Identification of Mutations Conferring Tryptanthrin Resistance to Mycobacterium smegmatis

 ,
,
Abstract
:1. Introduction
2. Results
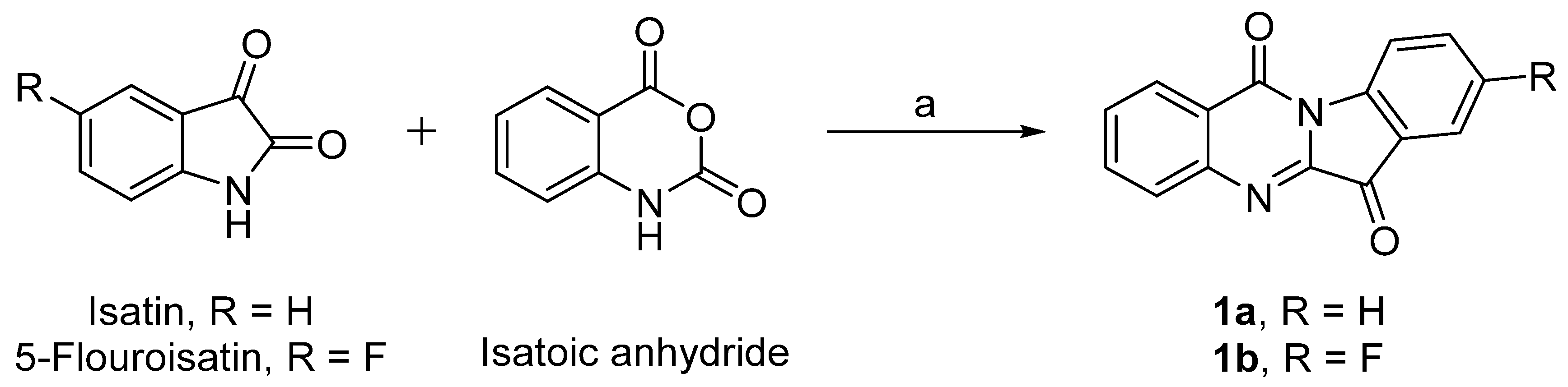
2.1. Synthesis of Tryptanthrin Analogs
2.2. Tryptanthrins May Be Subjected to MmpS5-MmpL5 Mediated Efflux
2.3. Mutations in Transcriptional Regulators and a Transporter Gene May Confer Resistance to Tryptanthrins
3. Discussion
4. Materials and Methods
4.1. Synthetic Procedures of Tryptanthrin Analogs
4.2. Bacterial Strains and Growth Conditions
4.3. MIC Determination
4.4. Generation of Resistant Mutants and Their Phenotype Characterization
4.5. Mycobacterial DNA Extraction
4.6. Whole-Genomic Sequencing
4.7. Whole-Genomic Data Analysis
4.8. Data Availability
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- World Health Organization. World Health Organization Global Tuberculosis Report 2020; World Health Organization: Geneva, Switzerland, 2020; pp. 1–232. [Google Scholar]
- Hameed, H.M.A.; Islam, M.M.; Chhotaray, C.; Wang, C.; Liu, Y.; Tan, Y.; Li, X.; Tan, S.; Delorme, V.; Yew, W.W.; et al. Molecular Targets Related Drug Resistance Mechanisms in MDR-, XDR-, and TDR-Mycobacterium tuberculosis Strains. Front. Cell. Infect. Microbiol. 2018, 8, 114. [Google Scholar] [CrossRef] [PubMed]
- Caminero, J.A.; Sotgiu, G.; Zumla, A.; Migliori, G.B. Best drug treatment for multidrug-resistant and extensively drug-resistant tuberculosis. Lancet Infect. Dis. 2010, 10, 621–629. [Google Scholar] [CrossRef]
- Gandhi, N.R.; Nunn, P.; Dheda, K.; Schaaf, H.S.; Zignol, M.; van Soolingen, D.; Jensen, P.; Bayona, J. Multidrug-resistant and extensively drug-resistant tuberculosis: A threat to global control of tuberculosis. Lancet 2010, 375, 1830–1843. [Google Scholar] [CrossRef]
- Klopper, M.; Warren, R.M.; Hayes, C.; van Pittius, N.C.G.; Streicher, E.M.; Müller, B.; Sirgel, F.A.; Chabula-Nxiweni, M.; Hoosain, E.; Coetzee, G.; et al. Emergence and Spread of Extensively and Totally Drug-Resistant Tuberculosis, South Africa. Emerg. Infect. Dis. 2013, 19, 449–455. [Google Scholar] [CrossRef] [Green Version]
- Velayati, A.A.; Farnia, P.; Farahbod, A.M. Overview of drug-resistant tuberculosis worldwide. Int. J. Mycobacteriol. 2016, 5 (Suppl. 1), S161. [Google Scholar] [CrossRef] [Green Version]
- Honda, G.; Tabata, M. Isolation of antifungal principle tryptanthrin, from Strobilanthes cusia O. Kuntze. Planta Med. 1979, 36, 85–90. [Google Scholar] [CrossRef]
- Honda, G.; Tosirisuk, V.; Tabata, M. Isolation of an antidermatophytic, tryptanthrin, from indigo plants, Polygonum tinctorium and Isatis tinctoria. Planta Med. 1980, 38, 275–276. [Google Scholar] [CrossRef]
- Scovill, J.; Blank, E.; Konnick, M.; Nenortas, E.; Shapiro, T. Antitrypanosomal activities of tryptanthrins. Antimicrob. Agents Chemother. 2002, 46, 882–883. [Google Scholar] [CrossRef] [Green Version]
- Kimoto, T.; Hino, K.; Koya-Miyata, S.; Yamamoto, Y.; Takeuchi, M.; Nishizaki, Y.; Micallef, M.J.; Ushio, S.; Iwaki, K.; Ikeda, M.; et al. Cell differentiation and apoptosis of monocytic and promyelocytic leukemia cells (U-937 and HL-60) by tryptanthrin, an active ingredient of Polygonum tinctorium Lour. Pathol. Int. 2001, 51, 315–325. [Google Scholar] [CrossRef]
- Sharma, V.M.; Prasanna, P.; Seshu, K.V.A.; Renuka, B.; Rao, C.V.L.; Kumar, G.S.; Narasimhulu, C.P.; Babu, P.A.; Puranik, R.C.; Subramanyam, D.; et al. Novel indolo[2,1-b]quinazoline analogues as cytostatic agents: Synthesis, biological evaluation and structure-activity relationship. Bioorg. Med. Chem. Lett. 2002, 12, 2303–2307. [Google Scholar] [CrossRef]
- Koya-Miyata, S.; Kimoto, T.; Micallef, M.J.; Hino, K.; Taniguchi, M.; Ushio, S.; Iwaki, K.; Ikeda, M.; Kurimoto, M. Prevention of azoxymethane-induced intestinal tumors by a crude ethyl acetate-extract and tryptanthrin extracted from Polygonum tinctorium Lour. Anticancer Res. 2001, 21, 3295–3300. [Google Scholar] [PubMed]
- Yang, S.; Li, X.; Hu, F.; Li, Y.; Yang, Y.; Yan, J.; Kuang, C.; Yang, Q. Discovery of tryptanthrin derivatives as potent inhibitors of indoleamine 2,3-dioxygenase with therapeutic activity in Lewis lung cancer (LLC) tumor-bearing mice. J. Med. Chem. 2013, 56, 8321–8331. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.; Zhang, S.; Fang, X.; Guo, L.; Hu, N.; Guo, Z.; Li, X.; Yang, S.; He, J.C.; Kuang, C.; et al. N-Benzyl/Aryl Substituted Tryptanthrin as Dual Inhibitors of Indoleamine 2,3-Dioxygenase and Tryptophan 2,3-Dioxygenase. J. Med. Chem. 2019, 62, 9161–9174. [Google Scholar] [CrossRef]
- Micallef, M.J.; Iwaki, K.; Ishihara, T.; Ushio, S.; Aga, M.; Kunikata, T.; Koya-Miyata, S.; Kimoto, T.; Ikeda, M.; Kurimoto, M. The natural plant product tryptanthrin ameliorates dextran sodium sulfate-induced colitis in mice. Int. Immunopharmacol. 2002, 2, 565–578. [Google Scholar] [CrossRef]
- Takei, Y.; Kunikata, T.; Aga, M.; Inoue, S.-I.; Ushio, S.; Iwaki, K.; Ikeda, M.; Kurimoto, M. Tryptanthrin inhibits interferon-gamma production by Peyer’s patch lymphocytes derived from mice that had been orally administered staphylococcal enterotoxin. Biol. Pharm. Bull. 2003, 26, 365–367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iwaki, K.; Ohashi, E.; Arai, N.; Kohno, K.; Ushio, S.; Taniguchi, M.; Fukuda, S. Tryptanthrin inhibits Th2 development, and IgE-mediated degranulation and IL-4 production by rat basophilic leukemia RBL-2H3 cells. J. Ethnopharmacol. 2011, 134, 450–459. [Google Scholar] [CrossRef]
- Mitscher, L.A.; Baker, W. Tuberculosis: A search for novel therapy starting with natural products. Med. Res. Rev. 1998, 18, 363–374. [Google Scholar] [CrossRef]
- Tripathi, A.; Wadia, N.; Bindal, D.; Jana, T. Docking Studies on Novel Alkaloid Tryptanthrin and Its Analogues against Enoyl-acyl Carrier Protein Reductase (InhA) of Mycobacterium tuberculosis; NISCAIR-CSIR: New Delhi, India, 2012. [Google Scholar]
- Banerjee, A.; Dubnau, E.; Quemard, A.; Balasubramanian, V.; Um, K.S.; Wilson, T.; Collins, D.; de Lisle, G.; Jacobs, W.R. inhA, a gene encoding a target for isoniazid and ethionamide in Mycobacterium tuberculosis. Science 1994, 263, 227–230. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Heym, B.; Allen, B.; Young, D.; Cole, S. The catalase-peroxidase gene and isoniazid resistance of Mycobacterium tuberculosis. Nature 1992, 358, 591–593. [Google Scholar] [CrossRef]
- Seifert, M.; Catanzaro, D.; Catanzaro, A.; Rodwell, T.C. Genetic Mutations Associated with Isoniazid Resistance in Mycobacterium tuberculosis: A Systematic Review. PLoS ONE 2015, 10, e0119628. [Google Scholar] [CrossRef] [Green Version]
- Kaur, R.; Manjal, S.K.; Rawal, R.K.; Kumar, K. Recent synthetic and medicinal perspectives of tryptanthrin. Bioorg. Med. Chem. 2017, 25, 4533–4552. [Google Scholar] [CrossRef] [PubMed]
- Baker, W.R.; Mitscher, L.A.; Corp, P. Indolo[2,1-Biquinazoline-6,12-dione Antibacterial Compounds and Methods of Use Thereof. U.S. Patent 5,441,955, 15 August 1995. [Google Scholar]
- Milano, A.; Pasca, M.R.; Provvedi, R.; Lucarelli, A.P.; Manina, G.; de Ribeiro, A.L.J.L.; Manganelli, R.; Riccardi, G. Azole resistance in Mycobacterium tuberculosis is mediated by the MmpS5-MmpL5 efflux system. Tuberculosis 2009, 89, 84–90. [Google Scholar] [CrossRef] [PubMed]
- Halloum, I.; Viljoen, A.; Khanna, V.; Craig, D.; Bouchier, C.; Brosch, R.; Coxon, G.; Kremer, L. Resistance to Thiacetazone Derivatives Active against Mycobacterium abscessus Involves Mutations in the MmpL5 Transcriptional Repressor MAB_4384. Antimicrob. Agents Chemother. 2017, 61, NP246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maslov, D.A.; Shur, K.V.; Vatlin, A.A.; Danilenko, V.N. MmpS5-MmpL5 Transporters Provide Mycobacterium smegmatis Resistance to imidazo[1,2-b][1,2,4,5]tetrazines. Pathogens 2020, 9, 166. [Google Scholar] [CrossRef] [Green Version]
- Chawla, K.; Martinez, E.; Kumar, A.; Shenoy, V.P.; Sintchenko, V. Whole-genome sequencing reveals genetic signature of bedaquiline resistance in a clinical isolate of Mycobacterium tuberculosis. J. Glob. Antimicrob. Resist. 2018, 15, 103–104. [Google Scholar] [CrossRef]
- Shur, K.V.; Frolova, S.G.; Akimova, N.I.; Danilenko, V.N.; Maslov, D.A. A Test-System for in vitro Screening Antimycobacterial Drug Candidates for MmpS5-MmpL5 Mediated Drug Resistance. Russ. J. Genet. 2021, 57, 114–116, In Press. [Google Scholar]
- Sharma, K.; Gupta, M.; Krupa, A.; Srinivasan, N.; Singh, Y. EmbR, a regulatory protein with ATPase activity, is a substrate of multiple serine/threonine kinases and phosphatase in Mycobacterium tuberculosis. FEBS J. 2006, 273, 2711–2721. [Google Scholar] [CrossRef]
- Li, G.; Zhang, J.; Guo, Q.; Jiang, Y.; Wei, J.; Zhao, L.-L.; Zhao, X.; Lu, J.; Wan, K. Efflux pump gene expression in multidrug-resistant Mycobacterium tuberculosis clinical isolates. PLoS ONE 2015, 10, e0119013. [Google Scholar] [CrossRef] [Green Version]
- Narang, A.; Giri, A.; Gupta, S.; Garima, K.; Bose, M.; Varma-Basil, M. Contribution of putative efflux pump genes to isoniazid resistance in clinical isolates of Mycobacterium tuberculosis. Int. J. Mycobacteriol. 2017, 6, 177–183. [Google Scholar]
- Ghajavand, H.; Kamakoli, M.K.; Khanipour, S.; Dizaji, S.P.; Masoumi, M.; Jamnani, F.R.; Fateh, A.; Yaseri, M.; Siadat, S.D.; Vaziri, F. Scrutinizing the drug resistance mechanism of multi- and extensively-drug resistant Mycobacterium tuberculosis: Mutations versus efflux pumps. Antimicrob. Resist. Infect. Control. 2019, 8, 70–78. [Google Scholar] [CrossRef]
- AlMatar, M.; Var, I.; Kayar, B.; Köksal, F. Differential Expression of Resistant and Efflux Pump Genes in MDR-TB Isolates. Endocr. Metab. Immune Disord. Drug Targets 2020, 20, 271–287. [Google Scholar] [CrossRef] [PubMed]
- Umar, F.; Hatta, M.; Husain, D.R.; Natzir, R.; Dwiyanti, R.; Junita, A.R.; Primaguna, M.R. The effect of anti-tuberculosis drugs therapy on mRNA efflux pump gene expression of Rv1250 in Mycobacterium tuberculosis collected from tuberculosis patients. New Microbes New Infect. 2019, 32, 100609. [Google Scholar] [CrossRef] [PubMed]
- Kanji, A.; Hasan, R.; Ali, A.; Zaver, A.; Zhang, Y.; Imtiaz, K.; Shi, W.; Clark, T.G.; McNerney, R.; Phelan, J.; et al. Single nucleotide polymorphisms in efflux pumps genes in extensively drug resistant Mycobacterium tuberculosis isolates from Pakistan. Tuberculosis 2017, 107, 20–30. [Google Scholar] [CrossRef] [PubMed]
- Popov, A.; Klimovich, A.; Styshova, O.; Moskovkina, T.; Shchekotikhin, A.; Grammatikova, N.; Dezhenkova, L.; Kaluzhny, D.; Deriabin, P.; Gerasimenko, A.; et al. Design, synthesis and biomedical evaluation of mostotrin, a new water soluble tryptanthrin derivative. Int. J. Mol. Med. 2020, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Cooper, C.B. Development of Mycobacterium tuberculosis Whole Cell Screening Hits as Potential Antituberculosis Agents. J. Med. Chem. 2013, 56, 7755–7760. [Google Scholar] [CrossRef] [PubMed]
- Hartkoorn, R.C.; Uplekar, S.; Cole, S.T. Cross-resistance between clofazimine and bedaquiline through upregulation of MmpL5 in Mycobacterium tuberculosis. Antimicrob. Agents Chemother. 2014, 58, 2979–2981. [Google Scholar] [CrossRef] [Green Version]
- Ramos, J.L.; Martínez-Bueno, M.; Molina-Henares, A.J.; Terán, W.; Watanabe, K.; Zhang, X.; Gallegos, M.T.; Brennan, R.; Tobes, R. The TetR family of transcriptional repressors. Microbiol. Mol. Biol. Rev. 2005, 69, 326–356. [Google Scholar] [CrossRef] [Green Version]
- Li, P.; Gu, Y.; Li, J.; Xie, L.; Li, X.; Xie, J. Mycobacterium tuberculosis Major Facilitator Superfamily Transporters. J. Membr. Biol. 2017, 250, 573–585. [Google Scholar] [CrossRef]
- Fluman, N.; Bibi, E. Bacterial multidrug transport through the lens of the major facilitator superfamily. Biochim. Biophys. Acta Proteins Proteom. 2009, 1794, 738–747. [Google Scholar] [CrossRef]
- Kaur, P.; Agarwal, S.; Datta, S. Delineating bacteriostatic and bactericidal targets in mycobacteria using IPTG inducible antisense expression. PLoS ONE 2009, 4, e5923. [Google Scholar] [CrossRef] [Green Version]
- Belisle, J.T.; Mahaffey, S.B.; Hill, P.J. Isolation of Mycobacterium Species Genomic DNA. In Mycobacteria Protocols; Methods in Molecular Biology; Humana Press: Totowa, NJ, USA, 2010; Volume 465, pp. 1–12. [Google Scholar]
- Andrews, S. FastQC: A Quality Control Tool for High Throughput Sequence Data. 2010. Available online: http://www.bioinformatics.babraham.ac.uk/projects/fastqc (accessed on 18 April 2020).
- Li, H.; Durbin, R. Fast and accurate long-read alignment with Burrows-Wheeler transform. Bioinformatics 2010, 26, 589–595. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H. A statistical framework for SNP calling, mutation discovery, association mapping and population genetical parameter estimation from sequencing data. Bioinformatics 2011, 27, 2987–2993. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koboldt, D.C.; Zhang, Q.; Larson, D.E.; Shen, D.; McLellan, M.D.; Lin, L.; Miller, C.A.; Mardis, E.R.; Ding, L.; Wilson, R.K. VarScan 2: Somatic mutation and copy number alteration discovery in cancer by exome sequencing. Genome Res. 2012, 22, 568–576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
| Compound | M. smegmatis Strains MICs | ||||||
|---|---|---|---|---|---|---|---|
| Mw | mc2 1551 | Δmmp5 2 | atr9c3 | ||||
| µg/mL | µM | µg/mL | µM | µg/mL | µM | ||
| 1a | 248.24 | 4 | 16.1 | 4 | 16.1 | 16 | 64.4 |
| 1b | 266.23 | 4 | 15 | 1 | 3.8 | 4 | 15 |
| Strain | Gene | MICs, µg/mL | ||||||
|---|---|---|---|---|---|---|---|---|
| MSMEG_1963 | MSMEG_5597 | MSMEG_4427 | 1a | 1b | ||||
| Nucleotide | Amino Acid | Nucleotide | Amino Acid | Nucleotide | Amino Acid | |||
| mc2-8 | w.t. | w.t. | C49>CTG | frameshift | CTG>CGG | L236R | 16 | 16 |
| mc2-10 | CGT>TGT | R893C | w.t. | w.t. | CTG>CGG | L236R | >49 | >53 |
| mc2-16 | w.t. | w.t. | Transposon insertion | CTG>CGG | L236R | 16 | 8 | |
| mc2-18 | w.t. | w.t. | Transposon insertion | CTG>CGG | L236R | 32 | 8 | |
| Δmmp5-1 | CGC>TGC | R523C | w.t. | w.t. | w.t. | w.t. | >49 | >53 |
| Δmmp5-3 | CGC>TGC | R523C | w.t. | w.t. | w.t. | w.t. | >49 | >53 |
| Δmmp5-6 | CGC>CTC | R523L | w.t. | w.t. | w.t. | w.t. | >49 | >53 |
| Δmmp5-10 | w.t. | w.t. | TGA>CGA | Stop207R | w.t. | w.t. | 8 | 4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Frolova, S.G.; Klimina, K.M.; Kumar, R.; Vatlin, A.A.; Salunke, D.B.; Kendrekar, P.; Danilenko, V.N.; Maslov, D.A. Identification of Mutations Conferring Tryptanthrin Resistance to Mycobacterium smegmatis. Antibiotics 2021, 10, 6. https://doi.org/10.3390/antibiotics10010006
Frolova SG, Klimina KM, Kumar R, Vatlin AA, Salunke DB, Kendrekar P, Danilenko VN, Maslov DA. Identification of Mutations Conferring Tryptanthrin Resistance to Mycobacterium smegmatis. Antibiotics. 2021; 10(1):6. https://doi.org/10.3390/antibiotics10010006
Chicago/Turabian StyleFrolova, Svetlana G., Ksenia M. Klimina, Ravinder Kumar, Aleksey A. Vatlin, Deepak B. Salunke, Pravin Kendrekar, Valery N. Danilenko, and Dmitry A. Maslov. 2021. "Identification of Mutations Conferring Tryptanthrin Resistance to Mycobacterium smegmatis" Antibiotics 10, no. 1: 6. https://doi.org/10.3390/antibiotics10010006
APA StyleFrolova, S. G., Klimina, K. M., Kumar, R., Vatlin, A. A., Salunke, D. B., Kendrekar, P., Danilenko, V. N., & Maslov, D. A. (2021). Identification of Mutations Conferring Tryptanthrin Resistance to Mycobacterium smegmatis. Antibiotics, 10(1), 6. https://doi.org/10.3390/antibiotics10010006