High Throughput Screening of Transcription Factor LysG for Constructing a Better Lysine Biosensor

Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Enzymes and Chemicals
2.2. Cultivation Media
2.3. Plasmid Construction
2.4. Strain Construction
2.5. Screening of LysG Mutant from Random Mutation Library
2.6. Screening Lysine Overproducers from Mutant Libraries
2.7. Cell Growth and Lysine Fermentation Analysis
3. Results and Discussion
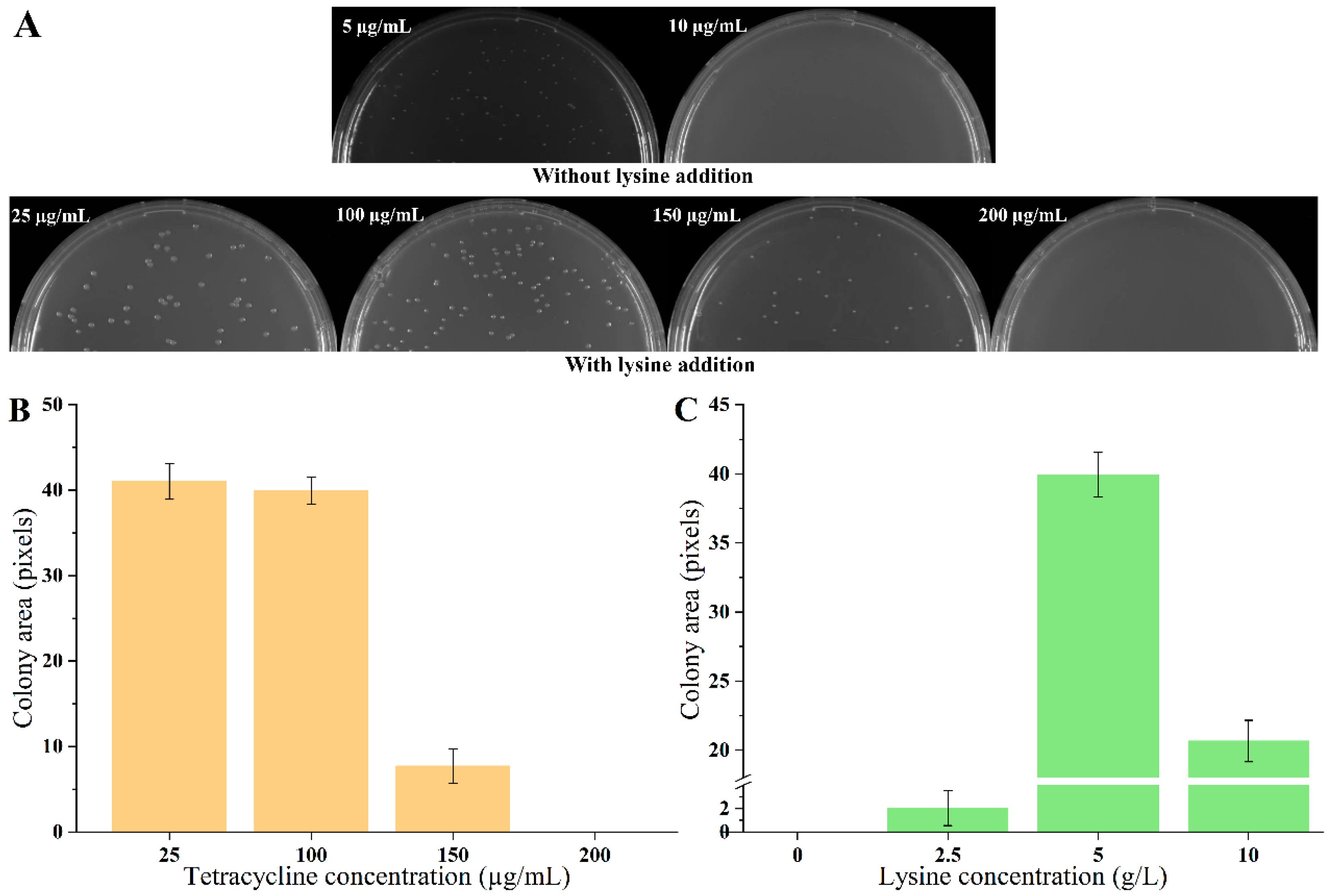
3.1. Design and Proof-of-Concept of the Biosensor Based on LysG
3.2. Optimization of the Biosensor through Screening LysG Mutant
3.3. Endogenous Lysine Overproduction Enhanced Growth of the Biosensor Strains
3.4. High Throughput Screening of Lysine Overproducers from Random Mutation Libraries
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Li, Q.; Liu, C.; He, J.; Liu, T.; Zhang, W.; Xie, Z.; Zong, J.; Li, Y.; Sun, X.; Lu, F. Construction and application of 3-fucosyllactose whole-cell biosensor for high-throughput screening of overproducers. Bioresour. Technol. 2024, 402, 130798. [Google Scholar] [CrossRef]
- Seok, J.Y.; Han, Y.H.; Yang, J.S.; Yang, J.N.; Lim, H.G.; Kim, S.G.; Seo, S.W.; Jung, G.Y. Synthetic biosensor accelerates evolution by rewiring carbon metabolism toward a specific metabolite. Cell Rep. 2021, 36, 109589. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Li, Q.; Wang, Y.; Zhou, W.; Guo, Y.; Chen, J.; Zheng, P.; Sun, J.; Ma, Y. Isoleucyl-tRNA synthetase mutant based whole-cell biosensor for high-throughput selection of isoleucine overproducers. Biosens. Bioelectron. 2021, 172, 112783. [Google Scholar] [CrossRef] [PubMed]
- Niu, F.-X.; He, X.; Huang, Y.-B.; Liu, J.-Z. Biosensor-guided atmospheric and room-temperature plasma mutagenesis and shuffling for high-level production of shikimic acid from sucrose in Escherichia coli. J. Agric. Food Chem. 2020, 68, 11765–11773. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Pang, Q.X.; Wang, Q.; Qi, Q.S.; Wang, Q. Modular tuning engineering and versatile applications of genetically encoded biosensors. Crit. Rev. Biotechnol. 2022, 42, 1010–1027. [Google Scholar] [CrossRef]
- Gu, M.B.; Mitchell, R.J.; Kim, B.C. Whole-cell-based biosensors for environmental biomonitoring and application. Adv. Biochem. Eng. Biot. 2004, 87, 269–305. [Google Scholar]
- Liu, Y.; Li, Q.; Zheng, P.; Zhang, Z.; Liu, Y.; Sun, C.; Cao, G.; Zhou, W.; Wang, X.; Zhang, D.; et al. Developing a high-throughput screening method for threonine overproduction based on an artificial promoter. Microb. Cell Fact. 2015, 14, 121. [Google Scholar] [CrossRef] [PubMed]
- Hossain, G.S.; Saini, M.; Miyake, R.; Ling, H.; Chang, M.W. Genetic biosensor design for natural product biosynthesis in microorganisms. Trends Biotechnol. 2020, 38, 797–810. [Google Scholar] [CrossRef]
- Mahr, R.; Gätgens, C.; Gätgens, J.; Polen, T.; Kalinowski, J.; Frunzke, J. Biosensor-driven adaptive laboratory evolution of L-valine production in Corynebacterium glutamicum. Metab. Eng. 2015, 32, 184–194. [Google Scholar] [CrossRef]
- Mahr, R.; Frunzke, J. Transcription factor-based biosensors in biotechnology: Current state and future prospects. Appl. Microbiol. Biot. 2016, 100, 79–90. [Google Scholar] [CrossRef]
- Binder, S.; Siedler, S.; Marienhagen, J.; Bott, M.; Eggeling, L. Recombineering in Corynebacterium glutamicum combined with optical nanosensors: A general strategy for fast producer strain generation. Nucleic Acids Res. 2013, 41, 6360–6369. [Google Scholar] [CrossRef] [PubMed]
- Pu, W.; Chen, J.Z.; Liu, P.; Shen, J.; Cai, N.Y.; Liu, B.Y.; Lei, Y.; Wang, L.X.; Ni, X.M.; Zhang, J.; et al. Directed evolution of linker helix as an efficient strategy for engineering LysR-type transcriptional regulators as whole-cell biosensors. Biosens. Bioelectron. 2023, 222, 115004. [Google Scholar] [CrossRef] [PubMed]
- Della Corte, D.; van Beek, H.L.; Syberg, F.; Schallmey, M.; Tobola, F.; Cormann, K.U.; Schlicker, C.; Baumann, P.T.; Krumbach, K.; Sokolowsky, S.; et al. Engineering and application of a biosensor with focused ligand specificity. Nat. Commun. 2020, 11, 4851. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Teng, J.; Yao, L.; Xu, J.; Liu, G. Selection of specific DNA aptamers for hetero-sandwich-based colorimetric determination of Campylobacter jejuni in food. J. Agric. Food Chem. 2020, 68, 8455–8461. [Google Scholar] [CrossRef]
- Pang, Q.X.; Han, H.; Liu, X.Q.; Wang, Z.G.; Liang, Q.F.; Hou, J.; Qi, Q.S.; Wang, Q. In vivo evolutionary engineering of riboswitch with high-threshold for N-acetylneuraminic acid production. Metab. Eng. 2020, 59, 36–43. [Google Scholar] [CrossRef]
- Wang, X.; Li, Q.; Sun, C.; Cai, Z.; Zheng, X.; Guo, X.; Ni, X.; Zhou, W.; Guo, Y.; Zheng, P.; et al. GREACE-assisted adaptive laboratory evolution in endpoint fermentation broth enhances lysine production by Escherichia coli. Microb. Cell Fact. 2019, 18, 106. [Google Scholar] [CrossRef]
- Bellmann, A.; Vrljic, M.; Pátek, M.; Sahm, H.; Krämer, R.; Eggeling, L. Expression control and specificity of the basic amino acid exporter LysE of Corynebacterium glutamicum. Microbiology 2001, 147, 1765–1774. [Google Scholar] [CrossRef]
- Binder, S.; Schendzielorz, G.; Stäbler, N.; Krumbach, K.; Hoffmann, K.; Bott, M.; Eggeling, L. A high-throughput approach to identify genomic variants of bacterial metabolite producers at the single-cell level. Genome Biol. 2012, 13, R40. [Google Scholar] [CrossRef]
- Sun, X.; Peng, Z.; Li, C.; Zheng, Y.; Cheng, Y.; Zong, J.; Lu, F.; Li, Y.; Li, Q. Combinatorial metabolic engineering and tolerance evolving of Escherichia coli for high production of 2′-fucosyllactose. Bioresour. Technol. 2023, 372, 128667. [Google Scholar] [CrossRef]
- Falco, S.C.; Guida, T.; Locke, M.; Mauvais, J.; Sanders, C.; Ward, R.T.; Webber, P. Transgenic canola and soybean seeds with increased lysine. Bio/Technology 1995, 13, 577–582. [Google Scholar] [CrossRef]
- Patte, J.C.; Loviny, T.; Cohen, G.N. Repression of meso-alpha, epsilon-diaminopimelic acid decarboxylase by L-lysine in Escherichia coli. Biochim. Biophys. Acta 1962, 58, 359–360. [Google Scholar] [CrossRef] [PubMed]
- Zhao, D.; Feng, X.; Zhu, X.; Wu, T.; Zhang, X.; Bi, C. CRISPR/Cas9-assisted gRNA-free one-step genome editing with no sequence limitations and improved targeting efficiency. Sci. Rep. 2017, 7, 16624. [Google Scholar] [CrossRef]
- Gioia, M.G.; Andreatta, P.; Boschetti, S.; Gatti, R. Development and validation of a liquid chromatographic method for the determination of branched-chain amino acids in new dosage forms. J. Pharm. Biomed. 2007, 45, 456–464. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Li, Q.; Zheng, P.; Guo, Y.; Wang, L.; Zhang, T.; Sun, J.; Ma, Y. Evolving the L-lysine high-producing strain of Escherichia coli using a newly developed high-throughput screening method. J. Ind. Microbiol. Biotechnol. 2016, 43, 1227–1235. [Google Scholar] [CrossRef] [PubMed]
- Mustafi, N.; Grünberger, A.; Kohlheyer, D.; Bott, M.; Frunzke, J. The development and application of a single-cell biosensor for the detection of L-methionine and branched-chain amino acids. Metab. Eng. 2012, 14, 449–457. [Google Scholar] [CrossRef] [PubMed]
- Møller, T.S.B.; Overgaard, M.; Nielsen, S.S.; Bortolaia, V.; Sommer, M.O.A.; Guardabassi, L.; Olsen, J.E. Relation between tetR and tetA expression in tetracycline resistant Escherichia coli. BMC Microbiol. 2016, 16, 39. [Google Scholar] [CrossRef]
- Qiu, C.; Huang, M.; Hou, Y.; Tao, H.; Zhao, J.; Shen, Y.; Bao, X.; Qi, Q.; Hou, J. Biosensor-coupled in vivo mutagenesis and omics analysis reveals reduced lysine and arginine synthesis to improve Malonyl-coenzyme a flux in Saccharomyces cerevisiae. mSystems 2022, 7, e01366-21. [Google Scholar] [CrossRef]
- Dykxhoorn, D.M.; Pierre, R.S.; Linn, T. A set of compatible tac promoter expression vectors. Gene 1996, 177, 133–136. [Google Scholar] [CrossRef]
- Arfman, N.; Worrell, V.; Ingram, L.O. Use of the tac promoter and lacIq for the controlled expression of Zymomonas mobilis fermentative genes in Escherichia coli and Zymomonas mobilis. J. Bacteriol. 1992, 174, 7370–7378. [Google Scholar] [CrossRef]
- Marbaniang, C.N.; Gowrishankar, J. Transcriptional cross-regulation between gram-negative and gram-positive bacteria, demonstrated using ArgP-argO of Escherichia coli and LysG-lysE of Corynebacterium glutamicum. J. Bacteriol. 2012, 194, 5657–5666. [Google Scholar] [CrossRef]
- Deng, C.; Wu, Y.K.; Lv, X.Q.; Li, J.H.; Liu, Y.F.; Du, G.C.; Chen, J.; Liu, L. Refactoring transcription factors for metabolic engineering. Biotechnol. Adv. 2022, 57, 107935. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Liu, Y.; Wang, M. Design, optimization and application of small molecule biosensor in metabolic engineering. Front. Microbiol. 2017, 8, 2012. [Google Scholar] [CrossRef]
- Li, X.; Ren, S.; Song, G.; Liu, Y.; Li, Y.; Lu, F. Novel detection method for evaluating the activity of an alkaline serine protease from Bacillus clausii. J. Agric. Food Chem. 2022, 70, 3765–3774. [Google Scholar] [CrossRef]
- Dumas, R.; Cobessi, D.; Robin, A.Y.; Ferrer, J.-L.; Curien, G. The many faces of aspartate kinases. Arch. Biochem. Biophys. 2012, 519, 186–193. [Google Scholar] [CrossRef]
- Ogawa-Miyata, Y.; Kojima, H.; Sano, K. Mutation analysis of the feedback inhibition site of aspartokinase III of Escherichia coli K-12 and its use in L-threonine production. Biosci. Biotech. Biochem. 2001, 65, 1149–1154. [Google Scholar] [CrossRef] [PubMed]
- Eggeling, L.; Oberle, S.; Sahm, H. Improved L-lysine yield with Corynebacterium glutamicum: Use of dapA resulting in increased flux combined with growth limitation. Appl. Microbiol. Biot. 1998, 49, 24–30. [Google Scholar] [CrossRef]
- Hudson, A.O.; Gilvarg, C.; Leustek, T. Biochemical and phylogenetic characterization of a novel diaminopimelate biosynthesis pathway in prokaryotes identifies a diverged form of LL-diaminopimelate aminotransferase. J. Bacteriol. 2008, 190, 3256–3263. [Google Scholar] [CrossRef] [PubMed]
- Schrumpf, B.; Schwarzer, A.; Kalinowski, J.; Pühler, A.; Eggeling, L.; Sahm, H. A functionally split pathway for lysine synthesis in Corynebacterium glutamicum. J. Bacteriol. 1991, 173, 4510–4516. [Google Scholar] [CrossRef]
- Zhou, S.H.; Yuan, S.F.; Nair, P.H.; Alper, H.S.; Deng, Y.; Zhou, J.W. Development of a growth coupled and multi-layered dynamic regulation network balancing malonyl-CoA node to enhance (2S)-naringenin biosynthesis in Escherichia coli. Metab. Eng. 2021, 67, 41–52. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Q.; Ren, H.; Liao, Z.; Xia, S.; Sun, X. High Throughput Screening of Transcription Factor LysG for Constructing a Better Lysine Biosensor. Biosensors 2024, 14, 455. https://doi.org/10.3390/bios14100455
Li Q, Ren H, Liao Z, Xia S, Sun X. High Throughput Screening of Transcription Factor LysG for Constructing a Better Lysine Biosensor. Biosensors. 2024; 14(10):455. https://doi.org/10.3390/bios14100455
Chicago/Turabian StyleLi, Qinggang, Haojie Ren, Zhenjiang Liao, Shuchang Xia, and Xue Sun. 2024. "High Throughput Screening of Transcription Factor LysG for Constructing a Better Lysine Biosensor" Biosensors 14, no. 10: 455. https://doi.org/10.3390/bios14100455
APA StyleLi, Q., Ren, H., Liao, Z., Xia, S., & Sun, X. (2024). High Throughput Screening of Transcription Factor LysG for Constructing a Better Lysine Biosensor. Biosensors, 14(10), 455. https://doi.org/10.3390/bios14100455