Abstract
Salmonella enterica is known as one of the most common foodborne pathogens worldwide. While salmonellosis is usually self-limiting, severe infections may require antimicrobial therapy. However, increasing resistance of Salmonella to antimicrobials, particularly fluoroquinolones and cephalosporins, is of utmost concern. The present study aimed to investigate the antimicrobial susceptibility of S. enterica isolated from pork, the major product in Philippine livestock production. Our results show that both the qnrS and the blaTEM antimicrobial resistance genes were present in 61.2% of the isolates. While qnrA (12.9%) and qnrB (39.3%) were found less frequently, co-carriage of blaTEM and one to three qnr subtypes was observed in 45.5% of the isolates. Co-carriage of blaTEM and blaCTX-M was also observed in 3.9% of the isolates. Antimicrobial susceptibility testing revealed that the majority of isolates were non-susceptible to ampicillin and trimethoprim/sulfamethoxazole, and 13.5% of the isolates were multidrug-resistant (MDR). MDR isolates belonged to either O:3,10, O:4, or an unidentified serogroup. High numbers of S. enterica carrying antimicrobial resistance genes (ARG), specifically the presence of isolates co-carrying resistance to both β-lactams and fluoroquinolones, raise a concern on antimicrobial use in the Philippine hog industry and on possible transmission of ARG to other bacteria.
1. Introduction
Salmonella infections, or salmonellosis, are commonly acquired through consumption of contaminated food of animal origin. In the Philippines, Salmonella enterica was shown to be the leading cause of foodborne disease outbreaks from 1995–2018 [1,2]. While the disease is usually self-limiting, it may require antimicrobial therapy when the infection becomes invasive. Fluoroquinolone ciprofloxacin and extended-spectrum cephalosporin (ESC) ceftriaxone are the current treatments of choice because the emergence of antimicrobial resistance (AMR) has rendered several drugs such as ampicillin, chloramphenicol, and trimethoprim/sulfamethoxazole obsolete in salmonellosis therapy [3,4].
Resistance to β-lactams, such as ESCs, is most commonly attributed to the bla genes of subtypes TEM, SHV, and CTX-M, which encode for β-lactamases that hydrolyze the β-lactam ring, thereby rendering the drug inactive [5,6]. In contrast to β-lactam resistance, fluoroquinolone resistance is typically attributed to chromosomal mutations in the quinolone targets DNA gyrase and topoisomerase IV, and overexpression of efflux pumps that reduce drug accumulation [7]. However, plasmid-mediated quinolone resistance (PMQR), such as qnr genes, may also occur. These genes are broadly distributed worldwide and are commonly found in association with genes encoding for β-lactamases [8,9,10,11]. Consequently, bla and qnr genes have been increasingly found in bacteria isolated from livestock animals [8,9,12,13,14,15,16,17]. If motile, resistance determinants may accelerate the spread of AMR when these are taken up by non-pathogenic or pathogenic bacteria alike.
There is evidence that substantial use of antimicrobials in food-producing animals may drive the emergence of drug-resistant strains [12,18,19]. While the use of certain antimicrobials such as nitrofurans and chloramphenicol has been banned in livestock production in several parts of the world, AMR in agriculture remains a global challenge [12,19]. Monitoring AMR development in livestock and meat allows early detection of AMR emergence and prevalence [20], which can be used to design interventions to improve antimicrobial therapy and reduce resistance selection pressure [21,22]. This is generally accomplished by antimicrobial susceptibility testing (AST) and detection of antimicrobial resistance genes (ARG).
In the Philippines, pork makes up the majority of livestock production and amounts to a 3.8 M USD industry [23]. The country’s rapidly growing population is expected to further increase pork consumption and production. If left unchecked, AMR may lead to challenges in food production, food security, food safety, economic losses to the hog industry, and AMR spillover to the surrounding environment [12,18,22]. Therefore, this study aimed to investigate the antimicrobial susceptibility and frequency of β-lactamase-encoding genes (blaCTX-M, blaSHV, and blaTEM) and plasmid-mediated quinolone resistance (qnrA, qnrB, and qnrS) in S. enterica from slaughtered pigs in Metro Manila, Philippines.
2. Results
In total, 178 isolates were analyzed in this study. Most isolates belonged to groups O:3,10 (38.8%) and O:7 (30.3%). These were followed by groups O:4 (21.3%), and O:8 (1.7%) and O:9 (1.7%). Eleven isolates (6.2%) were not grouped and were therefore not subtyped. All isolates belonging to group O:9 carried the serovar-specific gene encoding Sdf I, indicating presumptive S. enterica Enteritidis (Table 1).

Table 1.
Molecular characterization of S. enterica isolates.
Results from Vitek® 2 AST revealed that isolates were generally resistant to β-lactams but were susceptible to quinolones. A large number were non-susceptible to ampicillin (71.9%) and trimethoprim/sulfamethoxazole (70.8%). Non-susceptibility to key drugs, ceftazidime, ceftriaxone, and ciprofloxacin was observed in 8.4%, 7.9%, 15.7% of the isolates, respectively (Table 2). Multidrug resistance was observed in 24 (13.5%) isolates; most of which were non-susceptible to four classes of antimicrobial agents (Table 3). Although there are 15 and 14 isolates non-susceptible to the ESCs ceftazidime and ceftriaxone, respectively, Vitek® 2 AST reported only one isolate with an extended-spectrum β-lactamase (ESBL)-producing phenotype.
Table 2.
Non-susceptibility levels of 178 S. enterica isolates against different antimicrobial agents.
Table 3.
Multidrug resistance patterns of 24 S. enterica isolates.
Polymerase chain reaction (PCR) assays targeting bla genes revealed that 61.2% and 5.1% of the isolates harbored blaTEM and blaCTX-M genes, respectively. No isolate carried blaSHV. CTX-M variant typing revealed that 6/9 blaCTX-M-carrying isolates carried blaCTX-M-1, and 3/9 carried blaCTX-M-2. Co-carriage of blaCTX-M (four under the CTX-M-1 group, three under the CTX-M-2 group) and blaTEM was observed in seven isolates. For qnr genes, 12.9%, 39.3%, and 61.2% were harboring the qnrA, qnrB, and qnrS genes, respectively. Co-carriage of blaTEM and one to three qnr subtypes were found in 45.5% of the isolates (Figure 1).
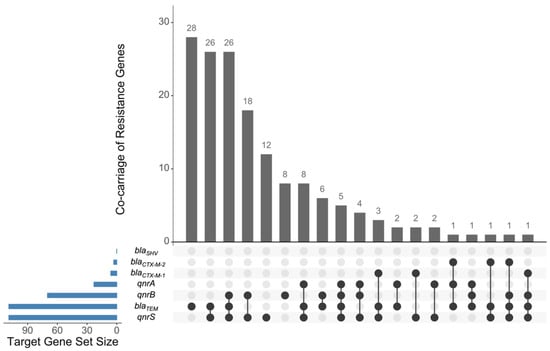
Figure 1.
S. enterica isolates carrying resistance genes. Horizontal bar graphs show the total number of isolates carrying a particular antimicrobial resistance gene (ARG); vertical bar graphs show the number of isolates carrying one or more ARG. Figure was generated using UpSetR [25].
3. Discussion
Since it has been established that ampicillin and trimethoprim/sulfamethoxazole have become obsolete in salmonellosis therapy, high non-susceptibility rates to these antimicrobials were expected. In many countries, aminopenicillins, which include ampicillin as well as trimethoprim, sulfamethoxazole, and trimethoprim/sulfonamide combinations are among the most frequently used antimicrobials in livestock production [12,26]. In the Philippines, amoxicillin, gentamicin, and trimethoprim/sulfamethoxazole are some common antimicrobials used specifically in hog farming. These antimicrobials are generally administered in all phases of pork production [26]. In this study, non-susceptibilities to ampicillin and trimethoprim/sulfamethoxazole were observed in 71.9% and 70.8% of S. enterica, respectively. This was similar to non-susceptibility rates reported in the Philippines in 2017, 70.5% and 80.3% to ampicillin and trimethoprim/sulfamethoxazole, respectively [27]. While non-susceptibility to the latter appeared to be common with all serogroups tested in the present study, non-susceptibility to ampicillin was not observed in the O:9 group. Phongaran et al. [13] reported that 69.0% of Salmonella isolated from hogs in Thailand were resistant to ampicillin. However, in this study, only 35.7% were resistant to trimethoprim/sulfamethoxazole. One study conducted among pork in Vietnam reported that 36.7% of Salmonella isolates were resistant to trimethoprim/sulfamethoxazole and only 41.3% to ampicillin [28]. Conversely, low rates of resistance (<5%) to ESC and ciprofloxacin were reported in both studies [13,28], while the present study reported rates that were almost twice as high (<10%) and that were mainly due to isolates from groups O:3,10 and O:4. However, only one of the isolates in this study, which belonged to the O:3,10 group, produced an ESBL phenotype. Non-susceptibility of other isolates to ESC must be due to broad-spectrum β-lactamases that are not affected by β-lactamase inhibitors. Three isolates of unidentified serogroups were non-susceptible to carbapenems, and one which showed the MDR pattern Pen, Pen/IB, Car, Flu. Carbapenems are not used in agriculture in Southeast Asia [12]; thus, this emerging non-susceptibility to carbapenems may warrant further investigation.
In this study, multidrug resistance was observed in 13.5% of S. enterica isolates from O:3,10 (11), O:4 (9), and unidentified serogroups (3). Reports of multidrug-resistant (MDR) Salmonella isolated from hogs in other Southeast Asian countries are higher (30–40%) [13,28]. In particular, multidrug resistance in the present study was five times lower than that reported in the Philippines in 2017 [27]. In other countries, higher rates (70–80%) of MDR Salmonella isolated from pork and the pork production chain were observed [15,29]. Of the 24 MDR S. enterica isolates in the present study, 15 and 8 were non-susceptible to ESC and fluoroquinolones, respectively, the current drug options in treating salmonellosis. Multidrug resistance is a challenge, as it narrows down the options for antimicrobial therapy.
The majority of studies on bla genes and livestock animals in Southeast Asian countries are focused on E. coli in which blaTEM and blaCTX-M are the most frequently identified bla genes [12]. In Salmonella, blaTEM appears to be the most common. Lalruatdiki et al. [14] observed that in India, 30% of Salmonella isolated from a pig population were carrying blaTEM, and 10% carried blaCTX-M. Similarly, a survey in Italy observed approximately 21% of Salmonella recovered from pork samples had the presence of blaTEM [30]. Co-carriage of blaCTX-M and blaTEM has also been observed in ESBL-producing Salmonella from pigs [14,31]. In the present study, co-carriage of blaTEM and blaCTX-M was found in seven (3.9%) isolates. However, none of these isolates were ESBL-producing strains which could suggest that they carry silent copies of bla genes. The presence of inactive ESBL genes has been reported in Klebsiella pneumoniae [32] and in Escherichia coli [33]. The only ESBL-producing Salmonella in this study was carrying only blaTEM. It is possible that this blaTEM subtype could encode ESBLs, or the isolate could be carrying other ESBL-encoding genes. Among isolates carrying blaTEM, 93.6% (102/109) were non-susceptible to ampicillin, but susceptible to ampicillin/clavulanic acid and piperacillin/tazobactam. Conversely, among isolates non-susceptible to ampicillin, 20.3% (26/128) were not carrying blaTEM. Furthermore, some of these isolates exhibited non-susceptibility to ampicillin/clavulanic acid and ceftriaxone. While most blaTEM in the study possibly confer only broad-spectrum β-lactam resistance considering the high rates of non-susceptibility to ampicillin, its presence in combination with other resistance determinants could render an isolate multidrug-resistant.
We report in this study that 71.3% of S. enterica isolates harbored PMQR. The genes qnrA, qnrB, and qnrS were observed in 12.9%, 39.3%, and 61.2% of the isolates, respectively. While Qnr proteins offer only low resistance against quinolones, these have been shown to broaden the mutant selection window in bacteria [7]. Lin et al. [16] demonstrated that ciprofloxacin resistance conferred by PMQR is even comparable to that of quinolone target mutations. However, Temmerman et al. [34] reported a limited role of PMQR in quinolone resistance. Nevertheless, investigating PMQR is significant because it often carries other ARG [9,10,11]. Prevalence rates of qnr genes appear to vary among samples and geographical locations. Cameron-Veas et al. [15] reported that 15% of S. enterica isolated from a pork production chain in Brazil were carrying qnrB, and none of them carried qnrA and qnrS. A separate study in China reported the prevalence of qnrD (3%), qnrB (16%), and qnrS (66%) [16] in foodborne Salmonella. In Thailand and in Laos, Sinwat et al. [17] found only 1–8% of S. enterica isolated from pork carried the same qnr genes. This highlights the importance of a national surveillance of ARG since it appears that individual countries seem to have different prevalence rates.
Several studies have also reported the association of qnr genes with bla genes. One MDR Salmonella isolated from a piglet in Spain carried both qnrB and blaCTX-M. Moawad et al. [8] found that 33% of Salmonella from poultry and beef in Egypt were carrying qnr genes and either blaCTX-M, blaTEM, or both. Whether qnr and bla genes reside within the same plasmid was not confirmed in either of the studies. However, Penha Filho et al. [9] recently isolated Salmonella from poultry in Brazil which carried both blaCTX-M-2 and qnrB in the same plasmid. In clinical isolates of S. enterica, Escherichia coli, and Klebsiella pneumoniae, qnr genes have also been found within the same plasmid as that of blaTEM or blaCTX-M [10,11]. In the present study, 81 blaTEM-carrying isolates and all 9 blaCTX-M-carrying isolates were harboring one to three qnr subtypes.
The increasing prevalence of MDR Salmonella in livestock animals has been widely reported [13,15,27,29] and is mainly attributed to the inappropriate use of antimicrobial agents in veterinary medicine [18,19]. MDR Salmonella in pork may potentially have an even larger impact on public health due to discharge of livestock waste into water sources. A recent survey in Ho Chi Minh City of surface water and produce indicated 17.5% of vegetable samples were positive for Salmonella, and of all isolates recovered, 26.5% were considered MDR Salmonella. The authors suggested that livestock runoff into surface water used for irrigation and agriculture processing were likely contamination sources [35]. We report that 89.4% of S. enterica isolated from slaughtered pork were non-susceptible to at least one antimicrobial agent and 13.5% were MDR. All MDR isolates belonged to either O:3,10, O:4, or a different serogroup that was not tested. Majority of the isolates were also harboring blaTEM that possibly encode broad-spectrum β-lactamases, and qnrS, which could facilitate emergence of mutations that target quinolone resistance.
While worldwide AMR surveillance has allowed the determination of the evolution of resistance, national surveillance will allow countries to create policies that would fit their needs. Generation of local information on AMR and antimicrobial consumption in the veterinary and agricultural sectors will allow the development of relevant approaches to tackle AMR [21,22]. This is highly important for low- and middle-income countries (LMICs) as strategies proven effective to work in developed countries may not be suitable for LMICs. Attention to AMR in the agricultural sector began in the Philippines only recently, and further surveillance is necessary to identify emerging resistant S. enterica in the pork production chain.
4. Materials and Methods
4.1. Sample Collection
The study population consisted of freshly slaughtered hogs from seven abattoirs across four districts of Metro Manila, Philippines. The abattoirs selected were all registered with the National Meat Inspection Service (NMIS) of the Philippines and were the major slaughtering facilities in each district. Informed consent was obtained from the NMIS; hence, ethics approval was waived for this particular study. Animal slaughter and evisceration were performed according to national regulations. Informed consent was also obtained from veterinarians in charge of the abattoirs, and farm owners for sample collection. Tissue samples from hog tonsils and jejunum were collected post-slaughter and under the supervision of a veterinarian. Sample collection was performed as previously described [27]. Briefly, tissues were collected from each hog upon evisceration using sterile forceps and scissors, and then immediately transferred into sterile bags. All samples were kept chilled upon collection and during transport and were immediately processed in the laboratory.
4.2. Bacterial Isolation and Identification
Bacterial isolates analyzed in this study included isolates from a previous study that were not tested for antimicrobial susceptibility, collected from June to December 2013 (n = 117) [27], with isolates collected from June to December 2014 (n = 61). Bacteria were first enriched prior to isolation as previously described [27]. Briefly, 25 g of each sample was transferred to 225 mL buffered peptone water (BPW) and incubated overnight at 35 °C. Afterward, 100 µL of pre-enriched bacterial culture in BPW was inoculated into 10 mL Rappaport-Vassiliadis broth (RVB), and then incubated overnight at 42 °C for selective enrichment of S. enterica. RVB cultures were inoculated onto brilliant green agar (BGA) and xylose lysine deoxycholate agar (XLD) and then incubated overnight at 35 °C for isolation. Presumptive S. enterica were then inoculated onto nutrient agar (NA) and incubated overnight at 35 °C for subsequent total DNA extraction.
Total DNA was extracted by harvesting colonies using a sterile 1 μL loop and suspending these in 100 μL TE buffer (10 mM Tris, 1 mM EDTA at pH 8.0). The suspension was boiled for 10 min, and pelleted at 6000 rpm for 5 min. The supernatant was collected and then stored at −20 °C until use. These DNA extracts were used in both PCR-based identification of S. enterica and detection of ARG.
Each PCR reaction for S. enterica identification contained 2 μL DNA, 10 pmol each of forward and reverse primers, and HiPi PCR Premix (Elpis Biotech, Daejeon, Korea) in a final volume of 20 μL. Amplification of a 244 bp region in the species-specific invA gene was performed as previously described [36]. PCR products were subsequently analyzed via capillary electrophoresis. S. enterica KCTC 2421 was used as a positive control.
4.3. Molecular Characterization of S. enterica
All isolates were subjected to a two-step PCR assay to characterize the somatic and flagellar antigens. The first step included 12 primers to identify the most common Salmonella serogroups: O:2, O:4, O:6,7, O:8, O:9, and O:3,10 and was based on a previously described protocol [37]. The second step included nine primers to identify the first-phase flagellar antigens (H1) and two to identify a fragment of the gene encoding the Salmonella difference fragment I (Sdf I) unique to S. enterica Enteritidis. The primers used are listed in Table 4.
Table 4.
Targets and primers used in molecular characterization of S. enterica.
Each reaction in the second step contained 2 μL DNA, 10 pmol each of forward and reverse primers, and 6.25 μL GoTaq® Green Master Mix (Promega, Madison, USA) in a final volume of 12.5 μL. For Sdf I, PCR was carried out under the following conditions: initial denaturation step at 95 °C for 3 min; 30 cycles of denaturation at 95 °C for 30 s, annealing at 58 °C for 30 s, extension at 72 °C for 1 min; and a final extension step at 72 °C for 5 min. For the H1 antigens, including G complex alleles, monoplex PCR for each target gene was carried out under the following conditions: initial denaturation step at 95 °C for 2 min; 35 cycles of denaturation at 95 °C for 30 s, annealing at 55 °C for 30 s, extension at 72 °C for 30 s; and a final extension step at 72 °C for 5 min.
Amplicons were analyzed in 1.5% (monoplex PCR products) or 2% (multiplex PCR products) agarose gels stained either with GelRed™ Nucleic Acid Gel Stain (Biotium, Fremont, USA) or SYBR® Safe DNA Gel Stain (Invitrogen, Carlsbad, USA) (1:10,000). Amplicons were allowed to separate at 100 V for 20–30 min and then viewed in a gel documentation system. KAPA™ Universal Ladder (KAPA Biosystems, Boston, USA) was used to estimate the molecular weights of the products.
4.4. Bacterial Storage and Recovery
Salmonella enterica isolates were maintained as glycerol stocks until further analyses. Glycerol stocks were prepared by gently mixing 300 μL of sterile 80% glycerol solution to 700 μL of overnight culture of S. enterica in tryptic soy broth (TSB) and then stored at −20 °C.
To recover glycerol stocks, 200 μL of the culture was inoculated into 800 μL TSB and then incubated overnight at 35 °C. TSB cultures were then inoculated onto XLD and incubated overnight at 35 °C to ensure purity of the culture. Typical Salmonella colonies were maintained in NA until subsequent antimicrobial susceptibility testing.
4.5. Antimicrobial Susceptibility Testing
The Vitek® 2 AST system was used to generate antimicrobial susceptibility profiles of the isolates. It automatically classifies isolates into susceptible, intermediate, or resistant to a particular antimicrobial agent based on the latest breakpoints provided by the Clinical and Laboratory Standards Institute (CLSI). Multidrug resistance was defined as non-susceptibility to at least one antimicrobial agent in three or more antimicrobial categories as recommended by Magiorakos et al. [24].
Inoculum preparation for the automated AST was followed as previously described [27]. Vitek® 2 AST-N261 cards were used which contain 15 antimicrobials including amikacin, amoxicillin/clavulanate, ampicillin, cefepime, cefoxitin, ceftazidime, ceftriaxone, ciprofloxacin, colistin, ertapenem, gentamicin, imipenem, meropenem, piperacillin/tazobactam, and trimethoprim/sulfamethoxazole and an ESBL test. Colistin was not tested because there are currently no CLSI breakpoints available for Salmonella spp. The ESBL test included cefepime, ceftriaxone, and ceftazidime alone and in combination with clavulanic acid. Vitek® 2 AST reports either a positive or negative ESBL test. Each test run was accompanied with AST for E. coli ATCC 25922 (negative control for ESBL test) and K. pneumoniae ATCC 600703 (positive control for ESBL test) (Supplementary Table S1).
4.6. Detection of bla and qnr Genes
S. enterica isolates were screened for β-lactamase-encoding genes (blaCTX-M, blaSHV, and blaTEM) and quinolone resistance genes (qnrA, qnrB, and qnrS) using monoplex PCR assays. The primers used are listed in Table 5. For bla genes, each reaction contained 2 μL DNA, 10 pmol each of forward and reverse primers, and AccuPower® PCR Premix (Bioneer, Daejeon, Korea) or Maxime PCR Premix (i-StarTaq™ GH) (iNtRON Biotechnology, Seongnam, Korea) in a final volume of 20 μL. PCR was carried out under the following conditions: initial denaturation step at 95 °C for 3 min; 30 cycles of denaturation at 95 °C for 30 s, annealing at 58 °C for blaCTX-M, 56 °C for blaSHV, and 50 °C for blaTEM for 30 s, extension at 72 °C for 1 min; and a final extension step at 72 °C for 10 min. For qnr genes, each reaction contained 2 μL DNA, 10 pmol each of forward and reverse primers, and 6.25 μL GoTaq® Green Master Mix in a final volume of 12.5 μL. PCR was carried out under the following conditions: initial denaturation step at 95 °C for 5 min; 33 cycles of denaturation at 95 °C for 1 min, annealing at 60 °C for 1 min, extension at 72 °C for 1 min; and a final extension step at 72 °C for 10 min.
Table 5.
Resistance gene targets and primers used in the present study.
S. enterica isolates carrying blaCTX-M were subjected to further PCR assays to identify CTX-M variants. The primers used in CTX-M variant typing are listed in Table 3. Each reaction contained 2 μL DNA, 10 pmol each of forward and reverse primers, and 6.25 μL GoTaq® Green Master Mix in a final volume of 12.5 μL. Amplification was performed as previously described [6]. Amplicons were analyzed as described above.
Supplementary Materials
The following are available online at https://www.mdpi.com/article/10.3390/antibiotics10121442/s1. Table S1: Minimum inhibitory concentrations of antimicrobials as determined by Vitek® 2 antimicrobial susceptibility testing. Table S2: Distribution of S. enterica into serogroups and H1/Sdf I typing.
Author Contributions
Conceptualization, A.M.B.C. and W.L.R.; methodology, A.M.B.C.; validation, K.W.W. and W.L.R.; formal analysis, A.M.B.C.; investigation, resources, K.W.W. and W.L.R.; data curation, A.M.B.C. and K.W.W.; writing—original draft preparation, A.M.B.C.; writing—review and editing, A.M.B.C., K.W.W. and W.L.R.; visualization, A.M.B.C.; supervision, K.W.W. and W.L.R.; project administration, W.L.R.; funding acquisition, A.M.B.C., K.W.W. and W.L.R. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Department of Agriculture—Biotechnology Program Office, Philippines (DABIOTECH-R1808); the UNU and GIST Joint Programme on Science and Technology for Sustainability, Gwangju Institute of Science and Technology, Korea; and the Office of the Vice Chancellor for Research and Development of the University of the Philippines Diliman through its Thesis and Dissertation Grant.
Institutional Review Board Statement
Ethical review and approval were waived for this study due to informed consent obtained from the Philippine National Meat Inspection Service. Animal slaughter and evisceration were performed according to national regulations. Informed consent was also obtained from veterinarians in charge of the abattoirs and farm owners for sample collection.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available in Supplementary Material.
Acknowledgments
We thank Phyllis Anne P. Paclibare for her invaluable assistance in sample collection and processing.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Azanza, M.P.V.; Membrebe, B.N.Q.; Sanchez, R.G.R.; Estilo, E.E.C.; Dollete, U.G.M.; Feliciano, R.J.; Garcia, N.K.A. Foodborne disease outbreaks in the Philippines (2005–2018). Philipp. J. Sci. 2019, 148, 317–336. [Google Scholar]
- Azanza, M.P.V. Philippine foodborne-disease outbreaks (1995–2004). J. Food Saf. 2006, 26, 92–102. [Google Scholar] [CrossRef]
- Britto, C.D.; Wong, V.K.; Dougan, G.; Pollard, A.J. A systematic review of antimicrobial resistance in Salmonella enterica serovar Typhi, the etiological agent of typhoid. PLoS Negl. Trop. Dis. 2018, 12, e0006779. [Google Scholar] [CrossRef] [PubMed]
- Gut, A.M.; Vasiljevic, T.; Yeager, T.; Donkor, O.N. Salmonella infection–Prevention and treatment by antibiotics and probiotic yeasts: A review. Microbiology 2018, 164, 1327–1344. [Google Scholar] [CrossRef]
- Monstein, H.-J.; Östholm-Balkhed, Å.; Nilsson, M.V.; Nilsson, M.; Dornbusch, K.; Nilsson, L.E. Multiplex PCR amplification assay for the detection of blaSHV, blaTEM and blaCTX-M genes in Enterobacteriaceae. APMIS 2007, 115, 1400–1408. [Google Scholar] [CrossRef]
- Dallenne, C.; Da Costa, A.; Decré, D.; Favier, C.; Arlet, G. Development of a set of multiplex PCR assays for the detection of genes encoding important β-lactamases in Enterobacteriaceae. J. Antimicrob. Chemother. 2010, 65, 490–495. [Google Scholar] [CrossRef] [PubMed]
- Jacoby, G.A. Mechanisms of resistance to quinolones. Clin. Infect. Dis. 2005, 41, S120–S126. [Google Scholar] [CrossRef] [PubMed]
- Moawad, A.A.; Hotzel, H.; Awad, O.; Tomaso, H.; Neubauer, H.; Hafez, H.M.; El-Adawy, H. Occurrence of Salmonella enterica and Escherichia coli in raw chicken and beef meat in northern Egypt and dissemination of their antibiotic resistance markers. Gut Pathog. 2017, 9, 57. [Google Scholar] [CrossRef]
- Penha Filho, R.A.C.; Ferreira, J.C.; Kanashiro, A.M.I.; Berchieri Junior, A.; da Costa Darini, A.L. Emergent multidrug-resistant nontyphoidal Salmonella serovars isolated from poultry in Brazil coharboring blaCTX-M-2 and qnrB or blaCMY-2 in large plasmids. Diagn. Microbiol. Infect. Dis. 2019, 95, 93–98. [Google Scholar] [CrossRef]
- Cheung, T.K.M.; Chu, Y.W.; Chu, M.Y.; Ma, C.H.; Yung, R.W.H.; Kam, K.M. Plasmid-mediated resistance to ciprofloxacin and cefotaxime in clinical isolates of Salmonella enterica serotype Enteritidis in Hong Kong. J. Antimicrob. Chemother. 2005, 56, 586–589. [Google Scholar] [CrossRef]
- Gutierrez, B.; Herrera-Leon, S.; Escudero, J.A.; Hidalgo, L.; Gonzalez-Sanz, R.; Arroyo, M.; San Millan, Á.; Echeita, M.A.; Gonzalez-Zorn, B. Novel genetic environment of qnrB2 associated with TEM-1 and SHV-12 on pB1004, an IncHI2 plasmid, in Salmonella Bredeney BB1047 from Spain. J. Antimicrob. Chemother. 2009, 64, 1334–1336. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Nhung, N.T.; Cuong, N.V.; Thwaites, G.; Carrique-Mas, J. Antimicrobial usage and antimicrobial resistance in animal production in Southeast Asia: A review. Antibiotics 2016, 5, 37. [Google Scholar] [CrossRef]
- Phongaran, D.; Khang-Air, S.; Angkititrakul, S. Molecular epidemiology and antimicrobial resistance of Salmonella isolates from broilers and pigs in Thailand. Vet. World 2019, 12, 1311–1318. [Google Scholar] [CrossRef]
- Lalruatdiki, A.; Dutta, T.K.; Roychoudhury, P.; Subudhi, P.K. Extended-spectrum β-lactamases producing multidrug resistance Escherichia coli, Salmonella and Klebsiella pneumoniae in pig population of Assam and Meghalaya, India. Vet. World 2018, 11, 868–873. [Google Scholar] [CrossRef]
- Cameron-Veas, K.; Fraile, L.; Napp, S.; Garrido, V.; Grilló, M.J.; Migura-Garcia, L. Multidrug resistant Salmonella enterica isolated from conventional pig farms using antimicrobial agents in preventative medicine programmes. Vet. J. 2018, 234, 36–42. [Google Scholar] [CrossRef] [PubMed]
- Lin, D.; Chen, K.; Chan, E.W.-C.; Chen, S. Increasing prevalence of ciprofloxacin-resistant food-borne Salmonella strains harboring multiple PMQR elements but not target gene mutations. Sci. Rep. 2015, 5, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Sinwat, N.; Angkittitrakul, S.; Coulson, K.F.; Pilapil, F.M.I.R.; Meunsene, D.; Chuanchuen, R. High prevalence and molecular characteristics of multidrug-resistant Salmonella in pigs, pork and humans in Thailand and Laos provinces. J. Med. Microbiol. 2016, 65, 1182–1193. [Google Scholar] [CrossRef] [PubMed]
- Khachatourians, G.G. Agricultural use of antibiotics and the evolution and transfer of antibiotic-resistant bacteria. Can. Med. Assoc. J. 1998, 159, 1129–1136. [Google Scholar]
- Barton, M.D. Impact of antibiotic use in the swine industry. Curr. Opin. Microbiol. 2014, 19, 9–15. [Google Scholar] [CrossRef]
- Reller, L.B.; Weinstein, M.; Jorgensen, J.H.; Ferraro, M.J. Antimicrobial susceptibility testing: A review of general principles and contemporary practices. Clin. Infect. Dis. 2009, 49, 1749–1755. [Google Scholar]
- Kerr, J.R. Antibiotic treatment and susceptibility testing. J. Clin. Pathol. 2005, 58, 786–787. [Google Scholar] [CrossRef]
- Kakkar, M.; Chatterjee, P.; Chauhan, A.S.; Grace, D.; Lindahl, J.; Beeche, A.; Jing, F.; Chotinan, S. Antimicrobial resistance in South East Asia: Time to ask the right questions. Glob. Health Action 2018, 11, 1483637. [Google Scholar] [CrossRef] [PubMed]
- Philippine Statistics Authority. Available online: http://openstat.psa.gov.ph/ (accessed on 16 October 2020).
- Magiorakos, A.-P.; Srinivasan, A.; Carey, R.B.; Carmeli, Y.; Falagas, M.E.; Giske, C.G.; Harbarth, S.; Hindler, J.F.; Kahlmeter, G.; Olsson-Liljequist, B.; et al. Multidrug-resistant, extensively drug-resistant and pandrug-resistant bacteria: An international expert proposal for interim standard definitions for acquired resistance. Clin. Microbiol. Infect. 2012, 18, 268–281. [Google Scholar] [CrossRef] [PubMed]
- Lex, A.; Gehlenborg, N.; Strobelt, H.; Vuillemot, R.; Pfister, H. UpSet: Visualization of intersecting sets. IEEE Trans. Vis. Comput. Graph. 2014, 20, 1983–1992. [Google Scholar] [CrossRef]
- Lekagul, A.; Tangcharoensathien, V.; Yeung, S. Patterns of antibiotic use in global pig production: A systematic review. Vet. Anim. Sci. 2019, 7, 100058. [Google Scholar] [CrossRef]
- Calayag, A.M.B.; Paclibare, P.A.P.; Santos, P.D.M.; Bautista, C.A.C.; Rivera, W.L. Molecular characterization and antimicrobial resistance of Salmonella enterica from swine slaughtered in two different types of Philippine abattoir. Food Microbiol. 2017, 65, 16. [Google Scholar] [CrossRef]
- Tu, L.T.P.; Hoang, N.V.M.; Cuong, N.V.; Campbell, J.; Bryant, J.E.; Hoa, N.T.; Kiet, B.T.; Thompson, C.; Duy, D.T.; Phat, V.V.; et al. High levels of contamination and antimicrobial-resistant non-typhoidal Salmonella serovars on pig and poultry farms in the Mekong Delta of Vietnam. Epidemiol. Infect. 2015, 143, 3074–3086. [Google Scholar] [CrossRef] [PubMed]
- Viana, C.; Sereno, M.J.; Pegoraro, K.; Yamatogi, R.S.; Call, D.R.; dos Santos Bersot, L.; Nero, L.A. Distribution, diversity, virulence genotypes and antibiotic resistance for Salmonella isolated from a Brazilian pork production chain. Int. J. Food Microbiol. 2019, 310, 108310. [Google Scholar] [CrossRef]
- Nguyen Thi, H.; Pham, T.-T.-T.; Turchi, B.; Fratini, F.; Ebani, V.V.; Cerri, D.; Bertelloni, F. Characterization of Salmonella spp. isolates from swine: Virulence and antimicrobial resistance. Animals 2020, 10, 2418. [Google Scholar] [CrossRef]
- Trongjit, S.; Angkititrakul, S.; Tuttle, R.E.; Poungseree, J.; Padungtod, P.; Chuanchuen, R. Prevalence and antimicrobial resistance in Salmonella enterica isolated from broiler chickens, pigs and meat products in Thailand–Cambodia border provinces. Microbiol. Immunol. 2017, 61, 23–33. [Google Scholar] [CrossRef]
- Li, X.; Yao, Z.; Yuan, L.; Qi, W.; Shuchang, A.; Jichao, C.; Yusheng, C.; Lin, L.; Jiabin, L.; Zhancheng, G. Identification of Klebsiella pneumoniae strains harboring inactive extended-spectrum beta-lactamase antibiotic-resistance genes. Chin. Med. J. 2014, 127, 3051–3057. [Google Scholar]
- Cruz, M.C.; Hedreyda, C.T. Detection of plasmid-borne β-lactamase genes in extended-spectrum β-lactamase (ESBL) and non-ESBL-producing Escherichia coli clinical isolates. Philipp. J. Sci 2017, 146, 167–175. [Google Scholar]
- Temmerman, R.; Garmyn, A.; Antonissen, G.; Vanantwerpen, G.; Vanrobaeys, M.; Haesebrouck, F.; Devreese, M. Evaluation of fluoroquinolone resistance in clinical avian pathogenic Escherichia coli isolates from Flanders (Belgium). Antibiotics 2020, 9, 800. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, D.T.A.; Awasthi, S.P.; Hoang, P.H.; Nguyen, P.D.; Jayedul, H.; Hatanaka, N.; Hinenoya, A.; Van Dang, C.; Faruque, S.M.; Yamasaki, S. Prevalence, serovar and antimicrobial resistance of nontyphoidal Salmonella in vegetable, fruit and water samples in Ho Chi Minh City, Vietnam. Foodborne Pathog. Dis. 2021, 18, 354–363. [Google Scholar] [CrossRef] [PubMed]
- Ng, K.C.S.; Rivera, W.L. Multiplex PCR-based serogrouping and serotyping of Salmonella enterica from tonsil and jejunum with jejunal lymph nodes of slaughtered swine in Metro Manila, Philippines. J. Food Prot. 2015, 78, 873–880. [Google Scholar] [CrossRef]
- Soguilon-Del Rosario, S.A.; Rivera, W.L. Incidence and molecular detection of Salmonella enterica serogroups and spvC virulence gene in raw and processed meats from selected wet markets in Metro Manila, Philippines. Int. J. Philipp. Sci. Tech. 2015, 8, 52–56. [Google Scholar] [CrossRef]
- Hong, Y.; Liu, T.; Lee, M.D.; Hofacre, C.L.; Maier, M.; White, D.G.; Ayers, S.; Wang, L.; Berghaus, R.; Maurer, J.J. Rapid screening of Salmonella enterica serovars Enteritidis, Hadar, Heidelberg and Typhimurium using a serologically-correlative allelotyping PCR targeting the O and H antigen alleles. BMC Microbiol. 2008, 8, 1–8. [Google Scholar] [CrossRef]
- Hirose, K.; Itoh, K.-I.; Nakajima, H.; Kurazono, T.; Yamaguchi, M.; Moriya, K.; Ezaki, T.; Kawamura, Y.; Tamura, K.; Watanabe, H. Selective amplification of tyv (rfbE), prt (rfbS), viaB, and fliC genes by multiplex PCR for identification of Salmonella enterica serovars Typhi and Paratyphi A. J. Clin. Microbiol. 2002, 40, 633–636. [Google Scholar] [CrossRef]
- Vanegas, R.A.; Joys, T.M. Molecular analyses of the phase-2 antigen complex 1, 2 of Salmonella spp. J. Bacteriol. 1995, 177, 3863–3864. [Google Scholar] [CrossRef] [PubMed]
- Herrera-León, S.; McQuiston, J.R.; Usera, M.A.; Fields, P.I.; Garaizar, J.; Echeita, M.A. Multiplex PCR for distinguishing the most common phase-1 flagellar antigens of Salmonella spp. J. Clin. Microbiol. 2004, 42, 2581–2586. [Google Scholar] [CrossRef]
- Agron, P.G.; Walker, R.L.; Kinde, H.; Sawyer, S.J.; Hayes, D.C.; Wollard, J.; Andersen, G.L. Identification by subtractive hybridization of sequences specific for Salmonella enterica serovar Enteritidis. Appl. Environ. Microbiol. 2001, 67, 4984–4991. [Google Scholar] [CrossRef] [PubMed]
- Paterson, D.L.; Hujer, K.M.; Hujer, A.M.; Yeiser, B.; Bonomo, M.D.; Rice, L.B.; Bonomo, R.A.; the International Klebsiella Study Group. Extended-spectrum β-lactamases in Klebsiella pneumoniae bloodstream isolates from seven countries: Dominance and widespread prevalence of SHV- and CTX-M-type β-lactamases. Antimicrob. Agents Chemother. 2003, 47, 3554–3560. [Google Scholar] [CrossRef]
- Boyd, D.A.; Tyler, S.; Christianson, S.; McGeer, A.; Muller, M.P.; Willey, B.M.; Bryce, E.; Gardam, M.; Nordmann, P.; Mulvey, M.R.; et al. Complete nucleotide sequence of a 92-Kilobase plasmid harboring the CTX-M-15 extended-spectrum beta-lactamase involved in an outbreak in long-term-care facilities in Toronto, Canada. Antimicrob. Agents Chemother. 2004, 48, 3758–3764. [Google Scholar] [CrossRef]
- Woodford, N.; Fagan, E.J.; Ellington, M.J. Multiplex PCR for rapid detection of genes encoding CTX-M extended-spectrum β-lactamases. J. Antimicrob. Chemother. 2006, 57, 154–155. [Google Scholar] [CrossRef] [PubMed]
- Cattoir, V.; Poirel, L.; Rotimi, V.; Soussy, C.-J.; Nordmann, P. Multiplex PCR for detection of plasmid-mediated quinolone resistance qnr genes in ESBL-producing enterobacterial isolates. J. Antimicrob. Chemother. 2007, 60, 394–397. [Google Scholar] [CrossRef] [PubMed]
- Cattoir, V.; Weill, F.-X.; Poirel, L.; Fabre, L.; Soussy, C.-J.; Nordmann, P. Prevalence of qnr genes in Salmonella in France. J. Antimicrob. Chemother. 2007, 59, 751–754. [Google Scholar] [CrossRef]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).