
Staphylococcus aureus Interferes with Streptococci Spatial Distribution and with Protein Expression of Species within a Polymicrobial Oral Biofilm


 , , , , , and
, , , , , and
Abstract
:1. Introduction
2. Experimental Section
2.1. Bacterial Strains and Growth Conditions
2.2. Construction of the Staphylococcus Aureus Mutant Strain
2.3. Multispecies Biofilm Formation and Harvesting
2.4. Image Analysis with Confocal Laser Scanning Microscopy (CLSM)
2.5. Culture Analyses
2.6. Bacterial and Biofilm Protein Extraction
2.7. Liquid Chromatography-Mass Spectrometry Analysis
2.8. Protein Identification and Label-Free Quantification
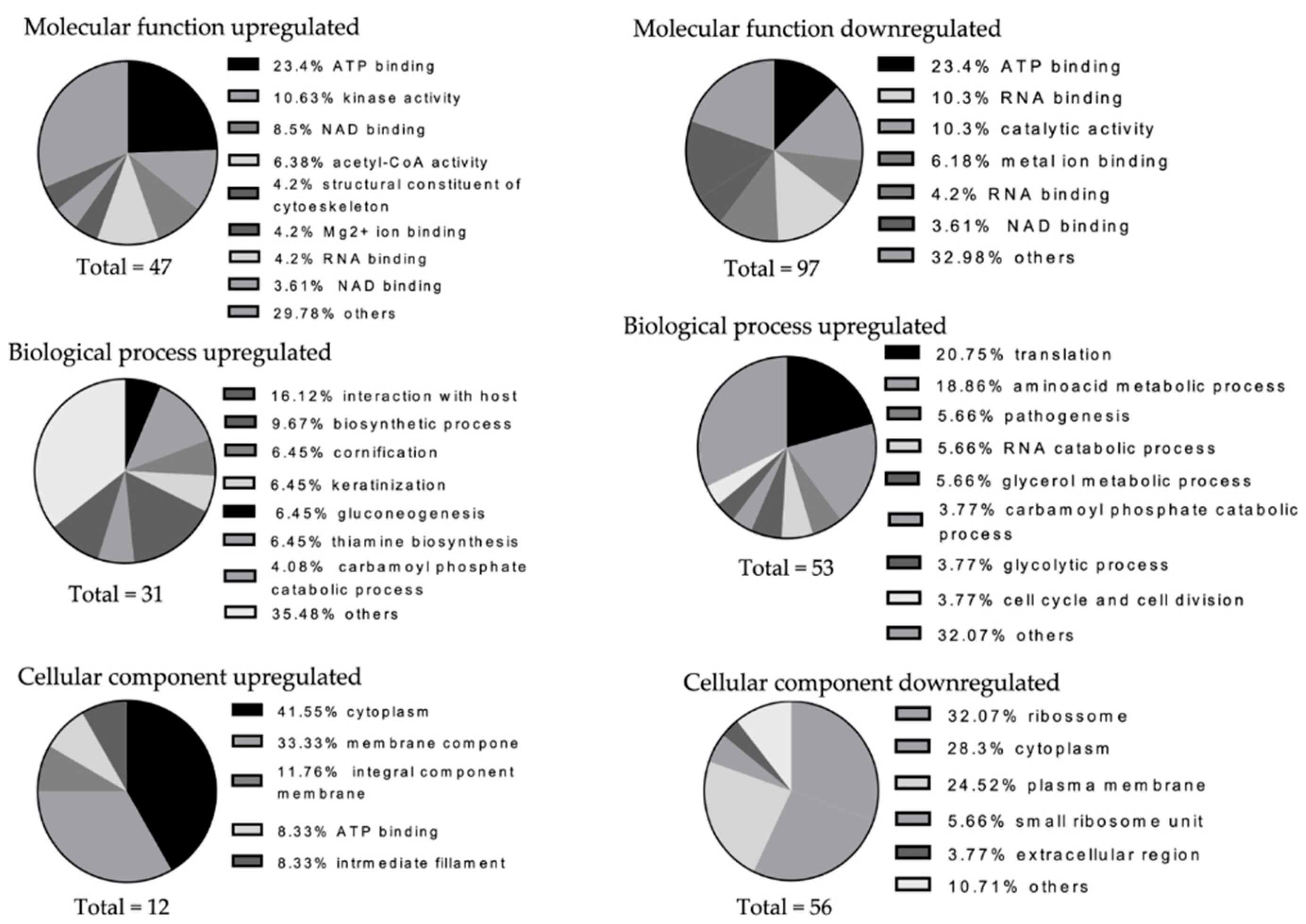
2.9. Functional and Ontology Analysis
2.10. Statistical Analysis
3. Results
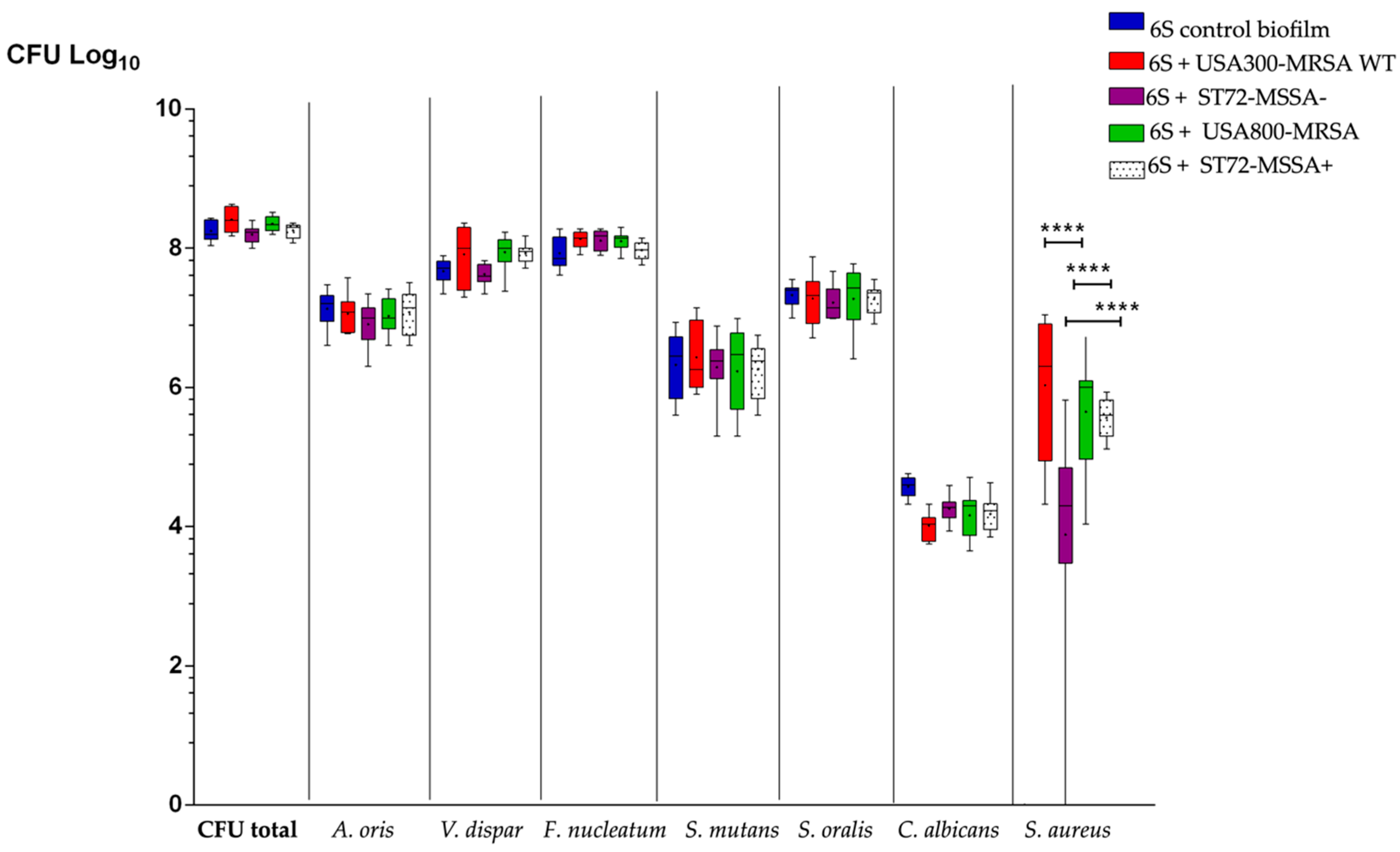
3.1. S. Aureus Strains Differently Affect the Number of Supragingival-Associated Species Grown on Hydroxyapatite Disks
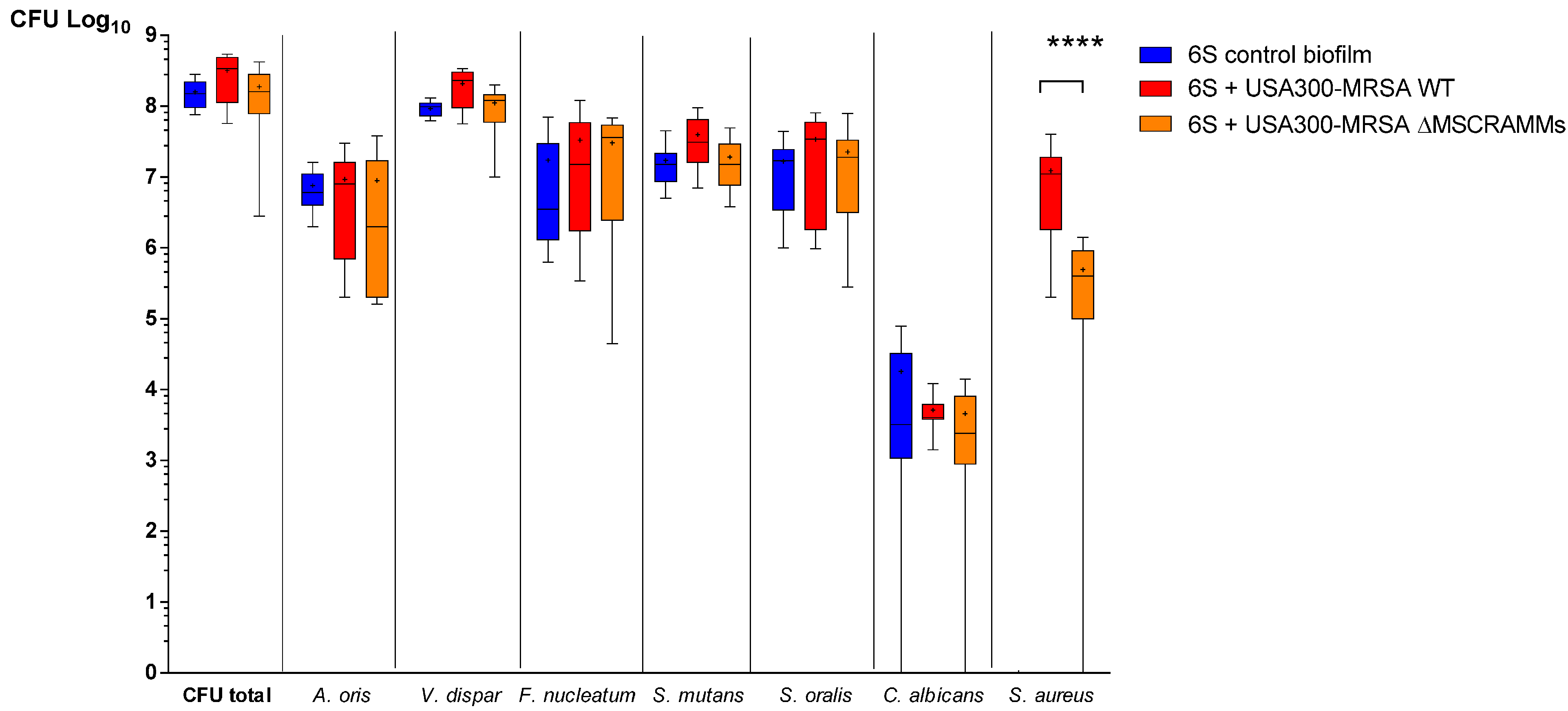
3.2. MSCRAMM Influence the Growth of S. aureus in Multispecies Oral Biofilms
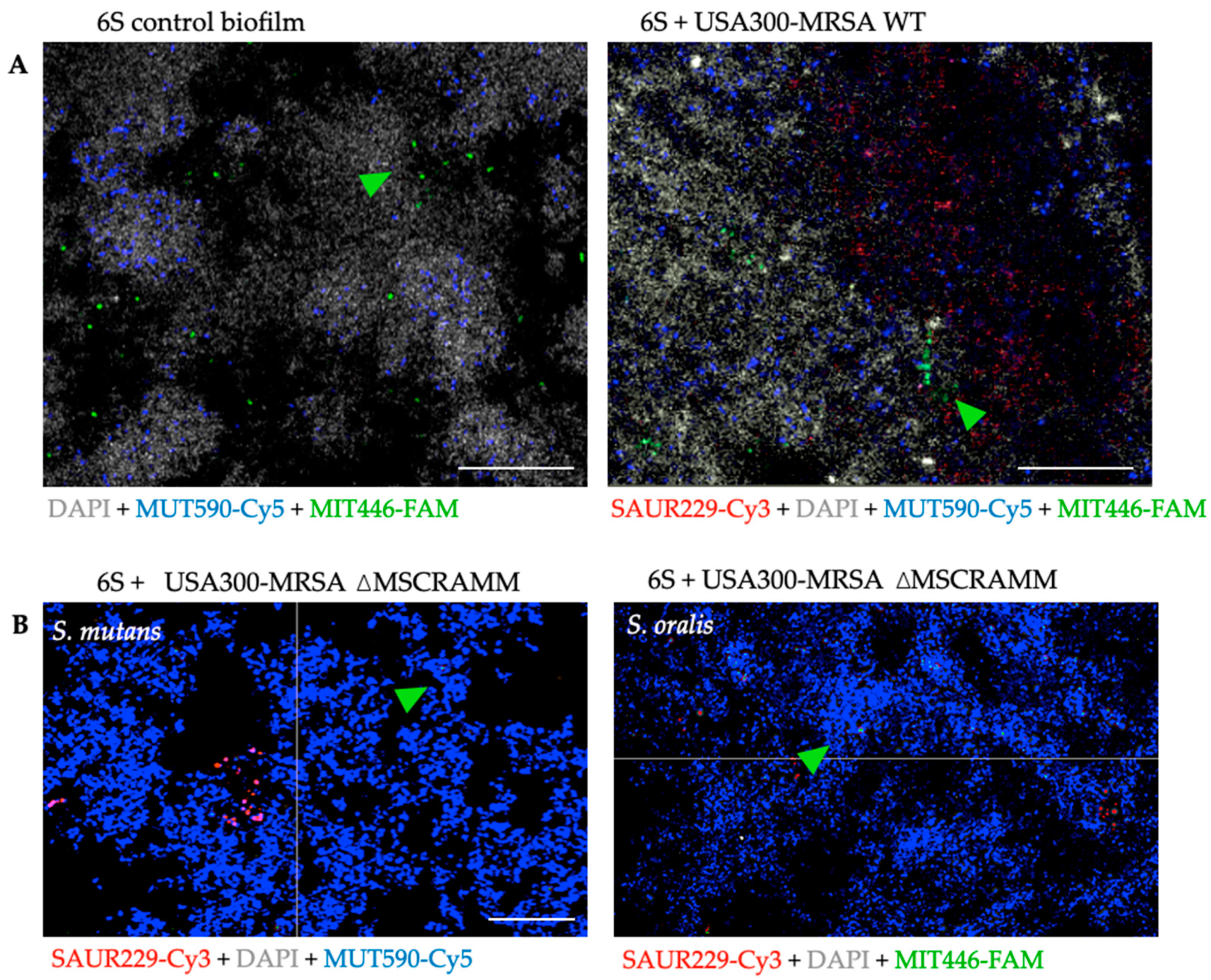
3.3. The Distribution of S. mutans and S. oralis is Modified When Staphylococcus Aureus MSCRAMM Genes are Deleted
3.4. Staphylococcus Aureus Causes Shifts in the Numbers of Other Species’ Detectable Proteins in the Six-Species Biofilm
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| FISH | Fluorescent in situ hybridization |
| CLSM | Confocal laser scanning microscopy |
| PIA | Extracellular polysaccharide intercellular adhesion |
| CWA | Cell-wall-anchored proteins |
| MSCRAMM | Microbial surface that recognizes adhesive matrix molecules |
| clf | Clumping factors |
| sdr | Serine aspartate repeats proteins |
| fnBP | Fibronectin-binding proteins |
| MRSA | Methicillin-resistant S. aureus |
| MSSA | Methicillin-sensitive S. aureus |
| icaADBC | Operon-encoded polysaccharide intercellular adhesin |
| eDNA | Extracellular DNA |
| MLST | Multilocus sequence type |
| SCC | Staphylococcal cassette chromosome |
| PVL | Panton-Valentine leukocidin (PVL)-encoding genes (lukF-PV and lukS-PV) |
| CBA | Columbia blood agar |
| TSB | Tryptic soy broth |
| SSmec | Staphylococcal cassette chromosome mec |
| mFUM | Fluid universal medium |
| CFU | culture forming units |
| HA | Hydroxyapatite disk |
| LC-MS | Liquid Chromatography-Mass Spectrometry |
| HIFU | High Intensity Focused Ultrasound |
| DDA | Data-dependent analysis |
| LIMS | Laboratory information management system |
| FDR | False discovery rates |
| GO | Gene ontology |
| ALS | Agglutinin-like sequence |
| DLL | Dock, lock and latch mechanism |
| CH | Collagen hug |
| Ssp | Streptococcal surface proteins |
| CBPs | Collagen-binding proteins |
| PBP2a | penicillin-binding protein 2a |
References
- Tong, S.Y.C.; Davis, J.S.; Eichenberger, E.; Holland, T.L.; Fowler, V.G. Staphylococcus aureus infections: Epidemiology, pathophysiology, clinical manifestations, and management. Clin. Microbiol. Rev. 2015, 28, 603–661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chan, L.C.; Basuino, L.; Dip, E.C.; Chambers, H.F. Comparative Efficacies of Tedizolid Phosphate, Vancomycin, and Daptomycin in a Rabbit Model of Methicillin-Resistant Staphylococcus aureus Endocarditis. Antimicrob. Agents Chemother. 2015, 59, 3252–3256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arciola, C.R.; Campoccia, D.; Montanaro, L. Implant infections: Adhesion, biofilm formation and immune evasion. Nat. Rev. Microbiol. 2018, 16, 397–409. [Google Scholar] [CrossRef] [PubMed]
- Sanz, M.; Beighton, D.; Curtis, M.A.; Cury, J.A.; Dige, I.; Dommisch, H.; Ellwood, R.; Giacaman, R.A.; Herrera, D.; Herzberg, M.C.; et al. Role of microbial biofilms in the maintenance of oral health and in the development of dental caries and periodontal diseases. Consensus report of group 1 of the Joint EFP/ORCA workshop on the boundaries between caries and periodontal disease. J. Clin. Periodontol. 2017, 44, 5–11. [Google Scholar] [CrossRef]
- Dewhirst, F.E.; Chen, T.; Izard, J.; Paster, B.J.; Tanner, A.C.R.; Yu, W.H.; Lakshmanan, A.; Wade, W.G. The human oral microbiome. J. Bacteriol. 2010, 192, 5002–5017. [Google Scholar] [CrossRef] [Green Version]
- Sultan, A.S.; Kong, E.F.; Rizk, A.M.; Jabra-Rizk, M.A. The oral microbiome: A Lesson in coexistence. PLoS Pathog. 2018, 14, e1006719. [Google Scholar] [CrossRef] [Green Version]
- Thurnheer, T.; Belibasakis, G.N. Integration of non-oral bacteria into in vitro oral biofilms. Virulence 2015, 6, 258–264. [Google Scholar] [CrossRef] [Green Version]
- Paharik, A.E.; Horswill, A.R. The Staphylococcal Biofilm: Adhesins, Regulation, and Host Response. Microbiol. Spectr. 2016, 4, 1–22. [Google Scholar] [CrossRef] [Green Version]
- Lamont, R.J.; Koo, H.; Hajishengallis, G. The oral microbiota: Dynamic communities and host interactions. Nat. Rev. Microbiol. 2018, 16, 745–759. [Google Scholar] [CrossRef]
- Thurnheer, T.; Gmür, R.; Guggenheim, B. Multiplex FISH analysis of a six-species bacterial biofilm. J. Microbiol. Methods. 2004, 56, 37–47. [Google Scholar] [CrossRef]
- Foulston, L.; Elsholz, A.K.W.; DeFrancesco, A.S.; Losick, R. The extracellular matrix of Staphylococcus aureus biofilms comprises cytoplasmic proteins that associate with the cell surface in response to decreasing pH. mBio 2014, 5, e01667-14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foster, T.J.; Geoghegan, J.A.; Ganesh, V.K.; Höök, M. Adhesion, invasion and evasion: The many functions of the surface proteins of Staphylococcus aureus. Nat. Rev. Microbiol. 2014, 12, 49–62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Archer, N.K.; Mazaitis, M.J.; Costerton, J.W.; Leid, J.G.; Powers, M.E.; Shirtliff, M.E. Staphylococcus aureus biofilms: Properties, regulation and roles in human disease. Virulence 2011, 2, 445–459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cardile, A.P.; Sanchez Jr, C.J.; Samberg, M.E.; Romano, D.R.; Hardy, S.K.; Wenke, J.C.; Murray, C.K.; Akers, K.S. Human plasma enhances the expression of Staphylococcal microbial surface components recognizing adhesive matrix molecules promoting biofilm formation and increases antimicrobial tolerance In Vitro. BMC Res. Notes 2014, 7, 457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raafat, D.; Otto, M.; Reppschläger, K.; Iqbal, J.; Holtfreter, S. Fighting Staphylococcus aureus Biofilms with Monoclonal Antibodies. Trends Microbiol. 2019, 27, 303–322. [Google Scholar] [CrossRef]
- McDevitt, D.; Nanavaty, T.; House-Pompeo, K.; Bell, E.; Turner, N.; McIntire, L.; Foster, T.; Höök, M. Characterization of the interaction between the Staphylococcus aureus clumping factor (ClfA) and fibrinogen. Eur. J. Biochem. 1997, 247, 416–424. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.Z.; Shih, M.H.; Tsai, P.J. ClfA (221–550), a fibrinogen-binding segment of Staphylococcus aureus clumping factor A, disrupts fibrinogen function. Thromb. Haemost. 2005, 94, 286–294. [Google Scholar] [CrossRef]
- Flick, M.J.; Du, X.; Prasad, J.M.; Raghu, H.; Palumbo, J.S.; Smeds, E.; Höök, M.; Degen, J.L. Genetic elimination of the binding motif on fibrinogen for the S. aureus virulence factor ClfA improves host survival in septicemia. Blood 2013, 121, 1783–1794. [Google Scholar] [CrossRef] [Green Version]
- Cheng, A.G.; Kim, H.K.; Burts, M.L.; Krausz, T.; Schneewind, O.; Missiakas, D.M. Genetic requirements for Staphylococcus aureus abscess formation and persistence in host tissues. FASEB J. 2009, 23, 3393–3404. [Google Scholar] [CrossRef] [Green Version]
- Moreillon, P.; Entenza, J.M.; Francioli, P.; McDevitt, D.; Foster, T.J.; Francois, P.; Vaudaux, P. Role of Staphylococcus aureus coagulase and clumping factor in pathogenesis of experimental endocarditis. Infect. Immun. 1995, 63, 4738–4743. [Google Scholar] [CrossRef] [Green Version]
- Fitzpatrick, F.; Humphreys, H.; O’Gara, J.P. Evidence for icaADBC-independent biofilm development mechanism in methicillin-resistant Staphylococcus aureus clinical isolates. J. Clin. Microbiol. 2005, 43, 1973–1976. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strommenger, B.; Braulke, C.; Pasemann, B.; Schmidt, C.; Witte, W. Multiplex PCR for rapid detection of Staphylococcus aureus isolates suspected to represent community-acquired strains. J. Clin. Microbiol. 2008, 46, 582–587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harmsen, D.; Claus, H.; Witte, W.; Rothganger, J.; Turnwald, D.; Vogel, U. Typing of methicillin-resistant Staphylococcus aureus in a university hospital setting by using novel software for spa repeat determination and database management. J. Clin. Microbiol. 2003, 41, 5442–5448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holtfreter, S.; Radcliff, F.J.; Grumann, D.; Read, H.; Johnson, S.; Monecke, S.; Ritchie, S.; Clow, F.; Goerke, C.; Bröker, B.M.; et al. Characterization of a Mouse-Adapted Staphylococcus aureus Strain. PLoS ONE 2013, 8, 71142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holtfreter, S.; Grumann, D.; Schmudde, M.; Nguyen, H.T.; Eichler, P.; Strommenger, B.; Kopron, K.; Kolata, J.; Giedrys-Kalemba, S.; Steinmetz, I.; et al. Clonal distribution of superantigen genes in clinical Staphylococcus aureus isolates. J. Clin. Microbiol. 2007, 45, 2669–2680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Enright, M.C.; Day, N.P.; Davies, C.E.; Peacock, S.J.; Spratt, B.G. Multilocus sequence typing for characterization of methicillin-resistant and methicillin-susceptible clones of Staphylococcus aureus. J. Clin. Microbiol. 2000, 38, 1008–1015. [Google Scholar] [CrossRef] [Green Version]
- Holtfreter, S.; Grumann, D.; Balau, V.; Barwich, A.; Kolata, J.; Goehler, A.; Weiss, S.; Holtfreter, B.; Bauerfeind, S.S.; Döring, P.; et al. Molecular Epidemiology of Staphylococcus aureus in the General Population in Northeast Germany: Results of the Study of Health in Pomerania (SHIP-TREND-0). J. Clin. Microbiol. 2016, 54, 2774–2785. [Google Scholar] [CrossRef] [Green Version]
- Diep, B.A.; Gill, S.R.; Chang, R.F.; Phan, T.H.; Chen, J.H.; Davidson, M.G.; Lin, F.; Lin, J.; Carleton, H.A.; Mongodin, E.F.; et al. Complete genome sequence of USA300, an epidemic clone of community-acquired meticillin-resistant Staphylococcus aureus. Lancet 2006, 367, 731–739. [Google Scholar] [CrossRef]
- Monecke, S.; Slickers, P.; Ehricht, R. Assignment of Staphylococcus aureus isolates to clonal complexes based on microarray analysis and pattern recognition. FEMS Immunol. Med. Microbiol. 2008, 53, 237–251. [Google Scholar] [CrossRef] [Green Version]
- Treangen, T.J.; Maybank, R.A.; Enke, S.; Friss, M.B.; Diviak, L.F.; Karaolis, D.K.; Koren, S.; Ondov, B.; Phillippy, A.M.; Bergman, N.H.; et al. Complete Genome Sequence of the Quality Control Strain Staphylococcus aureus subsp. aureus ATCC 25923. Genome Announc. 2014, 2, e01110–e01114. [Google Scholar] [CrossRef] [Green Version]
- Bae, T.; Schneewind, O. Allelic replacement in Staphylococcus aureus with inducible counter-selection. Plasmid 2006, 55, 58–63. [Google Scholar] [CrossRef] [PubMed]
- Diep, B.A.; Chan, L.; Tattevin, P.; Kajikawa, O.; Martin, T.R.; Basuino, L.; Mai, T.T.; Marbach, H.; Braughton, K.R.; Whitney, A.R.; et al. Polymorphonuclear leukocytes mediate Staphylococcus aureus Panton-Valentine leukocidin-induced lung inflammation and injury. Proc. Natl. Acad. Sci. USA 2010, 107, 5587–5592. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thurnheer, T.; van der Ploeg, J.R.; Giertsen, E.; Guggenheim, B. Effects of Streptococcus mutans gtfC deficiency on mixed oral biofilms in vitro. Caries Res. 2006, 40, 163–171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guggenheim, B.; Giertsen, E.; Schüpbach, P.; Shapiro, S. Validation of an in vitro biofilm model of supragingival plaque. J. Dent. Res. 2001, 80, 363–370. [Google Scholar] [CrossRef] [PubMed]
- Thurnheer, T.; Belibasakis, G.N. Incorporation of staphylococci into titanium-grown biofilms: An in vitro "submucosal" biofilm model for peri-implantitis. Clin. Oral Implants Res. 2016, 27, 890–895. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quevedo, B.; Giertsen, E.; Zijnge, V.; Luthi-Schaller, H.; Guggenheim, B.; Thurnheer, T.; Gmur, R. Phylogenetic group- and species-specific oligonucleotide probes for single-cell detection of lactic acid bacteria in oral biofilms. BMC Microbiol. 2011, 11, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bao, K.; Bostanci, N.; Selevsek, N.; Thurnheer, T.; Belibasakis, G.N. Quantitative proteomics reveal distinct protein regulations caused by Aggregatibacter actinomycetemcomitans within subgingival biofilms. PLoS ONE 2015, 10, e0119222. [Google Scholar] [CrossRef] [PubMed]
- Turker, C.; Akal, F.; Joho, D.; Panse, C.; Barkow-Oesterreicher, S.; Rehrauer, H.; Schlapbach, H. B-Fabric: The Swiss Army Knife for Life Sciences. EDBT 2010, 426, 22–26. [Google Scholar] [CrossRef] [Green Version]
- Jürgen, C.; Mann, M. MaxQuant Enables High Peptide Identification Rates, Individualized P.p.b.-Range Mass Accuracies and Proteome-Wide Protein Quantification. Nat. Biotechnol. 2008, 26, 1367–1372. [Google Scholar] [CrossRef]
- Wolski, W.; Grossmann, J.; Panse, C. SRM Service-R-Package to Report Quantitative Mass Spectrometry Data. 2018. Available online: http://github.com/protViz/SRMService (accessed on 12 December 2020).
- Perlroth, J.; Choi, B.; Spellberg, B. Nosocomial fungal infections: Epidemiology, diagnosis, and treatment. Med. Mycol. 2007, 45, 321–346. [Google Scholar] [CrossRef]
- Adam, B.; Baillie, G.S.; Douglas, L.J. Mixed species biofilms of Candida albicans and Staphylococcus epidermidis. J. Med. Microbiol. 2002, 51, 344–349. [Google Scholar] [CrossRef] [Green Version]
- Klotz, S.A.; Chasin, B.S.; Powell, B.; Gaur, N.K.; Lipke, P.N. Polymicrobial bloodstream infections involving Candida species: Analysis of patients and review of the literature. Diagn. Microbiol. Infect. Dis. 2007, 59, 401–406. [Google Scholar] [CrossRef]
- Peters, B.M.; Jabra-Rizk, M.A.; O’May, G.A.; Costerton, J.W.; Shirtliff, M.E. Polymicrobial interactions: Impact on pathogenesis and human disease. Clin. Microbiol. Rev. 2012, 25, 193–213. [Google Scholar] [CrossRef] [Green Version]
- Nobbs, A.H.; Vickerman, M.M.; Jenkinson, H.F. Heterologous expression of Candida albicans cell wall-associated adhesins in Saccharomyces cerevisiae reveals differential specificities in adherence and biofilm formation and in binding oral Streptococcus gordonii. Eukaryot Cell 2010, 9, 1622–1634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Groot, P.W.; Bader, O.; de Boer, A.D.; Weig, M.; Chauhan, N. Adhesins in human fungal pathogens: Glue with plenty of stick. Eukaryot Cell 2013, 12, 470–481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beaussart, A.; Herman, P.; El-Kirat-Chatel, S.; Lipke, P.N.; Kucharikova, S.; Van Dijck, P.; Dufrene, Y.F. Single-cell force spectros-copy of the medically important Staphylococcus epidermidis-Candida albicans interaction. Nanoscale 2013, 5, 10894–10900. [Google Scholar] [CrossRef] [Green Version]
- Bowman, S.M.; Free, S.J. The structure and synthesis of the fungal cell wall. Bioessays 2006, 28, 799–808. [Google Scholar] [CrossRef] [PubMed]
- Patti, J.M.; Allen, B.L.; McGavin, M.J.; Höök, M. MSCRAMM-mediated adherence of microorganisms to host tissues. Annu. Rev. Microbiol. 1994, 48, 585–617. [Google Scholar] [CrossRef]
- Foster, T.J. The MSCRAMM Family of Cell-Wall-Anchored Surface Proteins of Gram-Positive Cocci. Trends in Microbiology. Trends Microbiol. 2019, 27, 927–941. [Google Scholar] [CrossRef]
- Goss, C.H.; Muhlebach, M.S. Review: Staphylococcus aureus and MRSA in cystic fibrosis. J. Cyst. Fibros. 2011, 10, 298–306. [Google Scholar] [CrossRef] [Green Version]
- Mann, E.E.; Rice, K.C.; Boles, B.R.; Endres, J.L.; Ranjit, D.; Chandramohan, L.; Tsang, L.H.; Smeltzer, M.S.; Horswill, A.R.; Bayles, K.W. Modulation of eDNA release and degradation affects Staphylococcus aureus biofilm maturation. PLoS ONE 2009, 4, e5822. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boles, B.R.; Horswill, A.R. Staphylococcal biofilm disassembly. Trends Microbiol. 2011, 19, 449–455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moormeier, D.E.; Bose, J.L.; Horswill, A.R.; Bayles, K.W. Temporal and stochastic control of Staphylococcus aureus biofilm development. mBio 2014, 5, e01341-14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hobley, L.; Harkins, C.; MacPhee, C.E.; Stanley-Wall, N.R. Giving structure to the biofilm matrix: An overview of individual strategies and emerging common themes. FEMS Microbiol. Rev. 2015, 39, 649–669. [Google Scholar] [CrossRef] [Green Version]
- Nyvad, B.; Kilian, M. Microbiology of the early colonization of human enamel and root surfaces in vivo. Scand. J. Dent. Res. 1987, 95, 369–380. [Google Scholar] [CrossRef]
- Gibbons, R.J. Bacterial adhesion to oral tissues: A model for infectious diseases. J. Dent. Res. 1989, 68, 750–760. [Google Scholar] [CrossRef]
- Salvi, G.E.; Furst, M.M.; Lang, N.P.; Persson, G.R. One-year bacterial colonization patterns of Staphylococcus aureus and other bacteria at implants and adjacent teeth. Clin. Oral Implant. Res. 2008, 19, 242–248. [Google Scholar] [CrossRef]
- Mazmanian, S.K.; Liu, G.; Ton-That, H.; Schneewind, O. Staphylococcus aureus sortase, an enzyme that anchors surface proteins to the cell wall. Science 1999, 285, 760–763. [Google Scholar] [CrossRef]
- Singh, B.; Fleury, C.; Jalalvand, F.; Riesbeck, K. Human pathogens utilize host extracellular matrix proteins laminin and collagen for adhesion and invasion of the host. FEMS Microbiol. 2012, 36, 1122–1180. [Google Scholar] [CrossRef] [Green Version]
- Ajdić, D.; McShan, W.M.; Laughlin, R.E.; Savić, G.; Chang, J.; Carson, M.B.; Primeaux, C.; Tian, R.; Kenton, S.; Jia, H.; et al. Genome sequence of Streptococcus mutans UA159, acariogenic dental pathogen. Proc. Natl. Acad. Sci. USA 2002, 99, 14434–14439. [Google Scholar] [CrossRef] [Green Version]
- Avilés-Reyes, A.; Miller, J.H.; Lemos, J.A.; Abranches, J. Collagen-binding proteins of Streptococcus mutans and related streptococci. Mol. Oral Microbiol. 2016, 32, 89–106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dao, M.L.; Chavez, C.; Hirachi, Y.; Ferretti, J.J. Molecular cloning of the Streptococcus mutans gene specifying antigen A. Infect. Immun. 1989, 57, 3372–3376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, T.K.; Zhang, C.; Dao, M.L. Identification and characterization of collagen-binding activity in Strep-tococcus mutans wall-associated protein: A possible implication in dental root caries and endocarditis. Bio-chem. Biophys. Res. Commun. 2006, 343, 787–792. [Google Scholar] [CrossRef] [PubMed]
- Sato, Y.; Okamoto, K.; Kagami, A.; Yamamoto, Y.; Ohta, K.; Igarashi, T.; Kizaki, H. Application of in vitro mutagenesis to identify the gene responsible for cold agglutination phenotype of Streptococcus mutans. Microbiol. Immunol. 2004, 48, 449–456. [Google Scholar] [CrossRef] [PubMed]
- Sato, Y.; Okamoto, K.; Kagami, A.; Yamamoto, Y.; Igara-shi, T.; Kizaki, H. Streptococcus mutans strains harboring collagen-binding adhesin. J. Dent. Res. 2004, 83, 534–539. [Google Scholar] [CrossRef] [Green Version]
- Nomura, R.; Nakano, K.; Naka, S.; Nemoto, H.; Masuda, K.; Lapirattanakul, J.; Alaluusua, S.; Matsumoto, M.; Kawabata, S.; Ooshima, T. Identification and characterization of a collagen-binding protein, Cbm, in Streptococcus mutans. Mol. Oral Microbiol. 2012, 27, 308–323. [Google Scholar] [CrossRef]
- Cornejo, O.E.; Lefébure, T.; Bitar, P.D.; Lang, P.; Richards, V.P.; Eilertson, K.; Do, T.; Beighton, D.; Zeng, L.; Ahn, S.J.; et al. Evolutionary and population genomics of the cavity causing bacteria Streptococcus mutans. Mol. Biol. Evol. 2013, 30, 881–893. [Google Scholar] [CrossRef] [Green Version]
- Palmer, S.R.; Miller, J.H.; Abranches, J. Phenotypic heterogeneity of genomically-diverse isolates of Streptococcus mutans. PLoS ONE 2013, 8, e61358. [Google Scholar] [CrossRef]
- Nomura, R.; Nakano, K.; Taniguchi, N.; Lapirattanakul, J.; Hirotoshi, N.; Grönroos, L.; Alaluusua, S.; Ooshima, T. Molecular and clinical analyses of the collagen-binding adhesin of Streptococcus mutans. J. Med. Microbiol. 2009, 58, 469–475. [Google Scholar] [CrossRef] [Green Version]
- Abranches, J.; Miller, J.H.; Martinez, A.R.; Simpson-Hai-Daris, P.J.; Burne, R.A.; Lemos, J.A. The collagen-binding protein Cnm is required for Streptococcus mutans adherence to and intracellular invasion of human coronary artery endothelial cells. Infect. Immun. 2011, 79, 2277–2284. [Google Scholar] [CrossRef] [Green Version]
- Nakano, K.; Hokamura, K.; Taniguchi, N.; Wada, K.; Kudo, C.; Nomura, R.; Kojima, A.; Naka, S.; Muranaka, Y.; Thura, M.; et al. The collagen-binding protein of Streptococcus mutans is involved in haemorrhagic stroke. Nat. Commun. 2011, 2, 485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nomura, R.; Naka, S.; Nemoto, H.; Inagaki, S.; Taniguchi, K.; Ooshima, T.; Nakano, K. Potential involvement of collagen-binding proteins of Streptococcus mutans in infective endocarditis. Oral Dis. 2013, 19, 387–393. [Google Scholar] [CrossRef] [PubMed]
- Avilés-Reyes, A.; Miller, J.H.; Simpson-Haidaris, P.J.; Lemos, J.A.; Abranches, J. Cnm is a major virulence factor of invasive Streptococcus mutans and part of a con-served three-gene locus. Mol. Oral Microbiol. 2014, 29, 11–23. [Google Scholar] [CrossRef] [PubMed]
- Miller, J.H.; Avilés-Reyes, A.; Scott-Anne, K.; Gregoire, S.; Watson, G.E.; Sampson, E.; Progulske-Fox, A.; Koo, H.; Bowen, W.H.; Lemos, J.A.; et al. The collagen binding protein Cnm contributes to oral colonization and cariogenicity of Streptococcus mutans OMZ175. Infect. Immun. 2015, 83, 2001–2010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Falsetta, M.L.; Klein, M.I.; Colonne, P.M.; Scott-Anne, K.; Gregoire, S.; Pai, C.H.; Gonzalez-Begne, M.; Watson, G.; Krysan, D.J.; Bowen, W.H.; et al. Symbiotic relationship between Streptococcus mutans and Candida albicans synergizes virulence of plaque biofilms in vivo. Infect. Immun. 2014, 82, 1968–1981. [Google Scholar] [CrossRef] [Green Version]
- McCarthy, H.; Rudkin, J.K.; Black, N.S.; Gallagher, L.; O’Neill, E.; O’Gara, J.P. Methicillin resistance and the biofilm phenotype in Staphylococcus aureus. Front. Cell. Infect. Microbiol. 2015, 5, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Nadell, C.D.; Drescher, K.; Foster, K.R. Spatial structure, cooperation and competition in biofilms. Nat. Rev. Microbiol. 2016, 14, 589–600. [Google Scholar] [CrossRef]
- Lohse, M.B.; Gulati, M.; Johnson, A.D.; Nobile, C.J. Development and regulation of single- and multi-species Candida albicans biofilms. Nat. Rev. Microbiol. 2018, 16, 19–31. [Google Scholar] [CrossRef] [Green Version]
- Boles, B.R.; Horswill, A.R. Agr-mediated dispersal of Staphylococcus aureus biofilms. PLoS Pathog. 2008, 4, e1000052. [Google Scholar] [CrossRef]
- Speziale, P.; Pietrocola, G.; Foster, T.J.; Geoghegan, J.A. Protein-based biofilm matrices in Staphylococci. Front. Cell. Infect. Microbiol. 2014, 4, 171. [Google Scholar] [CrossRef] [Green Version]
- Kırmusaoğlu, S. Staphylococcal biofilms: Pathogenicity, mechanism and regulation of biofilm formation by quorum-sensing system and antibiotic resistance mechanisms of biofilm-embedded microorganisms. In Microbial Biofilms—Importance and Applications, 1st ed.; Dhanasekaran, D., Thajuddin, N., Eds.; InTech: Rijeka, Croatia, 2016; pp. 189–209. [Google Scholar] [CrossRef]
- Pozzi, C.; Waters, E.M.; Rudkin, J.K.; Schaeffer, C.R.; Lohan, A.J.; Tong, P.; Loftus, P.J.; Pier, G.B.; Fey, P.D.; Massey, R.C.; et al. Methicillin resistance alters the biofilm phenotype and attenuates virulence in Staphylococcus aureus device-associated infections. PLoS Pathog. 2012, 8, e1002626. [Google Scholar] [CrossRef] [PubMed]
- Rudkin, J.K.; Edwards, A.M.; Bowden, M.G.; Brown, E.L.; Pozzi, C.; Waters, E.M.; Chan, W.C.; Williams, P.; O’Gara, J.P.; Massey, R.C. Methicillin resistance reduces the virulence of healthcare-associated methicillin-resistant Staphylococcus aureus by interfering with the agr quorum sensing system. J. Infect. Dis. 2012, 205, 798–806. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Queck, S.Y.; Khan, B.A.; Wang, R.; Bach, T.H.; Kretschmer, D.; Chen, L.; Kreiswirth, B.N.; Peschel, A.; DeLeo, F.R.; Otto, M. Mobile genetic element-encoded cytolysin connects virulence to methicillin resistance in MRSA. PLoS Pathog. 2009, 5, e1000533. [Google Scholar] [CrossRef] [PubMed]
- Salam, A.M.; Quave, C.L. Targeting Virulence in Staphylococcus aureus by Chemical Inhibition of the Accessory Gene Regulator System In Vivo. mSphere 2018, 3, e00500-17. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| S. aureus [Terminology] | Place of Isolation, Description | Relevant Characteristics | PVL | Source |
|---|---|---|---|---|
| SF8300_USA300 wild type [USA300-MRSA WT] | San Francisco, California, USA, CA-MRSA from an abscess | Multi-resistant 1 USA300, CC8, SCCmec IV 2 | + | [28] |
| HU13N [USA800-MRSA] | Rio de Janeiro, Brazil, CA-MRSA nasal isolate | Non-multi-resistant 3, USA800, CC5, SCCmec IV 2 | − | [29] This study |
| PN35 [ST72-MSSA-] | Rio de Janeiro, Brazil, MSSA, periodontal pocket isolate | Methicillin-sensitive 4, ST72/CC8- | − | This study |
| OMZ1122 [ST72-MSSA+] | Seattle, USA, ATCC 25923 | Methicillin-sensitive, ST72 | + | [30] |
| SF8300_ΔMSCRAMM [USA300-MRSA ΔMSCRAMM] | Mutant strain from SF8300_USA300 | In-frame deletions of clfA, clfB, sdrC, sdrD, sdrE, fnbA-fnbB | + | This study |
| Organism | Name | FA 1 (%) | WB 2 (mM) | Sequence (5′, 3′) | Source |
|---|---|---|---|---|---|
| S. aureus | Saur229 | 40 | 46 | CTAATGCAGCGCGGATCC | [7] |
| S. oralis | MIT446 | 25 | 149 | ACACYCGTTCTTCTCTTACAA | [10] |
| S. mutans | MUT590 | 30 | 112 | ACTCCAGACTTTCCTGAC | [36] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schnurr, E.; Paqué, P.N.; Attin, T.; Nanni, P.; Grossmann, J.; Holtfreter, S.; Bröker, B.M.; Kohler, C.; Diep, B.A.; Ribeiro, A.d.A.; et al. Staphylococcus aureus Interferes with Streptococci Spatial Distribution and with Protein Expression of Species within a Polymicrobial Oral Biofilm. Antibiotics 2021, 10, 116. https://doi.org/10.3390/antibiotics10020116
Schnurr E, Paqué PN, Attin T, Nanni P, Grossmann J, Holtfreter S, Bröker BM, Kohler C, Diep BA, Ribeiro AdA, et al. Staphylococcus aureus Interferes with Streptococci Spatial Distribution and with Protein Expression of Species within a Polymicrobial Oral Biofilm. Antibiotics. 2021; 10(2):116. https://doi.org/10.3390/antibiotics10020116
Chicago/Turabian StyleSchnurr, Etyene, Pune N. Paqué, Thomas Attin, Paolo Nanni, Jonas Grossmann, Silva Holtfreter, Barbara M. Bröker, Christian Kohler, Binh An Diep, Apoena de Aguiar Ribeiro, and et al. 2021. "Staphylococcus aureus Interferes with Streptococci Spatial Distribution and with Protein Expression of Species within a Polymicrobial Oral Biofilm" Antibiotics 10, no. 2: 116. https://doi.org/10.3390/antibiotics10020116
APA StyleSchnurr, E., Paqué, P. N., Attin, T., Nanni, P., Grossmann, J., Holtfreter, S., Bröker, B. M., Kohler, C., Diep, B. A., Ribeiro, A. d. A., & Thurnheer, T. (2021). Staphylococcus aureus Interferes with Streptococci Spatial Distribution and with Protein Expression of Species within a Polymicrobial Oral Biofilm. Antibiotics, 10(2), 116. https://doi.org/10.3390/antibiotics10020116