
Natural and Enantiopure Alkylglycerols as Antibiofilms Against Clinical Bacterial Isolates and Quorum Sensing Inhibitors of Chromobacterium violaceum ATCC 12472


 ,
,
Abstract
:1. Introduction
2. Results
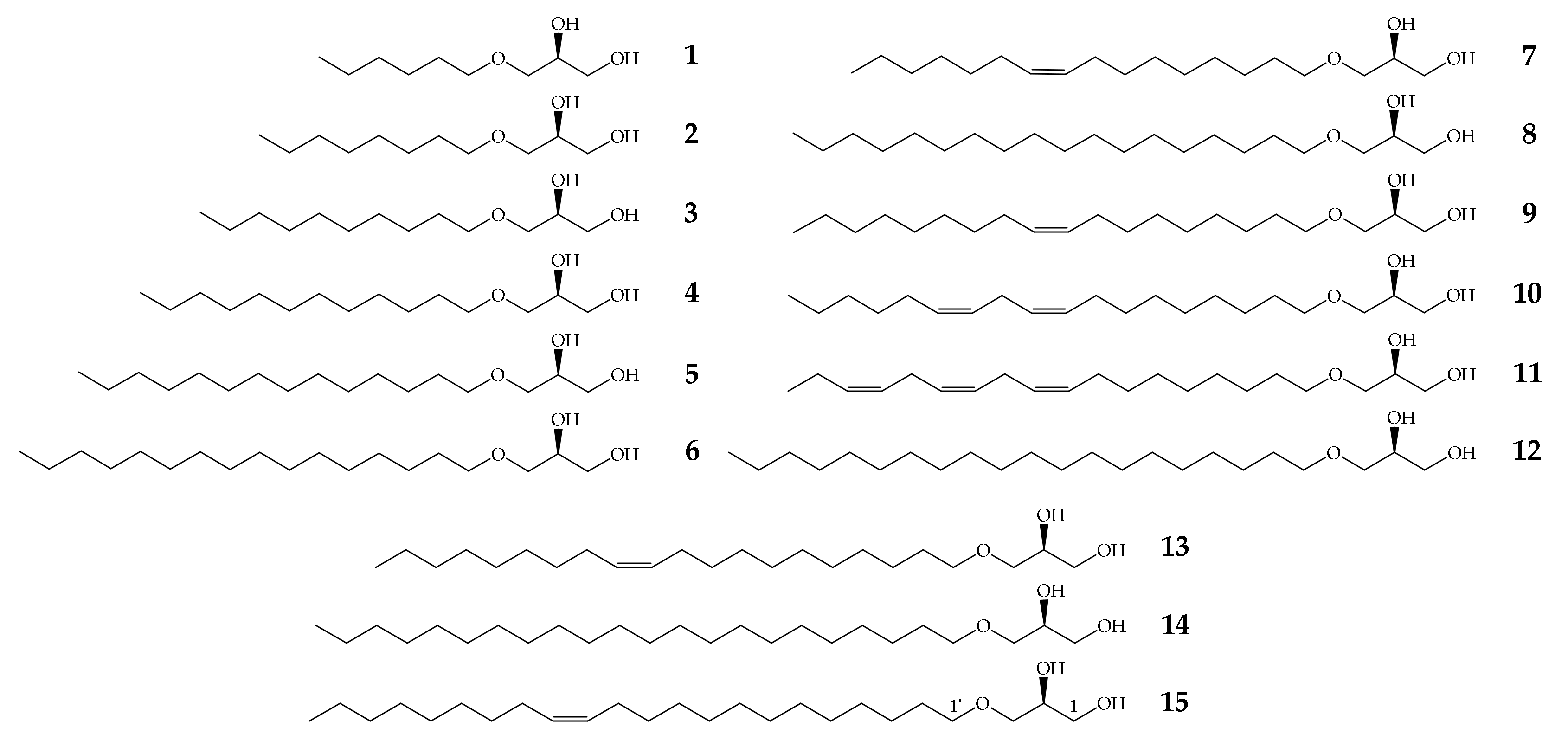
2.1. Structures of Natural and Enantiopure Alkylglycerols
2.2. Minimal Inhibitory Concentration (MIC) of Alkylglycerols
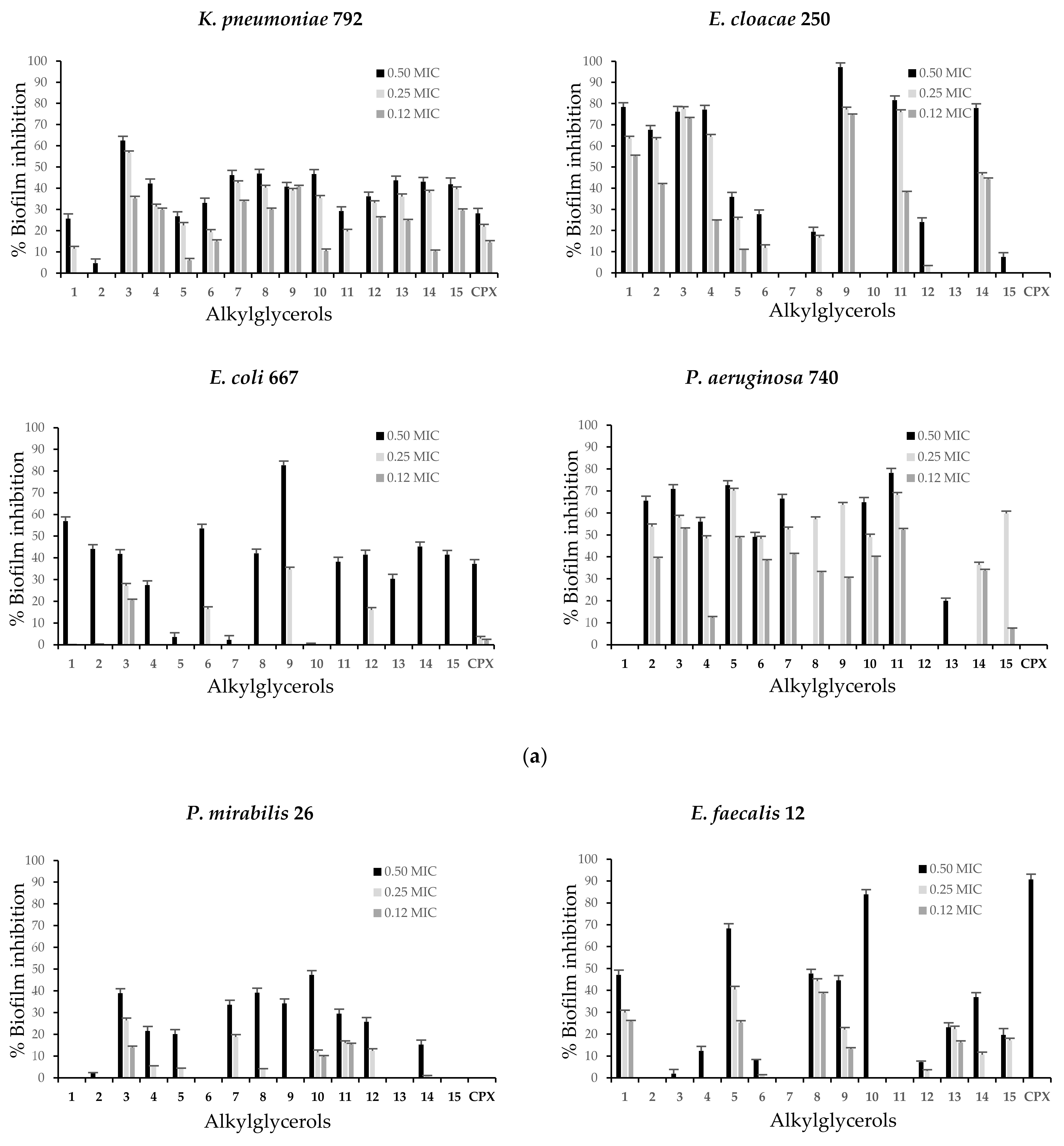
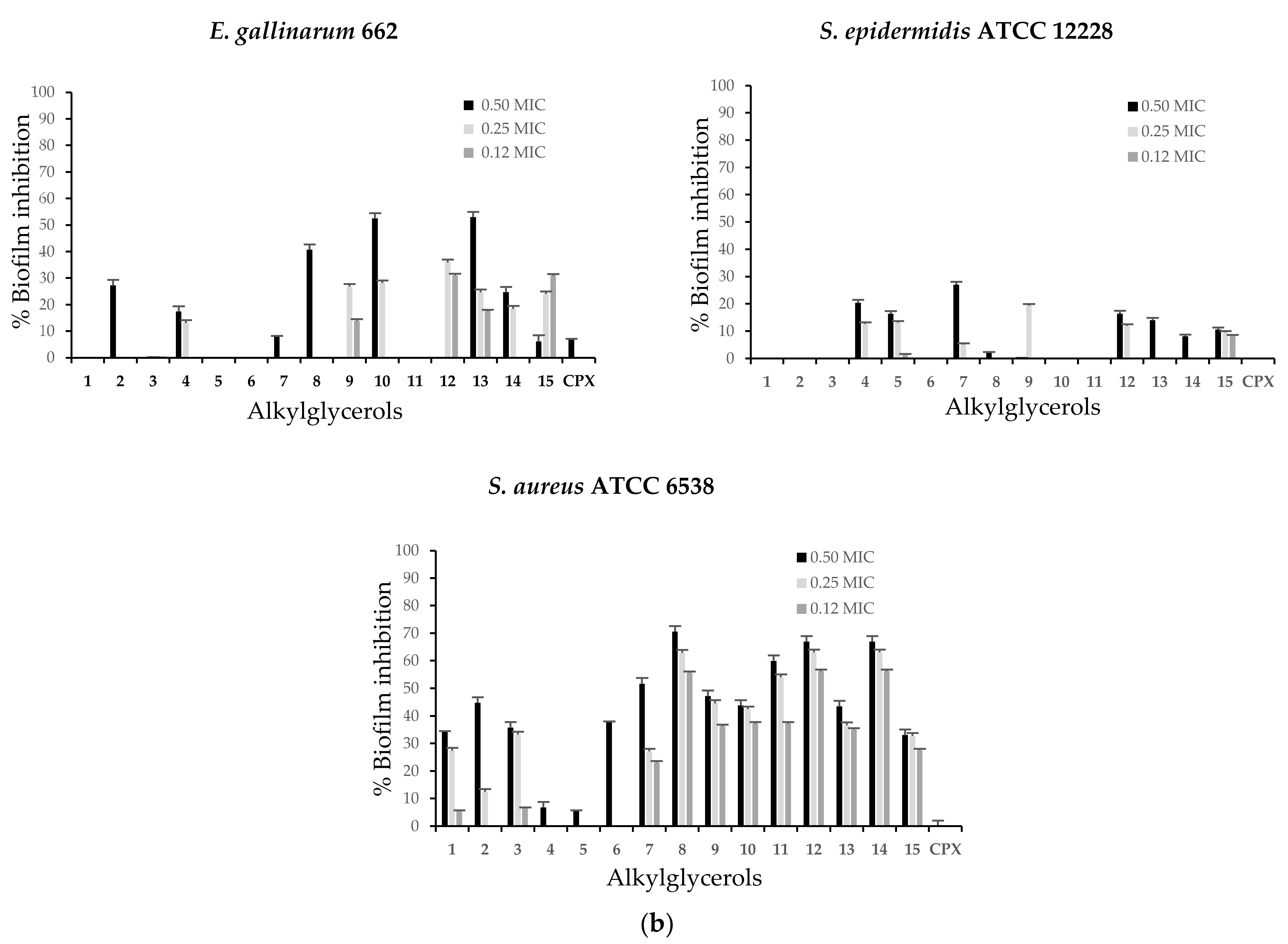
2.3. Alkylglycerols Ability to Inhibit of Biofilm Formation
2.4. Quorum Sensing Inhibition Activity of Natural Alkylglycerols
3. Discussion
4. Materials and Methods
4.1. General
4.1.1. Screened Compounds
4.1.2. Bacterial Strains and Culture Conditions
4.2. Determination of the Minimum Inhibitory Concentration of Screened Compounds
4.3. Effectiveness of Alkylglycerols on the Inhibition of Biofilm Formation
4.4. Screening for Quorum Sensing Inhibition Activity
4.5. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Aydemir, D.H.; Cifci, G.; Aviyente, V.; Tinaz, G.B. Quorum-sensing inhibitor potential of trans-anethole against Pseudomonas aeruginosa. J. Appl. Microbiol. 2018, 125, 731–739. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.; Wang, P.C.; Ma, H.M.; Chen, S.Y.; Fu, Y.H.; Liu, Y.Y.; Wang, X.; Yu, G.C.; Huang, T.; Hibbs, D.E.; et al. Design, synthesis and evaluation of halogenated furanone derivatives as quorum sensing inhibitors in Pseudomonas aeruginosa. Eur. J. Pharm. Sci. 2019, 140, 105058. [Google Scholar] [CrossRef] [PubMed]
- Pacios, O.; Blasco, L.; Bleriot, I.; Fernandez, L.G.; González, M.B.; Ambroa, A.; López, M.; Bou, G.; Tomás, M. Strategies to combat multidrug-resistant and persistent infectious diseases. Antibiotics 2020, 9, 65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reina, J.C.; Pérez, I.V.; Martín, J.; Llamas, I. A quorum-sensing inhibitor strain of Vibrio alginolyticus blocks Qs-controlled phenotypes in Chromobacterium violaceum and Pseudomonas aeruginosa. Mar. Drugs 2019, 17, 494. [Google Scholar] [CrossRef] [Green Version]
- Fong, J.; Yuan, M.; Jakobsen, T.H.; Mortensen, K.T.; Delos Santos, M.M.; Chua, S.L.; Yang, L.; Tan, C.H.; Nielsen, T.E.; Givskov, M. Disulfide bond-containing ajoene analogs as novel quorum sensing inhibitors of Pseudomonas aeruginosa. J. Med. Chem. 2017, 60, 215–227. [Google Scholar] [CrossRef]
- Xu, X.J.; Zeng, T.; Huang, Z.X.; Xu, X.F.; Lin, J.; Chen, W.M. Synthesis and biological evaluation of cajaninstilbene acid and amorfrutins A and B as inhibitors of the Pseudomonas aeruginosa quorum sensing system. J. Nat. Prod. 2018, 81, 2621–2629. [Google Scholar] [CrossRef]
- Li, C.; Jiang, C.; Jing, H.; Jiang, C.; Wang, H.; Du, X.; Lou, Z. Separation of phenolics from peony flowers and their inhibitory activities and action mechanism on bacterial biofilm. Appl. Microbiol. Biotechnol. 2020, 104, 4321–4332. [Google Scholar] [CrossRef]
- Rajkumari, J.; Borkotoky, S.; Murali, A.; Suchiang, K.; Mohanty, S.K.; Busi, S. Attenuation of quorum sensing controlled virulence factors and biofilm formation in Pseudomonas aeruginosa by pentacyclic triterpenes, betulin and betulinic acid. Microb. Pathog. 2018, 118, 48–60. [Google Scholar] [CrossRef]
- Parasuraman, P.; Devadatha, B.; Sarma, V.V.; Ranganathan, S.; Ampasala, D.R.; Siddhardha, B. Anti-quorum sensing and antibiofilm activities of Blastobotrys parvus PPR3 against Pseudomonas aeruginosa PAO1. Microb. Pathog. 2020, 138, 103811. [Google Scholar] [CrossRef]
- Vestby, L.K.; Grønseth, T.; Simm, R.; Nesse, L.L. Bacterial biofilm and its role in the pathogenesis of disease. Antibiotics 2020, 9, 59. [Google Scholar] [CrossRef] [Green Version]
- Borges, A.; Sousa, P.; Gaspar, A.; Vilar, S.; Borges, F.; Simões, M. Furvina inhibits the 3-oxo-C12-HSL-based quorum sensing system of Pseudomonas aeruginosa and QS-dependent phenotypes. Biofouling 2017, 33, 156–168. [Google Scholar] [CrossRef]
- Ohta, T.; Fukumoto, A.; Iizaka, Y.; Kato, F.; Koyama, Y.; Anzai, Y. Quorum sensing inhibitors against Chromobacterium violaceum CV026 derived from an actinomycete metabolite library. Biol. Pharm. Bull. 2020, 43, 179–183. [Google Scholar] [CrossRef] [Green Version]
- Paguigan, N.D.; Rivera, J.C.; Stempin, J.J.; Augustinović, M.; Noras, A.I.; Raja, H.A.; Todd, D.A.; Triplett, K.D.; Day, C.; Figueroa, M.; et al. Prenylated diresorcinols inhibit bacterial quorum sensing. J. Nat. Prod. 2019, 82, 550–558. [Google Scholar] [CrossRef]
- Chang, A.; Sun, S.; Li, L.; Dai, X.; Li, H.; He, Q.; Zhu, H. Tyrosol from marine fungi, a novel quorum sensing inhibitor against Chromobacterium violaceum and Pseudomonas aeruginosa. Bioorg. Chem. 2019, 91, 103140. [Google Scholar] [CrossRef]
- Abraham, W.R. Going beyond the control of quorum-sensing to combat biofilm infections. Antibiotics 2016, 5, 3. [Google Scholar] [CrossRef]
- Newman, D.J.; Cragg, G.M. Natural products as sources of new drugs over the nearly four decades from 01/1981 to 09/2019. J. Nat. Prod. 2020, 83, 770–803. [Google Scholar] [CrossRef]
- Linthorne, J.S.; Chang, B.J.; Flematti, G.R.; Ghisalberti, E.L.; Sutton, D.C. A direct pre-screen for marine bacteria producing compounds inhibiting quorum sensing reveals diverse planktonic bacteria that are bioactive. Mar. Biotechnol. 2015, 17, 33–42. [Google Scholar] [CrossRef]
- Carroll, A.R.; Copp, B.R.; Davis, R.A.; Keyzers, R.A.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep. 2020, 37, 175–223. [Google Scholar] [CrossRef]
- Deniau, A.L.; Mosset, P.; Pédrono, F.; Mitre, R.; Le Bot, D.; Legrand, A.B. Multiple beneficial health effects of natural alkylglycerols from shark liver oil. Mar. Drugs 2010, 8, 2175–2184. [Google Scholar] [CrossRef] [Green Version]
- Magnusson, C.D.; Haraldsson, G.G. Ether lipids. Chem. Phys. Lipids 2011, 164, 315–340. [Google Scholar] [CrossRef]
- Pemha, R.; Pegnyemb, D.E.; Mosset, P. Synthesis of (Z)-(20R)-1-O-(20-methoxynonadec-100-enyl)-sn-glycerol, a new analog of bioactive ether lipids. Tetrahedron 2012, 68, 2973–2983. [Google Scholar] [CrossRef]
- Magnusson, C.D.; Gudmundsdottir, A.V.; Hansen, K.A.; Haraldsson, G.G. Synthesis of enantiopure reversed structured ether lipids of the 1-O-alkyl-sn-2,3-diacylglycerol type. Mar. Drugs 2015, 13, 173–201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dean, J.M.; Lodhi, I.J. Structural and functional roles of ether lipids. Protein Cell 2018, 9, 196–206. [Google Scholar] [CrossRef] [PubMed]
- Iannitti, T.; Palmieri, B. An update on the therapeutic role of alkylglycerols. Mar. Drugs 2010, 8, 2267–2300. [Google Scholar] [CrossRef] [Green Version]
- Watschinger, K.; Werner, E.R. Orphan enzymes in ether lipid metabolism. Biochimie 2013, 95, 59–65. [Google Scholar] [CrossRef] [Green Version]
- Sutter, M.; Silva, E.D.; Duguet, N.; Raoul, Y.; Métay, E.; Lemaire, M. Glycerol ether synthesis: A bench test for green chemistry concepts and technologies. Chem. Rev. 2015, 115, 8609–8651. [Google Scholar] [CrossRef]
- Tyrtyshnaia, A.; Manzhulo, I.; Kipryushina, Y.; Ermolenko, E. Neuroinflammation and adult hipocampal neurogenesis in neuropathic pain and alkyl glycerol ethers treatment in aged mice. Int. J. Mol. Med. 2019, 43, 2153–2163. [Google Scholar] [CrossRef] [Green Version]
- Yu, H.; Dilbaz, S.; Coßmann, J.; Hoang, A.C.; Diedrich, V.; Herwig, A.; Harauma, A.; Hoshi, Y.; Moriguchi, T.; Landgraf, K.; et al. Breast milk alkylglycerols sustain beige adipocytes through adipose tissue macrophages. J. Clin. Investig. 2019, 129, 2485–2499. [Google Scholar] [CrossRef] [Green Version]
- Haynes, M.P.; Buckley, H.R.; Higgins, M.L.; Pieringer, R.A. Synergism between the antifungal agents amphotericin B and alkylglycerol ethers. Antimicrob. Agents Chemother. 1994, 38, 1523–1529. [Google Scholar] [CrossRef] [Green Version]
- Ved, H.S.; Gustow, E.; Mahadevans, V.; Pieringer, R.A. Dodecylglycerol a new type of antibacterial agent which stimulates autolysin activity in Streptococcus faecium ATCC 9790. J. Biol. Chem. 1984, 259, 8115–8121. [Google Scholar] [CrossRef]
- Brissette, J.L.; Cabacungan, E.A.; Pieringer, R.A. Studies on the antibacterial activity of dodecylglycerol. Its limited metabolism and inhibition of glycerolipid and lipoteichoic acid biosynthesis in Streptococcus mutans BHT. J. Biol. Chem. 1986, 261, 6338–6345. [Google Scholar] [CrossRef]
- Ved, H.S.; Gustow, E.; Pieringer, R.A. Synergism between penicillin G and the antimicrobial ether lipid, rac-l-dodecylglycerol, acting below its critical micelle concentration. Lipids 1990, 25, 119–121. [Google Scholar] [CrossRef]
- Lampe, M.F.; Ballweber, L.M.; Isaacs, C.E.; Patton, D.L.; Stamm, W.E. Killing of Chlamydia trachomatis by novel antimicrobial lipids adapted from compounds in human breast milk. Antimicrob. Agents Chemother. 1998, 42, 1239–1244. [Google Scholar] [CrossRef] [Green Version]
- Qi, S.H.; Zhang, S.; Yang, L.H.; Qian, P.Y. Antifouling and antibacterial compounds from the gorgonians Subergorgia suberosa and Scirpearia gracilis. Nat. Prod. Rep. 2008, 22, 154–166. [Google Scholar] [CrossRef]
- Lin, Y.C.; Schlievert, P.M.; Anderson, M.J.; Fair, C.L.; Schaefers, M.M.; Muthyala, R.; Peterson, M.L. Glycerol monolaurate and dodecylglycerol effects on Staphylococcus aureus and toxic shock syndrome toxin-1 in vitro and in vivo. PLoS ONE 2009, 4, e7499. [Google Scholar] [CrossRef] [Green Version]
- Castellanos, L.H.; Mayorga, H.W.; Duque, C.B. Estudio de la composición química y actividad antifouling del extracto de la esponja marina Cliona delitrix. Vitae 2010, 17, 209–224. [Google Scholar]
- Mayorga, H.; Urrego, N.F.; Castellanos, L.; Duque, C. Cembradienes from the Caribbean Sea whip Eunicea sp. Tetrahedron Lett. 2011, 52, 2515–2518. [Google Scholar] [CrossRef]
- Martínez, Y.D.; Vanegas, G.L.; Reina, L.G.; Mayorga, H.W.; Arévalo-Ferro, C.; Ramos, F.R.; Duque, C.B.; Castellanos, L.H. Biofilm inhibition activity of compounds isolated from two Eunicea species collected at the Caribbean Sea. Rev. Bras. Farmacogn. 2015, 25, 605–611. [Google Scholar] [CrossRef] [Green Version]
- Othmani, A.; Bunet, R.; Bonnefont, J.L.; Briand, J.F.; Culioli, G. Settlement inhibition of marine biofilm bacteria and barnacle larvae by compounds isolated from the Mediterranean brown alga Taonia atomaria. J. Appl. Phycol. 2015, 28, 1975–1986. [Google Scholar] [CrossRef]
- Nascimento, T.S.; Monteiro, L.G.; Braga, E.F.; Batista, W.R.; Albert, A.L.; Chantre, L.G.; Machado, S.P.; Lopes, R.S.; Lopes, C.C. Synthesis of natural ether lipids and 1-O-hexadecylglycero-arylboronates via an epoxidering opening approach: Potential antifouling additives to marine paint coatings. Int. J. Adv. Res. Sci. Eng. Technol. 2018, 5, 326–332. [Google Scholar] [CrossRef] [Green Version]
- Barragán, C.A.; Silva, E.G.; Moreno, B.M.; Mayorga, H.W. Inhibition of quorum sensing by compounds from two Eunicea species and synthetic saturated alkylglycerols. Vitae 2018, 25, 92–103. [Google Scholar] [CrossRef] [Green Version]
- Fernández, D.M.; Contreras, L.J.; Moreno, B.M.; Silva, E.G.; Mayorga, H.W. Enantiomeric synthesis of natural alkylglycerols and their antibacterial and antibiofilm activities. Nat. Prod. Res. 2019, 1–7. [Google Scholar] [CrossRef]
- Qiu, M.N.; Wang, F.; Chen, S.Y.; Wang, P.C.; Fu, Y.H.; Liu, Y.Y.; Wang, X.; Wang, F.B.; Wang, C.; Yang, H.W.; et al. Novel 2, 8-bit derivatives of quinolines attenuate Pseudomonas aeruginosa virulence and biofilm formation. Bioorg. Med. Chem. Lett. 2019, 29, 749–754. [Google Scholar] [CrossRef]
- Kumar, P.; Lee, J.H.; Beyenal, H.; Lee, J. Fatty acids as antibiofilm and antivirulence agents. Trends Microbiol. 2020, 28, 753–768. [Google Scholar] [CrossRef]
- Rabin, N.; Zheng, Y.; Opoku, C.T.; Du, Y.; Bonsu, E.; Sintim, H.O. Agents that inhibit bacterial biofilm formation. Future Med. Chem. 2015, 7, 647–671. [Google Scholar] [CrossRef]
- Nunes, L.S.; Rigon, Z.K.; Macedo, A.J.; Silva, D.T. Plant natural products targeting bacterial virulence factors. Chem. Rev. 2016, 116, 9162–9236. [Google Scholar] [CrossRef]
- Townsley, L.; Shank, E.A. Natural-product antibiotics: Cues for modulating bacterial biofilm formation. Trends Microbiol. 2017, 25, 1016–1026. [Google Scholar] [CrossRef]
- Song, X.; Xia, Y.X.; He, Z.D.; Zhang, H.J. A review of natural products with anti-biofilm activity. Curr. Org. Chem. 2018, 22, 788–816. [Google Scholar] [CrossRef]
- Melander, R.J.; Basak, A.K.; Melander, C. Natural products as inspiration for the development of bacterial antibiofilm agents. Nat. Prod. Rep. 2020, 37, 1454–1477. [Google Scholar] [CrossRef]
- López, Y.; Soto, S.M. The usefulness of microalgae compounds for preventing biofilm infections. Antibiotics 2020, 9, 9. [Google Scholar] [CrossRef] [Green Version]
- Asfour, H.Z. Anti-quorum sensing natural compounds. J. Microsc. Ultrastruct. 2018, 6, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Deryabin, D.; Galadzhieva, A.; Kosyan, D.; Duskaev, G. Plant-derived inhibitors of AHL-mediated quorum sensing in bacteria: Modes of action. Int. J. Mol. Sci. 2019, 20, 5588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saurav, K.; Costantino, V.; Venturi, V.; Steindler, L. Quorum sensing inhibitors from the sea discovered using bacterial N-acyl-homoserine lactone-based biosensors. Mar. Drugs 2017, 15, 53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, J.; Li, X.; Hou, X.; Quan, C.; Chen, M. Widespread existence of quorum sensing inhibitors in marine bacteria: Potential drugs to combat pathogens with novel strategies. Mar. Drugs 2019, 17, 275. [Google Scholar] [CrossRef] [Green Version]
- Cadavid, E.; Echeverri, F. The search for natural inhibitors of biofilm formation and the activity of the autoinductor C6-AHL in Klebsiella pneumoniae ATCC 13884. Biomolecules 2019, 9, 49. [Google Scholar] [CrossRef] [Green Version]
- Dos Reis Ponce, A.; Martin, M.L.; De Araujo, E.F.; Mantovani, H.L.; Vanetti, M.C. AiiA quorum-sensing quenching controls proteolytic activity and biofilm formation by Enterobacter cloacae. Curr. Microbiol. 2012, 65, 758–763. [Google Scholar] [CrossRef]
- Qvortrup, K.; Hultqvist, L.D.; Nilsson, M.; Jakobsen, T.H.; Jansen, C.U.; Uhd, J.; Andersen, J.B.; Nielsen, T.E.; Givskov, M.; Nielsen, T.T. Small molecule anti-biofilm agents developed on the basis of mechanistic understanding of biofilm formation. Front. Chem. 2019, 7, 742. [Google Scholar] [CrossRef] [Green Version]
- Wasfi, R.; Hamed, S.M.; Amer, M.A.; Fahmy, L.I. Proteus mirabilis biofilm: Development and therapeutic strategies. Front. Cell. Infect. Microbiol. 2020, 10, 414. [Google Scholar] [CrossRef]
- Wang, H.; Chu, W.; Ye, C.; Gaeta, B.; Tao, H.; Wang, M.; Qiu, Z. Chlorogenic acid attenuates virulence factors and pathogenicity of Pseudomonas aeruginosa by regulating quorum sensing. Appl. Microbiol. Biotechnol. 2019, 103, 903–915. [Google Scholar] [CrossRef]
- Li, Y.; Pan, J.; Wu, D.; Tian, Y.; Zhang, J.; Fang, J. Regulation of Enterococcus faecalis biofilm formation and quorum sensing related virulence factors with ultra-low dose reactive species produced by plasma activated water. Plasma Chem. Plasma Process. 2019, 39, 35–49. [Google Scholar] [CrossRef]
- Monticelli, J.; Knezevich, A.; Luzzati, R.; Di Bella, S. Clinical management of non-faecium non-faecalis vancomycin-resistant enterococci infection. Focus on Enterococcus gallinarum and Enterococcus casseliflavus/flavescens. J. Infect. Chemother. 2018, 24, 237–246. [Google Scholar] [CrossRef] [Green Version]
- Lönn, J.S.; Naemi, A.O.; Benneche, T.; Petersen, F.C.; Scheie, A.A. Thiophenones inhibit Staphylococcus epidermidis biofilm formation at nontoxic concentrations. FEMS Immunol. Med. Microbiol. 2012, 65, 326–334. [Google Scholar] [CrossRef] [Green Version]
- Nakagawa, S.; Hillebrand, G.G.; Nunez, G. Rosmarinus officinalis L. (Rosemary) extracts containing carnosic acid and carnosol are potent quorum sensing inhibitors of Staphylococcus aureus Virulence. Antibiotics 2020, 9, 149. [Google Scholar] [CrossRef] [Green Version]
- Stepanović, S.; Vuković, D.; Dakić, I.; Savić, B.; Svabić-Vlahovic, M. A modified microtiter-plate test for quantification of staphylococcal biofilm formation. J. Microbiol. Methods 2000, 40, 175–179. [Google Scholar] [CrossRef]
- Taylor, J.R. An Introduction to Error Analysis. The Study of Uncertainties in Physical Measurements, 2nd ed.; University Science Books: Sausalito, CA, USA, 1997; pp. 93–170. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Strains | Alkylglycerols a | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| CPX b | 2 | 3 | 4 | 5 | 6 | 7 | 9 | 10 | 11 | |
| K. pneumoniae 792 | 0.09 | >1224 | >1076 | >960 | >867 | >790 | >795 | >730 | >734 | >738 |
| E. cloacae 250 | 48.2 | >1224 | >1076 | >960 | >867 | 395 | >795 | >730 | >734 | >738 |
| E. coli 667 | 0.09 | >1224 | >1076 | >960 | >867 | >790 | >795 | >730 | >734 | >738 |
| P. aeruginosa 740 | 1.51 | >1224 | >1076 | >960 | >867 | >790 | >795 | >730 | >734 | >738 |
| P. mirabilis 26 | 0.09 | >1224 | 1076 | >960 | >867 | >790 | >795 | >730 | >734 | >738 |
| C. violaceum ATCC 12472 | 0.18 | >1224 | 269 | >960 | >867 | >790 | >795 | 730 | >734 | >738 |
| E. faecalis 12 | 0.51 | >1224 | 538 | 119 | 433 | >790 | 199 | 365 | 734 | >738 |
| E. gallinarum 662 | 48.2 | >1224 | >1076 | 119 | >867 | >790 | >795 | >730 | >734 | >738 |
| S. epidermidis ATCC 12228 | 0.39 | >1224 | 538 | 119 | 108 | >790 | 795 | >730 | 734 | >738 |
| S. aureus ATCC 6538 | 0.18 | 612 | 67 | 30 | 27 | >790 | 99 | >730 | 367 | 369 |
| Values. µM a | Alkylglycerols | ||||||||||||||
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | |
| 709 | 611 | 135 | 120 | 109 | 197 | 795 | 363 | 365 | 92 | 369 | 335 | 337 | 312 | 20 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chaverra Daza, K.E.; Silva Gómez, E.; Moreno Murillo, B.D.; Mayorga Wandurraga, H. Natural and Enantiopure Alkylglycerols as Antibiofilms Against Clinical Bacterial Isolates and Quorum Sensing Inhibitors of Chromobacterium violaceum ATCC 12472. Antibiotics 2021, 10, 430. https://doi.org/10.3390/antibiotics10040430
Chaverra Daza KE, Silva Gómez E, Moreno Murillo BD, Mayorga Wandurraga H. Natural and Enantiopure Alkylglycerols as Antibiofilms Against Clinical Bacterial Isolates and Quorum Sensing Inhibitors of Chromobacterium violaceum ATCC 12472. Antibiotics. 2021; 10(4):430. https://doi.org/10.3390/antibiotics10040430
Chicago/Turabian StyleChaverra Daza, Klauss E., Edelberto Silva Gómez, Bárbara D. Moreno Murillo, and Humberto Mayorga Wandurraga. 2021. "Natural and Enantiopure Alkylglycerols as Antibiofilms Against Clinical Bacterial Isolates and Quorum Sensing Inhibitors of Chromobacterium violaceum ATCC 12472" Antibiotics 10, no. 4: 430. https://doi.org/10.3390/antibiotics10040430
APA StyleChaverra Daza, K. E., Silva Gómez, E., Moreno Murillo, B. D., & Mayorga Wandurraga, H. (2021). Natural and Enantiopure Alkylglycerols as Antibiofilms Against Clinical Bacterial Isolates and Quorum Sensing Inhibitors of Chromobacterium violaceum ATCC 12472. Antibiotics, 10(4), 430. https://doi.org/10.3390/antibiotics10040430