
Synergistic Activity of Equol and Meropenem against Carbapenem-Resistant Escherichia coli


Abstract
:1. Introduction
2. Results
2.1. Detection of Carbapenem-Resistant E. coli
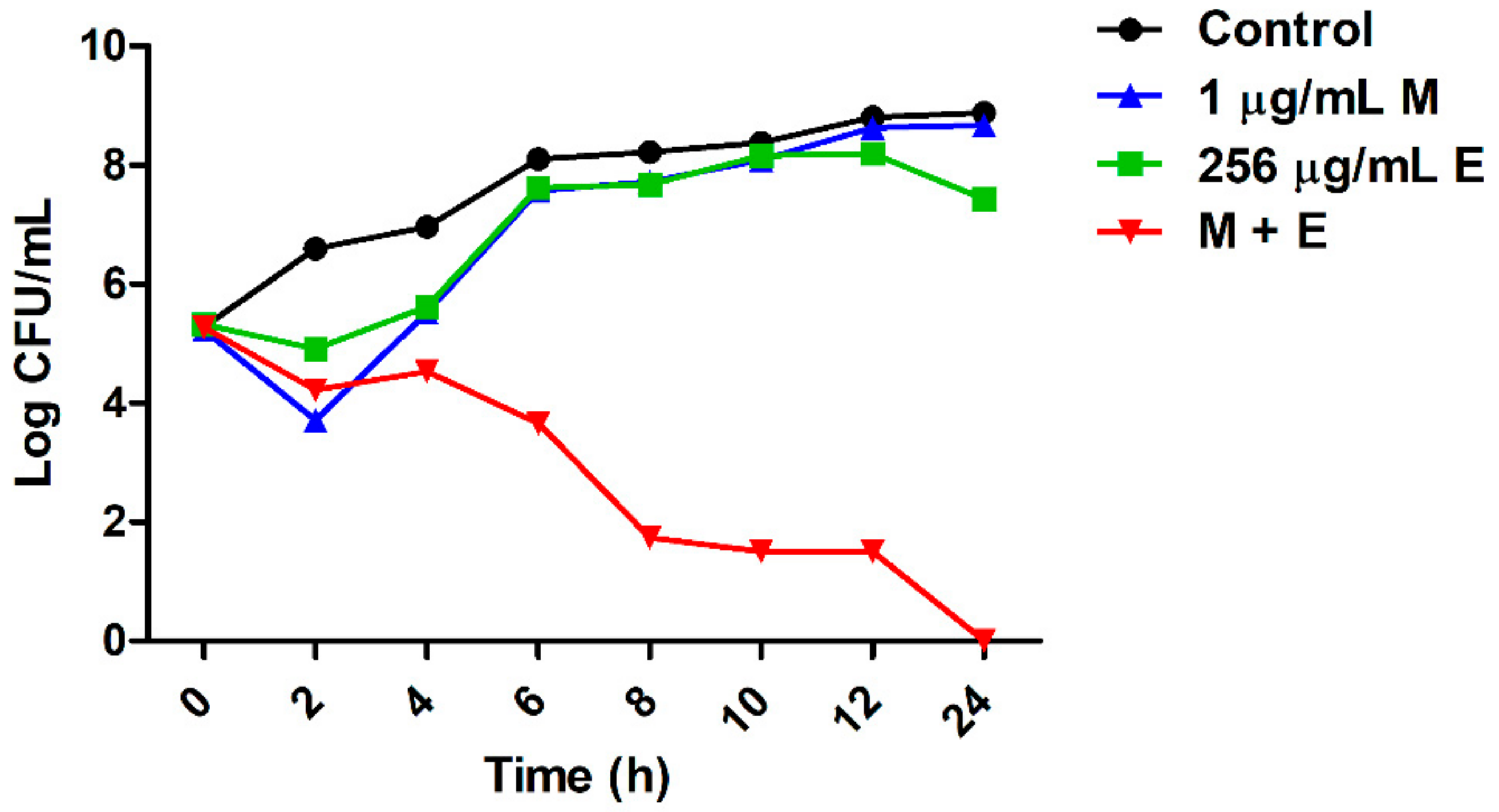
2.2. Equol Restores the Susceptibility of Carbapenem-Resistant E. coli to Meropenem
2.3. Anti-Biofilm Effects of Equol against Carbapenem-Resistant E. coli
2.4. Equol Suppresses the Motility of Carbapenem-Resistant E. coli
2.5. Equol Suppressed the Expression Levels of Carbapenemase- and Virulence-Related Genes in E. coli
3. Discussion
4. Materials and Methods
4.1. Organisms, Culture Conditions, and Reagents
4.2. Minimum Inhibitory Concentration Assay
4.3. DNA Extraction and Polymerase Chain Reaction (PCR) Amplification
4.4. Synergy Checkerboard Assay
4.5. Time–Kill Assay
4.6. Biofilm Inhibition and Eradication Assay
4.7. Motility Inhibition Assay
4.8. RNA Isolation
4.9. Quantitative Polymerase Chain Reaction (qPCR)
4.10. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Van Duin, D.; Doi, Y. The global epidemiology of carbapenemase-producing Enterobacteriaceae. Virulence 2017, 8, 460–469. [Google Scholar] [CrossRef]
- Baroud, M.; Dandache, I.; Araj, G.F.; Wakim, R.; Kanj, S.; Kanafani, Z.; Khairallah, M.; Sabra, A.; Shehab, M.; Dbaibo, G.; et al. Underlying mechanisms of carbapenem resistance in extended-spectrum beta-lactamase-producing Klebsiella pneumoniae and Escherichia coli isolates at a tertiary care centre in Lebanon: Role of OXA-48 and NDM-1 carbapenemases. Int. J. Antimicrob. Agents. 2013, 41, 75–79. [Google Scholar] [CrossRef]
- Suay-Garcia, B.; Pérez-Gracia, M.T. Present and Future of Carbapenem-resistant Enterobacteriaceae (CRE) Infections. Antibiotics 2019, 8, 122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Logan, L.K.; Weinstein, R.A. The Epidemiology of Carbapenem-Resistant Enterobacteriaceae: The Impact and Evolution of a Global Menace. J. Infect. Dis. 2017, 215, S28–S36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Queenan, A.M.; Bush, K. Carbapenemases: The versatile beta-lactamases. Clin. Microbiol. Rev. 2007, 20, 440–458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tenaillon, O.; Skurnik, D.; Picard, B.; Denamur, E. The population genetics of commensal Escherichia coli. Nat. Rev. Genet. 2010, 8, 207–217. [Google Scholar] [CrossRef]
- Kaper, J.B.; Nataro, J.P.; Mobley, H.L.T. Pathogenic Escherichia coli. Nat. Rev. Genet. 2004, 2, 123–140. [Google Scholar] [CrossRef]
- Gophna, U.; Barlev, M.; Seijffers, R.; Oelschlager, T.A.; Hacker, J.; Ron, E.Z. Curli Fibers Mediate Internalization ofEscherichia coli by Eukaryotic Cells. Infect. Immun. 2001, 69, 2659–2665. [Google Scholar] [CrossRef] [Green Version]
- Kao, C.Y.; Lin, W.H.; Tseng, C.C.; Wu, A.B.; Wang, M.C.; Wu, J.-J. The complex interplay among bacterial motility and virulence factors in different Escherichia coli infections. Eur. J. Clin. Microbiol. Infect. Dis. 2014, 33, 2157–2162. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Wen, Y. The role of bacterial biofilm in persistent infections and control strategies. Int. J. Oral Sci. 2011, 3, 66–73. [Google Scholar] [CrossRef] [Green Version]
- Li, X.-H.; Lee, J.-H. Antibiofilm agents: A new perspective for antimicrobial strategy. J. Microbiol. 2017, 55, 753–766. [Google Scholar] [CrossRef] [PubMed]
- Donlan, R.M. Biofilms: Microbial Life on Surfaces. Emerg. Infect. Dis. 2002, 8, 881–890. [Google Scholar] [CrossRef]
- Poirel, L.; Madec, J.Y.; Lupo, A.; Schink, A.K.; Kieffer, N.; Nordmann, P.; Schwarz, S. Antimicrobial resistance in Escherichia coli. Microbiol Spectr. 2018, 6. [Google Scholar] [CrossRef] [Green Version]
- Rizzo, K.; Horwich-Scholefield, S.; Epson, E. Carbapenem and Cephalosporin Resistance among Enterobacteriaceae in Healthcare-Associated Infections, California, USA. Emerg. Infect. Dis. 2019, 25, 1389–1393. [Google Scholar] [CrossRef] [Green Version]
- Sanhueza, L.; Melo, R.; Montero, R.; Maisey, K.; Mendoza, L.; Wilkens, M. Synergistic interactions between phenolic compounds identified in grape pomace extract with antibiotics of different classes against Staphylococcus aureus and Escherichia coli. PLoS ONE 2017, 12, e0172273. [Google Scholar] [CrossRef] [PubMed]
- Teethaisong, Y.; Autarkool, N.; Sirichaiwetchakoon, K.; Krubphachaya, P.; Kupittayanant, S.; Eumkeb, G. Synergistic activity and mechanism of action of Stephania suberosa Forman extract and ampicillin combination against ampicillin-resistant Staphylococcus aureus. J. Biomed. Sci. 2014, 21, 90. [Google Scholar] [CrossRef] [Green Version]
- Moussaoui, F.; Alaoui, T. Evaluation of antibacterial activity and synergistic effect between antibiotic and the essential oils of some medicinal plants. Asian Pac. J. Trop. Biomed. 2016, 6, 32–37. [Google Scholar] [CrossRef] [Green Version]
- Aiyegoro, O.A.; Afolayan, A.J.; Okoh, A.I. Synergistic interaction of Helichrysum pedunculatum leaf extracts with antibiotics against wound infection associated bacteria. Biol. Res. 2009, 42, 327–338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adwan, G.; Mhanna, M. Synergistic effects of plant extracts and antibiotics on Staphylococcus aureus strains isolated from clinical specimens. Asian Pac. J. Trop Med. 2009, 2, 46–51. [Google Scholar]
- Hossain, M.A.; Park, H.C.; Park, S.W.; Park, S.C.; Seo, M.G.; Her, M.; Kang, J. Synergism of the combination of traditional antibiotics and novel phenolic compounds against Escherichia coli. Pathogens 2020, 9, 811. [Google Scholar] [CrossRef]
- Setchell, K.D.R.; Brown, N.M.; Desai, P.; Zimmer-Nechemias, L.; Wolfe, B.E.; Brashear, W.T.; Kirschner, A.S.; Cassidy, A.; Heubi, J.E. Bioavailability of Pure Isoflavones in Healthy Humans and Analysis of Commercial Soy Isoflavone Supplements. J. Nutr. 2001, 131, 1362S–1375S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mayo, B.; Vázquez, L.; Flórez, A.B. Equol: A Bacterial Metabolite from the Daidzein Isoflavone and Its Presumed Beneficial Health Effects. Nutrients 2019, 11, 2231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hwang, J.; Wang, J.; Morazzoni, P.; Hodis, H.N.; Sevanian, A. The phytoestrogen equol increases nitric oxide availability by inhibiting superoxide production: An antioxidant mechanism for cell-mediated LDL modification. Free. Radic. Biol. Med. 2003, 34, 1271–1282. [Google Scholar] [CrossRef]
- Setchell, K.; Clerici, C.; Lephart, E.D.; Cole, S.J.; Heenan, C.; Castellani, D.; Wolfe, B.E.; Nechemias-Zimmer, L.; Brown, N.M.; Lund, T.D.; et al. S-Equol, a potent ligand for estrogen receptor β, is the exclusive enantiomeric form of the soy isoflavone metabolite produced by human intestinal bacterial flora. Am. J. Clin. Nutr. 2005, 81, 1072–1079. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jackson, R.L.; Greiwe, J.S.; Schwen, R.J. Emerging evidence of the health benefits of S-equol, an estrogen receptor β agonist. Nutr. Rev. 2011, 69, 432–448. [Google Scholar] [CrossRef]
- Chacko, B.K.; Chandler, R.T.; Mundhekar, A.; Khoo, N.; Pruitt, H.M.; Kucik, D.F.; Parks, D.A.; Kevil, C.G.; Barnes, S.; Patel, R.P. Revealing anti-inflammatory mechanisms of soy isoflavones by flow: Modulation of leukocyte-endothelial cell interactions. Am. J. Physiol. Circ. Physiol. 2005, 289, H908–H915. [Google Scholar] [CrossRef] [Green Version]
- Itsumi, M.; Shiota, M.; Takeuchi, A.; Kashiwagi, E.; Inokuchi, J.; Tatsugami, K.; Kajioka, S.; Uchiumi, T.; Naito, S.; Eto, M.; et al. Equol inhibits prostate cancer growth through degradation of androgen receptor by S-phase kinase-associated protein 2. Cancer Sci. 2016, 107, 1022–1028. [Google Scholar] [CrossRef] [PubMed]
- Charalambous, C.; Pitta, C.A.; Constantinou, A.I. Equol enhances tamoxifen’s anti-tumor activity by induction of caspase-mediated apoptosis in MCF-7 breast cancer cells. BMC Cancer 2013, 13, 238. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, Y.; Kimura, S.; Ishii, Y.; Tateda, K.; Yumi, T.; Soichiro, K.; Yoshikazu, I.; Kazuhiro, T. Equol inhibits growth and spore formation of Clostridioides difficile. J. Appl. Microbiol. 2019, 127, 932–940. [Google Scholar] [CrossRef]
- Lee, J.A.; Chee, H.Y. In VitroAntifungal Activity of Equol againstCandida albicans. Mycobiology 2010, 38, 328–330. [Google Scholar] [CrossRef] [Green Version]
- Roca, I.; Akova, M.; Baquero, F.; Carlet, J.; Cavaleri, M.; Coenen, S.; Cohen, J.H.M.; Findlay, D.; Gyssens, I.C.J.; Heure, O.E.; et al. The global threat of antimicrobial resistance: Science for intervention. New Microbes New Infect. 2015, 6, 22–29. [Google Scholar] [CrossRef] [Green Version]
- Xu, Y.; Gu, B.; Huang, M.; Liu, H.; Xu, T.; Xia, W.; Wang, T. Epidemiology of carbapenem resistant Enterobacteriaceae (CRE) during 2000-2012 in Asia. J. Thorac. Dis. 2015, 7, 376–385. [Google Scholar] [PubMed]
- Tyers, M.; Wright, G.D. Drug combinations: A strategy to extend the life of antibiotics in the 21st century. Nat. Rev. Genet. 2019, 17, 141–155. [Google Scholar] [CrossRef] [PubMed]
- Torella, J.P.; Chait, R.; Kishony, R. Optimal drug synergy in antimicrobial treatments. PLoS Comput Biol. 2010, 6, e1000796. [Google Scholar] [CrossRef]
- Lomovskaya, O.; Warren, M.S.; Lee, A.; Galazzo, J.; Fronko, R.; Lee, M.; Blais, J.; Cho, D.; Chamberland, S.; Renau, T.; et al. Identification and Characterization of Inhibitors of Multidrug Resistance Efflux Pumps in Pseudomonas aeruginosa: Novel Agents for Combination Therapy. Antimicrob. Agents Chemother. 2001, 45, 105–116. [Google Scholar] [CrossRef] [Green Version]
- González-Bello, C. Antibiotic adjuvants—A strategy to unlock bacterial resistance to antibiotics. Bioorganic Med. Chem. Lett. 2017, 27, 4221–4228. [Google Scholar] [CrossRef]
- Mahmood, H.Y.; Jamshidi, S.; Sutton, J.M.; Rahman, K.M. Current Advances in Developing Inhibitors of Bacterial Multidrug Efflux Pumps. Curr. Med. Chem. 2016, 23, 1062–1081. [Google Scholar] [CrossRef]
- Allen, R.C.; Popat, R.; Diggle, S.P.; Brown, S.P. Targeting virulence: Can we make evolution-proof drugs? Nat. Rev. Microbiol. 2014, 12, 300–308. [Google Scholar] [CrossRef]
- Yoon, E.-J.; Yang, J.W.; Kim, J.O.; Lee, H.; Lee, K.J.; Jeong, S.H. Carbapenemase-producing Enterobacteriaceae in South Korea: A report from the National Laboratory Surveillance System. Futur. Microbiol. 2018, 13, 771–783. [Google Scholar] [CrossRef]
- Park, S.H.; Kim, J.S.; Kim, H.S.; Yu, J.K.; Han, S.H.; Kang, M.J.; Hong, C.K.; Lee, S.M.; Oh, Y.H. Prevalence of carbapenem-resistant Enterobacteriaceae in Seoul, Korea. J. Bacteriol. Virol. 2020, 50, 107–116. [Google Scholar]
- Rondevaldova, J.; Hummelova, J.; Tauchen, J.; Kokoska, L. In vitro anti-staphylococcal synergistic effect of isoflavone metabolite demethyltexasin with amoxicillin and oxacillin. Microb. Drug Resist. 2018, 24, 24–29. [Google Scholar] [CrossRef] [PubMed]
- Stewart, P.S.; Costerton, J.W. Antibiotic resistance of bacteria in biofilms. Lancet 2001, 358, 135–138. [Google Scholar] [CrossRef]
- Barnhart, M.M.; Chapman, M.R. Curli Biogenesis and Function. Annu. Rev. Microbiol. 2006, 60, 131–147. [Google Scholar] [CrossRef] [Green Version]
- Arnqvist, A.; Olsén, A.; Pfeifer, J.; Russell, D.G.; Normark, S. The Crl protein activates cryptic genes for curli formation and fibronectin binding in Escherichia coli HB101. Mol. Microbiol. 2006, 6, 2443–2452. [Google Scholar] [CrossRef]
- Robbe-Saule, V.; Jaumouille, V.; Prevost, M.C.; Guadagnini, S.; Talhouarne, C.; Mathout, H.; Kolb, A.; Norel, F. Crl activates transcription initiation of RpoS-regulated genes involved in the multicellular behavior of Salmonella enterica serovar Typhimurium. J. Bacteriol. 2006, 188, 3983–3994. [Google Scholar] [CrossRef] [Green Version]
- Fahrner, K.A.; Berg, H.C. Mutations that stimulate flhDC expression in Escherichia coli K-12. J. Bacteriol. 2015, 197, 3087–3096. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fitzgerald, D.M.; Bonocora, R.P.; Wade, J.T. Comprehensive Mapping of the Escherichia coli Flagellar Regulatory Network. PLoS Genet. 2014, 10, e1004649. [Google Scholar] [CrossRef] [Green Version]
- Gauger, E.J.; Leatham, M.P.; Mercado-Lubo, R.; Laux, D.C.; Conway, T.; Cohen, P.S. Role of Motility and the flhDC Operon in Escherichia coli MG1655 Colonization of the Mouse Intestine. Infect. Immun. 2007, 75, 3315–3324. [Google Scholar] [CrossRef] [Green Version]
- Clinical and Laboratory Standards Institute (CLSI). Methods for Dilution Antimicrobial Susceptibility Tests for Bacteria that Grow Aerobically; CLSI: Wayne, PA, USA, 2012. [Google Scholar]
- Kim, H.R.; Eom, Y.B. Antifungal and anti-biofilm effects of 6-shogaol against Candida auris. J. Appl. Microbiol. 2020. [Google Scholar] [CrossRef]
- Poirel, L.; Walsh, T.R.; Cuvillier, V.; Nordmann, P. Multiplex PCR for detection of acquired carbapenemase genes. Diagn. Microbiol. Infect. Dis. 2011, 70, 119–123. [Google Scholar] [CrossRef]
- Hindiyeh, M.; Smollen, G.; Grossman, Z.; Ram, D.; Davidson, Y.; Mileguir, F.; Vax, M.; Ben David, D.; Tal, I.; Rahav, G.; et al. Rapid Detection of blaKPC Carbapenemase Genes by Real-Time PCR. J. Clin. Microbiol. 2008, 46, 2879–2883. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smiljanic, M.; Kaase, M.; Ahmad-Nejad, P.; Ghebremedhin, B. Comparison of in-house and commercial real time-PCR based carbapenemase gene detection methods in Enterobacteriaceae and non-fermenting gram-negative bacterial isolates. Ann. Clin. Microbiol. Antimicrob. 2017, 16, 48. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.-H.; Kim, Y.-G.; Raorane, C.J.; Ryu, S.Y.; Shim, J.-J.; Lee, J. The anti-biofilm and anti-virulence activities of trans-resveratrol and oxyresveratrol against uropathogenic Escherichia coli. Biofouling 2019, 35, 758–767. [Google Scholar] [CrossRef] [PubMed]
- Knobl, T.; Moreno, A.M.; Paixao, R.; Gomes, T.A.; Vieira, M.A.; da Silva Leite, D.; Blanco, J.E.; Ferreira, A.J. Prevalence of avian pathogenic Escherichia coli (APEC) clone harboring sfa gene in Brazil. Sci. World J. 2012, 2012, 437342. [Google Scholar] [CrossRef] [Green Version]
- Kakkanat, A.; Phan, M.-D.; Lo, A.W.; Beatson, S.A.; Schembri, M.A. Novel genes associated with enhanced motility of Escherichia coli ST131. PLoS ONE 2017, 12, e0176290. [Google Scholar] [CrossRef] [Green Version]
- Lane, M.C.; Simms, A.N.; Mobley, H.L.T. Complex Interplay between Type 1 Fimbrial Expression and Flagellum-Mediated Motility of Uropathogenic Escherichia coli. J. Bacteriol. 2007, 189, 5523–5533. [Google Scholar] [CrossRef] [Green Version]
- Shin, D.-S.; Eom, Y.-B. Antimicrobial and antibiofilm activities of Clostridium butyricum supernatant against Acinetobacter baumannii. Arch. Microbiol. 2020, 202, 1059–1068. [Google Scholar] [CrossRef]
- Eumkeb, G.; Siriwong, S.; Thumanu, K. Synergistic activity of luteolin and amoxicillin combination against amoxicillin-resistant Escherichia coli and mode of action. J. Photochem. Photobiol. B Biol. 2012, 117, 247–253. [Google Scholar] [CrossRef] [PubMed]
- Odds, F.C. Synergy, antagonism, and what the chequerboard puts between them. J. Antimicrob. Chemother. 2003, 52, 1. [Google Scholar] [CrossRef]
- Woo, S.-G.; Lee, S.-Y.; Lim, K.-H.; Ha, E.-J.; Eom, Y.-B. Activity of novel inhibitors of Staphylococcus aureus biofilms. Folia Microbiol. 2017, 62, 157–167. [Google Scholar] [CrossRef] [PubMed]
- Jang, H.; Eom, Y.-B. Repurposing auranofin to combat uropathogenic Escherichia coli biofilms. J. Appl. Microbiol. 2019, 127, 459–471. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.-R.; Shin, D.-S.; Jang, H.-I.; Eom, Y.-B. Anti-biofilm and anti-virulence effects of zerumbone against Acinetobacter baumannii. Microbiology 2020, 166, 717–726. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(T)(-Delta Delta C) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Carbapenem | MIC (μg/mL) | |
|---|---|---|
| E. coli KBN12P05816 | E. coli KBN12P06081 | |
| Meropenem | 16 | 16 |
| Imipenem | ≥4 | ≥4 |
| Ertapenem | 64 | 32 |
| Genes of Carbapenemase | Bacterial Strains | |
|---|---|---|
| E. coli KBN12P05816 | E. coli KBN12P06081 | |
| blaKPC | + | + |
| blaIMP | − | − |
| blaVIM | − | − |
| blaNDM | − | − |
| blaOXA-48 | + | + |
| Strains | MIC (μg/mL) | Meropenem + Equol | |||
|---|---|---|---|---|---|
| Meropenem | Equol | Meropenem + Equol | FIC Index | Interpretation | |
| E. coli KBN12P05816 | 16 | 1024 | 1 + 256 | 0.31 | S |
| E. coli KBN12P06081 | 16 | 1024 | 1 + 256 | 0.31 | S |
| Primers | Target Gene | Primer Sequence (5’-3’) | Annealing Temp. (°C) | References |
|---|---|---|---|---|
| PCR primers | blaKPC | F: CGTCTAGTTCTGCTGTCTTG | 54 | [51] |
| R: CTTGTCATCCTTGTTAGGCG | ||||
| blaIMP | F: GGAATAGAGTGGCTTAAYTC | 51 | [51] | |
| R: TCGGTTTAAYAAAACAACCACC | ||||
| blaVIM | F: GATGGTGTTTGGTCGCATA | 56 | [51] | |
| R: CGAATGCGCAGCACCAG | ||||
| blaNDM | F: GGTTTGGCGATCTGGTTTTC | 56 | [51] | |
| R: CGGAATGGCTCATCACGATC | ||||
| blaOXA-48 | F: GCGTGGTTAAGGATGAACAC | 55 | [51] | |
| R: CATCAAGTTCAACCCAACCG | ||||
| qPCR primers | blaKPC | F: GATACCACGTTCCGTCTGG | 57 | [52] |
| R: GCAGGTTCCGGTTTTGTCTC | ||||
| blaOXA-48 | F: GGCACGTATGAGCAAGATGC | 59 | [53] | |
| R: GTTTGACAATACGCTGGCTGC | ||||
| csgA | F: GGGCTCAGATGACAGCTCAAT | 59 | [54] | |
| R: GCCGTTCCACTGATCAAGAGTAG | ||||
| csgB | F: CATAATTGGTCAAGCTGGGACTAA | 55 | [54] | |
| R: GCAACAACCGCCAAAAGTTT | ||||
| crl | F: TTTCGATTGTCTGGCTGTATG | 54 | [55] | |
| R: CTTCAGATTCAGCGTCGTC | ||||
| motA | F: ACAGGTAGCGCGTTCTCACT | 58 | [54] | |
| R: AGCGTGGATAAACCGATACG | ||||
| flhD | F: ACTTGCACAGCGTCTGATTG | 55 | [56] | |
| R: AGCTTAACCATTTGCGGAAG | ||||
| fliC | F: ACAGCCTCTCGCTGATCACTCAAA | 61 | [57] | |
| R: GCGCTGTTAATACGCAAGCCAGAA | ||||
| 16SrRNA | F: CAGCTCGTGTCGTGAGATGT | 60 | [58] | |
| R: CGTAAGGGCCATGATGACTT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, H.-R.; Eom, Y.-B. Synergistic Activity of Equol and Meropenem against Carbapenem-Resistant Escherichia coli. Antibiotics 2021, 10, 161. https://doi.org/10.3390/antibiotics10020161
Kim H-R, Eom Y-B. Synergistic Activity of Equol and Meropenem against Carbapenem-Resistant Escherichia coli. Antibiotics. 2021; 10(2):161. https://doi.org/10.3390/antibiotics10020161
Chicago/Turabian StyleKim, Hye-Rim, and Yong-Bin Eom. 2021. "Synergistic Activity of Equol and Meropenem against Carbapenem-Resistant Escherichia coli" Antibiotics 10, no. 2: 161. https://doi.org/10.3390/antibiotics10020161