
Carbapenemase Producing Klebsiella pneumoniae (KPC): What Is the Best MALDI-TOF MS Detection Method




Abstract
:1. Introduction
2. Results and Discussion
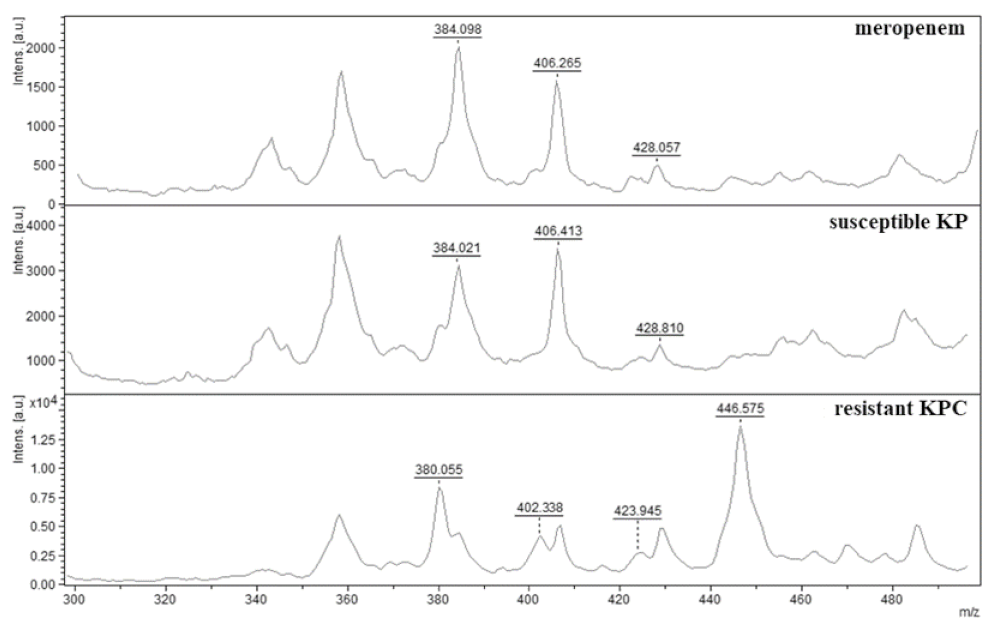
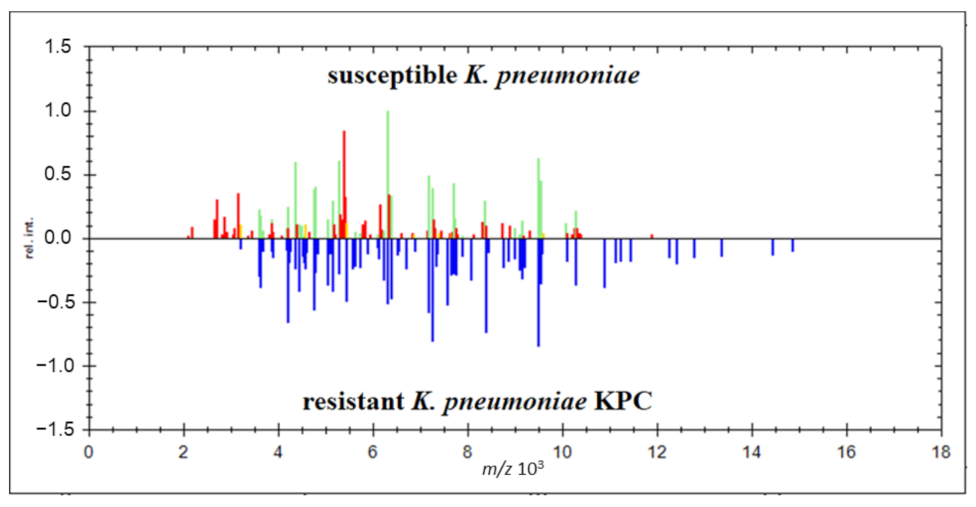
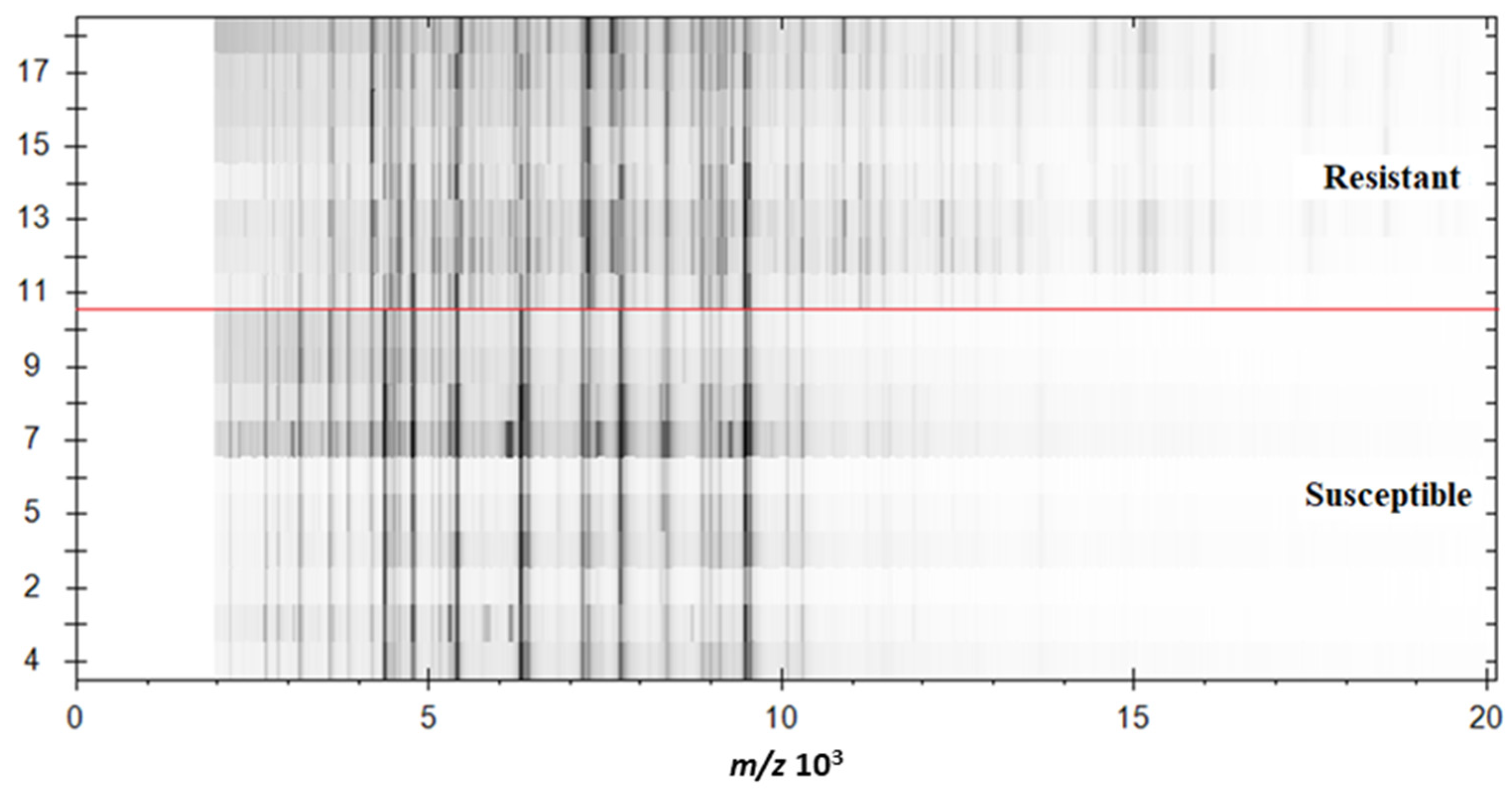
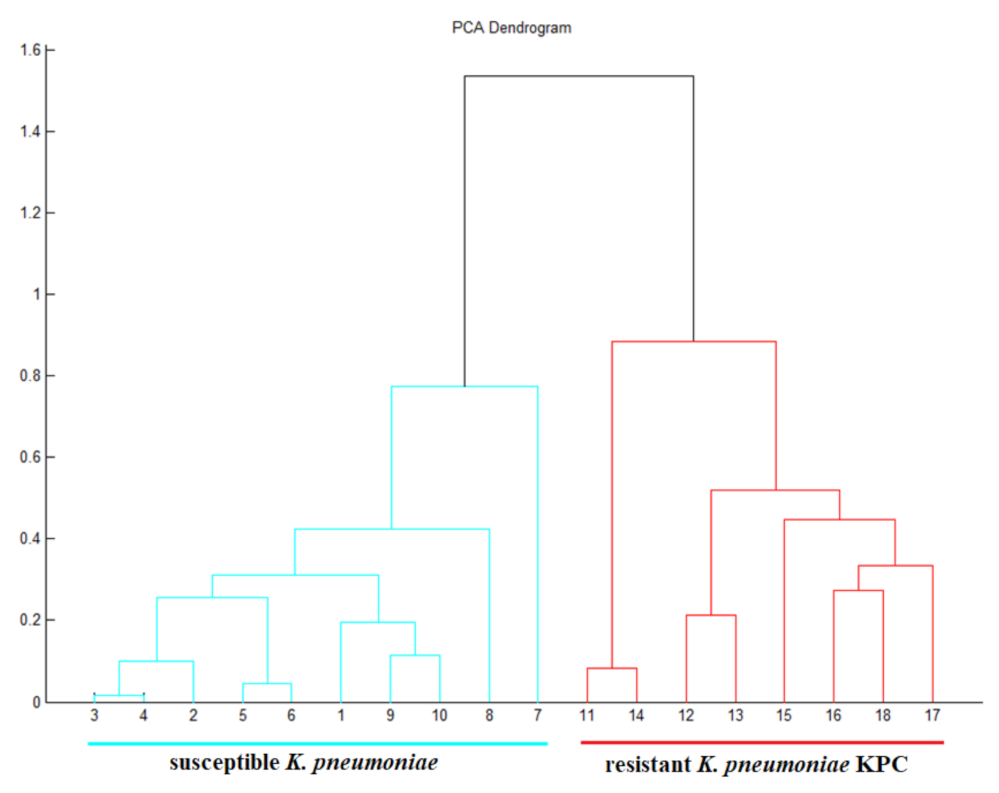
2.1. Verification of Carbapenemase-Producing Klebsiella pneumoniae (KPC)
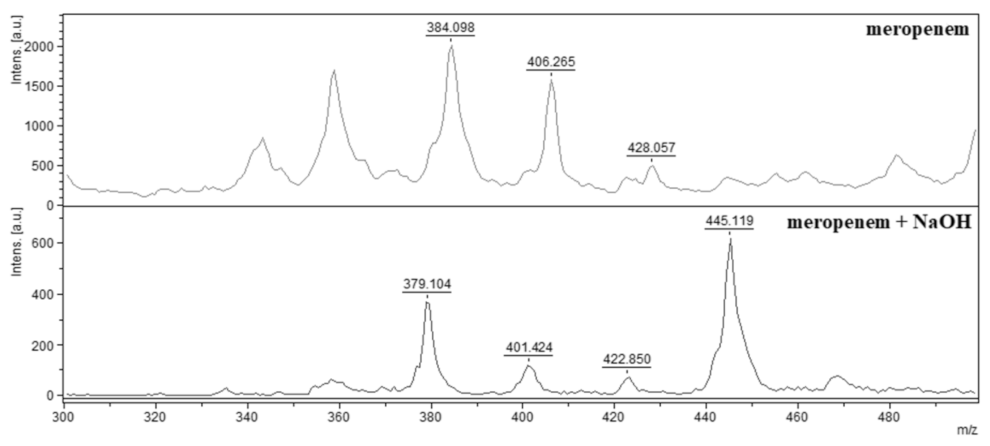
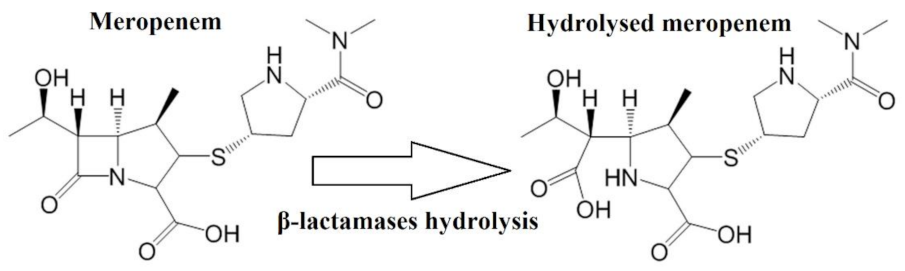
2.2. Indirect Detection of Enzymatic Hydrolysis
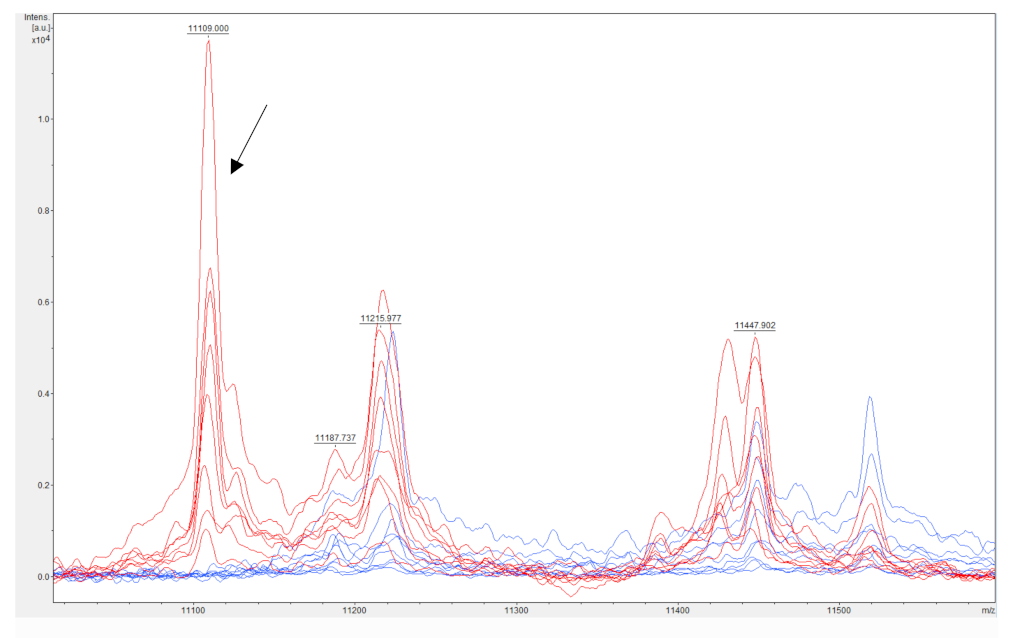
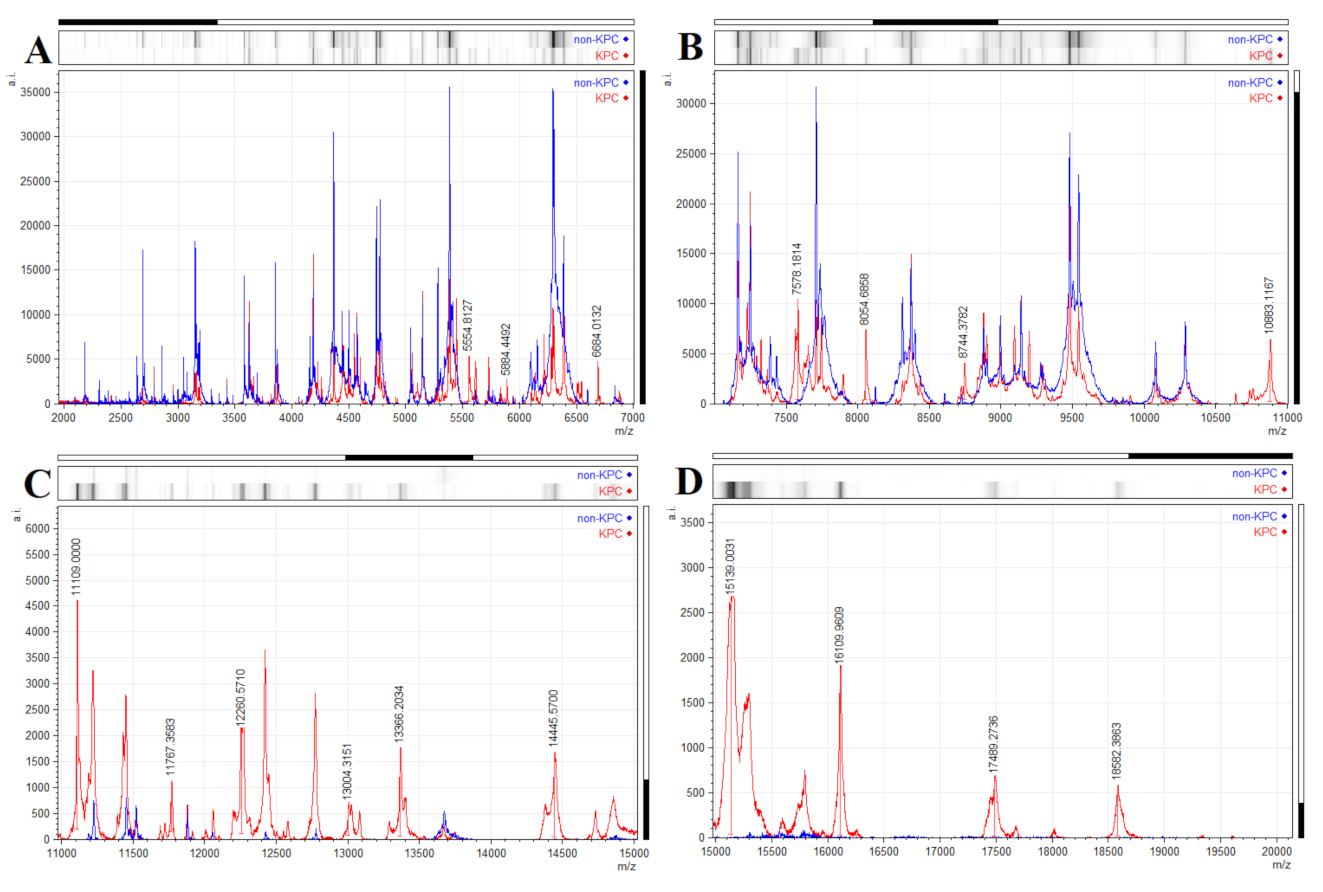
2.3. Identification of blaKPC Accompanying Peak with 11,109 Da
2.4. Identification Based on the Proteins Fingerprint Database
3. Materials and Methods
3.1. Bacterial Isolates and Cultivation
3.2. Identification of Klebsiella pneumoniae
3.3. Testing of Klebsiella pneumoniae Carbapenem Resistance
3.3.1. Antimicrobial Susceptibility Testing
3.3.2. Carbapenemase-Production Screening
3.3.3. KPC Genetic Confirmation
3.3.4. Indirect Detection of Enzymatic Hydrolysis
3.3.5. MALDI-TOF MS Analysis of Meropenem
3.3.6. Ampicillin and Meropenem Hydrolysis Assay
3.3.7. Analysis of Spectra
3.3.8. Calibration of MALDI-TOF MS
3.3.9. Identification of blaKPC Accompanying Peak with 11,109 Da
3.3.10. MALDI-TOF MS Samples Preparing
3.3.11. MALDI-TOF MS Spectrum Obtaining
3.3.12. Mass Spectra Analysis
3.3.13. Identification Based on the Proteins Fingerprint Database
3.3.14. Analysis of Mass Spectra
3.3.15. Creation of Local K. pneumoniae Database and Verification of Reproducibility
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Arnold, R.S.; Thom, K.A.; Sharma, S.; Phillips, M.; Kristie Johnson, J.; Morgan, D.J. Emergence of Klebsiella Pneumoniae Carbapenemase-Producing Bacteria. South. Med. J. 2011, 104, 40–45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, H.S.; Chon, J.W.; Kim, Y.J.; Kim, D.H.; Kim, M.S.; Seo, K.H. Prevalence and Characterization of Extended-Spectrum-β-Lactamase-Producing Escherichia Coli and Klebsiella Pneumoniae in Ready-to-Eat Vegetables. Int. J. Food Microbiol. 2015, 207, 83–86. [Google Scholar] [CrossRef] [PubMed]
- Romyasamit, C.; Sornsenee, P.; Chimplee, S.; Yuwalaksanakun, S.; Wongprot, D.; Saengsuwan, P. Prevalence and Characterization of Extended-Spectrum -Lactamaseproducing Escherichia Coli and Klebsiella Pneumoniae Isolated from Raw Vegetables Retailed in Southern Thailand. PeerJ 2021, 9, e11787. [Google Scholar] [CrossRef]
- Zekar, F.M.; Granier, S.A.; Touati, A.; Millemann, Y. Occurrence of Third-Generation Cephalosporins-Resistant Klebsiella Pneumoniae in Fresh Fruits and Vegetables Purchased at Markets in Algeria. Microb. Drug Resist. 2020, 26. [Google Scholar] [CrossRef]
- Struve, C.; Krogfelt, K.A. Pathogenic Potential of Environmental Klebsiella Pneumoniae Isolates. Environ. Microbiol. 2004, 6, 353–359. [Google Scholar] [CrossRef] [PubMed]
- Siu, L.K.; Yeh, K.M.; Lin, J.C.; Fung, C.P.; Chang, F.Y. Klebsiella Pneumoniae Liver Abscess: A New Invasive Syndrome. Lancet Infect. Dis. 2012, 12, 881–887. [Google Scholar] [CrossRef]
- Ghaith, D.M.; Zafer, M.M.; Said, H.M.; Elanwary, S.; Elsaban, S.; Al-Agamy, M.H.; Bohol, M.F.F.; Bendary, M.M.; Al-Qahtani, A.; Al-Ahdal, M.N. Genetic Diversity of Carbapenem-Resistant Klebsiella Pneumoniae Causing Neonatal Sepsis in Intensive Care Unit, Cairo, Egypt. Eur. J. Clin. Microbiol. Infect. Dis. 2020, 39, 583–591. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.F.; Anderson, D.J.; Paterson, D.L. Overview of the Epidemiology and the Threat of Klebsiella Pneumoniae Carbapenemases (KPC) Resistance. Infect. Drug Resist. 2012, 5, 133–141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coulson, R.A.A.; Frere, F.; Ghuysen, J.; Joris, B.; Forsman, M.; Levesque, R.; Tiraby, G.; Waley, S. A Standard Numbering Scheme for the Class A Beta-Lactamases. Biochem. J. 1991, 276, 1145. [Google Scholar]
- Doern, C.D.; Michael Dunne, W.; Burnham, C.A.D. Detection of Klebsiella Pneumoniae Carbapenemase (KPC) Production in Non-Klebsiella Pneumoniae Enterobacteriaceae Isolates by Use of the Phoenix, Vitek 2, and Disk Diffusion Methods. J. Clin. Microbiol. 2011, 49, 1143–1147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giske, C.G.; Gezelius, L.; Samuelsen, M.; Warner, M.; Sundsfjord, A.; Woodford, N. A Sensitive and Specific Phenotypic Assay for Detection of Metallo-β-Lactamases and KPC in Klebsiella Pneumoniae with the Use of Meropenem Disks Supplemented with Aminophenylboronic Acid, Dipicolinic Acid and Cloxacillin. Clin. Microbiol. Infect. 2011, 17, 552–556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dortet, L.; Poirel, L.; Nordmann, P. Rapid Identification of Carbapenemase Types in Enterobacteriaceae and Pseudomonas Spp. by Using a Biochemical Test. Antimicrob. Agents Chemother. 2012, 56, 6437–6440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van der Zwaluw, K.; de Haan, A.; Pluister, G.N.; Bootsma, H.J.; de Neeling, A.J.; Schouls, L.M. The Carbapenem Inactivation Method (CIM), a Simple and Low-Cost Alternative for the Carba NP Test to Assess Phenotypic Carbapenemase Activity in Gram-Negative Rods. PLoS ONE 2015, 10, e0123690. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hrabák, J.; Študentová, V.; Walková, R.; Žemličková, H.; Jakubů, V.; Chudáčková, E.; Gniadkowski, M.; Pfeifer, Y.; Perry, J.D.; Wilkinson, K.; et al. Detection of NDM-1, VIM-1, KPC, OXA-48, and OXA-162 Carbapenemases by Matrix-Assisted Laser Desorption Ionization-Time of Flight Mass Spectrometry. J. Clin. Microbiol. 2012, 50, 2441–2443. [Google Scholar] [CrossRef] [Green Version]
- Pasteran, F.; Denorme, L.; Ote, I.; Gomez, S.; de Belder, D.; Glupczynski, Y.; Bogaerts, P.; Ghiglione, B.; Power, P.; Mertens, P.; et al. Rapid Identification of OXA-48 and OXA-163 Subfamilies in Carbapenem-Resistant Gram-Negative Bacilli with a Novel Immunochromatographic Lateral Flow Assay. J. Clin. Microbiol. 2016, 54, 2832–2836. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giske, C.G.; Martinez-Martinez, L.; Cantón Spain, R.; Stefani, S.; Skov, R.; Glupczynski, Y.; Nordmann, P.; Wootton, M.; Miriagou, V.; Skov Simonsen, G. EUCAST Guidelines for Detection of Resistance Mechanisms and Specific Resistances of Clinical and/or Epidemiological Importance. Eur. Comm. Antimicrob. Susceptibility Test. 2017, 11, 1423. [Google Scholar]
- Yaman, G.; Akyar, I.; Can, S. Evaluation of the MALDI TOF-MS Method for Identification of Candida Strains Isolated from Blood Cultures. Diagn. Microbiol. Infect. Dis. 2012, 73, 65–67. [Google Scholar] [CrossRef]
- Karger, A. Current Developments to Use Linear MALDI-TOF Spectra for the Identification and Typing of Bacteria and the Characterization of Other Cells/Organisms Related to Infectious Diseases. Proteom.-Clin. Appl. 2016, 10, 982–993. [Google Scholar] [CrossRef] [PubMed]
- Johnson, J.S.; Spakowicz, D.J.; Hong, B.Y.; Petersen, L.M.; Demkowicz, P.; Chen, L.; Leopold, S.R.; Hanson, B.M.; Agresta, H.O.; Gerstein, M.; et al. Evaluation of 16S RRNA Gene Sequencing for Species and Strain-Level Microbiome Analysis. Nat. Commun. 2019, 10, 5029. [Google Scholar] [CrossRef] [Green Version]
- Homolka, S.; Projahn, M.; Feuerriegel, S.; Ubben, T.; Diel, R.; Nübel, U.; Niemann, S. High Resolution Discrimination of Clinical Mycobacterium Tuberculosis Complex Strains Based on Single Nucleotide Polymorphisms. PLoS ONE 2012, 7, e39855. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leonard, S.R.; Mammel, M.K.; Lacher, D.W.; Elkins, C.A. Strain-Level Discrimination of Shiga Toxin-Producing Escherichia Coli in Spinach Using Metagenomic Sequencing. PLoS ONE 2016, 11, e0167870. [Google Scholar] [CrossRef]
- Quainoo, S.; Coolen, J.P.M.; van Hijum, S.A.F.T.; Huynen, M.A.; Melchers, W.J.G.; van Schaik, W.; Wertheim, H.F.L. Whole-Genome Sequencing of Bacterial Pathogens: The Future of Nosocomial Outbreak Analysis. Clin. Microbiol. Rev. 2017, 30, 112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jolley, K.A.; Bliss, C.M.; Bennett, J.S.; Bratcher, H.B.; Brehony, C.; Colles, F.M.; Wimalarathna, H.; Harrison, O.B.; Sheppard, S.K.; Cody, A.J.; et al. Ribosomal Multilocus Sequence Typing: Universal Characterization of Bacteria from Domain to Strain. Microbiology 2012, 158, 1005–1015. [Google Scholar] [CrossRef] [PubMed]
- Manzoor, S.; Moncayo, S.; Navarro-Villoslada, F.; Ayala, J.A.; Izquierdo-Hornillos, R.; de Villena, F.J.M.; Caceres, J.O. Rapid Identification and Discrimination of Bacterial Strains by Laser Induced Breakdown Spectroscopy and Neural Networks. Talanta 2014, 121, 65–70. [Google Scholar] [CrossRef]
- Alamer, S.; Eissa, S.; Chinnappan, R.; Herron, P.; Zourob, M. Rapid Colorimetric Lactoferrin-Based Sandwich Immunoassay on Cotton Swabs for the Detection of Foodborne Pathogenic Bacteria. Talanta 2018, 185, 275–280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vidic, J.; Vizzini, P.; Manzano, M.; Kavanaugh, D.; Ramarao, N.; Zivkovic, M.; Radonic, V.; Knezevic, N.; Giouroudi, I.; Gadjanski, I. Point-of-Need DNA Testing for Detection of Foodborne Pathogenic Bacteria. Sensors 2019, 19, 1100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kántor, A.; Hutková, J.; Petrová, J.; Hleba, L.; Kačániová, M. Antimicrobial Activity of Pulcherrimin Pigment Produced by Metschnikowia Pulcherrima against Various Yeast Species. J. Microbiol. Biotechnol. Food Sci. 2016, 5, 282–285. [Google Scholar] [CrossRef] [Green Version]
- Hleba, L.; Kmeť, V.; Tóth, T.; Kačániová, M. Resistance in Bacteria and Indirect Beta-Lactamase Detection in E. Coli Isolated from Culex Pipiens Detected by Matrix-Assisted Laser Desorption Ionization Time of Flight Mass Spectrometry. J. Environ. Sci. Health-Part B Pestic. Food Contam. Agric. Wastes 2017, 52, 64–69. [Google Scholar] [CrossRef] [PubMed]
- Hleba, L.; Charousova, I.; Cisarova, M.; Kovacik, A.; Kormanec, J.; Medo, J.; Bozik, M.; Javorekova, S. Rapid Identification of Streptomyces Tetracycline Producers by MALDI-TOF Mass Spectrometry. J. Environ. Sci. Health-Part A Toxic/Hazard. Subst. Environ. Eng. 2018, 53, 1083–1093. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Shan, M.; Zhu, Z.; Mao, X.; Yan, M.; Chen, Y.; Zhu, Q.; Li, H.; Gu, B. Application of MALDI-TOF MS to Rapid Identification of Anaerobic Bacteria. BMC Infect. Dis. 2019, 19, 4584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hou, T.Y.; Chiang-Ni, C.; Teng, S.H. Current Status of MALDI-TOF Mass Spectrometry in Clinical Microbiology. J. Food Drug Anal. 2019, 27, 404–414. [Google Scholar] [CrossRef]
- Siegrist, T.J.; Anderson, P.D.; Huen, W.H.; Kleinheinz, G.T.; McDermott, C.M.; Sandrin, T.R. Discrimination and Characterization of Environmental Strains of Escherichia Coli by Matrix-Assisted Laser Desorption/Ionization Time-of-Flight Mass Spectrometry (MALDI-TOF-MS). J. Microbiol. Methods 2007, 68, 554–562. [Google Scholar] [CrossRef] [PubMed]
- Hettick, J.M.; Kashon, M.L.; Slaven, J.E.; Ma, Y.; Simpson, J.P.; Siegel, P.D.; Mazurek, G.N.; Weissman, D.N. Discrimination of Intact Mycobacteria at the Strain Level: A Combined MALDI-TOF MS and Biostatistical Analysis. Proteomics 2006, 6, 6416–6425. [Google Scholar] [CrossRef]
- Vargha, M.; Takáts, Z.; Konopka, A.; Nakatsu, C.H. Optimization of MALDI-TOF MS for Strain Level Differentiation of Arthrobacter Isolates. J. Microbiol. Methods 2006, 66, 399–409. [Google Scholar] [CrossRef] [PubMed]
- Lasch, P.; Fleige, C.; Stämmler, M.; Layer, F.; Nübel, U.; Witte, W.; Werner, G. Insufficient Discriminatory Power of MALDI-TOF Mass Spectrometry for Typing of Enterococcus Faecium and Staphylococcus Aureus Isolates. J. Microbiol. Methods 2014, 100, 58–69. [Google Scholar] [CrossRef]
- Takahashi, N.; Nagai, S.; Fujita, A.; Ido, Y.; Kato, K.; Saito, A.; Moriya, Y.; Tomimatsu, Y.; Kaneta, N.; Tsujimoto, Y.; et al. Discrimination of Psychrotolerant Bacillus Cereus Group Based on MALDI-TOF MS Analysis of Ribosomal Subunit Proteins. Food Microbiol. 2020, 91, 103542. [Google Scholar] [CrossRef]
- Novais, Â.; Sousa, C.; de Dios Caballero, J.; Fernandez-Olmos, A.; Lopes, J.; Ramos, H.; Coque, T.M.; Cantón, R.; Peixe, L. MALDI-TOF Mass Spectrometry as a Tool for the Discrimination of High-Risk Escherichia Coli Clones from Phylogenetic Groups B2 (ST131) and D (ST69, ST405, ST393). Eur. J. Clin. Microbiol. Infect. Dis. 2014, 33, 1391–1399. [Google Scholar] [CrossRef] [PubMed]
- Nagy, E.; Urbán, E.; Becker, S.; Kostrzewa, M.; Vörös, A.; Hunyadkürti, J.; Nagy, I. MALDI-TOF MS Fingerprinting Facilitates Rapid Discrimination of Phylotypes I, II and III of Propionibacterium Acnes. Anaerobe 2013, 20, 20–26. [Google Scholar] [CrossRef] [PubMed]
- Sakarikou, C.; Ciotti, M.; Dolfa, C.; Angeletti, S.; Favalli, C. Rapid Detection of Carbapenemase-Producing Klebsiella Pneumoniae Strains Derived from Blood Cultures by Matrix-Assisted Laser Desorption Ionization-Time of Flight Mass Spectrometry (MALDI-TOF MS). BMC Microbiol. 2017, 17, 9523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, J.; Liu, J.; Li, Y.; Yu, J.; Zhu, W.; Liu, Y.; Shen, L. Rapid Detection of Carbapenemase Activity of Enterobacteriaceae Isolated from Positive Blood Cultures by MALDI-TOF MS. Ann. Clin. Microbiol. Antimicrob. 2018, 17, 22. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Han, C.; Sui, W.; Wang, M.; Lu, X. MALDI-TOF MS Applied to Indirect Carbapenemase Detection: A Validated Procedure to Clearly Distinguish between Carbapenemase-Positive and Carbapenemase-Negative Bacterial Strains. Anal. Bioanal. Chem. 2013, 405, 5259–5266. [Google Scholar] [CrossRef] [PubMed]
- Hrabák, J.; Chudác ková, E.; Walková, R. Matrix-Assisted Laser Desorption Ionization-Time of Flight (MALDITOF) Mass Spectrometry for Detection of Antibiotic Resistance Mechanisms: From Research to Routine Diagnosis. Clin. Microbiol. Rev. 2013, 26, 103–114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sparbier, K.; Schubert, S.; Weller, U.; Boogen, C.; Kostrzewa, M. Matrix-Assisted Laser Desorption Ionization-Time of Flight Mass Spectrometry-Based Functional Assay for Rapid Detection of Resistance against β-Lactam Antibiotics. J. Clin. Microbiol. 2012, 50, 927–937. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mirande, C.; Canard, I.; Buffet Croix Blanche, S.; Charrier, J.P.; van Belkum, A.; Welker, M.; Chatellier, S. Rapid Detection of Carbapenemase Activity: Benefits and Weaknesses of MALDI-TOF MS. Eur. J. Clin. Microbiol. Infect. Dis. 2015, 34, 2225–2234. [Google Scholar] [CrossRef]
- Rotova, V.; Papagiannitsis, C.C.; Skalova, A.; Chudejova, K.; Hrabak, J. Comparison of Imipenem and Meropenem Antibiotics for the MALDI-TOF MS Detection of Carbapenemase Activity. J. Microbiol. Methods 2017, 137, 30–33. [Google Scholar] [CrossRef] [PubMed]
- Calderaro, A.; Buttrini, M.; Piergianni, M.; Montecchini, S.; Martinelli, M.; Covan, S.; Piccolo, G.; Medici, M.C.; Arcangeletti, M.C.; Chezzi, C.; et al. Evaluation of a Modified Meropenem Hydrolysis Assay on a Large Cohort of KPC and VIM Carbapenemase-Producing Enterobacteriaceae. PLoS ONE 2017, 12, e0174908. [Google Scholar] [CrossRef] [PubMed]
- Miltgen, G.; Plésiat, P.; Mille, A.; Chatelain, P.; Fournier, D. Detection of Carbapenemase Activity in Pseudomonas Aeruginosa by Matrix-Assisted Laser Desorption Ionization-Time of Flight Mass Spectrometry (MALDI-TOF MS). J. Microbiol. Methods 2018, 145, 66–68. [Google Scholar] [CrossRef] [PubMed]
- Hrabák, J.; Niemczyková, J.; Chudáčková, E.; Fridrichová, M.; Študentová, V.; Červená, D.; Urbášková, P.; Žemličková, H. KPC-2-Producing Klebsiella Pneumoniae Isolated from a Czech Patient Previously Hospitalized in Greece and in Vivo Selection of Colistin Resistance. Folia Microbiol. 2011, 56, 361–365. [Google Scholar] [CrossRef] [PubMed]
- Lasserre, C.; de Martin, L.; Cuzon, G.; Bogaerts, P.; Lamar, E.; Glupczynski, Y.; Naas, T.; Tandé, D. Efficient Detection of Carbapenemase Activity in Enterobacteriaceae by Matrix-Assisted Laser Desorption Ionization-Time of Flight Mass Spectrometry in Less than 30 Minutes. J. Clin. Microbiol. 2015, 53, 2163–2171. [Google Scholar] [CrossRef] [Green Version]
- Kulkarni, M.; Zurita, A.N.; Pyka, J.S.; Murray, T.S.; Hodsdon, M.E.; Peaper, D.R. Use of Imipenem to Detect KPC, NDM, OXA, IMP, and VIM Carbapenemase Activity from Gram-Negative Rods in 75 Minutes Using Liquid Chromatography-Tandem Mass Spectrometry. J. Clin. Microbiol. 2014, 52, 2500–2505. [Google Scholar] [CrossRef] [Green Version]
- Monteferrante, C.G.; Sultan, S.; ten Kate, M.T.; Dekker, L.J.M.; Sparbier, K.; Peer, M.; Kostzrewa, M.; Luider, T.M.; Goessens, W.H.F.; Burgers, P.C. Evaluation of Different Pretreatment Protocols to Detect Accurately Clinical Carbapenemase-Producing Enterobacteriaceae by MALDI-TOF. J. Antimicrob. Chemother. 2016, 71, 2856–2867. [Google Scholar] [CrossRef]
- Rodrigues, C.; Sousa, C.; Ramos, H.; Coque, T.M.; Cantón, R.; Lopes, J.A.; Peixe, L. Elucidating Constraints for Differentiation of Major Human Klebsiella Pneumoniae Clones Using MALDI-TOF MS. Eur. J. Clin. Microbiol. Infect. Dis. 2017, 36, 2856–2867. [Google Scholar] [CrossRef] [PubMed]
- Giordano, C.; Barnini, S. Rapid Detection of Colistin-Resistant Klebsiella Pneumoniae Using MALDI-TOF MS Peak-Based Assay. J. Microbiol. Methods 2018, 155, 27–33. [Google Scholar] [CrossRef]
- Idelevich, E.A.; Sparbier, K.; Kostrzewa, M.; Becker, K. Rapid Detection of Antibiotic Resistance by MALDI-TOF Mass Spectrometry Using a Novel Direct-on-Target Microdroplet Growth Assay. Clin. Microbiol. Infect. 2018, 24, 738–743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodrigues, C.; Passet, V.; Rakotondrasoa, A.; Brisse, S. Identification of Klebsiella Pneumoniae, Klebsiella Quasipneumoniae, Klebsiella Variicola and Related Phylogroups by MALDI-TOF Mass Spectrometry. Front. Microbiol. 2018, 9. [Google Scholar] [CrossRef] [Green Version]
- Lau, A.F.; Wang, H.; Weingarten, R.A.; Drake, S.K.; Suffredini, A.F.; Garfield, M.K.; Chen, Y.; Gucek, M.; Youn, J.H.; Stock, F.; et al. A Rapid Matrix-Assisted Laser Desorption Ionization-Time of Flight Mass Spectrometry-Based Method for Single-Plasmid Tracking in an Outbreak of Carbapenem-Resistant Enterobacteriaceae. J. Clin. Microbiol. 2014, 52, 2804–2812. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gato, E.; Constanso, I.P.; Rodiño-Janeiro, B.K.; Guijarro-Sánchez, P.; Alioto, T.; Arroyo, M.J.; Méndez, G.; Mancera, L.; Gut, M.; Gut, I.; et al. Occurrence of the P019 Gene in the BlaKPC-Harboring Plasmids: Adverse Clinical Impact for Direct Tracking of KPC-Producing Klebsiella Pneumoniae by Matrix-Assisted Laser Desorption Ionization-Time of Flight Mass Spectrometry. J. Clin. Microbiol. 2021, 59, 21. [Google Scholar] [CrossRef]
- Cordovana, M.; Kostrzewa, M.; Glandorf, J.; Bienia, M.; Ambretti, S.; Pranada, A.B. A Full MALDI-Based Approach to Detect Plasmid-Encoded KPC-Producing Klebsiella Pneumoniae. Front. Microbiol. 2018, 9, 2854. [Google Scholar] [CrossRef] [PubMed]
- Centonze, A.R.; Bragantini, M.; Lucchini, E.; Mazzariol, A. Laboratory Validation of a KPC-Producing Strain Identification Method Based on the Detection of a Specific 11,109 Da Peak via Maldi-Tof-Vitek MS in an Endemic Area. New Microbiol. 2019, 42, 114–117. [Google Scholar] [PubMed]
- Neonakis, I.K.; Spandidos, D.A. Detection of Carbapenemase Producers by Matrix-Assisted Laser Desorption-Ionization Time-of-Flight Mass Spectrometry (MALDI-TOF MS). Eur. J. Clin. Microbiol. Infect. Dis. 2019, 38, 1795–1801. [Google Scholar] [CrossRef] [PubMed]
- Migliorini, L.B.; de Sales, R.O.; Koga, P.C.M.; Doi, A.M.; Poehlein, A.; Toniolo, A.R.; Menezes, F.G.; Martino, M.D.V.; Gales, A.C.; Brüggemann, H.; et al. Prevalence of Blakpc-2, Blakpc-3 and Blakpc-30—Carrying Plasmids in Klebsiella Pneumoniae Isolated in a Brazilian Hospital. Pathogens 2021, 10, 332. [Google Scholar] [CrossRef] [PubMed]
- Partridge, S.R. Tn4401 Carrying BlaKPC Is Inserted within Another Insertion in PKpQIL and Related Plasmids. J. Clin. Microbiol. 2014, 52, 4448–4449. [Google Scholar] [CrossRef] [Green Version]
- Yao, H.; Cheng, J.; Li, A.; Yu, R.; Zhao, W.; Qin, S.; Du, X.D. Molecular Characterization of an IncFIIk Plasmid Co-Harboring BlaIMP–26 and Tet(A) Variant in a Clinical Klebsiella Pneumoniae Isolate. Front. Microbiol. 2020, 11, 1610. [Google Scholar] [CrossRef]
- Rada, A.M.; de la Cadena, E.; Agudelo, C.; Capataz, C.; Orozco, N.; Pallares, C.; Dinh, A.Q.; Panesso, D.; Ríos, R.; Diaz, L.; et al. Dynamics of BlaKPC-2 Dissemination from Non-CG258 Klebsiella Pneumoniae to Other Enterobacterales via IncN Plasmids in an Area of High Endemicity. Antimicrob. Agents Chemother. 2020, 64, 174320. [Google Scholar] [CrossRef] [PubMed]
- Gootz, T.D.; Lescoe, M.K.; Dib-Hajj, F.; Dougherty, B.A.; He, W.; Della-Latta, P.; Huard, R.C. Genetic Organization of Transposase Regions Surrounding BlaKPC Carbapenemase Genes on Plasmids from Klebsiella Strains Isolated in a New York City Hospital. Antimicrob. Agents Chemother. 2009, 53, 1998–2004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Espinal, P.; Nucleo, E.; Caltagirone, M.; Mattioni Marchetti, V.; Fernandes, M.R.; Biscaro, V.; Rigoli, R.; Carattoli, A.; Migliavacca, R.; Villa, L. Genomics of Klebsiella Pneumoniae ST16 Producing NDM-1, CTX-M-15, and OXA-232. Clin. Microbiol. Infect. 2019, 25, 385.e1–. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, L.; Chavda, K.D.; al Laham, N.; Melano, R.G.; Jacobs, M.R.; Bonomo, R.A.; Kreiswirth, B.N. Complete Nucleotide Sequence of a BlaKPC-Harboring IncI2 Plasmid and Its Dissemination in New Jersey and New York Hospitals. Antimicrob. Agents Chemother. 2013, 57, 5019–5025. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Lu, J.; Zhang, R.; Cai, J. Antibiotic Resistance and Virulence Characteristics Analysis of a Carbapenem-Resistant Hypervirulent Klebsiella Pneumoniae. Natl. Med J. China 2021, 101, 3143. [Google Scholar] [CrossRef]
- Ho, P.L.; Cheung, Y.Y.; Lo, W.U.; Li, Z.; Chow, K.H.; Lin, C.H.; Chan, J.F.W.; Cheng, V.C.C. Molecular Characterization of an Atypical IncX3 Plasmid PKPC-NY79 Carrying Bla KPC-2 in a Klebsiella Pneumoniae. Curr. Microbiol. 2013, 67, 493–498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Q.; Zhou, J.; Wu, S.; Yang, Y.; Yu, D.; Wang, X.; Wu, M. Characterization of the IncX3 Plasmid Producing BlaNDM–7 From Klebsiella Pneumoniae ST34. Front. Microbiol. 2020, 11, 1885. [Google Scholar] [CrossRef] [PubMed]
- Figueroa-Espinosa, R.; Costa, A.; Cejas, D.; Barrios, R.; Vay, C.; Radice, M.; Gutkind, G.; di Conza, J. MALDI-TOF MS Based Procedure to Detect KPC-2 Directly from Positive Blood Culture Bottles and Colonies. J. Microbiol. Methods 2019, 159, 120–127. [Google Scholar] [CrossRef] [PubMed]
- Yoon, E.J.; Lee, E.H.; Hwang, D.H.; Lee, H.; Baek, J.H.; Jeong, S.H. Direct Detection of Intact Klebsiella Pneumoniae Carbapenemases Produced by Enterobacterales Using MALDI-TOF MS. J. Antimicrob. Chemother. 2020, 75, 1174–1181. [Google Scholar] [CrossRef] [PubMed]
- Hleba, L.; Hlebová, M.; Kováčik, A.; Šmehýl, P.; Hricáková, N.; Petrová, J.; Shariati, M.A.; Čuboň, J. Escherichia Coli as a Carrier of Tetracyclines and Penicillins Resistance in Wild Pheasant (Phasianus Colchicus). J. Environ. Sci. Health-Part A Toxic/Hazard. Subst. Environ. Eng. 2020, 55, 1201–1209. [Google Scholar] [CrossRef]
- EUCAST. EUCAST Disk Diffusion Method for Antimicrobial Susceptibility Testing; ESCMID: Basel, Switzerland, 2017; Volume 6. [Google Scholar]
- EUCAST. Breakpoint Tables for Interpretation of MICs and Zone Diameters. Version 11.0. 2021. Available online: http://www.eucast.org (accessed on 1 January 2021).
- Pillai, D.R.; Melano, R.; Rawte, P.; Lo, S.; Tijet, N.; Fuksa, M.; Roda, N.; Farrell, D.J.; Krajden, S. Klebsiella Pneumoniae Carbapenemase, Canada. Emerg. Infect. Dis. 2009, 15, 827–829. [Google Scholar] [CrossRef] [PubMed]
- Niedermeyer, T.H.J.; Strohalm, M. MMass as a Software Tool for the Annotation of Cyclic Peptide Tandem Mass Spectra. PLoS ONE 2012, 7, e44913. [Google Scholar] [CrossRef] [PubMed] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| C-ATB | Native Forms of ATB (m/z) | Hydrolyzed Forms of ATB (m/z) | ||||||
|---|---|---|---|---|---|---|---|---|
| [M + H]+ | [M + Na]+ | [M + 2Na]+ | [M + 3Na]+ | [M + H]+ | [M + Na]+ | [M + 2Na]+ | [M + 3Na]+ | |
| Meropenem | 382 1,2,3 | 405 1,2,3 | 427 1,2,3 | 401 2,3 | 423 2,3 | 445 2,3 | 467 2 | |
| Ertapenem | 476 1,4 | 498 1,4,5 | 520 1,4,5 | 542 1,4 | 494 1,4 | 516 1,4 | 538 1,4 | 560 4 |
| Faropenem | 286 4 | 308 4 | 330 4 | 352 4 | 304 4 | 326 4 | 348 4 | 370 4 |
| Native forms of ATB | Decarboxylated forms of ATB | |||||||
| [M + H]+ | [M + Na]+ | [M + 2Na]+ | [M + 3Na]+ | [M + H2O − CO2 + H]+ | [M + H2O − CO2 + Na]+ | |||
| Imipenem | 3001 | 3225 | 2745 | 2965 | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hleba, L.; Hlebová, M.; Kováčik, A.; Čuboň, J.; Medo, J. Carbapenemase Producing Klebsiella pneumoniae (KPC): What Is the Best MALDI-TOF MS Detection Method. Antibiotics 2021, 10, 1549. https://doi.org/10.3390/antibiotics10121549
Hleba L, Hlebová M, Kováčik A, Čuboň J, Medo J. Carbapenemase Producing Klebsiella pneumoniae (KPC): What Is the Best MALDI-TOF MS Detection Method. Antibiotics. 2021; 10(12):1549. https://doi.org/10.3390/antibiotics10121549
Chicago/Turabian StyleHleba, Lukáš, Miroslava Hlebová, Anton Kováčik, Juraj Čuboň, and Juraj Medo. 2021. "Carbapenemase Producing Klebsiella pneumoniae (KPC): What Is the Best MALDI-TOF MS Detection Method" Antibiotics 10, no. 12: 1549. https://doi.org/10.3390/antibiotics10121549
APA StyleHleba, L., Hlebová, M., Kováčik, A., Čuboň, J., & Medo, J. (2021). Carbapenemase Producing Klebsiella pneumoniae (KPC): What Is the Best MALDI-TOF MS Detection Method. Antibiotics, 10(12), 1549. https://doi.org/10.3390/antibiotics10121549