo et al. [160] |
| Ultrasensitive nanophotonic bimodal waveguide interferometer | Gram-negative bacteria | ~105 CFU/mL (28 aM) | 30 min | Maldonado et al. [161] |
| Lectin-conjugated porous silicon-based biosensor | E. coli and S. aureus | 103 cells/ mL | No data | Yaghoubi et al. [100] |
| Bimodal waveguide interferometer (BiMW) | P. aeruginosa and methicillin-resistant S. aureus | 49 and 29 CFU/mL (theoretical LOD) | 12 min | Maldonado et al. [101] |
| High-throughput aptamer based photo-irradiation colorimetric biosensor * | S. aureus | 81 CFU/mL | 5.5 h | Yu et al. [51] |
| Fluorescent supramolecular biosensors (fSBs) * | E. coli | 105 CFU/mL | No data | Jeong et al. [56] |
| Portable fluorescent biosensing * | E. coli O157:H7, L. monocytogenes, S. typhimurium | 102, 103, and 103 CFU/mL | 60 min | Xu et al. [53] |
| Fluorescent magnetic biosensor based on DNAzyme * | E. coli O157:H7 | 1.57 CFU/ mL | 1.5 h | Zhou et al. [50] |
| Graphene-DNAzyme-based fluorescent biosensor * | E. coli | 105 CFU/mL | 4 h | Liu et al. [55] |
| Aptamer-based fluorescent biosensor * | S. sonnei | 103 CFU/ mL | No data | Song et al. [54] |
| Optical biosensor with immunomagnetic separation * | L. monocytogenes | 102 CFU/mL | No data | Chen et al. [162] |
| Fluorescent biosensor * | E. coli O157:H7 | 14 CFU/mL | 2 h | Xue et al. [52] |
| Quantum dot nanobead-based biosensor * | S. typhimurium | 5 × 103 CFU/mL | 10 min | Hu et al. [163] |
| Nanobiosensor (AuNRs based sensor) * | E. coli, P. aeruginosa | 109 to 106 CFU/mL (measured concentrations) | No data | Kaushal et al. [82] |
| Graphene field effect transistors (G-FET) | S. aureus, colistin resistant A. baumannii | 104 CFU/mL (S. aureus), 105 CFU/mL (A.baumannii) | 5 min | Kumar et al. [164] |
| Colorimetric biosensor (paper-based magnetic nanoparticle-peptide probe) * | E. coli O157:H7 | 12 CFU/mL (broth samples), 30–300 CFU/mL (spiked complex food matrices) | 30 s | Suaifan et al. [60] |
| Colorimetric paper-based biosensor * | S. aureus (L. monocytogenesis 19115, E. coli O157:H7, MRSA, C. albicans P. aeruginosa 15692) | 7 CFU/mL (pure broth) 40 CFU/mL (inoculated in food produces) 100 CFU/mL (environmental samples) | 1 min | Suaifan et al. [35] |
| Optical immunosensor * | MRSA and non-MRSA bacteria (E. coli, S. aureus and S. epidermis) | 103 CFU/mL (visual observation) 29 CFU/mL (linear regression equation) | 5 min | Raji et al. [58] |
| Colorimetric biosensor (using magnetic nanoparticles) * | P. aeruginosa | 102 CFU/mL | <1 min | Alhogail et al. [59] |
| L. monocytogenes | 2.17 × 102 CFU/mL | <1 min | Alhogail et al. [61] |
| Biosensor Type | Bacteria | Resistance | Limit of Detection | Assay Time | Ref. |
|---|---|---|---|---|---|
| Micromechanical oscillators | E. coli (XL1-Blue) | Kanamycin, tetracycline | 100 cells on cantilever (50 pg/Hz) | 1 h (active growth) 2 h (selective growth) | Gfeller et al. [30] |
| Biofunctionalized silicon micropillar arrays | E. coli (K-12) | Gentamicin, ciprofloxacin, ampicillin, ceftriaxone, sulfamethoxazol-trimethoprim (1:19) | 103 cell/ml | 2–3 h | Leonard et al. [173] |
| Carbon screen-printed electrochemical sensor (EIS) | blaNDM (found in Enterobacteriaceae) | - | 200 nM blaNDM | No data | Obaje et al. [31] |
| QCM-D (with phage spheroids) | MRSA, MSSA (S.aureus), B. anthracis, S. typhmurium, S. flexneri, Y. enterocolitica, K.pneumoniae, B. substilis | Penicillin-binding protein antibody latex beads | 104 CFU/mL (phage capture in case of S. aureus) | 16 min/sample (spheroid-bacteria time-to-answer) | Guntupalli et al. [184] |
| Magnesium zinc oxide (MZO) nanostructuremodified QCM (MZOnano-QCM) | E. coli and S. cerevisiae | Ampicillin and tetracycline (E. coli), amphotericin B and miconazole (S. cerevisiae) | 4.8 × 104 to 0.9 × 104 CFU/mL (for ampicillin sensitive E. coli) 6.1 × 104 to 5.9 × 104 CFU/mL (resistant E. coli) | 10 min | Reyes et al. [172] |
| Electrochemical impedance spectroscopy (EIS) | E. coli K-12 | Kanamycin, tetracyclin, erythromycin | 7.1 × 103 CFU/mL | ~20 min | Saucedo et al. [176] |
| Bead-based biosensor via fluorescence imaging | E. coli (ATCC 25922 and 6937) | Ceftazidime, levofloxacin | 5 × 104 CFU/ mL | 60 min | Sabhachandan et al. [179] |
| LSPR | P. aeruginosa, E. coli | Ceftazidime, cefotaxime, ampicillin, amoxicillin, levofloxacin, doxycycline | 0.01 μg/mL in tap water, 0.5 μg/mL in 5% human serum (ceftazidimine). 105 CFU/mL (used bacteria concentration) | 3 h | Nag et al. [178] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Péter, B.; Farkas, E.; Kurunczi, S.; Szittner, Z.; Bősze, S.; Ramsden, J.J.; Szekacs, I.; Horvath, R. Review of Label-Free Monitoring of Bacteria: From Challenging Practical Applications to Basic Research Perspectives. Biosensors 2022, 12, 188. https://doi.org/10.3390/bios12040188
Péter B, Farkas E, Kurunczi S, Szittner Z, Bősze S, Ramsden JJ, Szekacs I, Horvath R. Review of Label-Free Monitoring of Bacteria: From Challenging Practical Applications to Basic Research Perspectives. Biosensors. 2022; 12(4):188. https://doi.org/10.3390/bios12040188
Chicago/Turabian StylePéter, Beatrix, Eniko Farkas, Sandor Kurunczi, Zoltán Szittner, Szilvia Bősze, Jeremy J. Ramsden, Inna Szekacs, and Robert Horvath. 2022. "Review of Label-Free Monitoring of Bacteria: From Challenging Practical Applications to Basic Research Perspectives" Biosensors 12, no. 4: 188. https://doi.org/10.3390/bios12040188