
Clonal Lineages, Antimicrobial Resistance, and PVL Carriage of Staphylococcus aureus Associated to Skin and Soft-Tissue Infections from Ambulatory Patients in Portugal

 ,
,  ,
,
Abstract
:1. Introduction
2. Results
2.1. Antimicrobial Susceptibility Profile and Correlation with Resistance Determinants
2.2. Efflux Activity
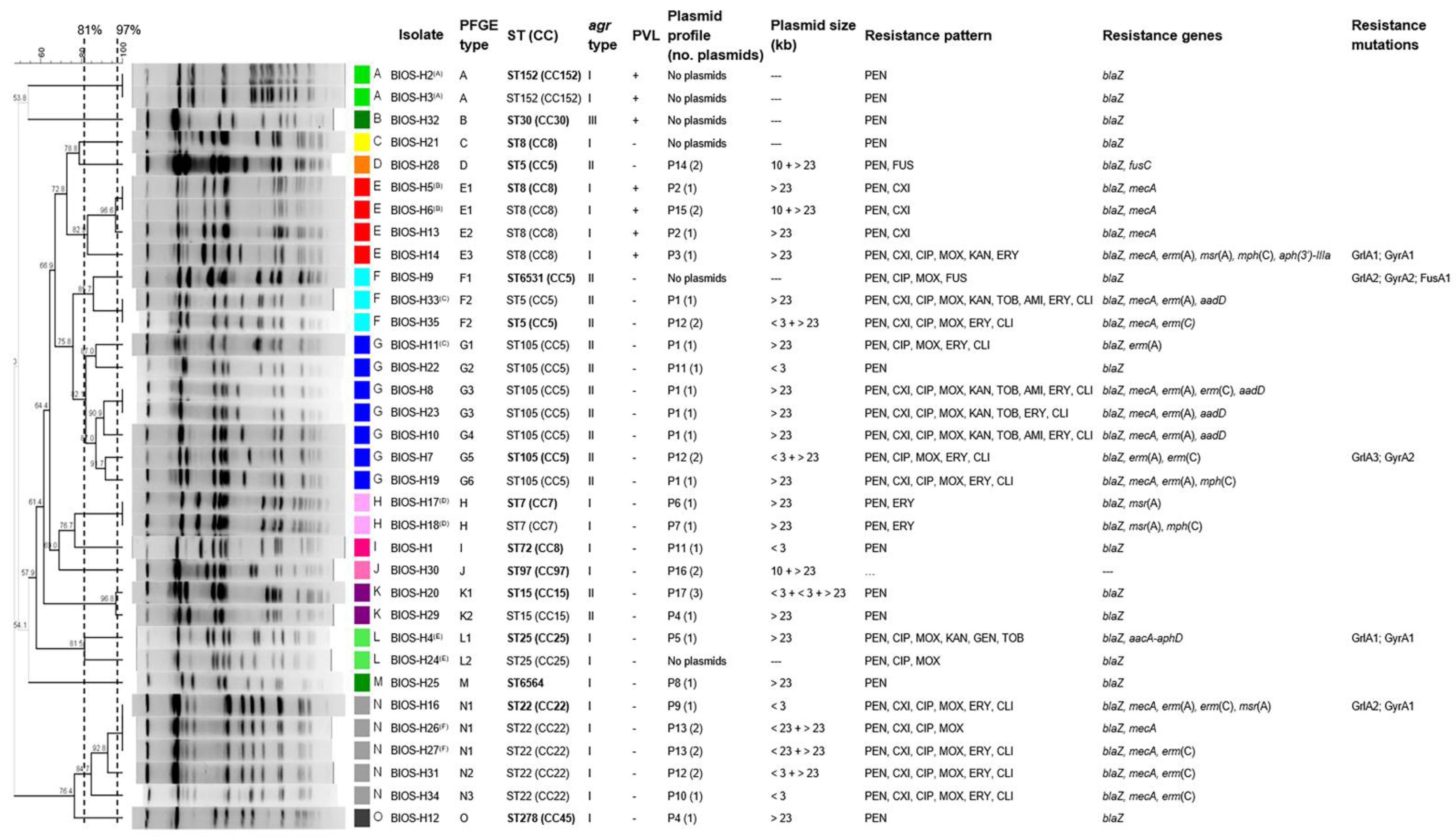
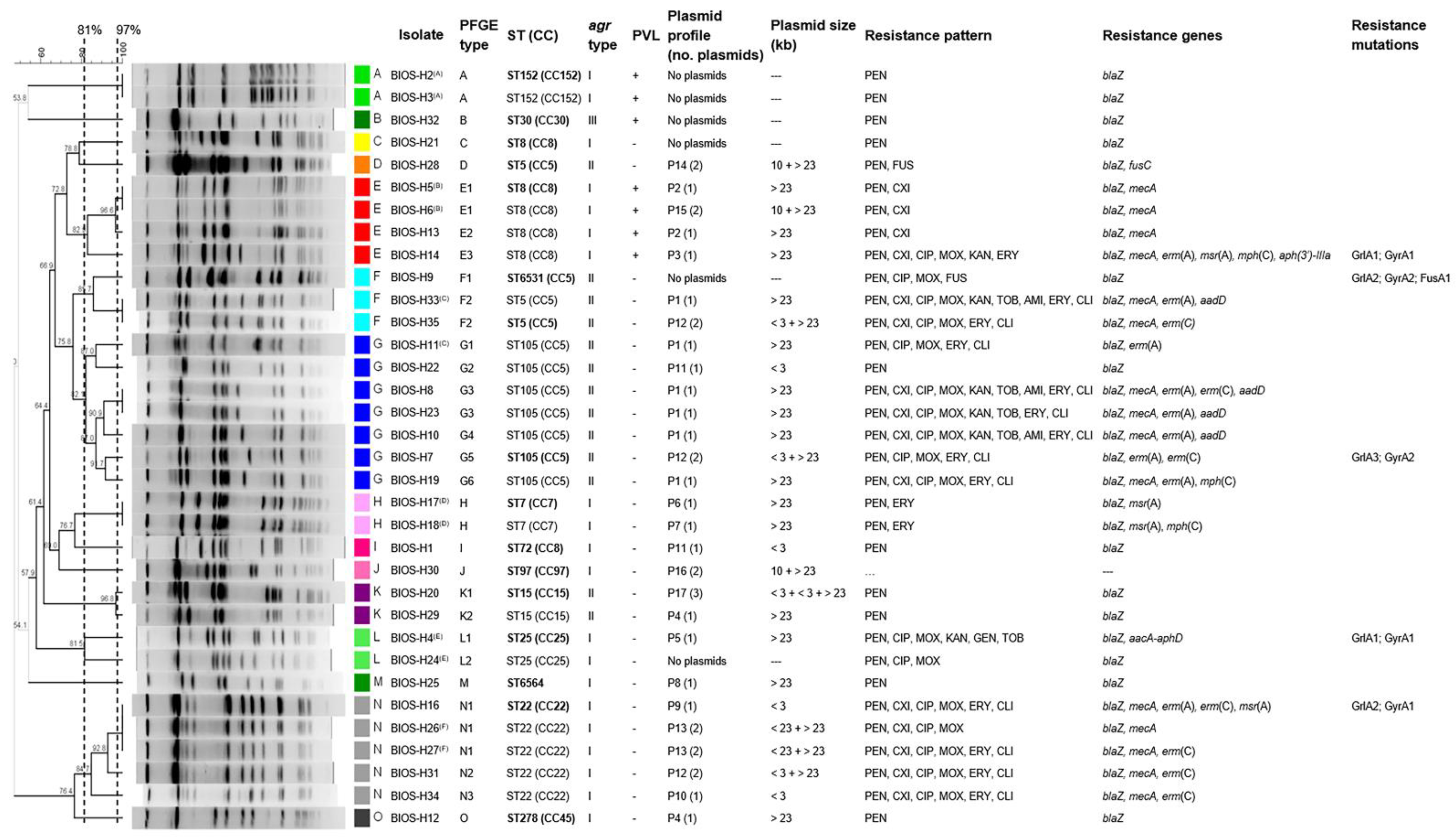
2.3. Main Clonal Lineages and Genetic Diversity of the S. aureus Isolates
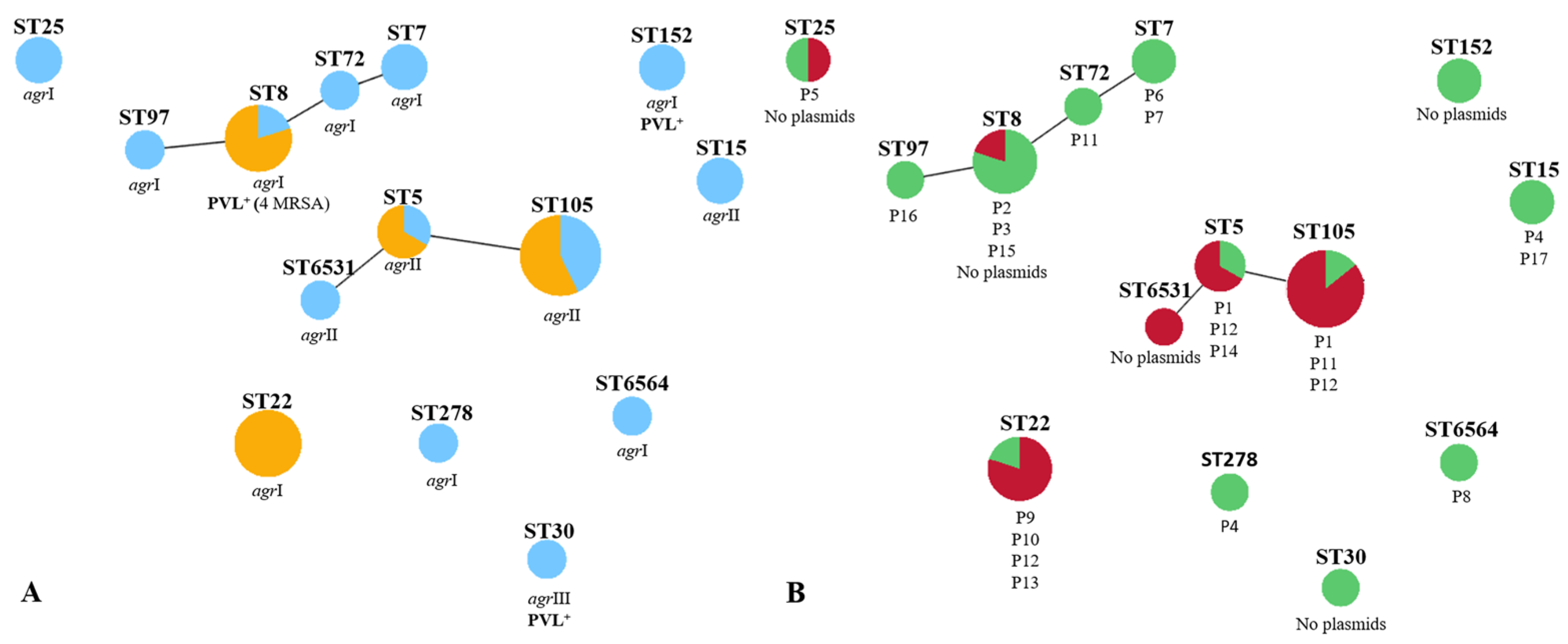
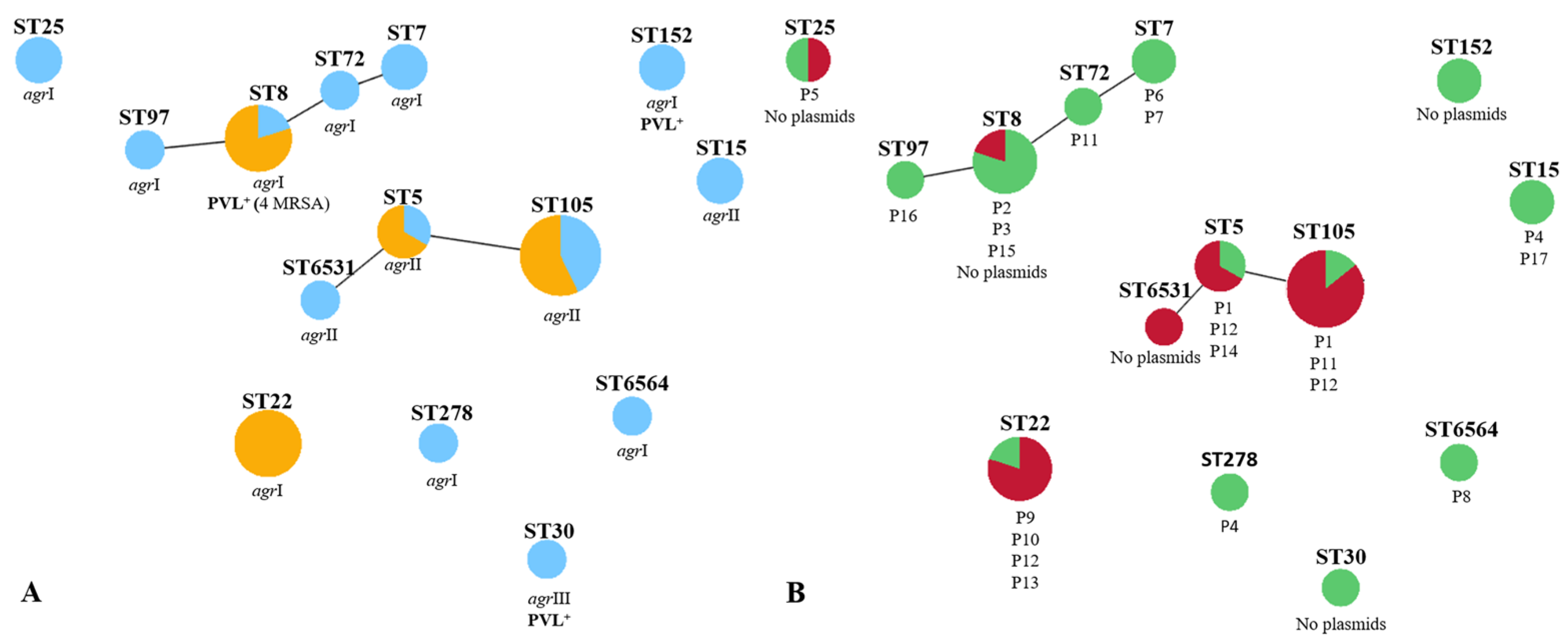
2.4. Correlation of Strain Lineage with agr Typing and PVL Carriage
2.5. Correlation of Strain Lineage with Antimicrobial Resistance and Plasmid Profiles
3. Discussion
4. Materials and Methods
4.1. Bacterial Isolates
4.2. Antimicrobial Susceptibility Testing
4.3. Detection of Resistance Genes by PCR
4.4. Screening of Mutations in grlA, gyrA, and fusA Genes
4.5. Evaluation of Efflux Activity
4.6. Plasmid DNA Extraction and Profiling
4.7. Detection of lukSF Genes
4.8. Molecular Typing
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Turner, N.A.; Sharma-Kuinkel, B.K.; Maskarinec, S.A.; Eichenberger, E.M.; Shah, P.P.; Carugati, M.; Holland, T.L.; Fowler, V.G., Jr. Methicillin-resistant Staphylococcus aureus: An overview of basic and clinical research. Nat. Rev. Microbiol. 2019, 17, 203–218. [Google Scholar] [CrossRef]
- Lee, A.S.; de Lencastre, H.; Garau, J.; Kluytmans, J.; Malhotra-Kumar, S.; Peschel, A.; Harbarth, S. Methicillin-resistant Staphylococcus aureus. Nat. Rev. Dis. Primers 2018, 4, 18033. [Google Scholar] [CrossRef]
- Foster, T. Antibiotic resistance in Staphylococcus aureus. Current status and future prospects. FEMS Microbiol. Rev. 2017, 41, 430–449. [Google Scholar] [CrossRef]
- Tacconelli, E.; Carrara, E.; Savoldi, A.; Harbarth, S.; Mendelson, M.; Monnet, D.L.; Pulcini, C.; Kahlmeter, G.; Kluytmans, J.; Carmeli, Y.; et al. Discovery, research, and development of new antibiotics: The WHO priority list of antibiotic-resistant bacteria and tuberculosis. Lancet Infect. Dis. 2018, 18, 318–327. [Google Scholar] [CrossRef]
- Olaniyi, R.; Pozzi, C.; Grimaldi, L.; Bagnoli, F. Staphylococcus aureus-associated skin and soft tissue infections: Anatomical localization, epidemiology, therapy and potential prophylaxis. Curr. Top. Microbiol. Immunol. 2016, 409, 199–227. [Google Scholar] [CrossRef]
- Liu, C.; Bayer, A.; Cosgrove, S.E.; Daum, R.S.; Fridkin, S.K.; Gorwitz, R.J.; Kaplan, S.L.; Karchmer, A.W.; Levine, D.P.; Murray, B.E.; et al. Clinical practice guidelines by the Infectious Diseases Society of America for the treatment of methicillin-resistant Staphylococcus aureus infections in adults and children. Clin. Infect. Dis. 2011, 52, e18–e55. [Google Scholar] [CrossRef] [PubMed]
- Esposito, S.; Bassetti, M.; Bonnet, E.; Bouza, E.; Chan, M.; De Simone, G.; Dryden, M.; Gould, I.; Lye, D.C.; Saeed, K.; et al. Hot topics in the diagnosis and management of skin and soft-tissue infections. Int. J. Antimicrob. Agents 2016, 48, 19–26. [Google Scholar] [CrossRef] [PubMed]
- Montravers, P.; Snauwaert, A.; Welsch, C. Current guidelines and recommendations for the management of skin and soft tissue infections. Curr. Opin. Infect. Dis. 2016, 29, 131–138. [Google Scholar] [CrossRef]
- Lee, G.C.; Dallas, S.D.; Wang, Y.; Olsen, R.J.; Lawson, K.A.; Wilson, J.; Frei, C.R. Emerging multidrug resistance in community-associated Staphylococcus aureus involved in skin and soft tissue infections and nasal colonization. J. Antimicrob. Chemother. 2017, 72, 2461–2468. [Google Scholar] [CrossRef]
- Davey, R.X.; Tong, S.Y.C. The epidemiology of Staphylococcus aureus skin and soft tissue infection in the southern Barkly region of Australia’s Northern Territory in 2017. Pathology 2019, 51, 308–312. [Google Scholar] [CrossRef]
- Stefanaki, C.; Ieronymaki, A.; Matoula, T.; Caroni, C.; Polythodoraki, E.; Chryssou, S.E.; Kontochristopoulos, G.; Antoniou, C. Six-year retrospective review of hospital data on antimicrobial resistance profile of Staphylococcus aureus isolated from skin infections from a single institution in Greece. Antibiotics 2017, 6, 39. [Google Scholar] [CrossRef]
- Watanabe, S.; Ohnishi, T.; Yuasa, A.; Kiyota, H.; Iwata, S.; Kaku, M.; Watanabe, A.; Sato, J.; Hanaki, H.; Manabe, M.; et al. The first nationwide surveillance of antibacterial susceptibility patterns of pathogens isolated from skin and soft-tissue infections in dermatology departments in Japan. J. Infect. Chemother. 2017, 23, 503–511. [Google Scholar] [CrossRef] [PubMed]
- Costa, S.S.; Viveiros, M.; Amaral, L.; Couto, I. Multidrug efflux pumps in Staphylococcus aureus: An update. Open Microbiol. J. 2013, 7, 59–71. [Google Scholar] [CrossRef] [PubMed]
- Lakhundi, S.; Zhang, K. Methicillin-Resistant Staphylococcus aureus: Molecular characterization, evolution, and epidemiology. Clin. Microbiol. Rev. 2018, 31. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Muir, T.W. Regulation of virulence in Staphylococcus aureus: Molecular mechanisms and remaining puzzles. Cell Chem. Biol. 2016, 23, 214–224. [Google Scholar] [CrossRef]
- Haaber, J.; Penadés, J.R.; Ingmer, H. Transfer of antibiotic resistance in Staphylococcus aureus. Trends Microbiol. 2017, 25, 893–905. [Google Scholar] [CrossRef]
- Jensen, S.O.; Lyon, B.R. Genetics of antimicrobial resistance in Staphylococcus aureus. Future Microbiol. 2009, 4, 565–582. [Google Scholar] [CrossRef]
- Faria, N.; Miragaia, M.; de Lencastre, H.; The Multi Laboratory Project Collaborators. Massive dissemination of methicillin resistant Staphylococcus aureus in bloodstream infections in a high MRSA prevalence country: Establishment and diversification of EMRSA-15. Microbial. Drug Resist. 2013, 19, 483–490. [Google Scholar] [CrossRef]
- Tavares, A.; Miragaia, M.; Rolo, J.; Coelho, C.; de Lencastre, H. High prevalence of hospital-associated methicillin-resistant Staphylococcus aureus in the community in Portugal: Evidence for the blurring of community–hospital boundaries. Eur. J. Clin. Microbiol. Infect. Dis. 2013, 32, 1269–1283. [Google Scholar] [CrossRef]
- Espadinha, D.; Faria, N.A.; Miragaia, M.; Lito, L.M.; Melo-Cristino, J.; de Lencastre, H.; Médicos Sentinela Network. Extensive dissemination of methicillin-resistant Staphylococcus aureus (MRSA) between the hospital and the community in a country with a high prevalence of nosocomial MRSA. PLoS ONE 2013, 8, e59960. [Google Scholar] [CrossRef]
- Conceição, T.; Diamantino, F.; Coelho, C.; de Lencastre, H.; Aires-de-Sousa, M. Contamination of public buses with MRSA in Lisbon, Portugal: A possible transmission route of major MRSA clones within the community. PLoS ONE 2013, 8, e77812. [Google Scholar] [CrossRef] [PubMed]
- Conceição, T.; Martins, H.; Rodrigues, S.; de Lencastre, H.; Aires-de-Sousa, M. Staphylococcus aureus nasal carriage among homeless population in Lisbon, Portugal. Eur. J. Clin. Microbiol. Infect. Dis. 2019, 38, 2037–2044. [Google Scholar] [CrossRef]
- Costa, S.S.; Junqueira, E.; Palma, C.; Viveiros, M.; Melo-Cristino, J.; Amaral, L.; Couto, I. Resistance to antimicrobials mediated by efflux pumps in Staphylococcus aureus. Antibiotics 2013, 2, 83–99. [Google Scholar] [CrossRef] [PubMed]
- European Centre for Disease Prevention and Control. Surveillance of Antimicrobial Resistance in Europe 2019; ECDC: Stockholm, Sweden, 2020. [Google Scholar]
- Conceição, T.; Aires-de-Sousa, M.; Pona, N.; Brito, M.J.; Barradas, C.; Coelho, R.; Sardinha, T.; Sancho, L.; de Sousa, G.; Machado, M.C.; et al. High prevalence of ST121 in community-associated methicillin-susceptible Staphylococcus aureus lineages responsible for skin and soft tissue infections in Portuguese children. Eur. J. Clin. Microbiol. Infect. Dis. 2011, 30, 293–297. [Google Scholar] [CrossRef]
- European Centre for Disease Prevention and Control. Antimicrobial Resistance Surveillance in Europe 2014. Annual Report of the European Antimicrobial Resistance Surveillance Network (EARS-NET); ECDC: Stockholm, Sweden, 2015. [Google Scholar]
- Livermore, D.M.; Wain, J. Revolutionising bacteriology to improve treatment outcomes and antibiotic stewardship. Infect. Chemother. 2013, 45, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Rolo, J.; Miragaia, M.; Turlej-Rogacka, A.; Empel, J.; Bouchami, O.; Faria, N.A.; Tavares, A.; Hryniewicz, W.; Fluit, A.C.; de Lencastre, H.; et al. High genetic diversity among community-associated Staphylococcus aureus in Europe: Results from a multicenter study. PLoS ONE 2012, 7, e34768. [Google Scholar] [CrossRef] [PubMed]
- Tavares, A.; Faria, N.A.; de Lencastre, H.; Miragaia, M. Population structure of methicillin-susceptible Staphylococcus aureus (MSSA) in Portugal over a 19-year period (1192-2011). Eur. J. Clin. Microbiol. Infect. Dis. 2014, 33, 423–432. [Google Scholar] [CrossRef]
- Sun, J.; Yang, M.; Sreevatsan, S.; Bender, J.B.; Singer, R.S.; Knutson, T.P.; Marthaler, D.G.; Davies, P.R. Longitudinal study of Staphylococcus aureus colonization and infection in a cohort of swine veterinarians in the United States. BMC Infect. Dis. 2017, 17, 690. [Google Scholar] [CrossRef]
- David, M.Z.; Siegel, J.D.; Henderson, J.; Leos, G.; Lo, K.; Iwuora, J.; Taylor, A.R.; Zychowski, D.L.; Porsa, E.; Boyle-Vavra, S.; et al. Hand and nasal carriage of discordant Staphylococcus aureus isolates among urban jail detainees. J. Clin. Microbiol. 2014, 52, 3422–3425. [Google Scholar] [CrossRef]
- Rasslan, O.S.E.; Khater, W.S.; Elnour, S.S.A.; Asaad, M.K. Fusidic acid resistance among Staphylococcus aureus causing community acquired skin and soft tissue infections. Egypt J. Med. Lab. Sci. 2016, 25, 21–31. [Google Scholar]
- Liu, Y.; Geng, W.; Yang, Y.; Wang, C.; Zheng, Y.; Shang, Y.; Wu, D.; Li, X.; Wang, L.; Yu, S.; et al. Susceptibility to and resistance determinants of fusidic acid in Staphylococcus aureus isolated from Chinese children with skin and soft tissue infections. FEMS Immunol. Med. Microbiol. 2012, 64, 212–218. [Google Scholar] [CrossRef]
- Bessa, G.R.; Quinto, V.P.; Machado, D.C.; Lipnharski, C.; Weber, M.B.; Bonamigo, R.R.; D’Azevedo, P.A. Staphylococcus aureus resistance to topical antimicrobials in atopic dermatitis. Anais Bras. Dermatol. 2016, 91, 604–610. [Google Scholar] [CrossRef]
- Stein, M.; Komerska, J.; Prizade, M.; Sheinberg, B.; Tasher, D.; Somekh, E. Clindamycin resistance among Staphylococcus aureus strains in Israel: Implications for empirical treatment of skin and soft tissue infections. Int. J. Infect. Dis. 2016, 46, 18–21. [Google Scholar] [CrossRef]
- Vicetti, M.C.P.; Mejias, A.; Leber, A.; Sanchez, P.J. A decade of antimicrobial resistance in Staphylococcus aureus: A single center experience. PLoS ONE 2019, 14, e0212029. [Google Scholar] [CrossRef]
- Mclaws, F.B.; Larsen, A.R.; Skov, R.L.; Chopra, I.; O’Neill, A.J. Distribution of fusidic acid resistance determinants in methicillin-resistant Staphylococcus aureus. Antimicrob. Agents Chemother. 2011, 55, 1173–1176. [Google Scholar] [CrossRef]
- Chen, H.J.; Hung, W.; Tseng, S.; Tsai, J.; Hsueh, P.; Teng, L. Fusidic acid resistance determinants in Staphylococcus aureus clinical isolates. Antimicrob. Agents Chemother. 2010, 54, 4985–4991. [Google Scholar] [CrossRef]
- Besier, S.; Ludwig, A.; Brade, V.; Wichelhaus, T.A. Molecular analysis of fusidic acid resistance in Staphylococcus aureus. Mol. Microbiol. 2003, 47, 463–469. [Google Scholar] [CrossRef]
- Castanheira, M.; Watters, A.A.; Bell, J.M.; Turnidge, J.; Jones, R.N. Fusidic acid resistance rates and prevalence of resistance mechanisms among Staphylococcus spp. isolated in North America and Australia, 2007–2008. Antimicrob. Agents Chemother. 2010, 54, 3614–3617. [Google Scholar] [CrossRef] [PubMed]
- Lannergård, T.; Norstrom, T.; Hughes, D. Genetic determinants of resistance to fusidic acid among clinical bacteremia isolates of Staphylococcus aureus. Antimicrob. Agents Chemother. 2009, 53, 2059–2065. [Google Scholar] [CrossRef] [PubMed]
- Ellington, M.J.; Reuterb, S.; Harrisb, S.R.; Holden, M.T.G.; Cartwright, E.J.; Greaves, D.; Gerver, S.M.; Hope, R.; Brown, N.M.; Török, M.E.; et al. Emergent and evolving antimicrobial resistance cassettes in community-associated fusidic acid and meticillin-resistant Staphylococcus aureus. Int. J. Antimicrob. Agents 2015, 45, 477–484. [Google Scholar] [CrossRef] [PubMed]
- Baines, S.L.; Howden, B.P.; Heffernan, H.; Stinear, T.P.; Carter, G.P.; Seeman, T.; Kwong, J.C.; Ritchie, S.R.; Williamson, D.A. Rapid emergence and evolution of Staphylococcus aureus clones harboring fusC-containing staphylococcal cassette chromosome elements. Antimicrob. Agents Chemother. 2016, 60, 2359–2365. [Google Scholar] [CrossRef]
- Schmitz, F.J.; Jones, M.E.; Hofmann, B.; Hansen, B.; Scheuring, S.; Luckefahr, M.; Fluit, A.; Verhoef, J.; Hadding, U.; Heinz, H.P.; et al. Characterization of grlA, grlB, gyrA, and gyrB Mutations in 116 unrelated isolates of Staphylococcus aureus and effects of mutations on ciprofloxacin MIC. Antimicrob. Agents Chemother. 1998, 42, 1249–1252. [Google Scholar] [CrossRef]
- Fitzgibbon, J.E.; John, J.F.; Delucia, J.L.; Dubin, D.T. Topoisomerase mutations in trovafloxacin-resistant Staphylococcus aureus. Antimicrob. Agents Chemother. 1998, 42, 2122–2124. [Google Scholar] [CrossRef]
- Costa, S.S.; Falcão, C.; Viveiros, M.; Machado, D.; Martins, M.; Melo-Cristino, J.; Amaral, L.; Couto, I. Exploring the contribution of efflux on the resistance to fluoroquinolones in clinical isolates of Staphylococcus aureus. BMC Microbiol. 2011, 11, 241. [Google Scholar] [CrossRef]
- Sierra, J.M.; Marco, F.; Ruiz, J.; de Anta, M.T.J.; Vila, J. Correlation between the activity of different fluoroquinolones and the presence of mechanisms of quinolone resistance in epidemiologically related and unrelated strains of methicillin-susceptible and -resistant Staphylococcus aureus. Clin. Microbiol. Infect. 2002, 8, 781–790. [Google Scholar] [CrossRef]
- Trong, H.N.; Prunier, A.L.; Leclercq, R. Hypermutable and fluoroquinolone-resistant clinical isolates of Staphylococcus aureus. Antimicrob. Agents Chemother. 2005, 49, 2098–2101. [Google Scholar] [CrossRef] [PubMed]
- Sanfilippo, C.M.; Hesje, C.K.; Haas, W.; Morris, T.W. Topoisomerase mutations that are associated with high-level resistance to earlier fluoroquinolones in Staphylococcus aureus have less effect on the antibacterial activity of besifloxacin. Chemotherapy 2011, 57, 363–371. [Google Scholar] [CrossRef]
- Marasa, B.S.; Iram, S.; Sung, k.; Kweon, O.; Cerniglia, C.E.; Khan, S. Molecular characterization of fluoroquinolone resistance of methicillin–resistant clinical Staphylococcus aureus isolates from Rawalpindi, Pakistan. Med. Res. Arch. 2015, 2. [Google Scholar] [CrossRef]
- Fuzi, M.; Szabo, D.; Csercsik, R. Double-serine fluoroquinolone resistance mutations advance major international clones and lineages of various multi-drug resistant bacteria. Front. Microbiol. 2017, 8, 2261. [Google Scholar] [CrossRef]
- Costa, S.S.; Palma, C.; Kladec, K.; Fessler, A.T.; Viveiros, M.; Melo-Cristino, J.; Schwarz, S.; Couto, I. Plasmid-borne antimicrobial resistance of Staphylococcus aureus isolated in a hospital in Lisbon, Portugal. Microb. Drug Resist. 2016, 22, 617–626. [Google Scholar] [CrossRef]
- David, M.Z.; Daum, R.S. Community-associated methicillin-resistant Staphylococcus aureus: Epidemiology and clinical consequences of an emerging epidemic. Clin. Microbiol. Rev. 2010, 23, 616–687. [Google Scholar] [CrossRef]
- Shallcross, L.J.; Fragaszy, E.; Johnson, A.M.; Hayward, A.C. The role of the Panton-Valentine leucocidin toxin in staphylococcal disease: A systematic review and meta-analysis. Lancet Infect. Dis. 2013, 13, 43–54. [Google Scholar] [CrossRef]
- Castro, A.; Komora, N.; Ferreira, V.; Lira, A.; Mota, M.; Silva, J.; Teixeira, P. Prevalence of Staphylococcus aureus from nares and hands on health care professionals in a Portuguese Hospital. J. Appl. Microbiol. 2016, 121, 831–839. [Google Scholar] [CrossRef] [PubMed]
- Mottola, C.; Semedo-Lemsaddek, T.; Mendes, J.J.; Melo-Cristino, J.; Tavares, L.; Cavaco-Silva, P.; Oliveira, M. Molecular typing, virulence traits and antimicrobial resistance of diabetic foot staphylococci. J. Biomed. Sci. 2016, 23, 33. [Google Scholar] [CrossRef]
- Poulsen, A.B.; Skov, R.; Pallesen, L.V. Detection of methicillin resistance in coagulase-negative staphylococci and in staphylococci directly from simulated blood cultures using the EVIGENE MRSA Detection Kit. J. Antimicrob. Chemother. 2003, 51, 419–421. [Google Scholar] [CrossRef]
- European Committee on Antimicrobial Susceptibility Testing. Breakpoint Tables for Interpretation of MICs and Zone Diameters; Version 10.0; 2020; Available online: http://www.eucast.org (accessed on 28 December 2020).
- Clinical and Laboratory Standards Institute. M100 Performance Standards for Antimicrobial Suscetibility Testing, 30th ed.; Clinical and Laboratory Standards Institute: Annapolis, MD, USA, 2020. [Google Scholar]
- Traczewski, M.M.; Brown, S.D. Proposed MIC and disk diffusion microbiological cutoffs and spectrum of activity of retapamulin, a novel topical antibiotic. Antimicrob. Agents Chemother. 2008, 52, 3863–3867. [Google Scholar] [CrossRef] [PubMed]
- Tenover, F.C. Mechanisms of antimicrobial resistance in bacteria. Am. J. Infect. Control 2006, 34, S3–S10. [Google Scholar] [CrossRef]
- Alexopoulou, K.; Foka, A.; Petinaki, E.; Jelastopulu, E.; Dimitracopoulos, G.; Spiliopoulou, I. Comparison of two commercial methods with PCR restriction fragment length polymorphism of the tuf gene in the identification of coagulase-negative staphylococci. Lett. Appl. Microbiol. 2006, 43, 450–454. [Google Scholar] [CrossRef]
- Patel, D.; Kosmidis, C.; Seo, S.M.; Kaatz, G.W. Ethidium bromide MIC screening for enhanced efflux pump gene expression or efflux activity in Staphylococcus aureus. Antimicrob. Agents Chemother. 2010, 54, 5070–5073. [Google Scholar] [CrossRef]
- Chung, M.; de Lencastre, H.; Matthews, P.; Tomasz, A.; Adamsson, I.; Aires-de-Sousa, M.; Camou, T.; Cocuzza, T.; Corso, A.; Couto, I.; et al. Molecular typing of methicillin-resistant Staphylococcus aureus by pulsed-field gel electrophoresis: Comparison of results obtained in a multilaboratory effort using identical protocols and MRSA strains. Microb. Drug Resist. 2000, 6, 189–198. [Google Scholar] [CrossRef]
- Carriço, J.A.; Pinto, F.R.; Simas, C.; Nunes, S.; Sousa, N.G.; Frazão, N.; de Lencastre, H.; Almeida, J.S. Assessment of band-based similarity coefficients for automatic type and subtype classification of microbial isolates analyzed by pulsed-field gel electrophoresis. J. Clin. Microbiol. 2005, 43, 5483–5490. [Google Scholar] [CrossRef]
- Carriço, J.A.; Silva-Costa, C.; Melo-Cristino, J.; Pinto, F.R.; de Lencastre, H.; Almeida, J.S.; Ramirez, M. Illustration of a common framework for relating multiple typing methods by application to macrolide-resistant Streptococcus pyogenes. J. Clin. Microbiol. 2006, 44, 2524–2532. [Google Scholar] [CrossRef]
- Enright, M.C.; Day, N.P.; Davies, C.E.; Peacock, S.J.; Spratt, B.G. Multilocus sequence typing for characterization of methicillin-resistant and methicillin-susceptible clones of Staphylococcus aureus. J. Clin. Microbiol. 2000, 38, 1008–1015. [Google Scholar] [CrossRef]
- Crisóstomo, M.I.; Westh, H.; Tomasz, A.; Chung, M.; Oliveira, D.C.; de Lencastre, H. The evolution of methicillin resistance in Staphylococcus aureus: Similarity of genetic backgrounds in historically early methicillin susceptible and resistant isolates and contemporary epidemic clones. Proc. Natl. Acad. Sci. USA 2001, 98, 9865–9870. [Google Scholar] [CrossRef]
- Francisco, A.P.; Vaz, C.; Monteiro, P.T.; Melo-Cristino, J.; Ramirez, M.; Carriço, J.A. PHYLOViZ: Phylogenetic inference and data visualization for sequence based typing methods. BMC Bioinform. 2012, 13, 87. [Google Scholar] [CrossRef]
- Lina, G.; Boutite, F.; Tristan, A.; Bes, M.; Etienne, J.; Vandenesch, F. Bacterial competition for human nasal cavity colonization: Role of Staphylococcal agr alleles. Appl. Environ. Microbiol. 2003, 69, 18–23. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
| Class | Antibiotic | Resistant Isolates (%) | Resistance Determinants (No. Isolates); [Mutations] |
|---|---|---|---|
| β-lactams | PEN | 33 (97.1%) | blaZ (33) mecA (15) |
| CXI | 15 (44.1%) | ||
| Fluoroquinolones | CIP | 17 (50%) | Mutations in GrlA [S80Y, E84G, S80F] and GyrA [S84L, E88K] |
| MOX | 17 (50%) | ||
| Macrolides/Lincosamides | ERY | 15 (44.1%) | erm(A) (9), erm(C) (7) msr(A) (4), mph(C) (3) |
| CLI | 12 (35.3%) | ||
| Aminoglycosides | KAN | 6 (17.6%) | aadD (4), aacA-aphD (1) aph(3′)-IIIa (1) |
| TOB | 5 (14.7%) | ||
| AMI | 3 (8.8%) | ||
| GEN | 1 (2.9%) | ||
| Fusidanes | FUS | 2 (5.9%) | fusC (1) FusA mutations [A71V, H457Q, G476C] |
| Isolate | MIC (μg/mL) | |||||
|---|---|---|---|---|---|---|
| EtBr | EtBr + TZ | EtBr + VER | CIP | CIP + TZ | CIP + VER | |
| BIOS-H4 | 16 | 4 | 2 | 16 | 16 | 16 |
| BIOS-H7 | 16 | 8 | 8 | 512 | 256 | 256 |
| BIOS-H8 | 16 | 8 | 4 | 512 | 256 | 256 |
| BIOS-H10 | 16 | 8 | 4 | 512 | 256 | 256 |
| BIOS-H11 | 16 | 4 | 2 | 256 | 128 | 128 |
| BIOS-H14 | 16 | 4 | 2 | 128 | 64 | 64 |
| BIOS-H19 | 16 | 4 | 2 | 512 | 256 | 256 |
| BIOS-H23 | 16 | 4 | 4 | 256 | 128 | 256 |
| BIOS-H24 | 16 | 2 | 2 | 32 | 16 | 16 |
| BIOS-H31 | 16 | 4 | 2 | 512 | 128 | 128 |
| BIOS-H33 | 16 | 4 | 2 | 512 | 256 | 256 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ferreira, C.; Costa, S.S.; Serrano, M.; Oliveira, K.; Trigueiro, G.; Pomba, C.; Couto, I. Clonal Lineages, Antimicrobial Resistance, and PVL Carriage of Staphylococcus aureus Associated to Skin and Soft-Tissue Infections from Ambulatory Patients in Portugal. Antibiotics 2021, 10, 345. https://doi.org/10.3390/antibiotics10040345
Ferreira C, Costa SS, Serrano M, Oliveira K, Trigueiro G, Pomba C, Couto I. Clonal Lineages, Antimicrobial Resistance, and PVL Carriage of Staphylococcus aureus Associated to Skin and Soft-Tissue Infections from Ambulatory Patients in Portugal. Antibiotics. 2021; 10(4):345. https://doi.org/10.3390/antibiotics10040345
Chicago/Turabian StyleFerreira, Carolina, Sofia Santos Costa, Maria Serrano, Ketlyn Oliveira, Graça Trigueiro, Constança Pomba, and Isabel Couto. 2021. "Clonal Lineages, Antimicrobial Resistance, and PVL Carriage of Staphylococcus aureus Associated to Skin and Soft-Tissue Infections from Ambulatory Patients in Portugal" Antibiotics 10, no. 4: 345. https://doi.org/10.3390/antibiotics10040345
APA StyleFerreira, C., Costa, S. S., Serrano, M., Oliveira, K., Trigueiro, G., Pomba, C., & Couto, I. (2021). Clonal Lineages, Antimicrobial Resistance, and PVL Carriage of Staphylococcus aureus Associated to Skin and Soft-Tissue Infections from Ambulatory Patients in Portugal. Antibiotics, 10(4), 345. https://doi.org/10.3390/antibiotics10040345