
Measurement of Rapid Amiloride-Dependent pH Changes at the Cell Surface Using a Proton-Sensitive Field-Effect Transistor


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
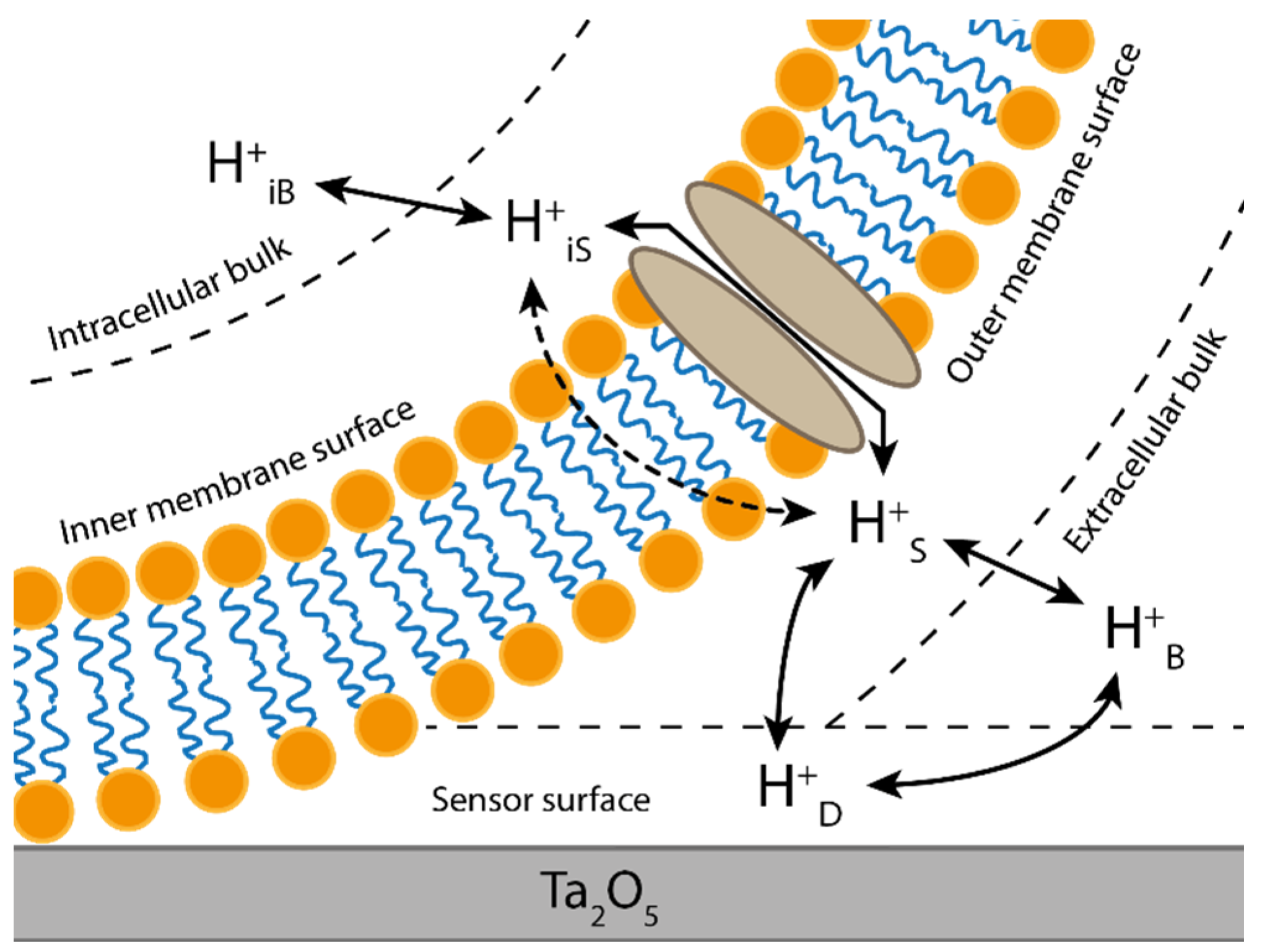
:1. Introduction
2. Materials and Methods
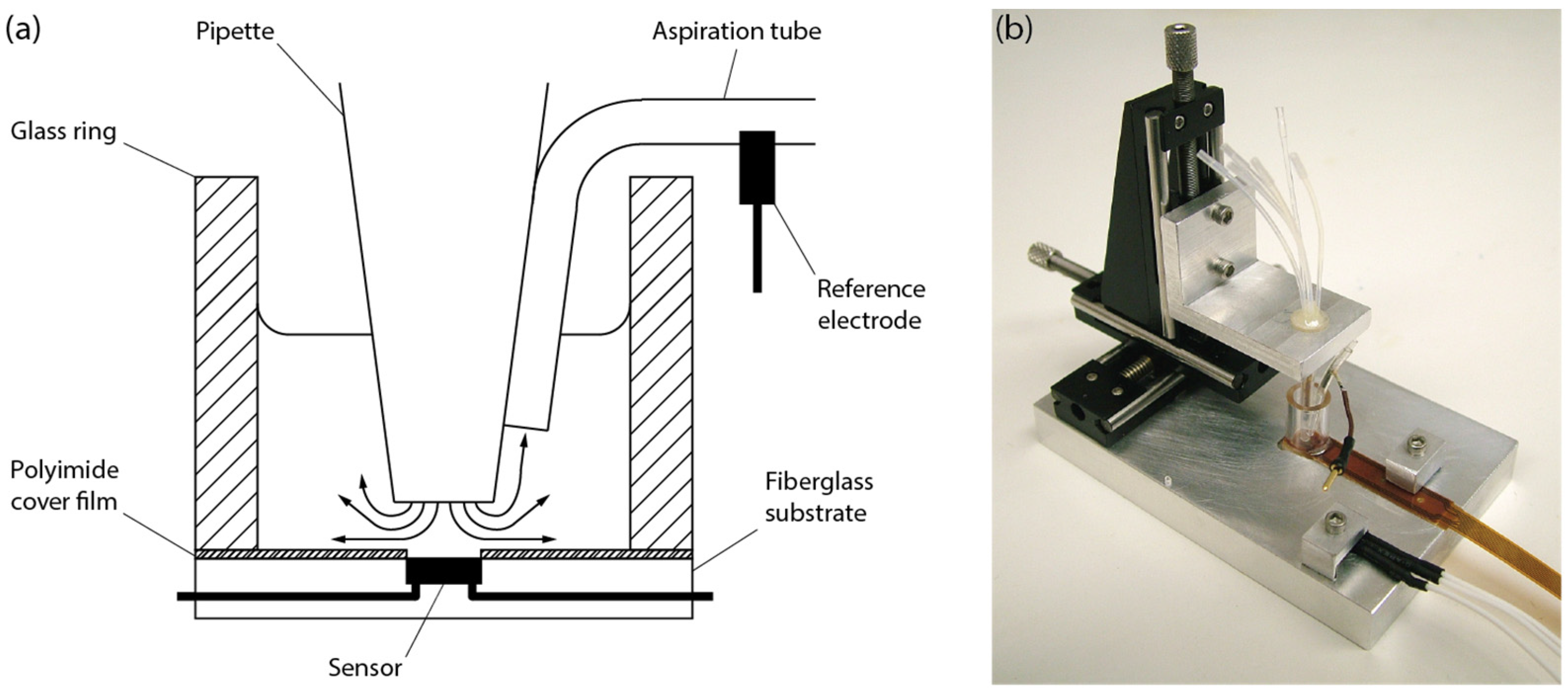
2.1. System Setup
2.2. Cell Culture
2.3. Solutions and Reagents
2.4. Sample Preparation and Experimental Procedure
3. Results and Discussion
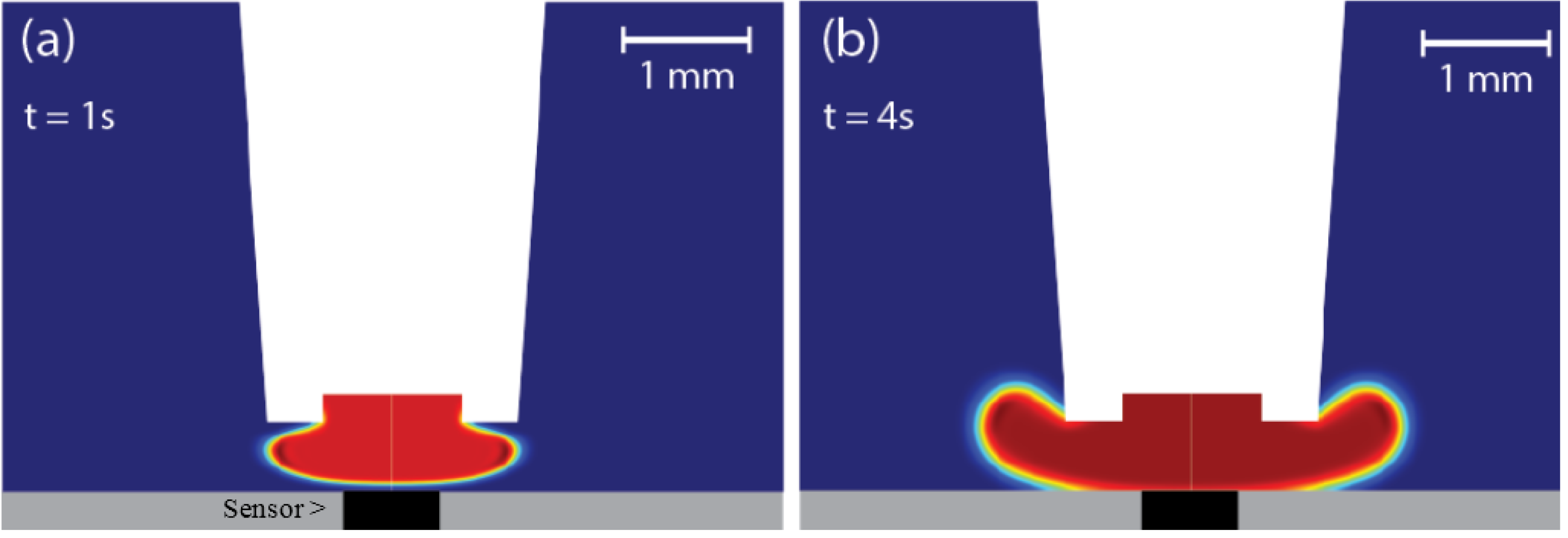
3.1. Optimization of the Superfusion
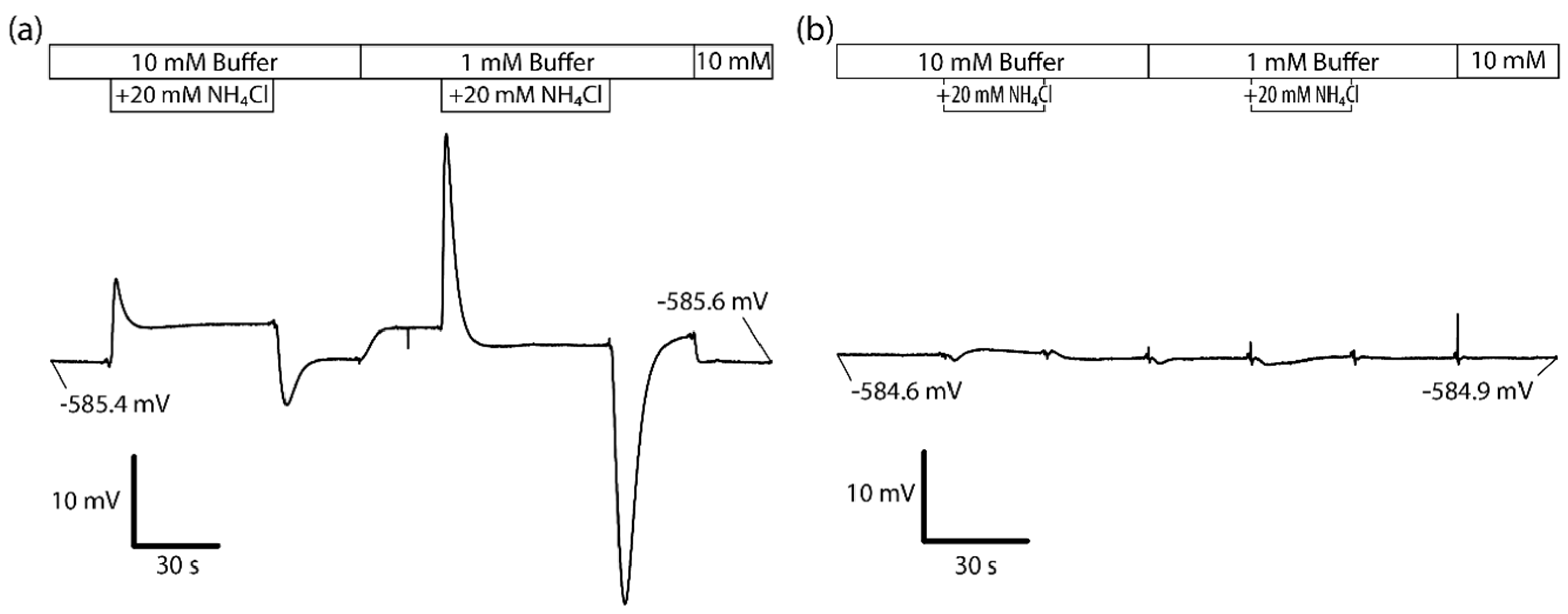
3.2. Ammonia Loading and Unloading
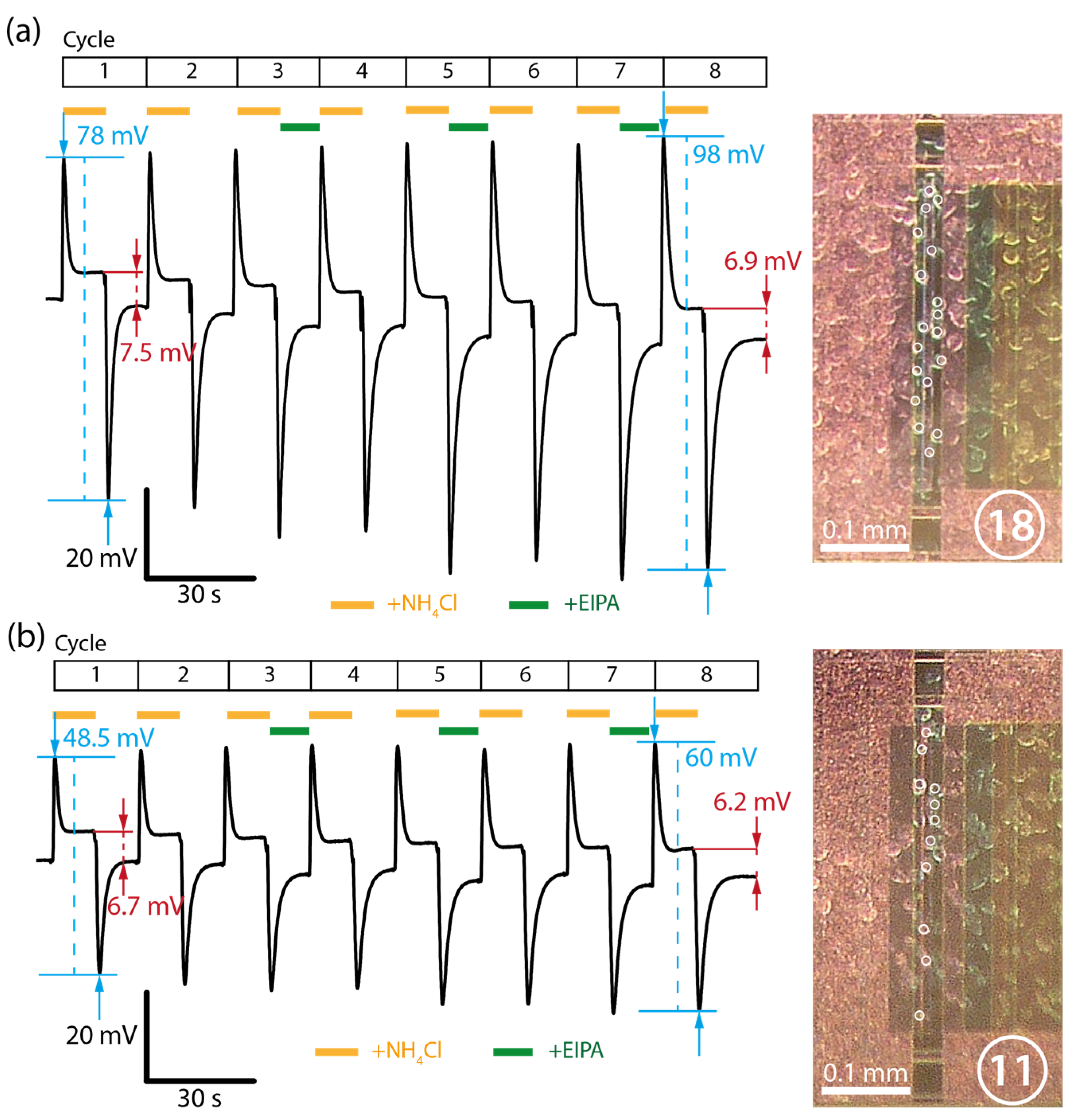
3.3. Repeated Ammonia Cycles and Dependence on Cell Density
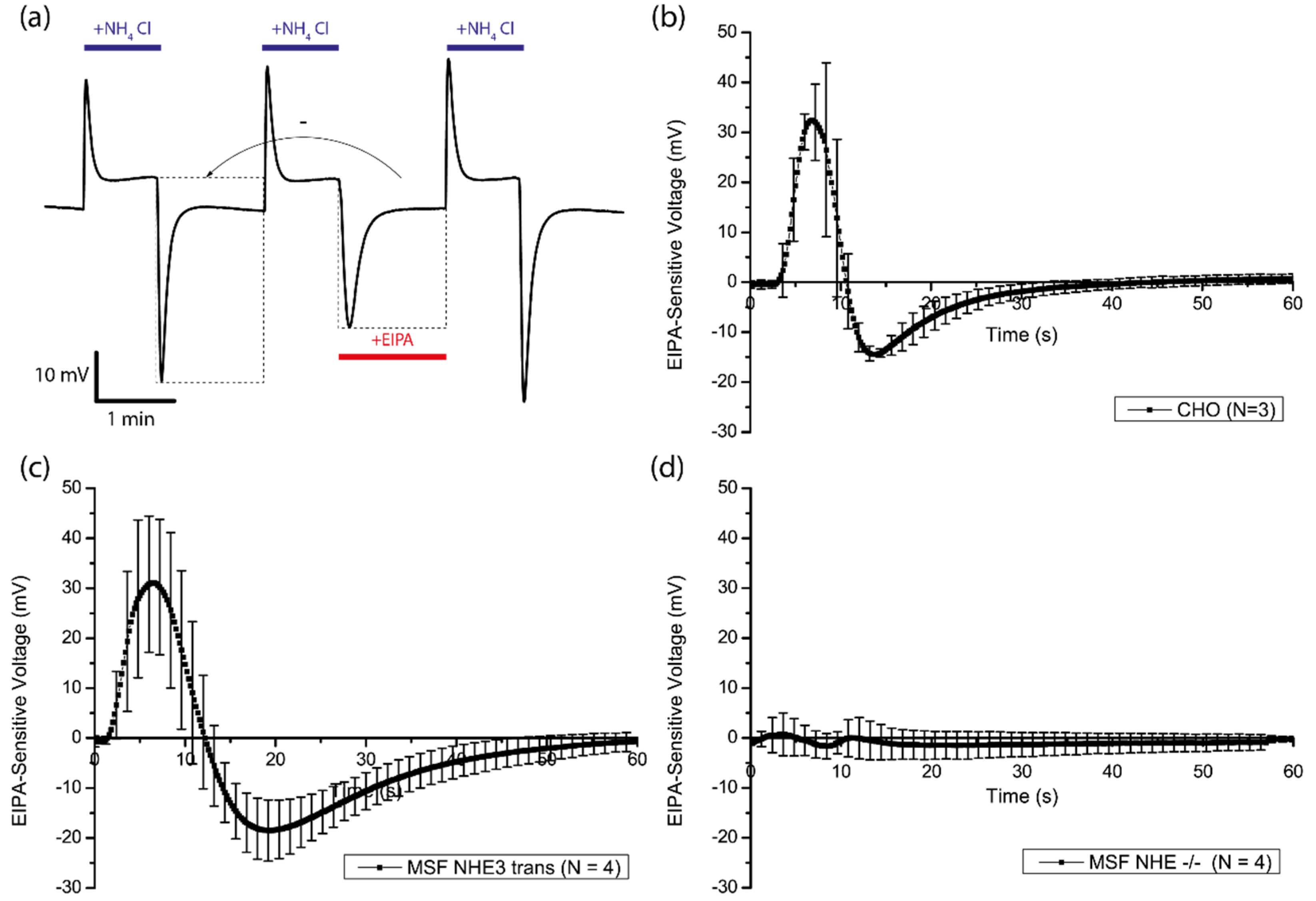
3.4. Amiloride-Sensitivity
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Mueckler, M.; Thorens, B. The SLC2 (GLUT) family of membrane transporters. ABCs Membr. Transp. Health Dis. SLC Ser. 2013, 34, 121–138. [Google Scholar]
- Orlowski, J.; Grinstein, S. Diversity of the mammalian sodium/proton exchanger SLC9 gene family. Pflüg. Arch. 2003, 447, 549–565. [Google Scholar]
- Fuster, D.; Moe, O.W.; Hilgemann, D.W. Lipid- and mechanosensitivities of sodium/hydrogen exchangers analyzed by electrical methods. Proc. Natl. Acad. Sci. USA 2004, 101, 10482–10487. [Google Scholar] [PubMed]
- Alexiev, U.; Mollaaghababa, R.; Scherrer, P.; Khorana, H.G.; Heyn, M.P. Rapid long-range proton diffusion along the surface of the purple membrane and delayed proton transfer into the bulk. Proc. Natl. Acad. Sci. USA 1995, 92, 372–376. [Google Scholar] [CrossRef] [PubMed]
- Antonenko, Y.N.; Pohl, P. Coupling of proton source and sink via H+-migration along the membrane surface as revealed by double patch-clamp experiments. FEBS Lett. 1998, 429, 197–200. [Google Scholar] [CrossRef]
- Medvedev, E.S.; Stuchebrukhov, A.A. Proton diffusion along biological membranes. J. Phys. Condens. Matter 2011, 23. [Google Scholar] [CrossRef] [PubMed]
- Schaffhauser, D.F.; Patti, M.; Goda, T.; Miyahara, Y.; Forster, I.C.; Dittrich, P.S. An Integrated Field-Effect Microdevice for Monitoring Membrane Transport in Xenopus laevis Oocytes via Lateral Proton Diffusion. PLoS ONE 2012, 7, e39238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Hal, R.E.G.; Eijkel, J.C.T.; Bergveld, P. A novel description of ISFET sensitivity with the buffer capacity and double-layer capacitance as key parameters. Sens. Actuators B Chem. 1995, 24, 201–205. [Google Scholar] [CrossRef]
- Lehmann, M.; Baumann, W.; Brischwein, M.; Ehret, R.; Kraus, M.; Schwinde, A.; Bitzenhofer, M.; Freund, I.; Wolf, B. Non-invasive measurement of cell membrane associated proton gradients by ion-sensitive field effect transistor arrays for microphysiological and bioelectronical applications. Biosens. Bioelectron. 2000, 15, 117–124. [Google Scholar] [CrossRef]
- Antonenko, Y.N.; Pohl, P.; Denisov, G.A. Permeation of ammonia across bilayer lipid membranes studied by ammonium ion selective microelectrodes. Biophys. J. 1997, 72, 2187–2195. [Google Scholar] [CrossRef]
- Mahnensmith, R.L.; Aronson, P.S. The plasma membrane sodium-hydrogen exchanger and its role in physiological and pathophysiological processes. Circ. Res. 1985, 56, 773–788. [Google Scholar] [CrossRef] [PubMed]
- Fuster, D.; Moe, O.W.; Hilgemann, D.W. Steady-state Function of the Ubiquitous Mammalian Na/H Exchanger (NHE1) in Relation to Dimer Coupling Models with 2Na/2H Stoichiometry. J. Gen. Physiol. 2008, 132, 465–480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bobulescu, I.A.; Di Sole, F.; Moe, O.W. Na+/H+ exchangers: Physiology and link to hypertension and organ ischemia. Curr. Opin. Nephrol. Hypertens. 2005, 14, 485–494. [Google Scholar] [CrossRef] [PubMed]
- Karmazyn, M. Amiloride enhances postischemic ventricular recovery: Possible role of Na+-H+ exchange. Am. J. Physiol. Heart Circ. Physiol. 1988, 255, H608–H615. [Google Scholar]
- Karmazyn, M.; Moffat, M.P. Role of Na+/H+ exchange in cardiac physiology and pathophysiology: Mediation of myocardial reperfusion injury by the pH paradox. Cardiovasc. Res. 1993, 27, 915–924. [Google Scholar] [CrossRef] [PubMed]
- Hale, S.L.; Kloner, R.A. Effect of combined KATP channel activation and Na+/H+ exchange inhibition on infarct size in rabbits. Am. J. Physiol. Heart Circ. Physiol. 2000, 279, H2673–H2677. [Google Scholar] [PubMed]
- Gumina, R.J.; Moore, J.; Schelling, P.; Beier, N.; Gross, G.J. Na+/H+ exchange inhibition prevents endothelial dysfunction after I/R injury. Am. J. Physiol. Heart Circ. Physiol. 2001, 281, H1260–H1266. [Google Scholar] [PubMed]
- Masereel, B.; Pochet, L.; Laeckmann, D. An overview of inhibitors of Na+/H+ exchanger. Eur. J. Med. Chem. 2003, 38, 547–554. [Google Scholar] [CrossRef]
- Casans, S.; Ramirez, D.; Navarro, A. Circuit provides constant current ISFETs/MEMFETs. EDN 2000, 45, 164. [Google Scholar]
- Schneider, M.; Marison, I.W.; von Stockar, U. The importance of ammonia in mammalian cell culture. J. Biotechnol. 1996, 46, 161–185. [Google Scholar] [CrossRef]
- Nagami, G.T. Luminal secretion of ammonia in the mouse proximal tubule perfused in vitro. J. Clin. Investig. 1988, 81, 159–164. [Google Scholar] [CrossRef] [PubMed]
- Weiner, I.D.; Hamm, L.L. Molecular Mechanisms of Renal Ammonia Transport. Annu. Rev. Physiol. 2007, 69, 317–340. [Google Scholar] [CrossRef] [PubMed]
- Weiner, I.D.; Verlander, J.W. Role of NH3 and NH4+ transporters in renal acid-base transport. Am. J. Physiol. Ren. Physiol. 2011, 300, F11–F23. [Google Scholar] [CrossRef] [PubMed]








© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schaffhauser, D.; Fine, M.; Tabata, M.; Goda, T.; Miyahara, Y. Measurement of Rapid Amiloride-Dependent pH Changes at the Cell Surface Using a Proton-Sensitive Field-Effect Transistor. Biosensors 2016, 6, 11. https://doi.org/10.3390/bios6020011
Schaffhauser D, Fine M, Tabata M, Goda T, Miyahara Y. Measurement of Rapid Amiloride-Dependent pH Changes at the Cell Surface Using a Proton-Sensitive Field-Effect Transistor. Biosensors. 2016; 6(2):11. https://doi.org/10.3390/bios6020011
Chicago/Turabian StyleSchaffhauser, Daniel, Michael Fine, Miyuki Tabata, Tatsuro Goda, and Yuji Miyahara. 2016. "Measurement of Rapid Amiloride-Dependent pH Changes at the Cell Surface Using a Proton-Sensitive Field-Effect Transistor" Biosensors 6, no. 2: 11. https://doi.org/10.3390/bios6020011
APA StyleSchaffhauser, D., Fine, M., Tabata, M., Goda, T., & Miyahara, Y. (2016). Measurement of Rapid Amiloride-Dependent pH Changes at the Cell Surface Using a Proton-Sensitive Field-Effect Transistor. Biosensors, 6(2), 11. https://doi.org/10.3390/bios6020011