
Determination of a Tentative Epidemiological Cut-Off Value (ECOFF) for Dalbavancin and Enterococcus faecium

 , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. Molecular Typing of the Strain Collection
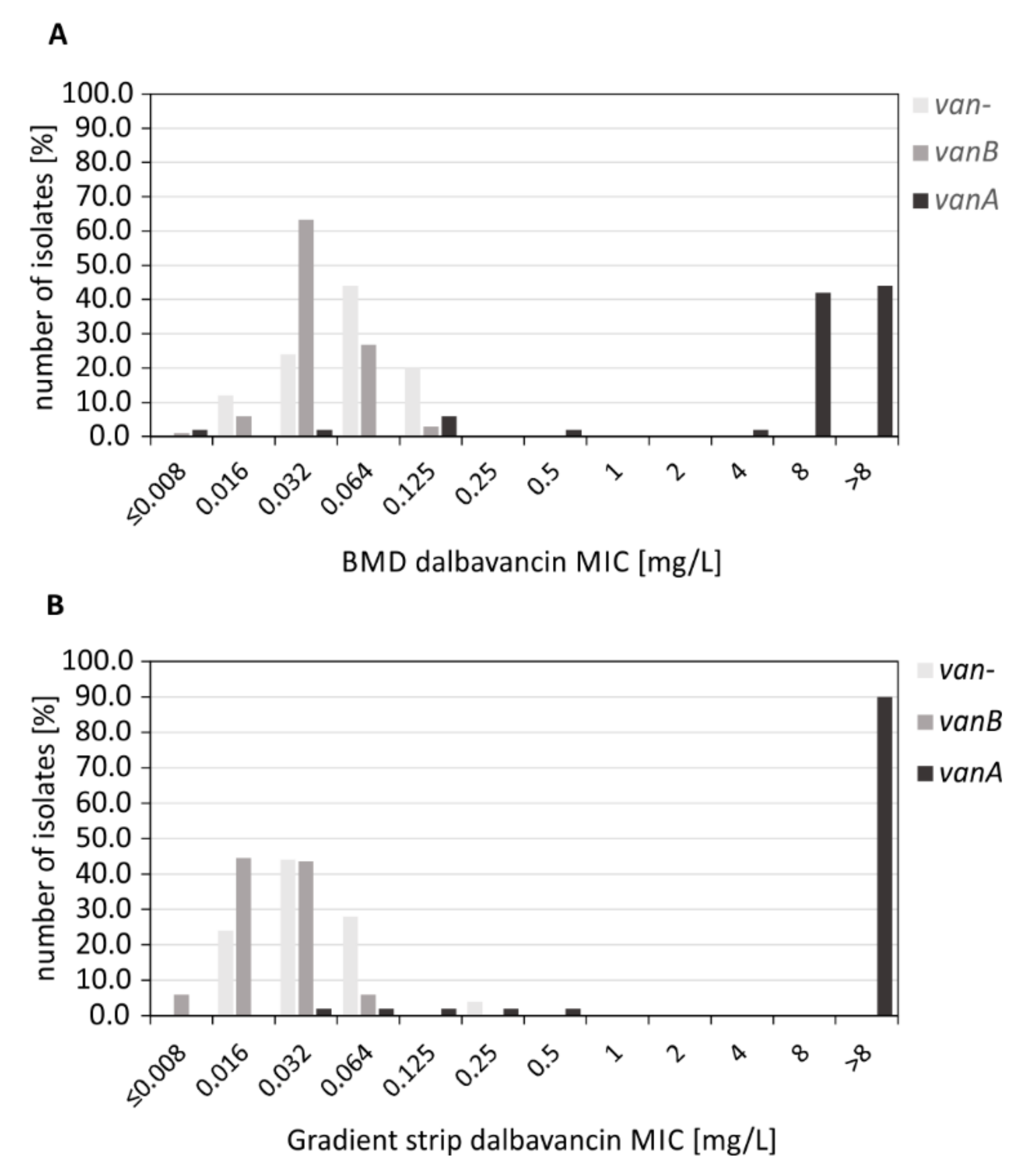
2.2. Determination of Dalbavancin MICs by BMD Testing
2.3. Determination of Dalbavancin MICs by MIC Gradient Strip Testing
2.4. Comparing Results of Broth Microdilution and MIC Test Strips
3. Discussion
4. Materials and Methods
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kramer, T.S.; Remschmidt, C.; Werner, S.; Behnke, M.; Schwab, F.; Werner, G.; Gastmeier, P.; Leistner, R. The importance of adjusting for enterococcus species when assessing the burden of vancomycin resistance: A cohort study including over 1000 cases of enterococcal bloodstream infections. Antimicrob. Resist. Infect. Control 2018, 7, 133. [Google Scholar] [CrossRef]
- European Centre for Disease Prevention and Control (ECDC). Surveillance of Antimicrobial Resistance in Europe 2018; ECDC: Stockholm, Sweden, 2019. [Google Scholar]
- Ayobami, O.; Willrich, N.; Reuss, A.; Eckmanns, T.; Markwart, R. The ongoing challenge of vancomycin-resistant Enterococcus faecium and Enterococcus faecalis in Europe: An epidemiological analysis of bloodstream infections. Emerg. Microbes Infect. 2020, 9, 1180–1193. [Google Scholar] [CrossRef] [PubMed]
- Buetti, N.; Wassilew, N.; Rion, V.; Senn, L.; Gardiol, C.; Widmer, A.; Marschall, J. Emergence of vancomycin-resistant enterococci in Switzerland: A nation-wide survey. Antimicrob. Resist. Infect. Control 2019, 8, 16. [Google Scholar] [CrossRef] [Green Version]
- Hammerum, A.M.; Justesen, U.S.; Pinholt, M.; Roer, L.; Kaya, H.; Worning, P.; Nygaard, S.; Kemp, M.; Clausen, M.E.; Nielsen, K.L.; et al. Surveillance of vancomycin-resistant enterococci reveals shift in dominating clones and national spread of a vancomycin-variable vanA Enterococcus faecium ST1421-CT1134 clone, Denmark, 2015 to March 2019. Eurosurveillance 2019, 24, 1900503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoffmann, M.; Yao, K.; Allard, M.; Sanchez, M.; Andersen, L.P.; Hasman, H.; Hammerum, A.M. Complete Genome Sequence of a Vancomycin-Resistant Sequence Type 203 Enterococcus faecium Strain with vanA Belonging to Complex Type 859. Microbiol. Resour. Announc. 2018, 7, e00815-18. [Google Scholar] [CrossRef] [Green Version]
- Eisenberger, D.; Tuschak, C.; Werner, M.; Bogdan, C.; Bollinger, T.; Hossain, H.; Friedrich, P.; Hussein, Z.; Pohlmann, C.; Wurstl, B.; et al. Whole-genome analysis of vancomycin-resistant Enterococcus faecium causing nosocomial outbreaks suggests the occurrence of few endemic clonal lineages in Bavaria, Germany. J. Antimicrob. Chemother. 2020, 75, 1398–1404. [Google Scholar] [CrossRef]
- Falgenhauer, L.; Fritzenwanker, M.; Imirzalioglu, C.; Steul, K.; Scherer, M.; Rhine-Main, V.s.g.; Heudorf, U.; Chakraborty, T. Near-ubiquitous presence of a vancomycin-resistant Enterococcus faecium ST117/CT71/vanB -clone in the Rhine-Main metropolitan area of Germany. Antimicrob Resist. Infect. Control. 2019, 8, 128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sadowy, E.; Gawryszewska, I.; Kuch, A.; Żabicka, D.; Hryniewicz, W. The changing epidemiology of VanB Enterococcus faecium in Poland. Eur. J. Clin. Microbiol. Infect. Dis. 2018, 37, 927–936. [Google Scholar] [CrossRef] [Green Version]
- Mischnik, A.; Werner, G.; Bender, J.; Mutters, N.T. Enterococci With Special Resistance Patterns—Epidemiology, Hygiene and Therapy. Deutsche Medizinische Wochenschrift 2019, 144, 553–560. [Google Scholar] [CrossRef]
- Bender, J.K.; Cattoir, V.; Hegstad, K.; Sadowy, E.; Coque, T.M.; Westh, H.; Hammerum, A.M.; Schaffer, K.; Burns, K.; Murchan, S.; et al. Update on prevalence and mechanisms of resistance to linezolid, tigecycline and daptomycin in enterococci in Europe: Towards a common nomenclature. Drug Resist. Updates 2018, 40, 25–39. [Google Scholar] [CrossRef] [PubMed]
- Klare, I.; Bender, J.K.; Markwart, R.; Reuss, A.; Abu SIn, M.; Eckmanns, T.; Werner, G. Properties, frequencies and distribution of vancomycin-resistant enterococci in Germany—Update 2017/2018. Epidemiol. Bull. 2019, 37, 365–372. (In German) [Google Scholar] [CrossRef]
- Klare, I.; Fleige, C.; Geringer, U.; Thurmer, A.; Bender, J.; Mutters, N.T.; Mischnik, A.; Werner, G. Increased frequency of linezolid resistance among clinical Enterococcus faecium isolates from German hospital patients. J. Glob. Antimicrob. Resist. 2015, 3, 128–131. [Google Scholar] [CrossRef] [PubMed]
- Bender, J.K.; Klare, I.; Fleige, C.; Werner, G. A Nosocomial Cluster of Tigecycline- and Vancomycin-Resistant Enterococcus faecium Isolates and the Impact of rpsJ and tet(M) Mutations on Tigecycline Resistance. Microb. Drug Resist. 2020, 26, 576–582. [Google Scholar] [CrossRef]
- Fiedler, S.; Bender, J.K.; Klare, I.; Halbedel, S.; Grohmann, E.; Szewzyk, U.; Werner, G. Tigecycline resistance in clinical isolates of Enterococcus faecium is mediated by an upregulation of plasmid-encoded tetracycline determinants tet(L) and tet(M). J. Antimicrob. Chemother. 2016, 71, 871–881. [Google Scholar] [CrossRef] [Green Version]
- Holmes, N.E.; Ballard, S.A.; Lam, M.M.; Johnson, P.D.; Grayson, M.L.; Stinear, T.P.; Howden, B.P. Genomic analysis of teicoplanin resistance emerging during treatment of vanB vancomycin-resistant Enterococcus faecium infections in solid organ transplant recipients including donor-derived cases. J. Antimicrob. Chemother. 2013, 68, 2134–2139. [Google Scholar] [CrossRef] [Green Version]
- Zhanel, G.G.; Calic, D.; Schweizer, F.; Zelenitsky, S.; Adam, H.; Lagacé-Wiens, P.R.; Rubinstein, E.; Gin, A.S.; Hoban, D.J.; Karlowsky, J.A. New lipoglycopeptides: A comparative review of dalbavancin, oritavancin and telavancin. Drugs 2010, 70, 859–886. [Google Scholar] [CrossRef]
- Arhin, F.F.; Belley, A.; McKay, G.A.; Moeck, G. Characterization of the in vitro activity of novel lipoglycopeptide antibiotics. Curr. Protoc. Microbiol. 2010, 16, 17.1.1–17.1.22. [Google Scholar] [CrossRef]
- Rosenthal, S.; Decano, A.G.; Bandali, A.; Lai, D.; Malat, G.E.; Bias, T.E. Oritavancin (Orbactiv): A New-Generation Lipoglycopeptide for the Treatment Of Acute Bacterial Skin and Skin Structure Infections. Pharm. Ther. 2018, 43, 143–179. [Google Scholar]
- Jones, R.N.; Flamm, R.K.; Sader, H.S. Surveillance of dalbavancin potency and spectrum in the United States (2012). Diagn. Microbiol. Infect. Dis. 2013, 76, 122–123. [Google Scholar] [CrossRef] [PubMed]
- Sader, H.S.; Mendes, R.E.; Pfaller, M.A.; Flamm, R.K. Antimicrobial activity of dalbavancin tested against Gram-positive organisms isolated from patients with infective endocarditis in US and European medical centres. J. Antimicrob. Chemother. 2019, 74, 1306–1310. [Google Scholar] [CrossRef]
- Raad, I.; Darouiche, R.; Vazquez, J.; Lentnek, A.; Hachem, R.; Hanna, H.; Goldstein, B.; Henkel, T.; Seltzer, E. Efficacy and safety of weekly dalbavancin therapy for catheter-related bloodstream infection caused by gram-positive pathogens. Clin. Infect. Dis. 2005, 40, 374–380. [Google Scholar] [CrossRef] [PubMed]
- Morata, L.; Cobo, J.; Fernandez-Sampedro, M.; Vasco, P.G.; Ruano, E.; Lora-Tamayo, J.; Somolinos, M.S.; Ruano, P.G.; Nieto, A.R.; Arnaiz, A.; et al. Safety and Efficacy of Prolonged Use of Dalbavancin in Bone and Joint Infections. Antimicrob. Agents Chemother. 2019, 63, e02280-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Wang, J.; Wang, R.; Li, Y.; Cai, Y. Efficacy and safety of dalbavancin in the treatment of Gram-positive bacterial infections. J. Glob. Antimicrob. Resist. 2020, 24, 72–80. [Google Scholar] [CrossRef] [PubMed]
- Rappo, U.; Puttagunta, S.; Shevchenko, V.; Shevchenko, A.; Jandourek, A.; Gonzalez, P.L.; Suen, A.; Casullo, V.M.; Melnick, D.; Miceli, R.; et al. Dalbavancin for the Treatment of Osteomyelitis in Adult Patients: A Randomized Clinical Trial of Efficacy and Safety. Open Forum Infect. Dis. 2019, 6, ofy331. [Google Scholar] [CrossRef] [PubMed]
- Biedenbach, D.J.; Bell, J.M.; Sader, H.S.; Turnidge, J.D.; Jones, R.N. Activities of dalbavancin against a worldwide collection of 81,673 gram-positive bacterial isolates. Antimicrob. Agents Chemother. 2009, 53, 1260–1263. [Google Scholar] [CrossRef] [Green Version]
- Kavanagh, A.; Ramu, S.; Gong, Y.; Cooper, M.A.; Blaskovich, M.A.T. Effects of Microplate Type and Broth Additives on Microdilution MIC Susceptibility Assays. Antimicrob. Agents Chemother. 2019, 63, e01760. [Google Scholar] [CrossRef] [Green Version]
- Cetinkaya, Y.; Falk, P.; Mayhall, C.G. Vancomycin-resistant enterococci. Clin. Microbiol. Rev. 2000, 13, 686–707. [Google Scholar] [CrossRef] [PubMed]
- Pfaller, M.A.; Flamm, R.K.; Castanheira, M.; Sader, H.S.; Mendes, R.E. Dalbavancin in-vitro activity obtained against Gram-positive clinical isolates causing bone and joint infections in US and European hospitals (2011–2016). Int. J. Antimicrob. Agents 2018, 51, 608–611. [Google Scholar] [CrossRef]
- Pfaller, M.A.; Mendes, R.E.; Sader, H.S.; Castanheira, M.; Flamm, R.K. Activity of dalbavancin tested against Gram-positive clinical isolates causing skin and skin-structure infections in paediatric patients from US hospitals (2014–2015). J. Glob. Antimicrob. Resist. 2017, 11, 4–7. [Google Scholar] [CrossRef] [PubMed]
- Turnidge, J.; Kahlmeter, G.; Kronvall, G. Statistical characterisation of bacterial wild-type MIC value distributions and the determination of epidemiological cut-off values. Clin. Microbiol. Infect. 2006, 12, 418–425. [Google Scholar] [CrossRef]
- Klare, I.; Konstabel, C.; Mueller-Bertling, S.; Werner, G.; Strommenger, B.; Kettlitz, C.; Borgmann, S.; Schulte, B.; Jonas, D.; Serr, A.; et al. Spread of ampicillin/vancomycin-resistant Enterococcus faecium of the epidemic-virulent clonal complex-17 carrying the genes esp and hyl in German hospitals. Eur. J. Clin. Microbiol. Infect. Dis. 2005, 24, 815–825. [Google Scholar] [CrossRef] [PubMed]
- Andrews, S.; Krueger, S.; Segonds-Pichon, F.; Biggins, L.; Krueger, C.; Wingett, S. FastQC: A Quality Control Tool for High Throughput Sequence Data; Babraham Institute: Cambridge, UK, 2012. [Google Scholar]
- Wood, D.E.; Salzberg, S.L. Kraken: Ultrafast metagenomic sequence classification using exact alignments. Genome Biol. 2014, 15, R46. [Google Scholar] [CrossRef] [Green Version]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.; Durbin, R. Fast and accurate short read alignment with Burrows-Wheeler transform. Bioinformatics 2009, 25, 1754–1760. [Google Scholar] [CrossRef] [Green Version]
- Camacho, C.; Coulouris, G.; Avagyan, V.; Ma, N.; Papadopoulos, J.; Bealer, K.; Madden, T.L. BLAST+: Architecture and applications. BMC Bioinform. 2009, 10, 421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed] [Green Version]

{kind=link}
| Genotype/Method | n | MIC [mg/L] | MIC50 [mg/L] | MIC90 [mg/L] | ECOFF * [99%] | CLSI ** [% S] | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ≤0.08 | 0.016 | 0.032 | 0.064 | 0.125 | 0.25 | 0.5 | 1 | 2 | 4 | 8 | >8 | ||||||
| vanA, BMD | 50 | 1 | 1 | 3 | 1 | 1 | 21 | 22 | 8 | >8 | - | 10 | |||||
| vanA, strip | 50 | 1 | 1 | 1 | 1 | 1 | 45 | >8 | >8 | - | 8 | ||||||
| vanB, BMD | 101 | 1 | 6 | 64 | 27 | 3 | 0.032 | 0.064 | 0.25 | 100 | |||||||
| vanB, strip | 101 | 6 | 45 | 44 | 6 | 0.016 | 0.032 | 0.125 | 100 | ||||||||
| van-, BMD | 25 | 3 | 6 | 11 | 5 | 0.064 | 0.125 | 0.5 | 100 | ||||||||
| van-, strip | 25 | 6 | 11 | 7 | 1 | 0.032 | 0.064 | 0.25 | 100 | ||||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Weber, R.E.; Fleige, C.; Layer, F.; Neumann, B.; Kresken, M.; Werner, G. Determination of a Tentative Epidemiological Cut-Off Value (ECOFF) for Dalbavancin and Enterococcus faecium. Antibiotics 2021, 10, 915. https://doi.org/10.3390/antibiotics10080915
Weber RE, Fleige C, Layer F, Neumann B, Kresken M, Werner G. Determination of a Tentative Epidemiological Cut-Off Value (ECOFF) for Dalbavancin and Enterococcus faecium. Antibiotics. 2021; 10(8):915. https://doi.org/10.3390/antibiotics10080915
Chicago/Turabian StyleWeber, Robert E., Carola Fleige, Franziska Layer, Bernd Neumann, Michael Kresken, and Guido Werner. 2021. "Determination of a Tentative Epidemiological Cut-Off Value (ECOFF) for Dalbavancin and Enterococcus faecium" Antibiotics 10, no. 8: 915. https://doi.org/10.3390/antibiotics10080915
APA StyleWeber, R. E., Fleige, C., Layer, F., Neumann, B., Kresken, M., & Werner, G. (2021). Determination of a Tentative Epidemiological Cut-Off Value (ECOFF) for Dalbavancin and Enterococcus faecium. Antibiotics, 10(8), 915. https://doi.org/10.3390/antibiotics10080915