
Pharmacological Properties, Volatile Organic Compounds, and Genome Sequences of Bacterial Endophytes from the Mangrove Plant Rhizophora apiculata Blume

 ,
,
Abstract
:1. Introduction
2. Results
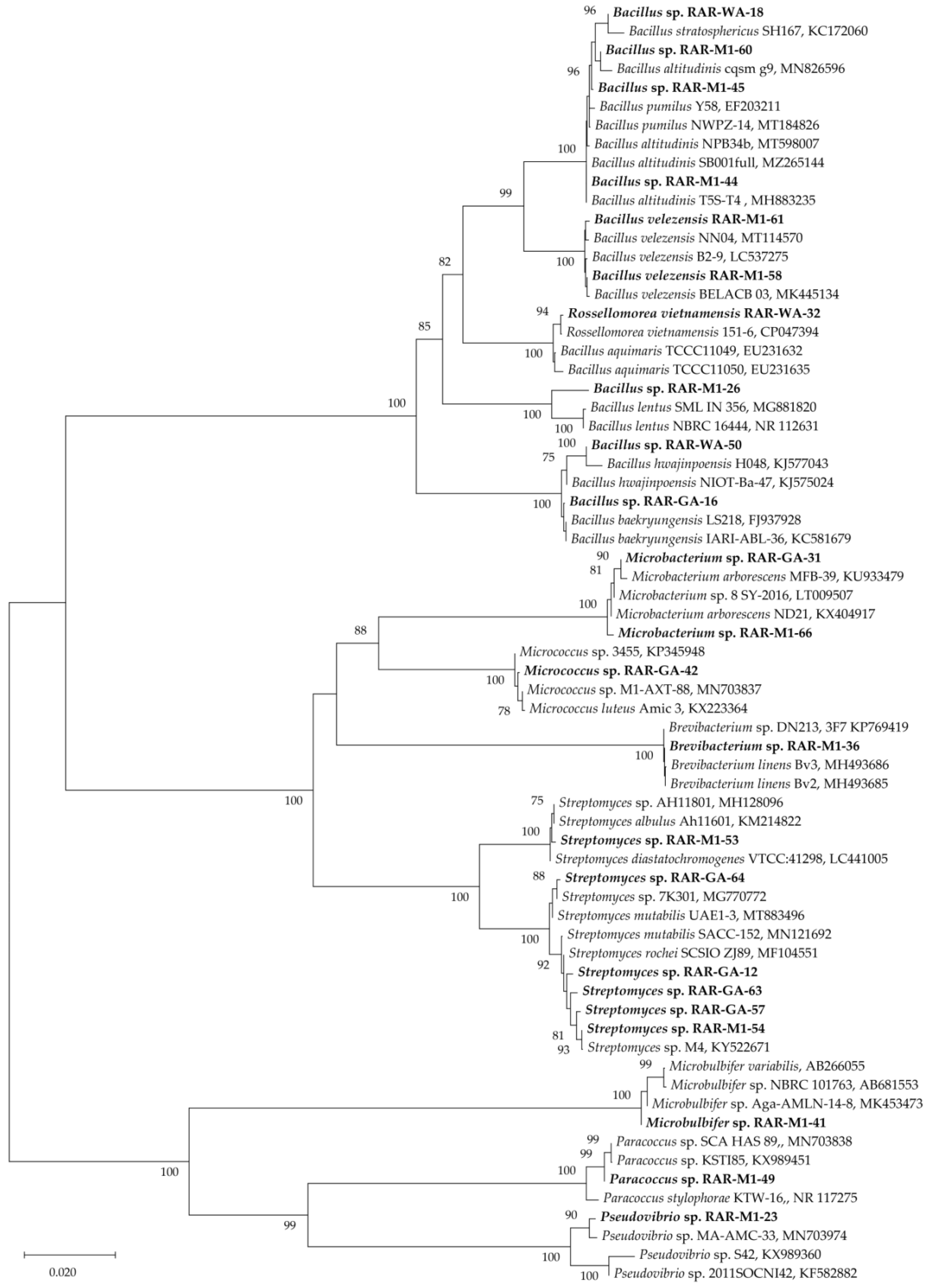
2.1. Isolation and Identification of Endophytic Bacteria
2.2. Pharmacological Evaluation of the Ethyl Acetate Extracts
2.2.1. Antimicrobial Activity
2.2.2. Antioxidant Activity
2.2.3. α-Amylase, α-Glucosidase, and Xanthine Oxidase Inhibitory Activities
2.2.4. Cytotoxic Activity
2.3. Volatile Chemical Composition of the Endophytic Bacterial Extracts by GC-MS
2.4. Genome Sequencing, Assembly, and Annotation of Biosynthesis Gene Clusters of Secondary Metabolites
3. Discussion
4. Materials and Methods
4.1. Plant Collection
4.2. Isolation and Identification of the Endophytic Bacteria
4.3. Preparation of Ethyl Acetate Extracts from the Culture Broths
4.4. Pharmacological Properties of Bacterial Crude Extracts
4.4.1. Antimicrobial Activity
4.4.2. Antioxidant Activity
4.4.3. α-Amylase and α-Glucosidase Activities
4.4.4. Xanthine Oxidase Inhibitory Activity
4.4.5. Cytotoxic Activity
4.5. Gas Chromatography-Mass Spectrometry (GC-MS) Analysis
4.6. Genome Sequencing, Assembly and Annotation of Biosynthesis Gene Clusters of Secondary Metabolites
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Afzal, I.; Shinwari, Z.K.; Sikandar, S.; Shahzad, S. Plant beneficial endophytic bacteria: Mechanisms, diversity, host range and genetic determinants. Microbiol. Res. 2019, 221, 36–49. [Google Scholar] [CrossRef] [PubMed]
- Cipollini, D.; Rigsby, C.M.; Barto, E.K. Microbes as Targets and Mediators of Allelopathy in Plants. J. Chem. Ecol. 2012, 38, 714–727. [Google Scholar] [CrossRef] [PubMed]
- Chuansheng, M.; Barry, S.F. The Use of Beneficial Microbial Endophytes for Plant Biomass and Stress Tolerance Improvement. Recent Pat. Biotechnol. 2010, 4, 81–95. [Google Scholar]
- Rosenblueth, M.; Martínez-Romero, E. Bacterial Endophytes and Their Interactions with Hosts. Mol. Plant-Microbe Interact. 2006, 19, 827–837. [Google Scholar] [CrossRef] [Green Version]
- Senthilkumar, M.; Anandham, R.; Madhaiyan, M.; Venkateswaran, V.; Sa, T. Endophytic Bacteria: Perspectives and Applications in Agricultural Crop Production. In Bacteria in Agrobiology: Crop Ecosystems; Maheshwari, D.K., Ed.; Springer: Berlin/Heidelberg, Germany, 2011; pp. 61–96. [Google Scholar]
- Kathiresan, K.; Bingham, B.L. Biology of mangroves and mangrove Ecosystems. In Advances in Marine Biology; Academic Press: Cambridge, MA, USA, 2001; Volume 40, pp. 81–251. [Google Scholar]
- Thatoi, H.; Behera, B.C.; Mishra, R.R.; Dutta, S.K. Biodiversity and biotechnological potential of microorganisms from mangrove ecosystems: A review. Ann. Microbiol. 2013, 63, 1–19. [Google Scholar] [CrossRef]
- Cadamuro, R.D.; da Silveira Bastos, I.M.A.; Silva, I.T.; da Cruz, A.C.C.; Robl, D.; Sandjo, L.P.; Alves, S., Jr.; Lorenzo, J.M.; Rodríguez-Lázaro, D.; Treichel, H.; et al. Bioactive Compounds from Mangrove Endophytic Fungus and Their Uses for Microorganism Control. J. Fungi 2021, 7, 455. [Google Scholar] [CrossRef]
- Kui-Wu, W.; Shi-Wei, W.; Bin, W.; Ji-Guang, W. Bioactive Natural Compounds from the Mangrove Endophytic Fungi. Mini-Rev. Med. Chem. 2014, 14, 370–391. [Google Scholar]
- Ancheeva, E.; Daletos, G.; Proksch, P. Lead Compounds from Mangrove-Associated Microorganisms. Mar. Drugs 2018, 16, 370–391. [Google Scholar] [CrossRef] [Green Version]
- Xu, D.-B.; Ye, W.-W.; Han, Y.; Deng, Z.-X.; Hong, K. Natural products from mangrove actinomycetes. Mar. Drugs 2014, 12, 2590–2613. [Google Scholar] [CrossRef] [Green Version]
- Takayama, K.; Tateishi, Y.; Kajita, T. Global phylogeography of a pantropical mangrove genus Rhizophora. Sci. Rep. 2021, 11, 7228. [Google Scholar] [CrossRef]
- Gao, M.-Z.; Yuan, X.-Y.; Cheng, M.-C.; Xiao, H.-B.; Bao, S.-X. A new diterpenoid from Rhizophora apiculata. J. Asian Nat. Prod. Res. 2011, 13, 776–779. [Google Scholar] [CrossRef]
- Selvaraj, G.; Kaliamurthi, S.; Thirugnasambandan, R. Effect of Glycosin alkaloid from Rhizophora apiculata in non-insulin dependent diabetic rats and its mechanism of action: In vivo and in silico studies. Phytomedicine 2016, 23, 632–640. [Google Scholar] [CrossRef]
- Moovendhan, M.; Ramasubburayan, R.; Vairamani, S.; Shanmugam, A.; Palavesam, A.; Immanuel, G. Antibiotic susceptibility of Genistein and Alkaloids from Rhizophora apiculata. Biocatal. Agric. Biotechnol. 2014, 3, 323–327. [Google Scholar] [CrossRef]
- Saxena, E.; Garg, H.S. Apiculol, 1β–hydroxy–13–epimanoyl Oxide from Roots of Rhizophora apiculata Mangrove. Nat. Prod. Lett. 1994, 4, 149–154. [Google Scholar] [CrossRef]
- Kokpol, U.; Chavasiri, W.; Chittawong, V.; Bruce, M.; Cunningham, G.N.; Miles, D.H. Long chain aliphatic alcohols and saturated carboxylic acids from heartwood of Rhizophora apiculata. Phytochemistry 1993, 33, 1129–1131. [Google Scholar] [CrossRef]
- Ramalingam, V.; Rajaram, R. Enhanced antimicrobial, antioxidant and anticancer activity of Rhizophora apiculata: An experimental report. 3 Biotech 2018, 8, 200. [Google Scholar] [CrossRef] [PubMed]
- Loo, A.Y.; Jain, K.; Darah, I. Antioxidant activity of compounds isolated from the pyroligneous acid, Rhizophora apiculata. Food Chem. 2008, 107, 1151–1160. [Google Scholar] [CrossRef]
- Gao, M.; Xiao, H. Activity-guided isolation of antioxidant compounds from Rhizophora apiculata. Molecules 2012, 17, 10675–10682. [Google Scholar] [CrossRef] [Green Version]
- Vijayavel, K.; Anbuselvam, C.; Balasubramanian, M.P. Free radical scavenging activity of the marine mangrove Rhizophora apiculata bark extract with reference to naphthalene induced mitochondrial dysfunction. Chem. Biol. Interact. 2006, 163, 170–175. [Google Scholar] [CrossRef]
- Premanathan, M.; Arakaki, R.; Izumi, H.; Kathiresan, K.; Nakano, M.; Yamamoto, N.; Nakashima, H. Antiviral properties of a mangrove plant, Rhizophora apiculata Blume, against human immunodeficiency virus. Antivir. Res. 1999, 44, 113–122. [Google Scholar] [CrossRef]
- Zhang, H.; Jacob, J.A.; Jiang, Z.; Xu, S.; Sun, K.; Zhong, Z.; Varadharaju, N.; Shanmugam, A. Hepatoprotective effect of silver nanoparticles synthesized using aqueous leaf extract of Rhizophora apiculata. Int. J. Nanomed. 2019, 14, 3517–3524. [Google Scholar] [CrossRef] [Green Version]
- Fan, Y.; Wang, Y.; Liu, P.; Fu, P.; Zhu, T.; Wang, W.; Zhu, W. Indole-Diterpenoids with Anti-H1N1 Activity from the Aciduric Fungus Penicillium camemberti OUCMDZ-1492. J. Nat. Prod. 2013, 76, 1328–1336. [Google Scholar] [CrossRef] [PubMed]
- Chaeprasert, S.; Piapukiew, J.; Whalley, A.J.S.; Sihanonth, P. Endophytic fungi from mangrove plant species of Thailand: Their antimicrobial and anticancer potentials. Bot. Mar. 2010, 53, 555–564. [Google Scholar] [CrossRef]
- Zhou, G.; Qiao, L.; Zhang, X.; Sun, C.; Che, Q.; Zhang, G.; Zhu, T.; Gu, Q.; Li, D. Fusaricates H-K and fusolanones A-B from a mangrove endophytic fungus Fusarium solani HDN15-410. Phytochemistry 2019, 158, 13–19. [Google Scholar] [CrossRef]
- UNEP. National Reports on Mangroves in the South China Sea; UNEP/GEF/SCS Technical Publication No. 14; UNEP: Nairobi, Kenya, 2008. [Google Scholar]
- Putra, M.Y.; Karim, F. Antibacterial and antioxidant activity-guided isolation studies on Fusarium sp. associated with the ascidian Botryllus schlosseri. AIP Conf. Proc. 2020, 2243, 020019. [Google Scholar]
- Ser, H.-L.; Palanisamy, U.D.; Yin, W.-F.; Abd Malek, S.N.; Chan, K.-G.; Goh, B.-H.; Lee, L.-H. Presence of antioxidative agent, Pyrrolo[1,2-a]pyrazine-1,4-dione, hexahydro- in newly isolated Streptomyces mangrovisoli sp. nov. Front. Microbiol. 2015, 6, 854. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhai, Y.; Shao, Z.; Cai, M.; Zheng, L.; Li, G.; Yu, Z.; Zhang, J. Cyclo(l-Pro–l-Leu) of Pseudomonas putida MCCC 1A00316 Isolated from Antarctic Soil: Identification and Characterization of Activity against Meloidogyne incognita. Molecules 2019, 24, 768. [Google Scholar] [CrossRef] [Green Version]
- Rhee, K.-H. Cyclic dipeptides exhibit synergistic, broad spectrum antimicrobial effects and have anti-mutagenic properties. Int. J. Antimicrob. Agents 2004, 24, 423–427. [Google Scholar] [CrossRef] [PubMed]
- Nishanth Kumar, S.; Mohandas, C.; Siji, J.V.; Rajasekharan, K.N.; Nambisan, B. Identification of antimicrobial compound, diketopiperazines, from a Bacillus sp. N strain associated with a rhabditid entomopathogenic nematode against major plant pathogenic fungi. J. Appl. Microbiol. 2012, 113, 914–924. [Google Scholar] [CrossRef]
- Casillas-Vargas, G.; Ocasio-Malavé, C.; Medina, S.; Morales-Guzmán, C.; Del Valle, R.G.; Carballeira, N.M.; Sanabria-Ríos, D.J. Antibacterial fatty acids: An update of possible mechanisms of action and implications in the development of the next-generation of antibacterial agents. Prog. Lipid Res. 2021, 82, 101093. [Google Scholar] [CrossRef]
- Chen, S.; Liu, J.; Gong, H.; Yang, D. Identification and antibacterial activity of secondary metabolites from Taxus endophytic fungus. Sheng Wu Gong Cheng Xue Bao 2009, 25, 368–374. [Google Scholar]
- Rahman, S.; Anwar, M.N. Fungitoxic and cytotoxic activity of a novel compound 1,2-benzenedicarboxylic acid, diiooctyl ester of Plumbago zeylanica Linn. Asian J. Microbiol. Biotechnol. Environ. Sci. 2006, 8, 461–464. [Google Scholar]
- Zheng, C.J.; Yoo, J.-S.; Lee, T.-G.; Cho, H.-Y.; Kim, Y.-H.; Kim, W.-G. Fatty acid synthesis is a target for antibacterial activity of unsaturated fatty acids. FEBS Lett. 2005, 579, 5157–5162. [Google Scholar] [CrossRef] [Green Version]
- Galbraith, H.; Miller, T.B.; Paton, A.M.; Thompson, J.K. Antibacterial Activity of Long Chain Fatty Acids and the Reversal with Calcium, Magnesium, Ergocalciferol and Cholesterol. J. Appl. Bacteriol. 1971, 34, 803–813. [Google Scholar] [CrossRef] [PubMed]
- Almoughrabie, S.; Ngari, C.; Guillier, L.; Briandet, R.; Poulet, V.; Dubois-Brissonnet, F. Rapid assessment and prediction of the efficiency of two preservatives against S. aureus in cosmetic products using High Content Screening—Confocal Laser Scanning Microscopy. PLoS ONE 2020, 15, e0236059. [Google Scholar] [CrossRef]
- Lucchini, J.J.; Corre, J.; Cremieux, A. Antibacterial activity of phenolic compounds and aromatic alcohols. Res. Microbiol. 1990, 141, 499–510. [Google Scholar]
- Zou, J.; Gao, P.; Hao, X.; Xu, H.; Zhan, P.; Liu, X. Recent progress in the structural modification and pharmacological activities of ligustrazine derivatives. Eur. J. Med. Chem. 2018, 147, 150–162. [Google Scholar] [CrossRef]
- Yan, S.; Yue, Y.-z.; Zong, Y.; Zeng, L. Tetramethylpyrazine Improves Postoperative Tissue Adhesion: A Drug Repurposing. Chin. J. Integr. Med. 2019, 25, 554–560. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.-t.; Zheng, X.-w.; Chen, S.; Shan, C.-S.; Xu, Q.-Q.; Zhu, J.-Z.; Bao, X.-Y.; Lin, Y.; Zheng, G.-q.; Wang, Y. Chinese herbal medicine for Alzheimer’s disease: Clinical evidence and possible mechanism of neurogenesis. Biochem. Pharmacol. 2017, 141, 143–155. [Google Scholar] [CrossRef] [PubMed]
- Corre, J.; Lucchini, J.J.; Mercier, G.M.; Cremieux, A. Antibacterial activity of phenethyl alcohol and resulting membrane alterations. Res. Microbiol. 1990, 141, 483–497. [Google Scholar] [CrossRef]
- Fraud, S.; Rees, E.L.; Mahenthiralingam, E.; Russell, A.D.; Maillard, J.Y. Aromatic alcohols and their effect on Gram-negative bacteria, cocci and mycobacteria. J. Antimicrob. Chemother. 2003, 51, 1435–1436. [Google Scholar] [CrossRef] [PubMed]
- Davidson, P.M.; Sofos, J.N.; Branen, A.L. Antimicrobials in Food, 3rd ed.; CRC Press: Boca Raton, FL, USA, 2005. [Google Scholar]
- Wu, J.-J.; Huang, J.-W.; Deng, W.-L. Phenylacetic Acid and Methylphenyl Acetate From the Biocontrol Bacterium Bacillus mycoides BM02 Suppress Spore Germination in Fusarium oxysporum f. sp. lycopersici. Front. Microbiol. 2020, 11, 3022. [Google Scholar] [CrossRef]
- Kleinwächter, I.S.; Pannwitt, S.; Centi, A.; Hellmann, N.; Thines, E.; Bereau, T.; Schneider, D. The Bacteriostatic Activity of 2-Phenylethanol Derivatives Correlates with Membrane Binding Affinity. Membranes 2021, 11, 254. [Google Scholar] [CrossRef] [PubMed]
- Hwang Byung, K.; Lim Song, W.; Kim Beom, S.; Lee Jung, Y.; Moon Surk, S. Isolation and In Vivo and In Vitro Antifungal Activity of Phenylacetic Acid and Sodium Phenylacetate from Streptomyces humidus. Appl. Environ. Microbiol. 2001, 67, 3739–3745. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.; Cho, J.-Y.; Kuk, J.-H.; Moon, J.-H.; Cho, J.-I.; Kim, Y.-C.; Park, K.-H. Identification and Antimicrobial Activity of Phenylacetic Acid Produced by Bacillus licheniformis Isolated from Fermented Soybean, Chungkook-Jang. Curr. Microbiol. 2004, 48, 312–317. [Google Scholar] [CrossRef]
- Narayana, K.J.P.; Prabhakar, P.; Vijayalakshmi, M.; Venkateswarlu, Y.; Krishna, P.S.J. Biological activity of phenylpropionic acid isolated from a terrestrial Streptomycetes. Pol. J. Microbiol. 2007, 56, 191–197. [Google Scholar]
- Varsha, K.K.; Devendra, L.; Shilpa, G.; Priya, S.; Pandey, A.; Nampoothiri, K.M. 2,4-Di-tert-butyl phenol as the antifungal, antioxidant bioactive purified from a newly isolated Lactococcus sp. Int. J. Food Microbiol. 2015, 211, 44–50. [Google Scholar] [CrossRef] [PubMed]
- Dharni, S.; Maurya, A.; Samad, A.; Srivastava, S.K.; Sharma, A.; Patra, D.D. Purification, Characterization, and in Vitro Activity of 2,4-Di-tert-butylphenol from Pseudomonas monteilii PsF84: Conformational and Molecular Docking Studies. J. Agric. Food Chem. 2014, 62, 6138–6146. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Zhao, X.; Deng, Y.; Bu, X.; Ye, H.; Guo, N. Antimicrobial potential of myristic acid against Listeria monocytogenes in milk. J. Antibiot. 2019, 72, 298–305. [Google Scholar] [CrossRef]
- Huang, C.B.; Alimova, Y.; Myers, T.M.; Ebersole, J.L. Short- and medium-chain fatty acids exhibit antimicrobial activity for oral microorganisms. Arch. Oral Biol. 2011, 56, 650–654. [Google Scholar] [CrossRef] [Green Version]
- Jeon, Y.-T.; Jun, E.-M.; Oh, K.-B.; Thu, P.Q.; Kim, S.-U. Identification of 12-methyltetradecanoic acid from endophytic Senotrophomonas maltophilia as inhibitor of appressorium formation of Magnaporthe oryzae. J. Korean Soc. Appl. Biol. Chem. 2010, 53, 578–583. [Google Scholar] [CrossRef]
- Yang, P.; Collin, P.; Madden, T.; Chan, D.; Sweeney-Gotsch, B.; McConkey, D.; Newman, R.A. Inhibition of proliferation of PC3 cells by the branched-chain fatty acid, 12-methyltetradecanoic acid, is associated with inhibition of 5-lipoxygenase. Prostate 2003, 55, 281–291. [Google Scholar] [CrossRef]
- Saeed, N.M.; El-Demerdash, E.; Abdel-Rahman, H.M.; Algandaby, M.M.; Al-Abbasi, F.A.; Abdel-Naim, A.B. Anti-inflammatory activity of methyl palmitate and ethyl palmitate in different experimental rat models. Toxicol. Appl. Pharmacol. 2012, 264, 84–93. [Google Scholar] [CrossRef]
- Shaaban, M.T.; Ghaly, M.F.; Fahmi, S.M. Antibacterial activities of hexadecanoic acid methyl ester and green-synthesized silver nanoparticles against multidrug-resistant bacteria. J. Basic Microbiol. 2021, 61, 557–568. [Google Scholar] [CrossRef] [PubMed]
- Dilika, F.; Bremner, P.D.; Meyer, J.J.M. Antibacterial activity of linoleic and oleic acids isolated from Helichrysum pedunculatum: A plant used during circumcision rites. Fitoterapia 2000, 71, 450–452. [Google Scholar] [CrossRef]
- Ström, K.; Sjögren, J.; Broberg, A.; Schnürer, J. Lactobacillus plantarum MiLAB 393 Produces the Antifungal Cyclic Dipeptides Cyclo(l-Phe-l-Pro) and Cyclo(l-Phe-trans-4-OH-l-Pro) and 3-Phenyllactic Acid. Appl. Environ. Microbiol. 2002, 68, 4322–4327. [Google Scholar] [CrossRef] [Green Version]
- Qi, S.-H.; Xu, Y.; Gao, J.; Qian, P.-Y.; Zhang, S. Antibacterial and antilarval compounds from marine bacterium Pseudomonas rhizosphaerae. Ann. Microbiol. 2009, 59, 229. [Google Scholar] [CrossRef]
- Brauns, S.C.; Milne, P.; Naudé, R.; Van de Venter, M. Selected cyclic dipeptides inhibit cancer cell growth and induce apoptosis in HT-29 colon cancer cells. Anticancer Res. 2004, 24, 1713–1719. [Google Scholar]
- Singh, V.K.; Mishra, A.; Jha, B. 3-Benzyl-Hexahydro-Pyrrolo[1,2-a]Pyrazine-1,4-Dione Extracted From Exiguobacterium indicum Showed Anti-biofilm Activity Against Pseudomonas aeruginosa by Attenuating Quorum Sensing. Front. Microbiol. 2019, 10, 1269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sastry, V.M.V.S.; Rao, G.R.K. Dioctyl phthalate, and antibacterial compound from the marine brown alga —Sargassum wightii. J. Appl. Phycol. 1995, 7, 185–186. [Google Scholar] [CrossRef]
- Kim, S.-K.; Kim, S.-S.; Kim, K.-S.; Chung, Y.-R.; Kim, C.-H. New Antibiotics Produced by Streptomyces melanosporofaciens II. Antimicrobial Activities and Isolation, Purification, and Structure Determination of the Active Compound. Kor. J. Microbiol. Biotechnol. 1991, 19, 235–241. [Google Scholar]
- Nguyen, D.T.M.; Nguyen, D.H.; Hwa-La, L.; Lee, H.; Shin, J.-H.; Kim, E.-I. Inhibition of Melanogenesis by Dioctyl Phthalate Isolated from Nigella glandulifera Freyn. J. Microbiol. Biotechnol. 2007, 17, 1585–1590. [Google Scholar]
- U.S. Department of Agriculture Agricultural Research Service Dr. Duke’s Phytochemical and Ethnobotanical Databases. Available online: http://phytochem.nal.usda.gov/ (accessed on 25 October 2021).
- Hamazaki, K.; Suzuki, N.; Kitamura, K.-i.; Hattori, A.; Nagasawa, T.; Itomura, M.; Hamazaki, T. Is vaccenic acid (18:1t n-7) associated with an increased incidence of hip fracture? An explanation for the calcium paradox. Prostaglandins Leukot. Essent. Fat. Acids 2016, 109, 8–12. [Google Scholar] [CrossRef]
- Bibi, S.N.; Gokhan, Z.; Rajesh, J.; Mahomoodally, M.F. Fungal endophytes associated with mangroves—Chemistry and biopharmaceutical potential. S. Afr. J. Bot. 2020, 134, 187–212. [Google Scholar] [CrossRef]
- Dat, T.T.H.; Oanh, P.T.T. In vitro antioxidant, α-amylase and α-glucosidase inhibitory activities of endophytic bacteria from the roots of the mangrove plant Rhizophora stylosa Griffith. Acad. J. Biol. 2021, 43, 125–135. [Google Scholar] [CrossRef]
- Sebastianes, F.L.S.; de Azevedo, J.L.; Lacava, P.T. Diversity and Biotechnological Potential of Endophytic Microorganisms Associated with Tropical Mangrove Forests. In Diversity and Benefits of Microorganisms from the Tropics; de Azevedo, J.L., Quecine, M.C., Eds.; Springer International Publishing: Cham, Switzerland, 2017; pp. 37–56. [Google Scholar]
- Castro, R.A.; Dourado, M.N.; Almeida, J.R.D.; Lacava, P.T.; Nave, A.; Melo, I.S.d.; Azevedo, J.L.D.; Quecine, M.C. Mangrove endophyte promotes reforestation tree (Acacia polyphylla) growth. Braz. J. Microbiol. 2018, 49, 59–66. [Google Scholar] [CrossRef] [PubMed]
- Dat, T.T.H.; Oanh, P.T.T.; Tam, V.T.T.; Anh, H.L.T. Antimicrobial and antioxidant activity of bacterial endophytes isolated from leaves of the mangrove plant Rhizophora stylosa. Acad. J. Biol. 2019, 41, 91–99. [Google Scholar] [CrossRef]
- Hernández-González, I.L.; Moreno-Hagelsieb, G.; Olmedo-Álvarez, G. Environmentally-driven gene content convergence and the Bacillus phylogeny. BMC Evol. Biol. 2018, 18, 148. [Google Scholar] [CrossRef] [PubMed]
- Ian, E.; Malko, D.B.; Sekurova, O.N.; Bredholt, H.; Rückert, C.; Borisova, M.E.; Albersmeier, A.; Kalinowski, J.; Gelfand, M.S.; Zotchev, S.B. Genomics of Sponge-Associated Streptomyces spp. Closely Related to Streptomyces albus J1074: Insights into Marine Adaptation and Secondary Metabolite Biosynthesis Potential. PLoS ONE 2014, 9, e96719. [Google Scholar]
- Versluis, D.; Nijsse, B.; Naim, M.A.; Koehorst, J.J.; Wiese, J.; Imhoff, J.F.; Schaap, P.J.; van Passel, M.W.J.; Smidt, H.; Sipkema, D. Comparative Genomics Highlights Symbiotic Capacities and High Metabolic Flexibility of the Marine Genus Pseudovibrio. Genome Biol. Evol. 2018, 10, 125–142. [Google Scholar] [CrossRef] [Green Version]
- Steil, L.; Hoffmann, T.; Budde, I.; Völker, U.; Bremer, E. Genome-Wide Transcriptional Profiling Analysis of Adaptation of Bacillus subtilis to High Salinity. J. Bacteriol. 2003, 185, 6358. [Google Scholar] [CrossRef] [Green Version]
- Tian, X.; Zhang, Z.; Yang, T.; Chen, M.; Li, J.; Chen, F.; Yang, J.; Li, W.; Zhang, B.; Zhang, Z.; et al. Comparative Genomics Analysis of Streptomyces Species Reveals Their Adaptation to the Marine Environment and Their Diversity at the Genomic Level. Front. Microbiol. 2016, 7, 998. [Google Scholar] [CrossRef]
- Bondarev, V.; Richter, M.; Romano, S.; Piel, J.; Schwedt, A.; Schulz-Vogt, H.N. The genus Pseudovibrio contains metabolically versatile bacteria adapted for symbiosis. Environ. Microbiol. 2013, 15, 2095–2113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belbahri, L.; Chenari Bouket, A.; Rekik, I.; Alenezi, F.N.; Vallat, A.; Luptakova, L.; Petrovova, E.; Oszako, T.; Cherrad, S.; Vacher, S.; et al. Comparative Genomics of Bacillus amyloliquefaciens Strains Reveals a Core Genome with Traits for Habitat Adaptation and a Secondary Metabolites Rich Accessory Genome. Front. Microbiol. 2017, 8, 1438. [Google Scholar] [CrossRef] [PubMed]
- Rukachaisirikul, V.; Rodglin, A.; Sukpondma, Y.; Phongpaichit, S.; Buatong, J.; Sakayaroj, J. Phthalide and Isocoumarin Derivatives Produced by an Acremonium sp. Isolated from a Mangrove Rhizophora apiculata. J. Nat. Prod. 2012, 75, 853–858. [Google Scholar] [CrossRef]
- Klaiklay, S.; Rukachaisirikul, V.; Tadpetch, K.; Sukpondma, Y.; Phongpaichit, S.; Buatong, J.; Sakayaroj, J. Chlorinated chromone and diphenyl ether derivatives from the mangrove-derived fungus Pestalotiopsis sp. PSU-MA69. Tetrahedron 2012, 68, 2299–2305. [Google Scholar] [CrossRef]
- Klaiklay, S.; Rukachaisirikul, V.; Phongpaichit, S.; Pakawatchai, C.; Saithong, S.; Buatong, J.; Preedanon, S.; Sakayaroj, J. Anthraquinone derivatives from the mangrove-derived fungus Phomopsis sp. PSU-MA214. Phytochem. Lett. 2012, 5, 738–742. [Google Scholar] [CrossRef]
- Klaiklay, S.; Rukachaisirikul, V.; Phongpaichit, S.; Buatong, J.; Preedanon, S.; Sakayaroj, J. Flavodonfuran: A new difuranylmethane derivative from the mangrove endophytic fungus Flavodon flavus PSU-MA201. Nat. Prod. Res. 2013, 27, 1722–1726. [Google Scholar] [CrossRef] [PubMed]
- Dat, T.T.H.; Cuc, N.T.; Cuong, P.V.; Smidt, H.; Sipkema, D. Diversity and Antimicrobial Activity of Vietnamese Sponge-Associated Bacteria. Mar. Drugs 2021, 19, 353. [Google Scholar] [CrossRef]
- Lane, D.J. 16S/23S rRNA Sequencing. In Nucleic Acid Techniques in Bacterial Systematics; Stackebrandt, E., Goodfellow, M., Eds.; John Wiley & Sons: New York, NY, USA, 1991; pp. 115–175. [Google Scholar]
- Nguyen, M.T.T.; Awale, S.; Tezuka, Y.; Tran, Q.L.; Watanabe, H.; Kadota, S. Xanthine Oxidase Inhibitory Activity of Vietnamese Medicinal Plants. Biol. Pharm. Bull. 2004, 27, 1414–1421. [Google Scholar] [CrossRef] [Green Version]
- Skehan, P.; Storeng, R.; Scudiero, D.; Monks, A.; McMahon, J.; Vistica, D.; Warren, J.T.; Bokesch, H.; Kenney, S.; Boyd, M.R. New Colorimetric Cytotoxicity Assay for Anticancer-Drug Screening. JNCI J. Natl. Cancer Inst. 1990, 82, 1107–1112. [Google Scholar] [CrossRef] [PubMed]
- Chin, C.-S.; Peluso, P.; Sedlazeck, F.J.; Nattestad, M.; Concepcion, G.T.; Clum, A.; Dunn, C.; O’Malley, R.; Figueroa-Balderas, R.; Morales-Cruz, A.; et al. Phased diploid genome assembly with single-molecule real-time sequencing. Nat. Methods 2016, 13, 1050–1054. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mikheenko, A.; Valin, G.; Prjibelski, A.; Saveliev, V.; Gurevich, A. Icarus: Visualizer for de novo assembly evaluation. Bioinformatics 2016, 32, 3321–3323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manni, M.; Berkeley, M.R.; Seppey, M.; Simão, F.A.; Zdobnov, E.M. BUSCO Update: Novel and Streamlined Workflows along with Broader and Deeper Phylogenetic Coverage for Scoring of Eukaryotic, Prokaryotic, and Viral Genomes. Mol. Biol. Evol. 2021, 38, 4647–4654. [Google Scholar] [CrossRef]
- Seemann, T. Prokka: Rapid prokaryotic genome annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef]
- Blin, K.; Shaw, S.; Steinke, K.; Villebro, R.; Ziemert, N.; Lee, S.Y.; Medema, M.H.; Weber, T. antiSMASH 5.0: Updates to the secondary metabolite genome mining pipeline. Nucleic Acids Res. 2019, 47, W81–W87. [Google Scholar] [CrossRef] [Green Version]
- Ziemert, N.; Podell, S.; Penn, K.; Badger, J.H.; Allen, E.; Jensen, P.R. The Natural Product Domain Seeker NaPDoS: A Phylogeny Based Bioinformatic Tool to Classify Secondary Metabolite Gene Diversity. PLoS ONE 2012, 7, e34064. [Google Scholar]
- Navarro-Muñoz, J.C.; Selem-Mojica, N.; Mullowney, M.W.; Kautsar, S.A.; Tryon, J.H.; Parkinson, E.I.; De Los Santos, E.L.C.; Yeong, M.; Cruz-Morales, P.; Abubucker, S.; et al. A computational framework to explore large-scale biosynthetic diversity. Nat. Chem. Biol. 2020, 16, 60–68. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| Strain ID | Identification by 16S rRNA Gene Sequence | NCBI BLAST Identity (%) | Accession Number |
|---|---|---|---|
| RAR_GA_12 | Streptomyces sp. | 99.8 | MT229086 |
| RAR_GA_16 | Bacillus sp. | 100 | OK649239 |
| RAR_WA_18 | Bacillus sp. | 99.8 | MT229092 |
| RAR_M1_23 | Pseudovibrio sp. | 99.7 | MT229094 |
| RAR_M1_26 | Bacillus sp. | 99.6 | MT229093 |
| RAR_GA_31 | Microbacterium sp. | 99.9 | MT229099 |
| RAR_WA_32 | Rossellomorea vietnamensis | 100 | OK649240 |
| RAR_M1_36 | Brevibacterium sp. | 99.9 | MT229096 |
| RAR_M1_41 | Microbulbifer sp. | 99.8 | MT229085 |
| RAR_GA_42 | Micrococcus sp. | 99.9 | MT229088 |
| RAR_M1_44 | Bacillus sp. | 100 | OK649238 |
| RAR_M1_45 | Bacillus sp. | 99.9 | MT229100 |
| RAR_M1_49 | Paracoccus sp. | 99.9 | MT229090 |
| RAR_WA_50 | Bacillus sp. | 99.8 | MT229104 |
| RAR_M1_53 | Streptomyces sp. | 99.9 | MT229089 |
| RAR_M1_54 | Streptomyces sp. | 100 | MT229103 |
| RAR_GA_57 | Streptomyces sp. | 99.9 | MT229098 |
| RAR_M1_58 | Bacillus velezensis | 100 | MT229097 |
| RAR_M1_60 | Bacillus sp. | 99.9 | MT229102 |
| RAR_M1_61 | Bacillus velezensis | 100 | MT229087 |
| RAR_GA_63 | Streptomyces sp. | 99.7 | MT229095 |
| RAR_GA_64 | Streptomyces sp. | 100 | MT229101 |
| RAR_M1_66 | Microbacterium sp. | 99.8 | MT229091 |
| Strain ID | S. aureus | E. faecalis | E. coli | P. aegurinosa | C. albicans |
|---|---|---|---|---|---|
| RAR_GA_12 | 64 | 128 | - | - | 64 |
| RAR_GA_16 | 16 | 32 | 256 | - | 32 |
| RAR_WA_18 | - | 64 | 128 | 256 | 256 |
| RAR_M1_23 | 128 | - | - | - | - |
| RAR_M1_26 | - | - | - | - | - |
| RAR_GA_31 | - | - | - | 64 | 128 |
| RAR_WA_32 | - | - | 32 | 64 | 128 |
| RAR_M1_36 | 64 | 32 | 128 | - | - |
| RAR_M1_41 | - | - | - | - | |
| RAR_GA_42 | - | - | - | - | - |
| RAR_M1_44 | 32 | 64 | 128 | - | 64 |
| RAR_M1_45 | - | 128 | 128 | 256 | - |
| RAR_M1_49 | - | - | - | - | - |
| RAR_WA_50 | 32 | - | - | 64 | 64 |
| RAR_M1_53 | - | 64 | - | 256 | - |
| RAR_M1_54 | - | 128 | - | - | - |
| RAR_GA_57 | - | 32 | - | 16 | 64 |
| RAR_M1_58 | - | 256 | - | - | |
| RAR_M1_60 | 128 | - | 128 | - | 32 |
| RAR_M1_61 | - | - | - | - | - |
| RAR_GA_63 | 64 | 256 | - | - | |
| RAR_GA_64 | - | - | 64 | - | |
| RAR_M1_66 | 128 | - | - | 64 | 128 |
| Ciprofloxacin | 1 | 2 | 0.5 | 0.5 | - |
| Fluconazole | 2 |
| Strain ID | DPPH Radical Scavenging Activity | ABTS Radical Scavenging Activity |
|---|---|---|
| RAR_GA_12 | >100 | >100 |
| RAR_GA_16 | 72.24 ± 5.26 | 56.29 ± 4.61 |
| RAR_WA_18 | >100 | >100 |
| RAR_M1_23 | 61.23 ± 3.26 | 76.32 ± 3.51 |
| RAR_M1_26 | >100 | >100 |
| RAR_GA_31 | 72.36 ± 3.95 | >100 |
| RAR_WA_32 | 43.52 ± 3.87 | 66.43 ± 5.25 |
| RAR_M1_36 | 44.34 ± 2.76 | 64.51 ± 3.26 |
| RAR_M1_41 | >100 | >100 |
| RAR_GA_42 | 51.64 ± 2.68 | 76.29 ± 3.75 |
| RAR_M1_44 | 81.34 ± 6.18 | 77.35 ± 5.82 |
| RAR_M1_45 | >100 | >100 |
| RAR_M1_49 | >100 | >100 |
| RAR_WA_50 | >100 | 81.45 ± 3.64 |
| RAR_M1_53 | >100 | >100 |
| RAR_M1_54 | >100 | >100 |
| RAR_GA_57 | >100 | 59.43 ± 2.68 |
| RAR_M1_58 | 88.32 ± 4.13 | 67.48 ± 2.46 |
| RAR_M1_60 | >100 | >100 |
| RAR_M1_61 | 65.23 ± 3.23 | 77.21 ± 3.24 |
| RAR_GA_63 | >100 | >100 |
| RAR_GA_64 | >100 | >100 |
| RAR_M1_66 | >100 | 71.35 ± 3.57 |
| Ascorbic acid | 29.14 ± 4.27 | 26.35 ± 3.24 |
| Strain ID | α-Amylase Inhibition | α-Glucosidase Inhibition | XO Inhibition |
|---|---|---|---|
| RAR_GA_12 | >200 | 74.26 ± 4.12 | >100 |
| RAR_GA_16 | 73.27 ± 3.45 | 85.73 ± 5.26 | 94.36 ± 4.74 |
| RAR_WA_18 | >200 | >200 | 54.57 ± 2.53 |
| RAR_M1_23 | 131.36 ± 5.41 | >200 | >100 |
| RAR_M1_26 | >200 | 73.64 ± 3.78 | >100 |
| RAR_GA_31 | 64.42 ± 3.35 | 81.32 ± 4.42 | >100 |
| RAR_WA_32 | 33.51 ± 4.62 | 53.68 ± 3.12 | 74.38 ± 3.81 |
| RAR_M1_36 | 51.67 ± 2.98 | >200 | 75.36 ± 3.76 |
| RAR_M1_41 | >200 | 0>200 | >100 |
| RAR_GA_42 | >200 | >200 | 82.35 ± 4.13 |
| RAR_M1_44 | 86.38 ± 5.39 | 94.14 ± 5.28 | 64.43 ± 4.56 |
| RAR_M1_45 | >200 | >200 | >100 |
| RAR_M1_49 | 58.34 ± 2.47 | 55.16 ± 4.39 | >100 |
| RAR_WA_50 | 77.64 ± 3.68 | >200 | 68.46 ± 3.47 |
| RAR_M1_53 | >200 | >200 | >100 |
| RAR_M1_54 | >200 | >200 | 74.38 ± 3.81 |
| RAR_GA_57 | 83.45 ± 4.25 | 57.38 ± 3.42 | >100 |
| RAR_M1_58 | 68.34 ± 3.42 | >200 | >100 |
| RAR_M1_60 | >200 | >200 | 71.44 ± 3.67 |
| RAR_M1_61 | >200 | 193.44 ± 6.73 | >100 |
| RAR_GA_63 | 128.34 ± 6.32 | >200 | >100 |
| RAR_GA_64 | >200 | >200 | 55.29 ± 3.27 |
| RAR_M1_66 | >200 | >200 | >100 |
| Acarbose | 89.34 ± 3.61 | 217.46 ± 6.38 | - |
| Allopurinol | - | - | 6.12 ± 0.61 |
| Strain ID | MCF-7 | A549 | Hela |
|---|---|---|---|
| RAR_GA_12 | >100 | >100 | >100 |
| RAR_GA_16 | 36.48 ± 2.63 | 89.53 ± 5.31 | 41.27 ± 3.42 |
| RAR_WA_18 | > 100 | >100 | >100 |
| RAR_M1_23 | >100 | >100 | >100 |
| RAR_M1_26 | >100 | >100 | >100 |
| RAR_GA_31 | >100 | >100 | >100 |
| RAR_WA_32 | >100 | 21.52 ± 3.22 | 57.67 ± 4.75 |
| RAR_M1_36 | 83.24 ± 4.51 | >100 | >100 |
| RAR_M1_41 | >100 | >100 | >100 |
| RAR_GA_42 | 53.17 ± 2.84 | >100 | >100 |
| RAR_M1_44 | 61.32 ± 3.22 | 51.32 ± 4.21 | 97.53 ± 5.31 |
| RAR_M1_45 | >100 | >100 | >100 |
| RAR_M1_49 | >100 | >100 | >100 |
| RAR_WA_50 | 61.32 ± 3.22 | 36.48 ± 2.32 | >100 |
| RAR_M1_53 | >100 | >100 | >100 |
| RAR_M1_54 | >100 | >100 | >100 |
| RAR_GA_57 | 77.36 ± 2.31 | >100 | >100 |
| RAR_M1_58 | >100 | >100 | >100 |
| RAR_M1_60 | >100 | >100 | >100 |
| RAR_M1_61 | >100 | >100 | >100 |
| RAR_GA_63 | >100 | >100 | >100 |
| RAR_GA_64 | >100 | >100 | >100 |
| RAR_M1_66 | >100 | >100 | >100 |
| Camptothecin | 4.75 ± 0.41 | 2.47 ± 0.36 | 3.56 ± 0.62 |
| No. | Composition | RT/min | Matching Quality | Quantity (%) | Biological Activity | Refs. |
|---|---|---|---|---|---|---|
| Bacillus sp. RAR_GA_16 | ||||||
| 1 | Hexahydro-pyrrolo[1,2-a]pyrazine-1,4-dione | 18.830 | 96 | 13.15 | Antimicrobial, antioxidant | [28,29] |
| 2 | 3-Isobutylhexahydropyrrolo[1,2-a]pyrazine-1,4-dione | 21.125 | 91 | 8.15 | Antimicrobial, nematicidal, and anti-mutagenic | [30,31,32] |
| 3 | Palmitic acid | 21.413 | 98 | 2.09 | Antibacterial | [33] |
| 4 | Diisooctyl phthalate | 22.022 | 95 | 6.90 | Antibacterial, antifungal | [34,35] |
| 5 | Linoleic acid | 23.454 | 99 | 1.57 | Antibacterial | [33,36] |
| 6 | 9(E)-Octadecenoic acid | 23.517 | 99 | 9.17 | Antibacterial | [37] |
| 7 | Stearic acid | 23.763 | 98 | 3.02 | Antibacterial | [33] |
| Rossellomorea vietnamensis RAR_WA_32 | ||||||
| 1 | Benzyl alcohol | 8.045 | 96 | 0.12 | Antibacterial | [38,39] |
| 2 | 2,3,5,6-Tetramethylpyrazine | 9.494 | 91 | 0.16 | Antioxidant, anticancer, anti-inflammatory | [40,41,42] |
| 3 | Phenylethyl alcohol | 10.225 | 97 | 0.16 | Antibacterial | [43,44] |
| 4 | Benzoic acid | 11.783 | 94 | 0.25 | Antimicrobial | [45] |
| 5 | Methyl phenylacetate | 12.022 | 94 | 0.19 | Antimicrobial | [46,47] |
| 6 | Benzeneacetic acid | 14.193 | 94 | 1.92 | Antimicrobial | [48,49] |
| 7 | Benzenepropanoic acid | 16.367 | 96 | 0.38 | Antimicrobial | [50] |
| 8 | Docosane | 20.327 | 91 | 0.09 | - | - |
| 9 | 2,4-Di-tert-butylphenol | 20.775 | 97 | 0.3 | Antifungal, antioxidant | [51,52] |
| 10 | Hexahydropyrrolo[1,2-a]pyrazine-1,4-dione | 26.144 | 96 | 0.38 | Antibacterial, antioxidant | [28,29] |
| 11 | Myristic acid | 26.351 | 94 | 0.08 | Antibacterial | [53,54] |
| 12 | Methyl 13-methylmyristate | 26.911 | 97 | 0.12 | - | - |
| 13 | 12-Methyltetradecanoic acid | 27.078 | 94 | 0.23 | Antifungal, anticancer | [55,56] |
| 14 | Pentadecanoic acid | 27.717 | 98 | 0.64 | - | - |
| 15 | Methyl palmitate | 29.702 | 95 | 0.76 | Antibacterial, anti-inflammatory | [57,58] |
| 16 | 3-Isobutylhexahydropyrrolo[1,2-a]pyrazine-1,4-dione | 29.836 | 95 | 4.56 | Antimicrobial, nematicidal, and anti-mutagenic | [30,31,32] |
| 17 | Palmitic acid | 30.482 | 99 | 5.56 | Antibacterial | [33] |
| 18 | Methyl 14-methylhexadecanoate | 31.084 | 92 | 0.18 | - | - |
| 19 | 11-Octadecenoic methyl ester | 33.090 | 95 | 0.08 | - | - |
| 20 | Methyl stearate | 33.458 | 96 | 0.14 | - | - |
| 21 | 9-Octadecenoic acid | 33.755 | 98 | 0.79 | Antibacterial | [59] |
| 22 | Stearic acid | 34.135 | 99 | 3.55 | Antibacterial | [33] |
| 23 | 1,3,3a,6,7,9a-Hexahydro-cis-cycloocta[c]furan | 35.701 | 90 | 0.23 | - | - |
| 24 | Cyclo(phe-pro) | 36.941 | 90 | 0.15 | Antimicrobial, anticancer | [60,61,62] |
| 25 | 3-Benzylhexahydropyrrolo[1,2-a]pyrazine-1,4-dione | 37.704 | 95 | 1.87 | Anti-biofilm, anti-quorum sensing | [63] |
| 26 | Dioctyl phthalate | 40.507 | 91 | 58.09 | Antibacterial, tyrosinase inhibitory | [64,65,66] |
| Bacillus sp. RAR_M1_44 | ||||||
| 1 | Palmitic acid | 21.416 | 97 | 0.51 | Antimicrobial, antioxidant | [33] |
| 2 | (Z,Z)-9,12-Octadecadienoic acid | 23.459 | 99 | 0.33 | Anti-inflammatory, nematicidal, and hepatoprotective | [67] |
| 3 | cis-Vaccenic acid | 23.521 | 99 | 1.93 | Antibacterial, hypolipidemic | [68] |
| 4 | Stearic acid | 23.764 | 99 | 0.34 | Antibacterial, antifungal | [33] |
| 5 | 1,2-Benzenedicarboxylic acid | 23.978 | 90 | 0.99 | Antifungal, anticancer | [33,36] |
| Genomic Features | RAR_GA_16 | RAR_WA_32 | RAR_M1_44 |
|---|---|---|---|
| Size of the genome assembly (bp) | 4,394,636 | 4,494,267 | 3,768,026 |
| GC content (%) | 40.69 | 44.09 | 41.38 |
| Contigs | 22 | 4 | 5 |
| Maximum Contig Length (bp) | 770,470 | 2,474,194 | 2,582,281 |
| N50 Contig Length (bp) | 501,856 | 2,474,194 | 2,582,281 |
| CDS | 4610 | 4529 | 3807 |
| rRNA | 27 | 22 | 24 |
| tRNA | 89 | 111 | 81 |
| tmRNA | 1 | 0 | 1 |
| Complete genome (%) | 96.6 | 99.5 | 99.8 |
| Cluster | Length | Types | Most Similar Known Cluster | MIBiG BGC-ID | Similarity |
|---|---|---|---|---|---|
| Bacillus sp. RAR_GA_16 | |||||
| Cluster 1 | 21,787 | LAP, bacteriocin | - | - | - |
| Cluster 2 | 14,207 | Lassopeptide | Paeninodin | BGC0001356 | 80% |
| Cluster 3 | 28,959 | Siderophore | - | - | - |
| Cluster 4 | 37,598 | T3PKS | - | - | - |
| Cluster 5 | 21,821 | Terpene | Butirosin A/butirosin B | BGC0000693 | 7% |
| Cluster 6 | 20,837 | Terpene | Carotenoid | BGC0000645 | 50% |
| Cluster 7 | 24,479 | - | S-layer glycan | BGC0000796 | 14% |
| Cluster 8 | 15,240 | - | Thaxteramide C | BGC0002025 | 7% |
| Cluster 9 | 4858 | - | Fengycin | BGC0001095 | 20% |
| Cluster 10 | 8288 | - | Capsular polysaccharide | BGC0000758 | 4% |
| Cluster 11 | 11,638 | - | - | - | - |
| Cluster 12 | 6299 | - | - | - | - |
| Cluster 13 | 3468 | - | - | - | - |
| Cluster 14 | 15,194 | - | - | - | - |
| Cluster 15 | 9297 | - | - | - | - |
| Cluster 16 | 15,779 | - | - | - | - |
| Cluster 17 | 11,263 | - | - | - | - |
| Cluster 18 | 9795 | - | - | - | - |
| Cluster 19 | 9448 | - | - | - | - |
| Cluster 20 | 16,830 | - | - | - | - |
| Cluster 21 | 7027 | - | - | - | - |
| Cluster 22 | 5280 | - | - | - | - |
| Rossellomorea vietnamensis RAR_WA_32 | |||||
| Cluster 1 | 21,869 | Terpene | Pyxidicycline A/pyxidicycline B | BGC0001940 | 6% |
| Cluster 2 | 20,443 | Saccharide | A40926 | BGC0000289 | 3% |
| Cluster 3 | 22,620 | Saccharide | Carotenoid | BGC0000645 | 33% |
| Cluster 4 | 121,332 | Saccharide | S-layer glycan | BGC0000796 | 14% |
| Cluster 5 | 50,793 | Fatty_acid, saccharide, terpene | Carotenoid | BGC0000645 | 50% |
| Cluster 6 | 19,105 | LAP, RiPP-like | - | - | - |
| Cluster 7 | 20,392 | Saccharide | - | - | - |
| Cluster 8 | 70,095 | Saccharide,T3PKS | - | - | - |
| Cluster 9 | 29,497 | Saccharide | - | - | - |
| Cluster 10 | 21,005 | Fatty_acid | - | - | - |
| Cluster 11 | 30,306 | Saccharide | - | - | - |
| Cluster 12 | 34,473 | Saccharide | - | - | - |
| Cluster 13 | 21,368 | Saccharide | - | - | - |
| Cluster 14 | 24,846 | Saccharide | - | - | - |
| Cluster 15 | 30,807 | Saccharide | - | - | - |
| Cluster 16 | 35,649 | Saccharide | - | - | - |
| Bacillus sp. RAR_M1_44 | |||||
| Cluster 1 | 28,756 | Siderophore, terpene | Carotenoid | BGC0000645 | 50% |
| Cluster 2 | 28,411 | Betalactone | Fengycin | BGC0001095 | 53% |
| Cluster 3 | 83,725 | NRPS | Lichenysin | BGC0000868 | 85% |
| Cluster 4 | 8013 | Other | Teichuronic acid | BGC0001184 | 75% |
| Cluster 5 | 41,421 | Other | Bacilysin | BGC0000381 | 85% |
| Cluster 6 | 21,877 | Terpene | - | - | - |
| Cluster 7 | 41,097 | T3PKS | - | - | - |
| Cluster 8 | 26,707 | Bacteriocin | |||
| Cluster 9 | 32,416 | Betalactone | |||
| Cluster 10 | 15,922 | - | - | - | - |
| Cluster 11 | 5255 | - | - | - | - |
| Cluster 12 | 13,700 | - | - | - | - |
| Cluster 13 | 8838 | - | - | - | - |
| Cluster 14 | 16,010 | - | - | - | - |
| Cluster 15 | 3876 | - | - | - | - |
| Cluster 16 | 9767 | - | - | - | - |
| KS/C Domain ID | Database Match ID | Percent Identity (%) | E-Value | Pathway Product |
|---|---|---|---|---|
| Bacillus sp. RAR_GA_16 | ||||
| KS1 | AmphI_Q93NX9_1mod | 42 | 4e-7 | Amphotericin |
| KS2 | SpnB_Q9ALM5_1KSB | 35 | 2e-7 | Spinosad |
| KS3 | FabF_Bacillus_FAS | 76 | 0 | Fatty acid |
| KS4 | DynE_AAN79725_ene10 | 45 | 9e-6 | Dynemicin |
| Rossellomorea vietnamensis RAR_WA_32 | ||||
| KS1 | KirAI_CAN89631_2T | 40 | 5e-7 | Kirromycin |
| KS2 | FabB_Streptomyces_FAS | 41 | 3e-6 | Fatty acid |
| KS3 | MycAIII_Q83WE8_2KSB | 44 | 1e-6 | Mycinamicin |
| KS4 | FabF_Bacillus_FAS | 71 | 0.0 | Fatty acid |
| KS5 | RifB_O52545_1mod | 26 | 1e-6 | Rifamycin |
| Bacillus sp. RAR_M1_44 | ||||
| KS1 | Myca_YP881572_1KSB | 40 | 2e-6 | Mycocerosic acid |
| KS2 | FabF_Bacillus_FAS | 48 | 1e-108 | Fatty acid |
| KS3 | FabF_Bacillus_FAS | 84 | 0.0 | Fatty acid |
| C1 | Liche1_C1_start | 51 | 3e-135 | Lychenicin |
| C2 | Liche1_C1_start | 24 | 4e-25 | Lychenicin |
| C3 | Liche1_C1_start | 22 | 6e-18 | Lychenicin |
| C4 | Surfa5_C1_DCL | 57 | 5e-151 | Surfactin |
| C5 | Surfa5_C1_DCL | 24 | 2e-24 | Surfactin |
| C6 | Surfa5_C1_DCL | 22 | 3e-12 | Surfactin |
| C7 | Surfa5_C1_DCL | 24 | 3e-8 | Surfactin |
| C8 | Liche3_C1_DCL | 58 | 5e-151 | Lychenicin |
| C9 | Act3_C3_LCL | 39 | 6e-86 | Actinomycin |
| C10 | Act3_C3_LCL | 23 | 2e-20 | Actinomycin |
| C11 | Liche1_C3_LCL | 38 | 2e-77 | Lychenicin |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dat, T.T.H.; Oanh, P.T.T.; Cuong, L.C.V.; Anh, L.T.; Minh, L.T.H.; Ha, H.; Lam, L.T.; Cuong, P.V.; Anh, H.L.T. Pharmacological Properties, Volatile Organic Compounds, and Genome Sequences of Bacterial Endophytes from the Mangrove Plant Rhizophora apiculata Blume. Antibiotics 2021, 10, 1491. https://doi.org/10.3390/antibiotics10121491
Dat TTH, Oanh PTT, Cuong LCV, Anh LT, Minh LTH, Ha H, Lam LT, Cuong PV, Anh HLT. Pharmacological Properties, Volatile Organic Compounds, and Genome Sequences of Bacterial Endophytes from the Mangrove Plant Rhizophora apiculata Blume. Antibiotics. 2021; 10(12):1491. https://doi.org/10.3390/antibiotics10121491
Chicago/Turabian StyleDat, Ton That Huu, Phung Thi Thuy Oanh, Le Canh Viet Cuong, Le Tuan Anh, Le Thi Hong Minh, Hoang Ha, Le Tung Lam, Pham Viet Cuong, and Hoang Le Tuan Anh. 2021. "Pharmacological Properties, Volatile Organic Compounds, and Genome Sequences of Bacterial Endophytes from the Mangrove Plant Rhizophora apiculata Blume" Antibiotics 10, no. 12: 1491. https://doi.org/10.3390/antibiotics10121491