
Alternatives to Fight Vancomycin-Resistant Staphylococci and Enterococci


Abstract
1. Introduction
2. Alternatives to Vancomycin
2.1. Traditional Antimicrobials
2.1.1. Conventional Antibiotics
2.1.2. Modified Antibiotics
2.1.3. Combinations of Antibiotics
2.2. Non-Traditional Antimicrobials
2.2.1. Antimicrobial Peptides and Bacteriocins
Antimicrobial Peptides
Bacteriocins
2.2.2. Bacteriophages
2.2.3. Nanoparticles
3. Conclusions
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- O’Neill, J. Tackling Drug-Resistant Infections Globally: Final Report and Recommendations; Government of the United Kingdom: London, UK, 2016.
- Touat, M.; Opatowski, M.; Brun-Buisson, C.; Cosker, K.; Guillemot, D.; Salomon, J.; Tuppin, P.; de Lagasnerie, G.; Watier, L. A Payer Perspective of the Hospital Inpatient Additional Care Costs of Antimicrobial Resistance in France: A Matched Case-Control Study. Appl. Health Econ. Health Policy 2019, 17, 381–389. [Google Scholar] [CrossRef] [PubMed]
- Cassini, A.; Högberg, L.D.; Plachouras, D.; Quattrocchi, A.; Hoxha, A.; Simonsen, G.S.; Colomb-Cotinat, M.; Kretzschmar, M.E.; Devleesschauwer, B.; Cecchini, M.; et al. Attributable Deaths and Disability-Adjusted Life-Years Caused by Infections with Antibiotic-Resistant Bacteria in the EU and the European Economic Area in 2015: A Population-Level Modelling Analysis. Lancet Infect. Dis. 2019, 19, 56–66. [Google Scholar] [CrossRef]
- Theuretzbacher, U.; Bush, K.; Harbarth, S.; Paul, M.; Rex, J.H.; Tacconelli, E.; Thwaites, G.E. Critical Analysis of Antibacterial Agents in Clinical Development. Nat. Rev. Microbiol. 2020, 18, 286–298. [Google Scholar] [CrossRef] [PubMed]
- McKenna, M. The Antibiotic Paradox: Why Companies Can’t Afford to Create Life-Saving Drugs. Nature 2020, 584, 338–341. [Google Scholar] [CrossRef] [PubMed]
- Tacconelli, E.; Carrara, E.; Savoldi, A.; Harbarth, S.; Mendelson, M.; Monnet, D.L.; Pulcini, C.; Kahlmeter, G.; Kluytmans, J.; Carmeli, Y.; et al. Discovery, Research, and Development of New Antibiotics: The WHO Priority List of Antibiotic-Resistant Bacteria and Tuberculosis. Lancet Infect. Dis. 2018, 18, 318–327. [Google Scholar] [CrossRef]
- Rex, J.H.; Lynch, H.F.; Cohen, I.G.; Darrow, J.J.; Outterson, K. Designing Development Programs for Non-Traditional Antibacterial Agents. Nat. Commun. 2019, 10, 3416. [Google Scholar] [CrossRef]
- Levine, D.P. Vancomycin: A History. Clin. Infect. Dis. 2006, 42 (Suppl. S1), S5–S12. [Google Scholar] [CrossRef]
- Arias, C.A.; Murray, B.E. The Rise of the Enterococcus: Beyond Vancomycin Resistance. Nat. Rev. Microbiol. 2012, 10, 266–278. [Google Scholar] [CrossRef]
- Kristich, C.J.; Rice, L.B.; Arias, C.A. Enterococcal Infection—Treatment and Antibiotic Resistance. In Enterococci: From Commensals to Leading Causes of Drug Resistant Infection; Gilmore, M.S., Clewell, D.B., Ike, Y., Shankar, N., Eds.; Massachusetts Eye and Ear Infirmary: Boston, MA, USA, 2014. [Google Scholar]
- Kaplan, A.H.; Gilligan, P.H.; Facklam, R.R. Recovery of Resistant Enterococci during Vancomycin Prophylaxis. J. Clin. Microbiol. 1988, 26, 1216–1218. [Google Scholar] [CrossRef] [PubMed]
- Leclercq, R.; Derlot, E.; Duval, J.; Courvalin, P. Plasmid-Mediated Resistance to Vancomycin and Teicoplanin in Enterococcus faecium. N. Engl. J. Med. 1988, 319, 157–161. [Google Scholar] [CrossRef]
- Gholizadeh, Y.; Courvalin, P. Acquired and Intrinsic Glycopeptide Resistance in Enterococci. Int. J. Antimicrob. Agents 2000, 16, 11–17. [Google Scholar] [CrossRef]
- Reynolds, P.E. Structure, Biochemistry and Mechanism of Action of Glycopeptide Antibiotics. Eur. J. Clin. Microbiol. Infect. Dis. 1989, 8, 943–950. [Google Scholar] [CrossRef] [PubMed]
- Schaechter, M. Desk Encyclopedia of Microbiology, 2nd ed.; Elsevier/AP: Amsterdam, The Netherlands, 2009; ISBN 978-0-12-374980-2. [Google Scholar]
- Nitanai, Y.; Kikuchi, T.; Kakoi, K.; Hanamaki, S.; Fujisawa, I.; Aoki, K. Crystal Structures of the Complexes between Vancomycin and Cell-Wall Precursor Analogs. J. Mol. Biol. 2009, 385, 1422–1432. [Google Scholar] [CrossRef]
- ECDC. Antimicrobial Resistance in the EU/EEA (EARS-Net)—Annual Epidemiological Report for 2019. Available online: ecdc.europa.eu/en/publications-data/surveillance-antimicrobial-resistance-europe-2019 (accessed on 1 January 2021).
- DiazGranados, C.A.; Zimmer, S.M.; Mitchel, K.; Jernigan, J.A. Comparison of Mortality Associated with Vancomycin-Resistant and Vancomycin-Susceptible Enterococcal Bloodstream Infections: A Meta-Analysis. Clin. Infect. Dis. 2005, 41, 327–333. [Google Scholar] [CrossRef]
- Reinseth, I.S.; Ovchinnikov, K.V.; Tønnesen, H.H.; Carlsen, H.; Diep, D.B. The Increasing Issue of Vancomycin-Resistant Enterococci and the Bacteriocin Solution. Probiot. Antimicrob. Proteins 2020, 12, 1203–1217. [Google Scholar] [CrossRef] [PubMed]
- Bender, J.K.; Cattoir, V.; Hegstad, K.; Sadowy, E.; Coque, T.M.; Westh, H.; Hammerum, A.M.; Schaffer, K.; Burns, K.; Murchan, S.; et al. Update on Prevalence and Mechanisms of Resistance to Linezolid, Tigecycline and Daptomycin in Enterococci in Europe: Towards a Common Nomenclature. Drug Resist. Updates 2018, 40, 25–39. [Google Scholar] [CrossRef] [PubMed]
- Contreras, G.A.; Munita, J.M.; Arias, C.A. Novel Strategies for the Management of Vancomycin-Resistant Enterococcal Infections. Curr. Infect. Dis. Rep. 2019, 21, 22. [Google Scholar] [CrossRef]
- Yim, J.; Smith, J.R.; Rybak, M.J. Role of Combination Antimicrobial Therapy for Vancomycin-Resistant Enterococcus faecium Infections: Review of the Current Evidence. Pharmacotherapy 2017, 37, 579–592. [Google Scholar] [CrossRef]
- Livermore, D.M. Linezolid in Vitro: Mechanism and Antibacterial Spectrum. J. Antimicrob. Chemother. 2003, 51, ii9–ii16. [Google Scholar] [CrossRef]
- Long, K.S.; Poehlsgaard, J.; Kehrenberg, C.; Schwarz, S.; Vester, B. The Cfr RRNA Methyltransferase Confers Resistance to Phenicols, Lincosamides, Oxazolidinones, Pleuromutilins, and Streptogramin A Antibiotics. Antimicrob. Agents Chemother. 2006, 50, 2500–2505. [Google Scholar] [CrossRef]
- Locke, J.B.; Zurenko, G.E.; Shaw, K.J.; Bartizal, K. Tedizolid for the Management of Human Infections: In Vitro Characteristics. Clin. Infect. Dis. 2014, 58, S35–S42. [Google Scholar] [CrossRef]
- Brenciani, A.; Morroni, G.; Vincenzi, C.; Manso, E.; Mingoia, M.; Giovanetti, E.; Varaldo, P.E. Detection in Italy of Two Clinical Enterococcus faecium Isolates Carrying Both the Oxazolidinone and Phenicol Resistance Gene optrA and a Silent Multiresistance Gene cfr: Table 1. J. Antimicrob. Chemother. 2016, 71, 1118–1119. [Google Scholar] [CrossRef]
- Sharkey, L.K.R.; O’Neill, A.J. Antibiotic Resistance ABC-F Proteins: Bringing Target Protection into the Limelight. ACS Infect. Dis. 2018, 4, 239–246. [Google Scholar] [CrossRef] [PubMed]
- Gawryszewska, I.; Żabicka, D.; Hryniewicz, W.; Sadowy, E. Linezolid-Resistant Enterococci in Polish Hospitals: Species, Clonality and Determinants of Linezolid Resistance. Eur. J. Clin. Microbiol. Infect. Dis. 2017, 36, 1279–1286. [Google Scholar] [CrossRef] [PubMed]
- Nannini, E.; Murray, B.E.; Arias, C.A. Resistance or Decreased Susceptibility to Glycopeptides, Daptomycin, and Linezolid in Methicillin-Resistant Staphylococcus aureus. Curr. Opin. Pharmacol. 2010, 10, 516–521. [Google Scholar] [CrossRef]
- Bi, R.; Qin, T.; Fan, W.; Ma, P.; Gu, B. The Emerging Problem of Linezolid-Resistant Enterococci. J. Glob. Antimicrob. Resist. 2018, 13, 11–19. [Google Scholar] [CrossRef]
- Krull, M.; Klare, I.; Ross, B.; Trenschel, R.; Beelen, D.W.; Todt, D.; Steinmann, E.; Buer, J.; Rath, P.-M.; Steinmann, J. Emergence of Linezolid- and Vancomycin-Resistant Enterococcus faecium in a Department for Hematologic Stem Cell Transplantation. Antimicrob. Resist. Infect. Control 2016, 5, 31. [Google Scholar] [CrossRef][Green Version]
- Heininger, A.; Zimmermann, S.; Bootsveld, C.; Boutin, S.; Nurjadi, D. Low Prevalence of Combined Linezolid- and Vancomycin-Resistant Enterococcus faecium from Hospital Admission Screening in an Endemic Region in Germany. J. Glob. Antimicrob. Resist. 2020, 22, 646–650. [Google Scholar] [CrossRef] [PubMed]
- Zhanel, G.G.; Love, R.; Adam, H.; Golden, A.; Zelenitsky, S.; Schweizer, F.; Gorityala, B.; Lagacé-Wiens, P.R.S.; Rubinstein, E.; Walkty, A.; et al. Tedizolid: A Novel Oxazolidinone with Potent Activity Against Multidrug-Resistant Gram-Positive Pathogens. Drugs 2015, 75, 253–270. [Google Scholar] [CrossRef] [PubMed]
- Zhanel, G.G. Antibiotic Activity against Urinary Tract Infection (UTI) Isolates of Vancomycin-Resistant Enterococci (VRE): Results from the 2002 North American Vancomycin Resistant Enterococci Susceptibility Study (NAVRESS). J. Antimicrob. Chemother. 2003, 52, 382–388. [Google Scholar] [CrossRef] [PubMed][Green Version]
- O’Driscoll, T.; Crank, C.W. Vancomycin-Resistant Enterococcal Infections: Epidemiology, Clinical Manifestations, and Optimal Management. Infect. Drug Resist. 2015, 8, 217–230. [Google Scholar] [CrossRef]
- Wang, Y.; Li, X.; Wang, Y.; Schwarz, S.; Shen, J.; Xia, X. Intracellular Accumulation of Linezolid and Florfenicol in OptrA-Producing Enterococcus faecalis and Staphylococcus aureus. Molecules 2018, 23, 3195. [Google Scholar] [CrossRef]
- Straus, S.K.; Hancock, R.E.W. Mode of Action of the New Antibiotic for Gram-Positive Pathogens Daptomycin: Comparison with Cationic Antimicrobial Peptides and Lipopeptides. Biochim. Biophys. Acta 2006, 1758, 1215–1223. [Google Scholar] [CrossRef] [PubMed]
- Chuang, Y.-C.; Wang, J.-T.; Lin, H.-Y.; Chang, S.-C. Daptomycin versus Linezolid for Treatment of Vancomycin-Resistant Enterococcal Bacteremia: Systematic Review and Meta-Analysis. BMC Infect. Dis 2014, 14, 687. [Google Scholar] [CrossRef]
- Werth, B.J.; Barber, K.E.; Tran, K.-N.T.; Nonejuie, P.; Sakoulas, G.; Pogliano, J.; Rybak, M.J. Ceftobiprole and Ampicillin Increase Daptomycin Susceptibility of Daptomycin-Susceptible and -Resistant VRE. J. Antimicrob. Chemother. 2015, 70, 489–493. [Google Scholar] [CrossRef] [PubMed]
- Miller, W.R.; Bayer, A.S.; Arias, C.A. Mechanism of Action and Resistance to Daptomycin in Staphylococcus aureus and Enterococci. Cold Spring Harb. Perspect. Med. 2016, 6, a026997. [Google Scholar] [CrossRef]
- Tran, T.T.; Munita, J.M.; Arias, C.A. Mechanisms of Drug Resistance: Daptomycin Resistance. Ann. N. Y. Acad. Sci. 2015, 1354, 32–53. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.; Davlieva, M.; Panesso, D.; Rincon, S.; Miller, W.R.; Diaz, L.; Reyes, J.; Cruz, M.R.; Pemberton, O.; Nguyen, A.H.; et al. Antimicrobial Sensing Coupled with Cell Membrane Remodeling Mediates Antibiotic Resistance and Virulence in Enterococcus faecalis. Proc. Natl. Acad. Sci. USA 2019, 116, 26925–26932. [Google Scholar] [CrossRef]
- Werth, B.J.; Steed, M.E.; Ireland, C.E.; Tran, T.T.; Nonejuie, P.; Murray, B.E.; Rose, W.E.; Sakoulas, G.; Pogliano, J.; Arias, C.A.; et al. Defining Daptomycin Resistance Prevention Exposures in Vancomycin-Resistant Enterococcus faecium and E. faecalis. Antimicrob. Agents Chemother. 2014, 58, 5253–5261. [Google Scholar] [CrossRef]
- Dönhöfer, A.; Franckenberg, S.; Wickles, S.; Berninghausen, O.; Beckmann, R.; Wilson, D.N. Structural Basis for TetM-Mediated Tetracycline Resistance. Proc. Natl. Acad. Sci. USA 2012, 109, 16900–16905. [Google Scholar] [CrossRef]
- Beabout, K.; Hammerstrom, T.G.; Perez, A.M.; Magalhães, B.F.; Prater, A.G.; Clements, T.P.; Arias, C.A.; Saxer, G.; Shamoo, Y. The Ribosomal S10 Protein Is a General Target for Decreased Tigecycline Susceptibility. Antimicrob. Agents Chemother. 2015, 59, 5561–5566. [Google Scholar] [CrossRef]
- Bender, J.K.; Klare, I.; Fleige, C.; Werner, G. A Nosocomial Cluster of Tigecycline- and Vancomycin-Resistant Enterococcus faecium Isolates and the Impact of RpsJ and Tet(M) Mutations on Tigecycline Resistance. Microb. Drug Resist. 2020, 26, 576–582. [Google Scholar] [CrossRef]
- Hamzah, A.M.C.; Yeo, C.C.; Puah, S.M.; Chua, K.H.; Rahman, N.I.A.; Abdullah, F.H.; Othman, N.; Chew, C.H. Tigecycline and Inducible Clindamycin Resistance in Clinical Isolates of Methicillin-Resistant Staphylococcus aureus from Terengganu, Malaysia. J. Med. Microbiol. 2019, 68, 1299–1305. [Google Scholar] [CrossRef]
- Bensaci, M.; Sahm, D. Surveillance of Tedizolid Activity and Resistance: In Vitro Susceptibility of Gram-Positive Pathogens Collected over 5 Years from the United States and Europe. Diagn. Microbiol. Infect. Dis. 2017, 87, 133–138. [Google Scholar] [CrossRef]
- Zhen, J.-B.; Kang, P.-W.; Zhao, M.-H.; Yang, K.-W. Silver Nanoparticle Conjugated Star PCL-b-AMPs Copolymer as Nanocomposite Exhibits Efficient Antibacterial Properties. Bioconjug. Chem. 2020, 31, 51–63. [Google Scholar] [CrossRef] [PubMed]
- Jain, R.K.; Trias, J.; Ellman, J.A. D-Ala-D-Lac Binding Is Not Required for the High Activity of Vancomycin Dimers against Vancomycin Resistant Enterococci. J. Am. Chem. Soc. 2003, 125, 8740–8741. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, O.; Nakamura, J.; Yamashiro, H.; Miura, K.; Hayashi, S.; Umetsu, K.; Xu, S.; Maki, H.; Arimoto, H. New Insight into the Mode of Action of Vancomycin Dimers in Bacterial Cell Wall Synthesis. Med. Chem. Commun. 2011, 2, 278. [Google Scholar] [CrossRef]
- Nakamura, J.; Yamashiro, H.; Hayashi, S.; Yamamoto, M.; Miura, K.; Xu, S.; Doi, T.; Maki, H.; Yoshida, O.; Arimoto, H. Elucidation of the Active Conformation of Vancomycin Dimers with Antibacterial Activity against Vancomycin-Resistant Bacteria. Chem. Eur. J. 2012, 18, 12681–12689. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Yoshida, O.; Hayashi, S.; Arimoto, H. Synthesis of Rigidly-Linked Vancomycin Dimers and Their in Vivo Efficacy against Resistant Bacteria. Chem. Commun. 2007, 3, 251–253. [Google Scholar] [CrossRef]
- Szűcs, Z.; Ostorházi, E.; Kicsák, M.; Nagy, L.; Borbás, A.; Herczegh, P. New Semisynthetic Teicoplanin Derivatives Have Comparable in Vitro Activity to That of Oritavancin against Clinical Isolates of VRE. J. Antibiot. 2019, 72, 524–534. [Google Scholar] [CrossRef]
- Malabarba, A.; Ciabatti, R. Glycopeptide Derivatives. Curr. Med. Chem. 2001, 8, 1759–1773. [Google Scholar] [CrossRef] [PubMed]
- Zeng, D.; Debabov, D.; Hartsell, T.L.; Cano, R.J.; Adams, S.; Schuyler, J.A.; McMillan, R.; Pace, J.L. Approved Glycopeptide Antibacterial Drugs: Mechanism of Action and Resistance. Cold Spring Harb. Perspect. Med. 2016, 6, a026989. [Google Scholar] [CrossRef]
- Jones, R.N.; Fritsche, T.R.; Sader, H.S.; Goldstein, B.P. Antimicrobial Spectrum and Potency of Dalbavancin Tested Against Clinical Isolates from Europe and North America (2003): Initial Results from an International Surveillance Protocol. J. Chemother. 2005, 17, 593–600. [Google Scholar] [CrossRef] [PubMed]
- Lozano, C.; Torres, C. Actualización en la resistencia antibiótica en Gram positivos. Enferm. Infecc. Microbiol. Clin. 2017, 35, 2–8. [Google Scholar] [CrossRef]
- Karaoui, L.R.; EL-Lababidi, R.; Chahine, E.B. Oritavancin: An Investigational Lipoglycopeptide Antibiotic. Am. J. Health Syst. Pharm. 2013, 70, 23–33. [Google Scholar] [CrossRef] [PubMed]
- Rosenthal, S.; Decano, A.G.; Bandali, A.; Lai, D.; Malat, G.E.; Bias, T.E. Oritavancin (Orbactiv): A New-Generation Lipoglycopeptide for the Treatment of Acute Bacterial Skin and Skin Structure Infections. Pharm. Ther. 2018, 43, 143–179. [Google Scholar]
- Okano, A.; Isley, N.A.; Boger, D.L. Peripheral Modifications of [Ψ[CH2NH]Tpg4] Vancomycin with Added Synergistic Mechanisms of Action Provide Durable and Potent Antibiotics. Proc. Natl. Acad. Sci. USA 2017, 114, 201704125. [Google Scholar] [CrossRef]
- Aktas, G. In Vitro Synergistic Effect of Vancomycin Combined with Daptomycin Against Vancomycin-Resistant Enterococci. Microb. Drug Resist. 2019, 25, 1484–1489. [Google Scholar] [CrossRef]
- de Oliveira, K.S.; Queiroz, P.R.M.; Fensterseifer, I.C.M.; Migliolo, L.; Oliveira, A.L.; Franco, O.L. Purified Citritin in Combination with Vancomycin Inhibits VRE in Vitro and in Vivo. Microbiology 2017, 163, 1525–1531. [Google Scholar] [CrossRef] [PubMed]
- Luther, M.K.; Arvanitis, M.; Mylonakis, E.; LaPlante, K.L. Activity of Daptomycin or Linezolid in Combination with Rifampin or Gentamicin against Biofilm-Forming Enterococcus faecalis or E. faecium in an In Vitro Pharmacodynamic Model Using Simulated Endocardial Vegetations and an In Vivo Survival Assay Using Galleria mellonella Larvae. Antimicrob. Agents Chemother. 2014, 58, 4612–4620. [Google Scholar] [CrossRef]
- Chen, H.; Du, Y.; Xia, Q.; Li, Y.; Song, S.; Huang, X. Role of Linezolid Combination Therapy for Serious Infections: Review of the Current Evidence. Eur. J. Clin. Microbiol. Infect. Dis. 2020, 39, 1043–1052. [Google Scholar] [CrossRef]
- Meyer, K.A.; Deraedt, M.F.; Harrington, A.T.; Danziger, L.H.; Wenzler, E. Efficacy of Oritavancin Alone and in Combination against Vancomycin-Susceptible and -Resistant Enterococci in an in-Vivo Galleria mellonella Survival Model. Int. J. Antimicrob. Agents 2019, 54, 197–201. [Google Scholar] [CrossRef]
- Qi, C.; Xu, S.; Wu, M.; Zhu, S.; Liu, Y.; Huang, H.; Zhang, G.; Li, J.; Huang, X. Pharmacodynamics of Linezolid-Plus-Fosfomycin Against Vancomycin-Susceptible and -Resistant Enterococci In Vitro and In Vivo of A Galleria mellonella Larval Infection Model. Infect. Drug Resist. 2019, 12, 3497–3505. [Google Scholar] [CrossRef]
- Tang, H.-J.; Chen, C.-C.; Zhang, C.-C.; Su, B.-A.; Li, C.-M.; Weng, T.-C.; Chiang, S.-R.; Ko, W.-C.; Chuang, Y.-C. In Vitro Efficacy of Fosfomycin-Based Combinations against Clinical Vancomycin-Resistant Enterococcus Isolates. Diagn. Microbiol. Infect. Dis. 2013, 77, 254–257. [Google Scholar] [CrossRef] [PubMed]
- Tang, H.-J.; Chen, C.-C.; Cheng, K.-C.; Toh, H.-S.; Su, B.-A.; Chiang, S.-R.; Ko, W.-C.; Chuang, Y.-C. In Vitro Efficacy of Fosfomycin-Containing Regimens against Methicillin-Resistant Staphylococcus aureus in Biofilms. J. Antimicrob. Chemother. 2012, 67, 944–950. [Google Scholar] [CrossRef]
- Smith, J.R.; Barber, K.E.; Raut, A.; Aboutaleb, M.; Sakoulas, G.; Rybak, M.J. β-Lactam Combinations with Daptomycin Provide Synergy against Vancomycin-Resistant Enterococcus faecalis and Enterococcus faecium. J. Antimicrob. Chemother. 2015, 70, 1738–1743. [Google Scholar] [CrossRef] [PubMed]
- Smith, J.R.; Yim, J.; Raut, A.; Rybak, M.J. Oritavancin Combinations with β-Lactams against Multidrug-Resistant Staphylococcus aureus and Vancomycin-Resistant Enterococci. Antimicrob. Agents Chemother. 2016, 60, 2352–2358. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.; Meyer, K.; Harrington, A.T.; Danziger, L.H.; Wenzler, E. In Vitro Activity of Oritavancin Alone or in Combination against Vancomycin-Susceptible and -Resistant Enterococci. J. Antimicrob. Chemother. 2019, 74, 1300–1305. [Google Scholar] [CrossRef] [PubMed]
- Mroue, N.; Arya, A.; Gandt, A.B.; Russell, C.; Han, A.; Gavrish, E.; LaFleur, M. Pharmacodynamics of ClpP-Activating Antibiotic Combinations against Gram-Positive Pathogens. Antimicrob. Agents Chemother. 2019, 64, e01554-19. [Google Scholar] [CrossRef] [PubMed]
- Gandt, A.B.; Griffith, E.C.; Lister, I.M.; Billings, L.L.; Han, A.; Tangallapally, R.; Zhao, Y.; Singh, A.P.; Lee, R.E.; LaFleur, M.D. In Vivo and in Vitro Effects of a ClpP-Activating Antibiotic against Vancomycin-Resistant Enterococci. Antimicrob. Agents Chemother. 2018, 62, e00424-18. [Google Scholar] [CrossRef]
- Mull, R.W.; Brennan, A.A.; Russ, B.R.; Tal-Gan, Y. Attenuating the Selection of Vancomycin Resistance Among Enterococci through the Development of Peptide-Based Vancomycin Antagonists. ACS Infect. Dis. 2020, 6, 2913–2925. [Google Scholar] [CrossRef]
- Liu, Y.; Jia, Y.; Yang, K.; Li, R.; Xiao, X.; Wang, Z. Antagonizing Vancomycin Resistance in Enterococcus by Surface Localized Antimicrobial Display-Derived Peptides. ACS Infect. Dis. 2020, 6, 761–767. [Google Scholar] [CrossRef]
- Thappeta, K.R.V.; Vikhe, Y.S.; Yong, A.M.H.; Chan-Park, M.B.; Kline, K.A. Combined Efficacy of an Antimicrobial Cationic Peptide Polymer with Conventional Antibiotics to Combat Multidrug-Resistant Pathogens. ACS Infect. Dis. 2020, 6, 1228–1237. [Google Scholar] [CrossRef] [PubMed]
- Shurko, J.F. Evaluation of LL-37 Antimicrobial Peptide Derivatives Alone and in Combination with Vancomycin against S. aureus. J. Antibiot. 2018, 71, 971–974. [Google Scholar] [CrossRef] [PubMed]
- Umstätter, F.; Domhan, C.; Hertlein, T.; Ohlsen, K.; Mühlberg, E.; Kleist, C.; Zimmermann, S.; Beijer, B.; Klika, K.D.; Haberkorn, U.; et al. Vancomycin Resistance Is Overcome by Conjugation of Polycationic Peptides. Angew. Chem. Int. Ed. 2020, 59, 8823–8827. [Google Scholar] [CrossRef]
- Kim, S.G.; Becattini, S.; Moody, T.U.; Shliaha, P.V.; Littmann, E.R.; Seok, R.; Gjonbalaj, M.; Eaton, V.; Fontana, E.; Amoretti, L.; et al. Microbiota-Derived Lantibiotic Restores Resistance against Vancomycin-Resistant Enterococcus. Nature 2019, 572, 665–669. [Google Scholar] [CrossRef]
- Lawton, E.; Ross, R.; Hill, C.; Cotter, P. Two-Peptide Lantibiotics: A Medical Perspective. MRMC 2007, 7, 1236–1247. [Google Scholar] [CrossRef] [PubMed]
- Kruszewska, D.; Sahl, H.-G.; Bierbaum, G.; Pag, U.; Hynes, S.O. Mersacidin Eradicates Methicillin-Resistant Staphylococcus aureus (MRSA) in a Mouse Rhinitis Model. J. Antimicrob. Chemother. 2004, 54, 648–653. [Google Scholar] [CrossRef]
- Piper, C.; Casey, P.G.; Hill, C.; Cotter, P.D.; Ross, R.P. The Lantibiotic Lacticin 3147 Prevents Systemic Spread of Staphylococcus aureus in a Murine Infection Model. Int. J. Microbiol. 2012, 2012, 806230. [Google Scholar] [CrossRef] [PubMed]
- Aunpad, R.; Na-Bangchang, K. Pumilicin 4, A Novel Bacteriocin with Anti-MRSA and Anti-VRE Activity Produced by Newly Isolated Bacteria Bacillus pumilus Strain WAPB4. Curr. Microbiol. 2007, 55, 308–313. [Google Scholar] [CrossRef] [PubMed]
- Ovchinnikov, K.V.; Kristiansen, P.E.; Straume, D.; Jensen, M.S.; Aleksandrzak-Piekarczyk, T.; Nes, I.F.; Diep, D.B. The Leaderless Bacteriocin Enterocin K1 Is Highly Potent against Enterococcus faecium: A Study on Structure, Target Spectrum and Receptor. Front. Microbiol. 2017, 8, 774. [Google Scholar] [CrossRef]
- Phumisantiphong, U.; Siripanichgon, K.; Reamtong, O.; Diraphat, P. A Novel Bacteriocin from Enterococcus faecalis 478 Exhibits a Potent Activity against Vancomycin-Resistant Enterococci. PLoS ONE 2017, 12, e0186415. [Google Scholar] [CrossRef]
- Regmi, S.; Choi, Y.H.; Choi, Y.S.; Kim, M.R.; Yoo, J.C. Antimicrobial Peptide Isolated from Bacillus amyloliquefaciens K14 Revitalizes Its Use in Combinatorial Drug Therapy. Folia Microbiol. 2017, 62, 127–138. [Google Scholar] [CrossRef]
- Montalbán-López, M.; Cebrián, R.; Galera, R.; Mingorance, L.; Martín-Platero, A.M.; Valdivia, E.; Martínez-Bueno, M.; Maqueda, M. Synergy of the Bacteriocin AS-48 and Antibiotics against Uropathogenic Enterococci. Antibiotics 2020, 9, 567. [Google Scholar] [CrossRef]
- Proença, D.; Fernandes, S.; Leandro, C.; Silva, F.A.; Santos, S.; Lopes, F.; Mato, R.; Cavaco-Silva, P.; Pimentel, M.; São-José, C. Phage Endolysins with Broad Antimicrobial Activity against Enterococcus faecalis Clinical Strains. Microb. Drug Resist. 2012, 18, 322–332. [Google Scholar] [CrossRef]
- Zhang, H.; Buttaro, B.A.; Fouts, D.E.; Sanjari, S.; Evans, B.S.; Stevens, R.H. Bacteriophage ΦEf11 ORF28 Endolysin, a Multifunctional Lytic Enzyme with Properties Distinct from All Other Identified Enterococcus faecalis Phage Endolysins. Appl. Environ. Microbiol. 2019, 85, e00555-19. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Jin, J.-S.; Choi, Y.-J.; Kim, J. LysSAP26, a New Recombinant Phage Endolysin with a Broad Spectrum Antibacterial Activity. Viruses 2020, 12, 1340. [Google Scholar] [CrossRef]
- Zhang, H.; Stevens, R.H. Intrinsic Resistance of Enterococcus faecalis Strains to ΦEf11 Phage Endolysin Is Associated with the Presence of ΦEf11 Prophage. Arch. Virol. 2020, 166, 249–258. [Google Scholar] [CrossRef] [PubMed]
- Głowacka-Rutkowska, A.; Ulatowska, M.; Empel, J.; Kowalczyk, M.; Boreczek, J.; Łobocka, M. A Kayvirus Distant Homolog of Staphylococcal Virulence Determinants and VISA Biomarker Is a Phage Lytic Enzyme. Viruses 2020, 12, 292. [Google Scholar] [CrossRef] [PubMed]
- Biswas, B.; Adhya, S.; Washart, P.; Paul, B.; Trostel, A.N.; Powell, B.; Carlton, R.; Merril, C.R. Bacteriophage Therapy Rescues Mice Bacteremic from a Clinical Isolate of Vancomycin-Resistant Enterococcus faecium. Infect. Immun. 2002, 70, 204–210. [Google Scholar] [CrossRef] [PubMed]
- Khalifa, L.; Shlezinger, M.; Beyth, S.; Houri-Haddad, Y.; Coppenhagen-Glazer, S.; Beyth, N.; Hazan, R. Phage Therapy against Enterococcus faecalis in Dental Root Canals. J. Oral Microbiol. 2016, 8, 32157. [Google Scholar] [CrossRef]
- Gelman, D.; Beyth, S.; Lerer, V.; Adler, K.; Poradosu-Cohen, R.; Coppenhagen-Glazer, S.; Hazan, R. Combined Bacteriophages and Antibiotics as an Efficient Therapy against VRE Enterococcus faecalis in a Mouse Model. Res. Microbiol. 2018, 169, 531–539. [Google Scholar] [CrossRef] [PubMed]
- Khalifa, L.; Gelman, D.; Shlezinger, M.; Dessal, A.L.; Coppenhagen-Glazer, S.; Beyth, N.; Hazan, R. Defeating Antibiotic- and Phage-Resistant Enterococcus faecalis Using a Phage Cocktail in Vitro and in a Clot Model. Front. Microbiol. 2018, 9, 326. [Google Scholar] [CrossRef] [PubMed]
- Shlezinger, M.; Coppenhagen-Glazer, S.; Gelman, D.; Beyth, N.; Hazan, R. Eradication of Vancomycin-Resistant Enterococci by Combining Phage and Vancomycin. Viruses 2019, 11, 954. [Google Scholar] [CrossRef] [PubMed]
- Górski, A.; Międzybrodzki, R.; Łobocka, M.; Głowacka-Rutkowska, A.; Bednarek, A.; Borysowski, J.; Jończyk-Matysiak, E.; Łusiak-Szelachowska, M.; Weber-Dąbrowska, B.; Bagińska, N.; et al. Phage Therapy: What Have We Learned? Viruses 2018, 10, 288. [Google Scholar] [CrossRef]
- Pelgrift, R.Y.; Friedman, A.J. Nanotechnology as a Therapeutic Tool to Combat Microbial Resistance. Adv. Drug Deliv. Rev. 2013, 65, 1803–1815. [Google Scholar] [CrossRef]
- Friedman, A.J.; Phan, J.; Schairer, D.O.; Champer, J.; Qin, M.; Pirouz, A.; Blecher-Paz, K.; Oren, A.; Liu, P.T.; Modlin, R.L.; et al. Antimicrobial and Anti-Inflammatory Activity of Chitosan-Alginate Nanoparticles: A Targeted Therapy for Cutaneous Pathogens. J. Investig. Dermatol. 2013, 133, 1231–1239. [Google Scholar] [CrossRef]
- Li, J.; Zhang, K.; Ruan, L.; Chin, S.F.; Wickramasinghe, N.; Liu, H.; Ravikumar, V.; Ren, J.; Duan, H.; Yang, L.; et al. Block Copolymer Nanoparticles Remove Biofilms of Drug-Resistant Gram-Positive Bacteria by Nanoscale Bacterial Debridement. Nano Lett. 2018, 18, 4180–4187. [Google Scholar] [CrossRef]
- Zhen, J.-B.; Zhao, M.-H.; Ge, Y.; Liu, Y.; Xu, L.-W.; Chen, C.; Gong, Y.-K.; Yang, K.-W. Construction, Mechanism, and Antibacterial Resistance Insight into Polypeptide-Based Nanoparticles. Biomater. Sci. 2019, 7, 4142–4152. [Google Scholar] [CrossRef]
- Wilson, D.N. Ribosome-Targeting Antibiotics and Mechanisms of Bacterial Resistance. Nat. Rev. Microbiol. 2014, 12, 35–48. [Google Scholar] [CrossRef]
- Mühlberg, E.; Umstätter, F.; Kleist, C.; Domhan, C.; Mier, W.; Uhl, P. Renaissance of Vancomycin: Approaches for Breaking Antibiotic Resistance in Multidrug-Resistant Bacteria. Can. J. Microbiol. 2020, 66, 11–16. [Google Scholar] [CrossRef]
- Mishra, N.M.; Stolarzewicz, I.; Cannaerts, D.; Schuermans, J.; Lavigne, R.; Looz, Y.; Landuyt, B.; Schoofs, L.; Schols, D.; Paeshuyse, J.; et al. Iterative Chemical Engineering of Vancomycin Leads to Novel Vancomycin Analogs with a High in Vitro Therapeutic Index. Front. Microbiol. 2018, 9, 1175. [Google Scholar] [CrossRef]
- Yoshida, O.; Yasukata, T.; Sumino, Y.; Munekage, T.; Narukawa, Y.; Nishitani, Y. Novel Semi-Synthetic Glycopeptide Antibiotics Active against Methicillin-Resistant Staphylococcus aureus (MRSA) and Vancomycin-Resistant Enterococci (VRE): Doubly-Modified Water-Soluble Derivatives of Chloroorienticin B. Bioorg. Med. Chem. Lett. 2002, 12, 3027–3031. [Google Scholar] [CrossRef]
- James, R.C.; Pierce, J.G.; Okano, A.; Xie, J.; Boger, D.L. Redesign of Glycopeptide Antibiotics—Back to the Future. ACS Chem. Biol. 2012, 7, 797–804. [Google Scholar] [CrossRef]
- Ma, N.; Jia, Y.; Liu, Z.; Gonzalez-Zamora, E.; Bois-Choussy, M.; Malabarba, A.; Brunati, C.; Zhu, J. Design and Synthesis of Macrocycles Active against Vancomycin-Resistant Enterococci (VRE): The Interplay between d-Ala-d-Lac Binding and Hydrophobic Effect. Bioorg. Med. Chem. Lett. 2005, 15, 743–746. [Google Scholar] [CrossRef] [PubMed]
- Plotkin, P.; Patel, K.; Uminski, A.; Marzella, N. Telavancin (Vibativ), a New Option for the Treatment of Gram-Positive Infections. Pharm. Ther. 2011, 36, 127–138. [Google Scholar]
- Zhang, J.; Keller, K.; Takemoto, J.Y.; Bensaci, M.; Litke, A.; Czyryca, P.G.; Chang, C.-W.T. Synthesis and Combinational Antibacterial Study of 5″-Modified Neomycin. J. Antibiot. 2009, 62, 539–544. [Google Scholar] [CrossRef] [PubMed]
- Mwangi, J.; Hao, X.; Lai, R.; Zhang, Z.-Y. Antimicrobial Peptides: New Hope in the War against Multidrug Resistance. Zool. Res. 2019, 40, 488–505. [Google Scholar] [CrossRef] [PubMed]
- Fedders, H.; Podschun, R.; Leippe, M. The Antimicrobial Peptide Ci-MAM-A24 Is Highly Active against Multidrug-Resistant and Anaerobic Bacteria Pathogenic for Humans. Int. J. Antimicrob. Agents 2010, 36, 264–266. [Google Scholar] [CrossRef]
- Bechinger, B.; Gorr, S.-U. Antimicrobial Peptides. J. Dent. Res. 2017, 96, 254–260. [Google Scholar] [CrossRef]
- Kumariya, R.; Garsa, A.K.; Rajput, Y.S.; Sood, S.K.; Akhtar, N.; Patel, S. Bacteriocins: Classification, Synthesis, Mechanism of Action and Resistance Development in Food Spoilage Causing Bacteria. Microb. Pathog. 2019, 128, 171–177. [Google Scholar] [CrossRef] [PubMed]
- Singh, M.; Chaudhary, S.; Sareen, D. Roseocin, a Novel Two-component Lantibiotic from an Actinomycete. Mol. Microbiol. 2020, 113, 326–337. [Google Scholar] [CrossRef] [PubMed]
- Shokri, D.; Zaghian, S.; Khodabakhsh, F.; Fazeli, H.; Mobasherizadeh, S.; Ataei, B. Antimicrobial Activity of a UV-Stable Bacteriocin-like Inhibitory Substance (BLIS) Produced by Enterococcus faecium Strain DSH20 against Vancomycin-Resistant Enterococcus (VRE) Strains. J. Microbiol. Immunol. Infect. 2014, 47, 371–376. [Google Scholar] [CrossRef] [PubMed]
- Inoue, T.; Tomita, H.; Ike, Y. Bac 32, a Novel Bacteriocin Widely Disseminated among Clinical Isolates of Enterococcus faecium. Antimicrob. Agents Chemother. 2006, 50, 1202–1212. [Google Scholar] [CrossRef] [PubMed]
- Todokoro, D.; Tomita, H.; Inoue, T.; Ike, Y. Genetic Analysis of Bacteriocin 43 of Vancomycin-Resistant Enterococcus faecium. Appl. Environ. Microbiol. 2006, 72, 6955–6964. [Google Scholar] [CrossRef] [PubMed]
- Basi-Chipalu, S.; Dischinger, J.; Josten, M.; Szekat, C.; Zweynert, A.; Sahl, H.-G.; Bierbaum, G. Pseudomycoicidin, a Class II Lantibiotic from Bacillus pseudomycoides. Appl. Environ. Microbiol. 2015, 81, 3419–3429. [Google Scholar] [CrossRef]
- Li, Y.; Ducasse, R.; Zirah, S.; Blond, A.; Goulard, C.; Lescop, E.; Giraud, C.; Hartke, A.; Guittet, E.; Pernodet, J.-L.; et al. Characterization of Sviceucin from Streptomyces Provides Insight into Enzyme Exchangeability and Disulfide Bond Formation in Lasso Peptides. ACS Chem. Biol. 2015, 10, 2641–2649. [Google Scholar] [CrossRef]
- Hegemann, J.D.; Zimmermann, M.; Xie, X.; Marahiel, M.A. Lasso Peptides: An Intriguing Class of Bacterial Natural Products. Acc. Chem. Res. 2015, 48, 1909–1919. [Google Scholar] [CrossRef]
- Nir-Paz, R.; Gelman, D.; Khouri, A.; Sisson, B.M.; Fackler, J.; Alkalay-Oren, S.; Khalifa, L.; Rimon, A.; Yerushalmy, O.; Bader, R.; et al. Successful Treatment of Antibiotic-Resistant, Poly-Microbial Bone Infection with Bacteriophages and Antibiotics Combination. Clin. Infect. Dis. 2019, 69, 2015–2018. [Google Scholar] [CrossRef]
- Leitner, L.; Sybesma, W.; Chanishvili, N.; Goderdzishvili, M.; Chkhotua, A.; Ujmajuridze, A.; Schneider, M.P.; Sartori, A.; Mehnert, U.; Bachmann, L.M.; et al. Bacteriophages for Treating Urinary Tract Infections in Patients Undergoing Transurethral Resection of the Prostate: A Randomized, Placebo-Controlled, Double-Blind Clinical Trial. BMC Urol. 2017, 17, 90. [Google Scholar] [CrossRef]
- Sarker, S.A.; Sultana, S.; Reuteler, G.; Moine, D.; Descombes, P.; Charton, F.; Bourdin, G.; McCallin, S.; Ngom-Bru, C.; Neville, T.; et al. Oral Phage Therapy of Acute Bacterial Diarrhea with Two Coliphage Preparations: A Randomized Trial in Children From Bangladesh. EBioMedicine 2016, 4, 124–137. [Google Scholar] [CrossRef]
- Otawa, K.; Hirakata, Y.; Kaku, M.; Nakai, Y. Bacteriophage Control of Vancomycin-Resistant Enterococci in Cattle Compost. J. Appl. Microbiol. 2012, 113, 499–507. [Google Scholar] [CrossRef]
- Barros, J.; Melo, L.D.R.; Poeta, P.; Igrejas, G.; Ferraz, M.P.; Azeredo, J.; Monteiro, F.J. Lytic Bacteriophages against Multidrug-Resistant Staphylococcus aureus, Enterococcus faecalis and Escherichia coli Isolates from Orthopaedic Implant-Associated Infections. Int. J. Antimicrob. Agents 2019, 54, 329–337. [Google Scholar] [CrossRef]
- Uchiyama, J.; Rashel, M.; Maeda, Y.; Takemura, I.; Sugihara, S.; Akechi, K.; Muraoka, A.; Wakiguchi, H.; Matsuzaki, S. Isolation and Characterization of a Novel Enterococcus faecalis Bacteriophage ΦEF24C as a Therapeutic Candidate. FEMS Microbiol. Lett. 2008, 278, 200–206. [Google Scholar] [CrossRef]
- Sofy, A.R.; Abd El Haliem, N.F.; Refaey, E.E.; Hmed, A.A. Polyvalent Phage CoNShP-3 as a Natural Antimicrobial Agent Showing Lytic and Antibiofilm Activities against Antibiotic-Resistant Coagulase-Negative Staphylococci Strains. Foods 2020, 9, 673. [Google Scholar] [CrossRef]
- Altamirano, F.L.G.; Barr, J.J. Phage Therapy in the Postantibiotic Era. Clin. Microbiol. Rev. 2019, 32, e00066-18. [Google Scholar] [CrossRef]
- Thierauf, A.; Perez, G.; Maloy, S. Generalized Transduction. In Bacteriophages; Clokie, M.R.J., Kropinski, A.M., Eds.; Methods in Molecular Biology; Humana Press: Totowa, NJ, USA, 2009; Volume 501, pp. 267–286. ISBN 978-1-58829-682-5. [Google Scholar]
- Mattila, S.; Ruotsalainen, P.; Jalasvuori, M. On-Demand Isolation of Bacteriophages Against Drug-Resistant Bacteria for Personalized Phage Therapy. Front. Microbiol. 2015, 6, 1271. [Google Scholar] [CrossRef] [PubMed]
- Kortright, K.E.; Chan, B.K.; Koff, J.L.; Turner, P.E. Phage Therapy: A Renewed Approach to Combat Antibiotic-Resistant Bacteria. Cell Host Microbe 2019, 25, 219–232. [Google Scholar] [CrossRef] [PubMed]
- Lourenço, M.; Chaffringeon, L.; Lamy-Besnier, Q.; Pédron, T.; Campagne, P.; Eberl, C.; Bérard, M.; Stecher, B.; Debarbieux, L.; De Sordi, L. The Spatial Heterogeneity of the Gut Limits Predation and Fosters Coexistence of Bacteria and Bacteriophages. Cell Host Microbe 2020, 28, 390–401.e5. [Google Scholar] [CrossRef] [PubMed]
- Fabijan, A.P.; Lin, R.C.Y.; Ho, J.; Maddocks, S.; Zakour, N.L.B.; Iredell, J.R.; Westmead Bacteriophage Therapy Team; Khalid, A.; Venturini, C.; Chard, R.; et al. Safety of Bacteriophage Therapy in Severe Staphylococcus aureus Infection. Nat. Microbiol. 2020, 5, 465–472. [Google Scholar] [CrossRef]
- Cisek, A.A.; Dąbrowska, I.; Gregorczyk, K.P.; Wyżewski, Z. Phage Therapy in Bacterial Infections Treatment: One Hundred Years After the Discovery of Bacteriophages. Curr. Microbiol. 2017, 74, 277–283. [Google Scholar] [CrossRef]
- Kilcher, S.; Studer, P.; Muessner, C.; Klumpp, J.; Loessner, M.J. Cross-Genus Rebooting of Custom-Made, Synthetic Bacteriophage Genomes in L-Form Bacteria. Proc. Natl. Acad. Sci. USA 2018, 115, 567–572. [Google Scholar] [CrossRef] [PubMed]
- Asghar, M.A.; Yousuf, R.I.; Shoaib, M.H.; Asghar, M.A. Antibacterial, Anticoagulant and Cytotoxic Evaluation of Biocompatible Nanocomposite of Chitosan Loaded Green Synthesized Bioinspired Silver Nanoparticles. Int. J. Biol. Macromol. 2020, 160, 934–943. [Google Scholar] [CrossRef] [PubMed]
- Gordon, O.; Slenters, T.V.; Brunetto, P.S.; Villaruz, A.E.; Sturdevant, D.E.; Otto, M.; Landmann, R.; Fromm, K.M. Silver Coordination Polymers for Prevention of Implant Infection: Thiol Interaction, Impact on Respiratory Chain Enzymes, and Hydroxyl Radical Induction. Antimicrob. Agents Chemother. 2010, 54, 4208–4218. [Google Scholar] [CrossRef] [PubMed]
- Lara, H.H.; Ayala-Núñez, N.V.; del Turrent, L.I.C.; Padilla, C.R. Bactericidal Effect of Silver Nanoparticles against Multidrug-Resistant Bacteria. World J. Microbiol. Biotechnol. 2010, 26, 615–621. [Google Scholar] [CrossRef]
- AlSalhi, M.S.; Devanesan, S.; Atif, M.; AlQahtani, W.S.; Nicoletti, M.; Serrone, P.D. Therapeutic Potential Assessment of Green Synthesized Zinc Oxide Nanoparticles Derived from Fennel Seeds Extract. Int. J. Nanomed. 2020, 15, 8045–8057. [Google Scholar] [CrossRef]
- Punjabi, K.; Mehta, S.; Chavan, R.; Chitalia, V.; Deogharkar, D.; Deshpande, S. Efficiency of Biosynthesized Silver and Zinc Nanoparticles Against Multi-Drug Resistant Pathogens. Front. Microbiol. 2018, 9, 2207. [Google Scholar] [CrossRef]
- Iram, S.; Akbar Khan, J.; Aman, N.; Nadhman, A.; Zulfiqar, Z.; Yameen, M.A. Enhancing the Anti-Enterococci Activity of Different Antibiotics by Combining with Metal Oxide Nanoparticles. Jundishapur J. Microbiol. 2016, 9, e31302. [Google Scholar] [CrossRef] [PubMed]
- Lai, H.-Z.; Chen, W.-Y.; Wu, C.-Y.; Chen, Y.-C. Potent Antibacterial Nanoparticles for Pathogenic Bacteria. ACS Appl. Mater. Interfaces 2015, 7, 2046–2054. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.-G.; Chen, Y.-C.; Chen, Y.-C. Antibacterial Gold Nanoparticle-Based Photothermal Killing of Vancomycin-Resistant Bacteria. Nanomedicine 2018, 13, 1405–1416. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, S.P.; Sahu, S.K.; Mahapatra, S.K.; Santra, S.; Bal, M.; Roy, S.; Pramanik, P. Nanoconjugated Vancomycin: New Opportunities for the Development of Anti-VRSA Agents. Nanotechnology 2010, 21, 105103. [Google Scholar] [CrossRef] [PubMed]
- Honary, S.; Ebrahimi, P.; Hadianamrei, R. Optimization of Particle Size and Encapsulation Efficiency of Vancomycin Nanoparticles by Response Surface Methodology. Pharm. Dev. Technol. 2014, 19, 987–998. [Google Scholar] [CrossRef] [PubMed]
- Bäumler, A.J.; Sperandio, V. Interactions between the Microbiota and Pathogenic Bacteria in the Gut. Nature 2016, 535, 85–93. [Google Scholar] [CrossRef] [PubMed]


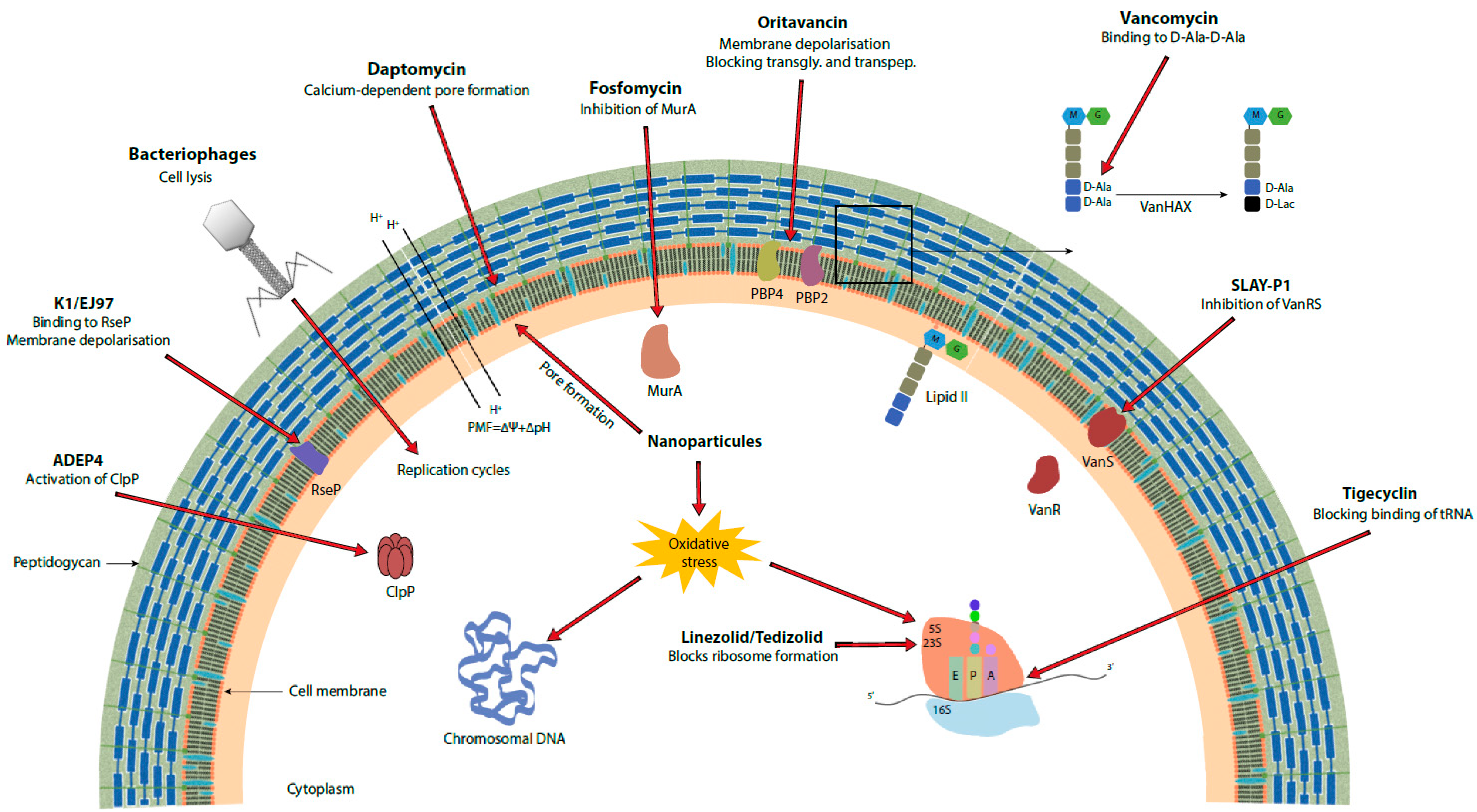{kind=link}
| Antimicrobial | Utilization | Target | Mechanism of Action | Pros | Cons | References |
|---|---|---|---|---|---|---|
| Available antibiotics | ||||||
| linezolid | clinical use | 23S ribosomal RNA | protein synthesis inhibition | - well known - industrial production | resistance | [20,21,22,23,24,25,26,28,29,30,31,32,36] |
| daptomycin | clinical use | calcium ions | membrane disruption | [20,22,29,35,37,38,39,40,41,42,43] | ||
| tigecycline | clinical use | 30S ribosomal subunit | protein synthesis inhibition | [20,44,45,46,47] | ||
| tedizolid | clinical use | 23S ribosomal RNA | protein synthesis inhibition | active against linezolid resistant bacteria (cfr gene) | [33,48] | |
| Modified antibiotics | ||||||
| vancomycin dimers | in vitro (in vivo on S. pneumoniae) [49] | Penicillin binding protein 2 | cell wall synthesis inhibition | new mechanism of action | resistance is still possible | [50,51,52,53] |
| teicoplanin derivatives | in vitro | lipid II | active on VanA phenotype | [54] | ||
| dalbavancin | clinical use | lipid II | - increasing MIC - resistance of VanA phenotype | [55,56,57,58] | ||
| oritavancin | clinical use | lipid II—membrane | - cell wall synthesis inhibition (transglycosylation and transpeptidation) - membrane disruption | 3 different mechanisms of action | resistance in vitro | [55,56,58,59,60] |
| vancomycin derivative | in vitro | lipid II (D-ala-D-ala and D-ala-D-lac)—membrane | - cell wall synthesis inhibition - membrane disruption | 3 different mechanisms of action | [61] | |
| Combination of antibiotics | ||||||
| various types and classes | clinical use | DNA/RNA/protein metabolism—membrane | multiple | - multiple mechanisms of action - overcome resistance | wide spectrum | [22,43,62,63,64,65,66,67,68,69,70,71,72,73,74] |
| Antimicrobial peptides | ||||||
| anti-vancomycin peptide | in vivo | vancomycin | vancomycin concentration decreasing | prevent the emergence of resistance | [75] | |
| SLAY-P1 | in vivo | VanRS | vancomycin resistance inhibition | - overcome resistance - stable in human serum | resistance is still possible | [76] |
| cationic peptides | in vivo | membrane | membrane disruption | - physical interactions - easy engineering | - stability in host - toxicity - production cost - resistance | [76] |
| peptides + antibiotics combination | in vivo | DNA/RNA/protein metabolism—membrane | multiple | - multiple mechanisms of action - overcome resistance - activity on biofilm | resistance is still possible | [76,77,78,79] |
| Bacteriocins | ||||||
| bacterial transplantation | in vivo | intestinal VRE | VRE elimination | resistant pathogen elimination before infection | - resistance is still possible - sensitive to proteases | [80] |
| mersacidin | in vivo | lipid II | cell wall synthesis inhibition | [81,82] | ||
| lacticin 3147 | in vivo | lipid II | membrane disruption | active against multiple pathogens of interest | [81,83] | |
| pumilicin 4 | (clinical use = B. pumilus probiotic) | unknown | membrane destabilization | heat resistant | [84] | |
| K1 and EJ97 | in vitro | RseP | membrane depolarization | attenuates bacteria’s pathogenicity | narrow spectrum | [19,85] |
| EF478 | in vitro | peptidoglycan | cell wall disruption | active against MDRE | [86] | |
| bacteriocins + antibiotics combination | in vitro | DNA/RNA/protein metabolism—membrane | multiple | - multiple mechanisms of action - overcome resistance - activity on biofilm - used in food for more than 50 years - generally stable - easy engineering - non toxic | [87,88] | |
| Lysins | ||||||
| in vivo | cell wall | peptidoglycan lysis | - act quickly = less resistance - heterologous production - easy engineering | resistance | [89,90,91,92] | |
| combination lysin bacteriocin | in vitro | cell wall | peptidoglycan lysis | [93] | ||
| Bacteriophages | ||||||
| ENB6 | in vivo | cell metabolism | [94] | |||
| EFDG1/EFLK1 | in vivo | cell metabolism | - can act on biofilm - overcome resistance | [95,96,97] | ||
| combination phages antibiotics | in vivo | - cell metabolism - DNA/RNA/protein metabolism—membrane | hijack of the cell metabolism + other depending on the antibiotic | - overcome resistance - reduce antibiotics concentrations | wide spectrum | [96,98] |
| all bacteriophages | clinical use | cell metabolism | hijack of the cell metabolism | - “smart therapy” - only one administration | - host immune response - possible toxicity with toxins - narrow spectrum = isolation needed | |
| Nanoparticles | ||||||
| silver nanoparticles | in vivo | - membrane - cell metabolism | - membrane disruption - inhibition of DNA replication - ROS production | multiple mechanisms of action | possible toxicity | [99] |
| chitosan nanoparticles | in vitro | - membrane - cell metabolism | - membrane disruption (P. acnes) - transcription inhibition - metal chelation | multiple mechanisms of action | [100,101] | |
| AgNPs@PCL-b-AMPs | in vivo | - membrane - cell metabolism | - membrane disruption - ROS production | - no toxicity to eukaryotic cells - no resistance in vitro | [49] | |
| DA95B5 | in vivo | cell surface | biofilm inhibition | - no toxicity - limitation of biofilm | need another treatment to kill bacteria | [102] |
| polypeptide-based nanoparticles | in vitro | membrane | membrane disruption | - no nanoparticles resistance observed - low toxicity | [103] | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Baëtz, B.; Boudrioua, A.; Hartke, A.; Giraud, C. Alternatives to Fight Vancomycin-Resistant Staphylococci and Enterococci. Antibiotics 2021, 10, 1116. https://doi.org/10.3390/antibiotics10091116
Baëtz B, Boudrioua A, Hartke A, Giraud C. Alternatives to Fight Vancomycin-Resistant Staphylococci and Enterococci. Antibiotics. 2021; 10(9):1116. https://doi.org/10.3390/antibiotics10091116
Chicago/Turabian StyleBaëtz, Benjamin, Abdelhakim Boudrioua, Axel Hartke, and Caroline Giraud. 2021. "Alternatives to Fight Vancomycin-Resistant Staphylococci and Enterococci" Antibiotics 10, no. 9: 1116. https://doi.org/10.3390/antibiotics10091116
APA StyleBaëtz, B., Boudrioua, A., Hartke, A., & Giraud, C. (2021). Alternatives to Fight Vancomycin-Resistant Staphylococci and Enterococci. Antibiotics, 10(9), 1116. https://doi.org/10.3390/antibiotics10091116