
S-PRG Filler Eluate Induces Oxidative Stress in Oral Microorganism: Suppression of Growth and Pathogenicity, and Possible Clinical Application


Abstract
:1. Introduction
2. Results
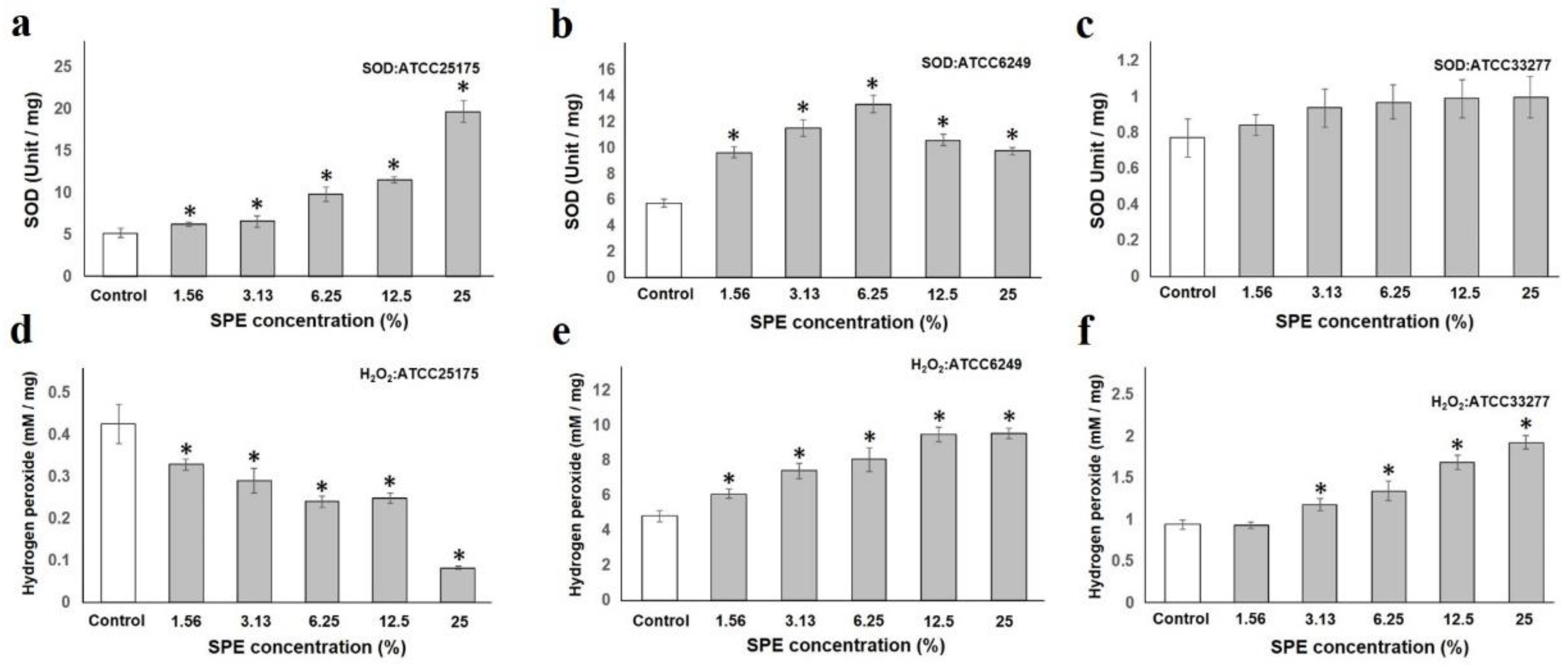
2.1. SPE Induces Oxidative Stress
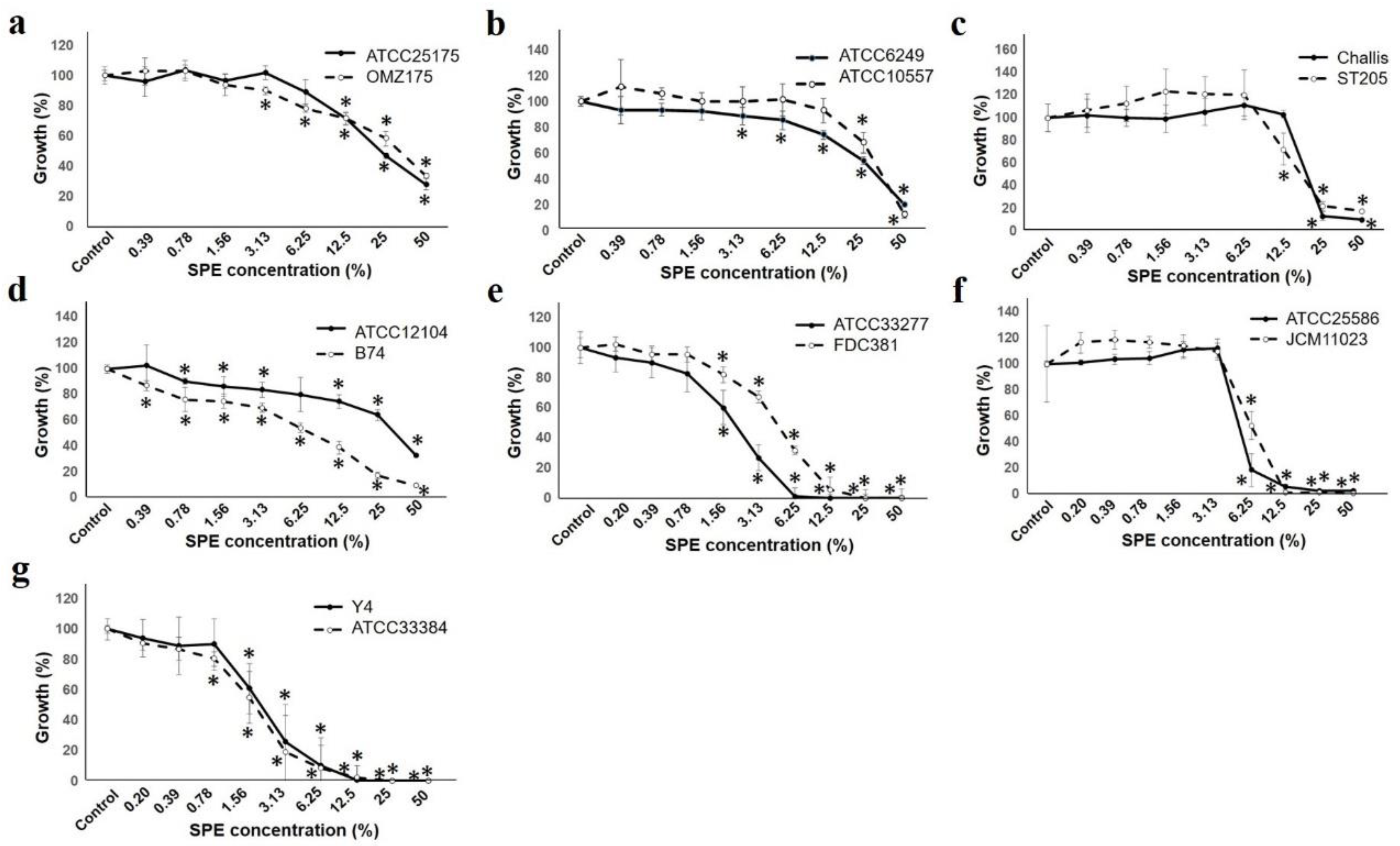
2.2. Bacterial Growth Evaluation
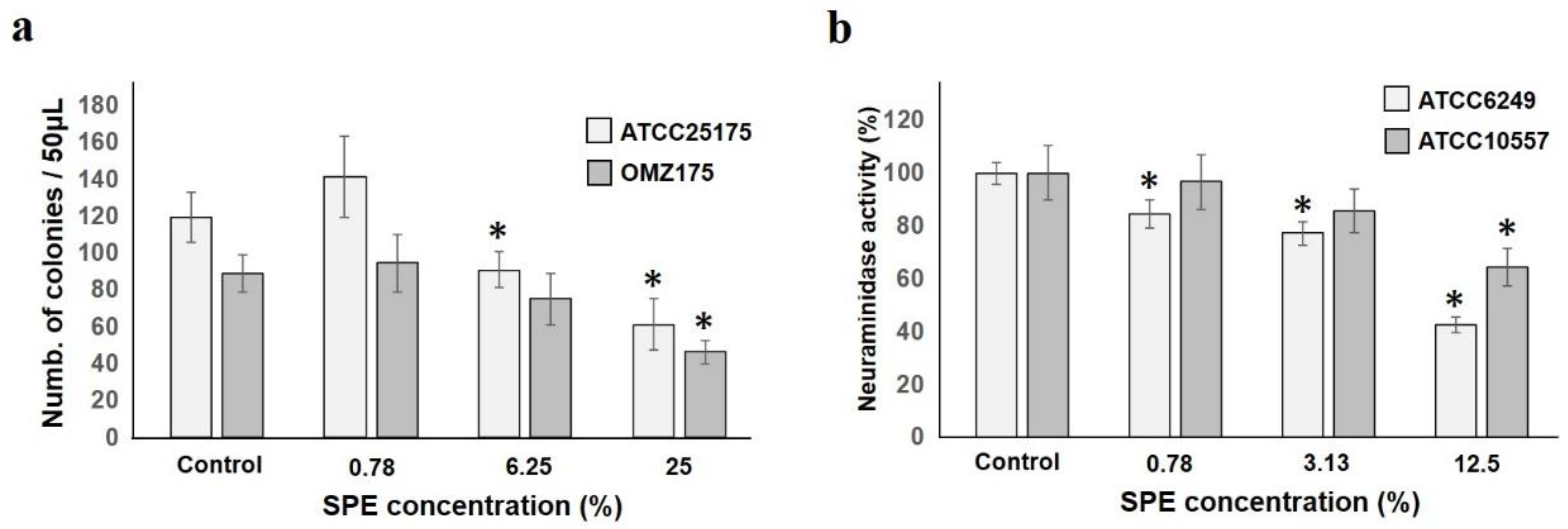
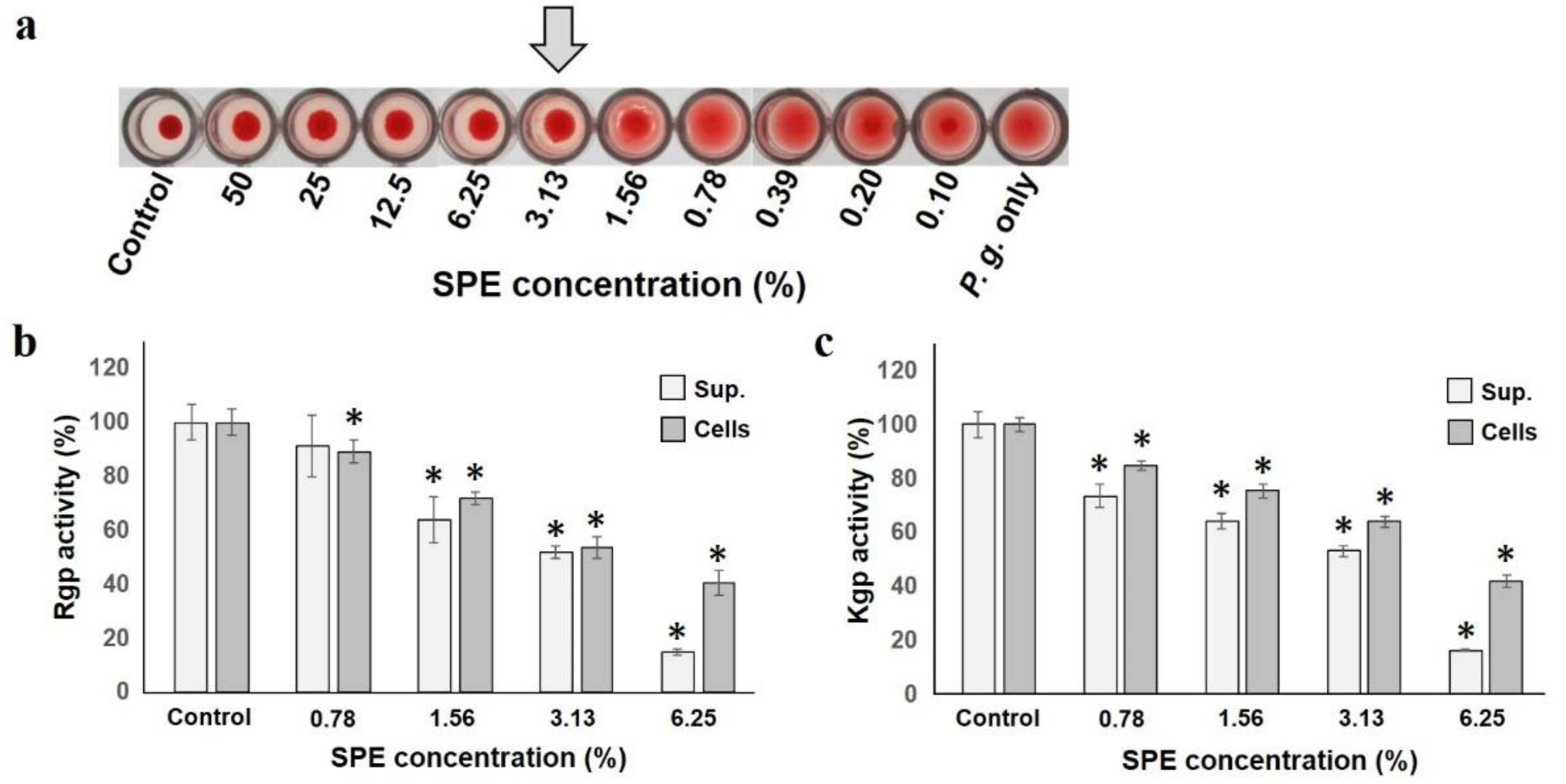
2.3. Effects on Adhesion and Pathogenic Factors of Test Strains
3. Discussion
4. Materials and Methods
4.1. Bacterial Cells and Materials
4.2. Superoxide Dismutase (SOD) and Hydrogen Peroxide Measurement
4.3. Bacterial Growth Evaluation
4.4. Effects on Adhesion, Hemagglutination, and Pathogenic Factors of Test Strains
4.5. Statistical Analyses
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Aas, J.A.; Paster, B.J.; Stokes, L.N.; Olsen, I.; Dewhirst, F.E. Defining the normal bacteria flora of the oral cavity. J. Clin. Microbiol. 2005, 43, 5721–5732. [Google Scholar] [CrossRef] [Green Version]
- Selwitz, R.H.; Ismail, A.I.; Pitts, N.B. Dental caries. Lancet 2007, 369, 51–59. [Google Scholar] [CrossRef]
- William, R.C. Periodontal disease. N. Engl. J. Med. 1990, 32, 373–382. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Kolltveit, R.M.; Traonstad, L.; Olsen, I. Systemic diseases caused by oral infection. Clin. Microbiol. Rev. 2000, 13, 547–558. [Google Scholar] [CrossRef] [PubMed]
- Seymour, G.J.; Ford, P.J.; Cullinan, M.P.; Leishman, S.; Yamazaki, K. Relationship between periodontal infections and systemic disease. Clin. Microbiol. Infect. 2007, 13, 3–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Slots, J.; Kamma, J.J. General health risk of periodontal disease. Int. Dent. J. 2001, 51, 417–427. [Google Scholar] [CrossRef] [PubMed]
- D’Aiuto, F.; Gable, D.; Syed, D.; Allen, Y.; Wanyonyi, K.L.; White, S.; Gallagher, J.E. Evidence summary: The relationship between oral diseases and diabetes. Br. Dent. J. 2017, 222, 944–948. [Google Scholar] [CrossRef] [Green Version]
- Dietrich, T.; Webb, I.; Stenhouse, L.; Pattni, A.; Ready, D.; Wanyonyi, K.L.; White, S.; Gallagher, J.E. Evidence summary: The relationship between oral and cardiovascular disease. Br. Dent. J. 2017, 222, 381–385. [Google Scholar] [CrossRef]
- Gomes-Filho, I.S.; Passos, J.S.; de Cruz, S.S. Respiratory disease and the role of oral bacteria. J. Oral Microbiol. 2010, 2, 5811. [Google Scholar] [CrossRef]
- Stein, P.S.; Scheff, S.; Dawson, D.R., III. Alzheimer’s disease and periodontal disease: Mechanisms underlying a potential bi-directional relationship. Grand Round Oral Syst. Med. 2006, 1, 14–24. [Google Scholar]
- Kolenbrander, P.E.; London, J. Adhere today, here tomorrow, oral bacterial adherence. J. Bacteriol. 1993, 175, 3247–3252. [Google Scholar] [CrossRef] [Green Version]
- Coulthwaite, L.; Verran, J. Potentioal pathogenic aspects of denture plaque. Br. J. Biomed. Sci. 2007, 64, 180–189. [Google Scholar] [CrossRef]
- Larsen, T.; Fiehn, N.E. Dental biofilm infections—An update. Apmis 2017, 123, 376–384. [Google Scholar] [CrossRef]
- Murakami, S.; Mealey, B.L.; Mariotti, A.; Chapple, I.L.C. Dental plaque–induced gingival conditions. J. Clin. Periodontol. 2018, 45, S17–S27. [Google Scholar] [CrossRef] [PubMed]
- Bin-Shuwaish, M.S. Effects and effectiveness of cavity disinfectants in operative dentistry: A literature review. J. Contemp. Dent. Pract. 2016, 17, 867–879. [Google Scholar] [CrossRef] [PubMed]
- Van Buul, L.W.; van ser Steen, J.T.; Veenhuizen, R.B.; Achterberg, W.P.; Schellevis, F.G.; Pharm, T.G.M.E.; van Benthem, B.H.B.; Pharm, S.N.; Hertogh, C.M.P.M. Antibiotic use and resistance in long term care facilities. J. Am. Med. Dir. Assoc. 2012, 13, 568.e1–568.e13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Othman, L.; Sleiman, A.; Abdel-Massih, R.M. Antimicrobial activity of polyphenols and alkaloids in middle eastern plants. Front. Microbiol. 2019, 10, 911. [Google Scholar] [CrossRef] [PubMed]
- Panpaliya, N.P.; Dahake, P.T.; Kale, Y.J.; Dadpe, M.V.; Kendre, S.B.; Siddiqi, A.G.; Maggavi, U.R. In vitro evaluation of antimicrobial property of silver nanoparticles and chlorhexidine against five different oral pathogenic bacteria. Saudi Dent. J. 2019, 31, 76–83. [Google Scholar] [CrossRef] [PubMed]
- Sarb, L.G.; Bahrin, L.G.; Babii, C.; Stefan, M.; Birsa, M.L. Synthetic flavonoids with antimicrobial activity: A review. J. Appl. Microbiol. 2019, 127, 1282–1290. [Google Scholar] [CrossRef] [Green Version]
- Tamura, M.; Saito, H.; Kikuchi, K.; Ishigami, T.; Toyama, Y.; Takami, M.; Ochiai, K. Antimicrobial activity of gel-entrapped catechins toward oral microorganisms. Biol. Pharm. Bull. 2011, 34, 638–643. [Google Scholar] [CrossRef] [Green Version]
- Fujimoto, Y.; Iwasa, M.; Murayama, R.; Miyazaki, M.; Nagafuji, A.; Nakatsuka, T. Detection of ions released from S-PRG fillers and their modulation effect. Dent. Mater. J. 2010, 29, 392–397. [Google Scholar] [CrossRef] [Green Version]
- Hotta, M.; Morikawa, T.; Tamura, D.; Kusakabem, S. Adherence of Streptococcus sanguinis and Streptococcus mutans to saliva-coated S-PRG resin blocks. Dent. Mater. J. 2014, 33, 261–267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaga, M.; Kakuda, S.; Ida, Y.; Toshima, H.; Hashimoto, M.; Endo, K.; Sano, H. Inhibition of enamel demineralization by buffering effect of S-PRG filler-containing dental sealant. Eur. J. Oral Sci. 2014, 122, 78–83. [Google Scholar] [CrossRef] [PubMed]
- Kotaku, M.; Murayama, R.; Shimamura, Y.; Takahashi, F.; Suzuki, T.; Kurokawa, H.; Miyazaki, M. Evaluation of the effects of fluoride-releasing varnish on dentin demineralization using optical coherence tomography. Dent. Mater. J. 2014, 33, 648–655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakamura, N.; Yamada, A.; Iwamoto, T.; Arakaki, M.; Tanaka, K.; Aizawa, S.; Nonaka, K.; Fukumoto, S. Two-year clinical evaluation of flowable composite resin containing pre-reacted glass-ionomer. Pediatr. Dent. J. 2009, 19, 89–97. [Google Scholar] [CrossRef] [Green Version]
- Shimizubata, M.; Inokoshi, M.; Wada, T.; Takahashi, R.; Uo, M.; Minakuchi, S. Basic properties of novel S-PRG filler-containing cement. Dent. Mater. J. 2020, 39, 963–969. [Google Scholar] [CrossRef]
- Takahashi, Y.; Okamoto, M.; Komichi, S.; Imazato, S.; Nakatsuka, T.; Sakamoto, S.; Kimoto, K.; Hayashi, M. Application of a direct pulp capping cement containing S-PRG filler. Clin. Oral Investig. 2019, 23, 1723–1731. [Google Scholar] [CrossRef]
- Storz, G.; Imlayt, J.A. Oxidative stress. Curr. Opin. Microbiol. 1999, 2, 188–194. [Google Scholar] [CrossRef]
- Da Silva, D.A.; Patterson, M.J.; Smith, D.A.; Maccallum, D.M.; Erwig, L.P.; Morgan, B.A.; Quinn, J. Thioredoxin regulates multiple hydrogen peroxide- induced signaling pathways in Candida albicans. Mol. Cell Biol. 2010, 30, 4550–4563. [Google Scholar] [CrossRef] [Green Version]
- Groeger, G.; Quiney, C.; Fotter, T.G. Hydrogen peroxide as a cell-survival signaling molecule. Antioxid. Redox Signal. 2009, 11, 2655–2671. [Google Scholar] [CrossRef]
- Pensinger, D.A.; Schaenzer, A.J.; Sauer, J.-D. Do shoot the messenger: PASTA kinases as virulence determinants and antibiotic targets. Trends Microbiol. 2018, 26, 56–69. [Google Scholar] [CrossRef] [PubMed]
- Cheng, X.; Redanz, S.; Treerat, P.; Qin, H.; Choi, D.; Zhou, X.; Xu, X.; Merritt, J.; Kreth, J. Magnesium-Dependent Promotion of H2O2 Production Increases Ecological Competitiveness of Oral Commensal Streptococci. J. Dent. Res. 2020, 90, 847–854. [Google Scholar] [CrossRef]
- Tamura, M.; Ochiai, K. Exploring the possible application of catechin (gel) for oral care of the elderly and disabled individuals. Jpn. Dent. Sci. Rev. 2012, 48, 126–134. [Google Scholar] [CrossRef] [Green Version]
- Fortuin, S.; Nel, A.J.M.; Blackburn, J.M.; Soares, N.C. Comparison between the proteome of Escherichia coli single colony and during liquid culture. J. Proteom. 2020, 228, 103929. [Google Scholar] [CrossRef]
- Tabaraie, B.; Ghasemian, E.; Tabaraie, T.; Parvizi, E.; Rezazarandi, M. Comparative evalution of cephalosporin-C production in solid state fermentation and sunmerged liquid culture. J. Microbiol. Biotech. Food Sci. 2012, 2, 83–94. [Google Scholar]
- Singh, A.K.; Woodiga, S.A.; Grau, M.A.; King, S.J. Streptococcus oralis neuraminidase modulates adherence to multiple carbohydrates on platelets. Infect. Immun. 2017, 85, e00774-16. [Google Scholar] [CrossRef] [Green Version]
- Holt, S.C.; Kesavalu, L.; Walker, S.; Genco, C.A. Virulence factors of Porphyromonas gingivalis. Periodontal 2000 1999, 20, 168–238. [Google Scholar] [CrossRef]
- Aroma, A.; Sairam, R.K.; Srivastava, G.C. Oxidative stress and antioxidative system in plants. Curr. Sci. 2002, 82, 1227–1238. [Google Scholar]
- Puppel, K.; Kapusta, A.; Kuczyriska, B. The etiology of oxidative stress in the various species of animals, a review. J. Sci. Food Agric. 2014, 95, 2179–2184. [Google Scholar] [CrossRef] [PubMed]
- Sies, H.; Berndt, C.; Jones, D.P. Oxidative stress. Ann. Rev. Biochem. 2017, 86, 715–748. [Google Scholar] [CrossRef]
- Sigler, K.; Chaloupka, J.; Brozmanova, J.; Stadler, N.; Höfer, M. Oxidative stress in microorganisms-I. Folia Microbiol. 1999, 44, 587–624. [Google Scholar] [CrossRef] [PubMed]
- Chlubek, D. Fluoride and oxidative stress. Fluoride 2003, 36, 217–228. [Google Scholar]
- Jebahi, S.; Oudadesse, H.; el Feki, H.; Rebai, T.; Keskes, H.; Pellen, P.; el Feki, A. Antioxidative/oxidative effects of strontium-doped bioactive glass as bone graft. In vivo assays in ovariectomised rats. J. Appl. Biomed. 2012, 10, 195–209. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.-H.; Khan, A.L.; Waqas, M.; Lee, I.-J. Silicon regulates antioxidant activities of crop plants under abiotic-induced oxidative stress: A review. Front. Plant Sci. 2017, 8, 510. [Google Scholar] [CrossRef] [Green Version]
- Lu, Y.; Luo, Q.; Cui, H.; Deng, H.; Kuang, P.; Liu, H.; Fang, J.; Zuo, Z.; Deng, J.; Li, Y.; et al. Sodium fluoride causes oxidative stress and apoptosis in the mouse liver. Aging 2017, 9, 1623–1639. [Google Scholar] [CrossRef] [Green Version]
- Praticò, D.; Uryu, K.; Sung, S.; Tang, S.; Trojanowski, J.Q.; Lee, V.M.-Y. Aluminum modulates brain amyloidosis through oxidative stress in APP transgenic mice. FASEB J. 2002, 16, 1138–1140. [Google Scholar] [CrossRef]
- Song, B.; Hao, X.; Wang, X.; Yang, S.; Dong, Y.; Ding, Y.; Wang, Q.; Wang, X.; Zhou, J. Boron stress inhibits beet (Beta vulgaris L.) growth through influencing endogenous hormones and oxidative stress response. Soil Sci. Plant Nutr. 2019, 65, 346–352. [Google Scholar] [CrossRef]
- Yalin, S.; Sagir, O.; Comelekoglu, U.; Berköz, M.; Eroglu, P. Strontium ranelate treatment improves oxidative damage in osteoporotic rat model. Pharm. Rep. 2012, 64, 396–402. [Google Scholar] [CrossRef]
- Tamura, M.; Cueno, M.E.; Abe, K.; Kamio, N.; Imai, K.; Ochiai, K. Ions released from a S-PRG filler induces oxidative stress in Candida albicans inhibiting its growth and pathogenicity. Cell Stress Chaperones 2018, 23, 1137–1143. [Google Scholar] [CrossRef]
- Miki, S.; Kitagawa, H.; Kitagawa, R.; Kiba, W.; Hayashi, M.; Imazato, S. Antibacterial activity of resin composites containing surface pre-reacted glass-ionomer (S-PRG) filler. Dent. Mater. 2016, 32, 1095–1102. [Google Scholar] [CrossRef]
- Kitada, K.; Ino, T. Effect of saliva viscosity on the co-aggregation between oral streptococci and Actinomyces naeslundii. Gerodontolgy 2012, 29, e981–e987. [Google Scholar] [CrossRef]
- Liu, P.-F.; Shi, W.; Zhu, W.; Smith, J.W.; Hsieh, S.-L.; Gallo, R.L.; Huang, C.-M. Vaccination targeting surface FomA of Fusobacterium nucleatum against bacterial co-aggregation: Implication for treatment of periodontal infection and halitosis. Vaccine 2010, 28, 3496–3505. [Google Scholar] [CrossRef] [Green Version]
- Banas, J.A. Virulence properties of Streptococcus mutans. Front. Biosci. 2004, 9, 1267–1277. [Google Scholar] [CrossRef]
- Hotta, M.; Tamura, D.; Kotake, H.; Kusakabe, S.; Gen, T.; Oike, K. Adherence of Streptococcus mutans and adsorption of salivary protein to resin composites containing S-PRG fillers. Open J. Stomatol. 2017, 7, 158–168. [Google Scholar] [CrossRef] [Green Version]
- Nomura, R.; Morita, Y.; Matayoshi, S.; Nakano, N. Inhibitory effect of surface pre-reacted glass-ionomer (S-PRG) eluate against adhesion and colonization by Streptococcus mutans. Sci. Rep. 2018, 8, 5056. [Google Scholar] [CrossRef]
- Kitagawa, H.; Kimi-Oka, S.; Mayanagi, G.; Abiko, Y.; Takahashi, N.; Imazato, S. Inhibitory effect of resin composite containing S-PRG filler on Streptococcus mutans glucose metabolism. J. Dent. 2018, 70, 92–96. [Google Scholar] [CrossRef]
- Colman, P.M. Influenza virus neuraminidase: Structure, antibodies, and inhibitors. Protein Sci. 1994, 3, 1687–1696. [Google Scholar] [CrossRef] [Green Version]
- Kamio, N.; Imai, K.; Shimizu, K.; Cueno, M.E.; Tamura, M.; Saito, Y.; Ochiai, K. Neuraminidase-producing oral mitis group streptococci potentially contribute to influenza viral infection and reduction in antiviral efficacy of zanamivir. Cell Mol. Life Sci. 2015, 72, 357–366. [Google Scholar] [CrossRef]
- Nishikawa, T.; Shimizu, K.; Tanaka, T.; Kuroda, K.; Takayama, T.; Yamamoto, T.; Hanada, N.; Hamada, Y. Bacterial neuraminidase rescues influenza virus replication from inhibition by a neuraminidase inhibitor. PLoS ONE 2012, 7, e45371. [Google Scholar] [CrossRef] [Green Version]
- Seth, S.; Batra, J.; Srinivasan, S. COVID-19: Targeting proteases in viral invasion and host immune response. Front. Mol. Biosci. 2020, 7, 215. [Google Scholar] [CrossRef]
- Sun, X.; Tse, L.V.; Ferguson, A.D.; Whittaker, G.R. Modifications to the hemagglutinin cleavage site control the virulence of a neurotropic H1N1 Influenza virus. J. Virol. 2010, 84, 8683–8690. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kolenbrander, P.E. Coaggregationsamong oral bacteria. Methods Enzymol. 1995, 253, 385–387. [Google Scholar]
- Eick, S.; Reissmann, A.; Rodel, J.; Schmidt, K.H.; Pfister, W. Porphyromonas gingivalis survives within KB cells and modulates inflammatory response. Oral Microbiol. Immunol. 2006, 21, 231–237. [Google Scholar] [CrossRef]
- Lewis, J.P.; Dawson, J.A.; Hannis, J.C.; Muddiman, D.; Macrina, F.L. Hemoglobinase activity of lysine gingipain protease (Kgp) of Porphyromonas gingivalis W83. J. Bacteriol. 1999, 181, 4905–4913. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Turbidity Method | Colony Forming Unit Method | |
|---|---|---|
| Gram Positives | ||
| S. mutans ATCC25175 | 22.9 | 9.1 |
| S. oralis ATCC6249 | 30.9 | 20.8 |
| S. gondorii Charlis | 21.0 | 14.5 |
| A. naeslundii ATCC12104 | 30.6 | 25.5 |
| Gram Negatives | ||
| P. ginigivalis ATCC33277 | 2.1 | 3.1 |
| F. nuckeatum ATCC25586 | 5.0 | 3.8 |
| A. actinomycetemcomitans Y4 | 2.7 | 4.5 |
| Minimum inhibitory concentration 50 | SPE concentration (%) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kono, Y.; Tamura, M.; Cueno, M.E.; Tonogi, M.; Imai, K. S-PRG Filler Eluate Induces Oxidative Stress in Oral Microorganism: Suppression of Growth and Pathogenicity, and Possible Clinical Application. Antibiotics 2021, 10, 816. https://doi.org/10.3390/antibiotics10070816
Kono Y, Tamura M, Cueno ME, Tonogi M, Imai K. S-PRG Filler Eluate Induces Oxidative Stress in Oral Microorganism: Suppression of Growth and Pathogenicity, and Possible Clinical Application. Antibiotics. 2021; 10(7):816. https://doi.org/10.3390/antibiotics10070816
Chicago/Turabian StyleKono, Yu, Muneaki Tamura, Marni E. Cueno, Morio Tonogi, and Kenichi Imai. 2021. "S-PRG Filler Eluate Induces Oxidative Stress in Oral Microorganism: Suppression of Growth and Pathogenicity, and Possible Clinical Application" Antibiotics 10, no. 7: 816. https://doi.org/10.3390/antibiotics10070816
APA StyleKono, Y., Tamura, M., Cueno, M. E., Tonogi, M., & Imai, K. (2021). S-PRG Filler Eluate Induces Oxidative Stress in Oral Microorganism: Suppression of Growth and Pathogenicity, and Possible Clinical Application. Antibiotics, 10(7), 816. https://doi.org/10.3390/antibiotics10070816