
The Role of the Environment in Dynamics of Antibiotic Resistance in Humans and Animals: A Modelling Study

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
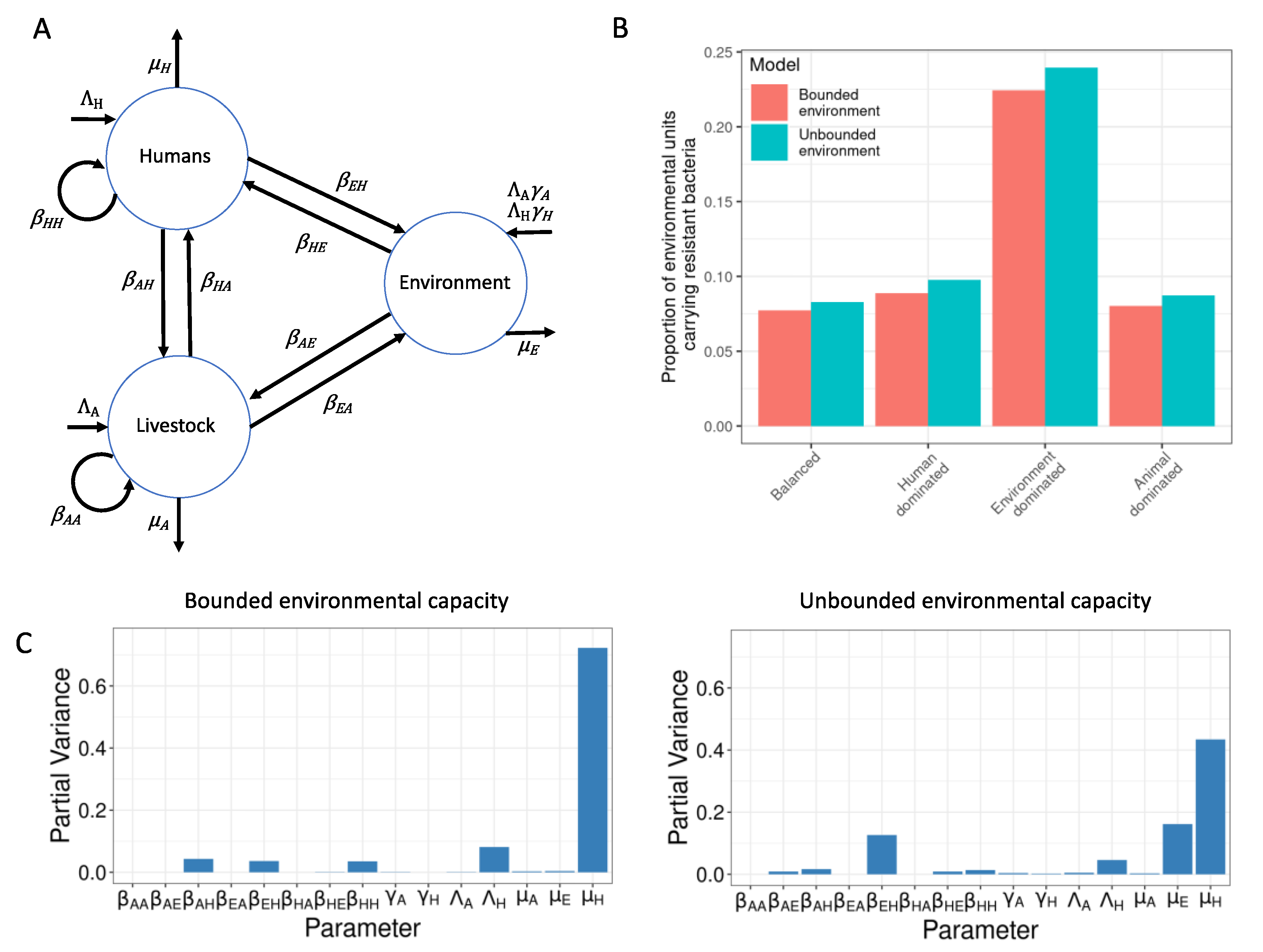
2. Results
2.1. Long-Term Dynamics of Resistance in Humans
2.1.1. Prevalence of Resistance in Humans
2.1.2. Sensitivity Analysis
2.2. Impact of Interventions to Reduce Resistance in Humans
2.2.1. Impact of Curtailing Antibiotic Usage in Animals
2.2.2. Reducing vs. Reducing
2.2.3. Effect of on Impact of Interventions to Reduce Antibiotic Consumption in Animals
3. Materials and Methods
3.1. Model Description
3.2. Capacity for Resistance in the Environment
3.3. Impact of Interventions on Resistance in Humans
3.4. Sensitivity Analysis
3.5. Parameterisation
3.6. Software
4. Discussion
4.1. Key Findings
4.2. Is Curtailing Antibiotic Usage in Animals an Effective Intervention to Reduce Human Resistance Levels?
4.3. Could the Environment Be an Effective Alternative Intervention Target?
4.4. Should the Environment Be Included in AMR Models?
4.5. Modelling the Environment Highlights Data Needs
4.6. Limitations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Woolhouse, M.; Ward, M.; van Bunnik, B.; Farrar, J. Antimicrobial Resistance in Humans, Livestock and the Wider Environment. Philos. Trans. R. Soc. B Biol. Sci. 2015, 370, 20140083. [Google Scholar] [CrossRef] [PubMed]
- Robinson, T.P.; Bu, D.P.; Carrique-Mas, J.; Fèvre, E.M.; Gilbert, M.; Grace, D.; Hay, S.I.; Jiwakanon, J.; Kakkar, M.; Kariuki, S.; et al. Antibiotic Resistance Is the Quintessential One Health Issue. Trans. R. Soc. Trop. Med. Hyg. 2016, 110, 377–380. [Google Scholar] [CrossRef] [Green Version]
- Collignon, P.J.; McEwen, S.A. One Health-Its Importance in Helping to Better Control Antimicrobial Resistance. Trop. Med. Infect. Dis. 2019, 4, 22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Bi, Z.; Ma, S.; Chen, B.; Cai, C.; He, J.; Schwarz, S.; Sun, C.; Zhou, Y.; Yin, J.; et al. Inter-Host Transmission of Carbapenemase-Producing Escherichia Coli among Humans and Backyard Animals. Environ. Health Perspect. 2019, 127, 107009. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singer, R.S.; Cox, L.A.; Dickson, J.S.; Hurd, H.S.; Phillips, I.; Miller, G.Y. Modeling the Relationship between Food Animal Health and Human Foodborne Illness. Prev. Vet. Med. 2007, 79, 186–203. [Google Scholar] [CrossRef]
- Larsson, D.G.J.; Andremont, A.; Bengtsson-Palme, J.; Brandt, K.K.; de Roda Husman, A.M.; Fagerstedt, P.; Fick, J.; Flach, C.F.; Gaze, W.H.; Kuroda, M.; et al. Critical Knowledge Gaps and Research Needs Related to the Environmental Dimensions of Antibiotic Resistance. Environ. Int. 2018, 117, 132–138. [Google Scholar] [CrossRef] [PubMed]
- Bürgmann, H.; Frigon, D.; Gaze, W.H.; Manaia, C.M.; Pruden, A.; Singer, A.C.; Smets, B.F.; Zhang, T. Water and Sanitation: An Essential Battlefront in the War on Antimicrobial Resistance. FEMS Microbiol. Ecol. 2018, 94, fiy-101. [Google Scholar] [CrossRef] [PubMed]
- Manaia, C.M. Antibiotic Resistance in Wastewater: Origins, Fate, and Risks. Prävention Gesundh. 2014, 9, 180–184. [Google Scholar] [CrossRef]
- Amos, G.C.A.; Gozzard, E.; Carter, C.E.; Mead, A.; Bowes, M.J.; Hawkey, P.M.; Zhang, L.; Singer, A.C.; Gaze, W.H.; Wellington, E.M.H. Validated Predictive Modelling of the Environmental Resistome. ISME J. 2015, 9, 1467–1476. [Google Scholar] [CrossRef] [Green Version]
- Leonard, A.F.C.; Zhang, L.; Balfour, A.J.; Garside, R.; Gaze, W.H. Human Recreational Exposure to Antibiotic Resistant Bacteria in Coastal Bathing Waters. Environ. Int. 2015, 82, 92–100. [Google Scholar] [CrossRef] [PubMed]
- Palacios, O.A.; Contreras, C.A.; Muñoz-Castellanos, L.N.; González-Rangel, M.O.; Rubio-Arias, H.; Palacios-Espinosa, A.; Nevárez-Moorillón, G.V. Monitoring of Indicator and Multidrug Resistant Bacteria in Agricultural Soils under Different Irrigation Patterns. Agric. Water Manag. 2017, 184, 19–27. [Google Scholar] [CrossRef]
- Araujo, S.; Henriques, I.S.; Leandro, S.M.; Alves, A.; Pereira, A.; Correia, A. Gulls Identified as Major Source of Fecal Pollution in Coastal Waters: A Microbial Source Tracking Study. Sci. Total Environ. 2014, 470, 84–91. [Google Scholar] [CrossRef] [PubMed]
- Swift, B.; Bennett, M.; Waller, K.; Dodd, C.; Murray, A.; Gomes, R.; Humphreys, B.; Hobman, J.; Jones, M.; Whitlock, S.; et al. Anthropogenic Environmental Drivers of Antimicrobial Resistance in Wildlife. Sci. Total Environ. 2019, 649, 12–20. [Google Scholar] [CrossRef]
- Call, D.R.; Matthews, L.; Subbiah, M.; Liu, J. Do Antibiotic Residues in Soils Play a Role in Amplification and Transmission of Antibiotic Resistant Bacteria in Cattle Populations? Front. Microbiol. 2013, 4, 193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cabello, F.C.; Godfrey, H.P.; Tomova, A.; Ivanova, L.; Dölz, H.; Millanao, A.; Buschmann, A.H. Antimicrobial Use in Aquaculture Re-Examined: Its Relevance to Antimicrobial Resistance and to Animal and Human Health. Environ. Microbiol. 2013, 15, 1917–1942. [Google Scholar] [CrossRef]
- Raven, K.E.; Ludden, C.; Gouliouris, T.; Blane, B.; Naydenova, P.; Brown, N.M.; Parkhill, J.; Peacock, S.J. Genomic Surveillance of Escherichia Coli in Municipal Wastewater Treatment Plants as an Indicator of Clinically Relevant Pathogens and Their Resistance Genes. Microb. Genom. 2019, 5, e000267. [Google Scholar] [CrossRef] [PubMed]
- Knight, G.M.; Davies, N.G.; Colijn, C.; Coll, F.; Donker, T.; Gifford, D.R.; Glover, R.E.; Jit, M.; Klemm, E.; Lehtinen, S.; et al. Mathematical Modelling for Antibiotic Resistance Control Policy: Do We Know Enough? BMC Infect. Dis. 2019, 19, 1011. [Google Scholar] [CrossRef] [PubMed]
- Niewiadomska, A.M.; Jayabalasingham, B.; Seidman, J.C.; Willem, L.; Grenfell, B.; Spiro, D.; Viboud, C. Population-Level Mathematical Modeling of Antimicrobial Resistance: A Systematic Review. BMC Med. 2019, 17, 81. [Google Scholar] [CrossRef] [Green Version]
- McBryde, E.S.; McElwain, D.L.S. A Mathematical Model Investigating the Impact of an Environmental Reservoir on the Prevalence and Control of Vancomycin- Resistant Enterococci. J. Infect. Dis. 2006, 193, 1473–1474. [Google Scholar] [CrossRef] [PubMed]
- Kouyos, R.D.; zur Wiesch, P.A.; Bonhoeffer, S. On Being the Right Size: The Impact of Population Size and Stochastic Effects on the Evolution of Drug Resistance in Hospitals and the Community. PLoS Pathog. 2011, 7, e1001334. [Google Scholar] [CrossRef] [PubMed]
- Græsbøll, K.; Nielsen, S.S.; Toft, N.; Christiansen, L.E. How Fitness Reduced, Antimicrobial Resistant Bacteria Survive and Spread: A Multiple Pig-Multiple Bacterial Strain Model. PLoS ONE 2014, 9, e100458. [Google Scholar] [CrossRef] [PubMed]
- Booton, R.D.; Meeyai, A.; Alhusein, N.; Buller, H.; Feil, E.; Lambert, H.; Mongkolsuk, S.; Pitchforth, E.; Reyher, K.K.; Sakcamduang, W.; et al. One Health Drivers of Antibacterial Resistance: Quantifying the Relative Impacts of Human, Animal and Environmental Use and Transmission. One Health 2021, 12, 100220. [Google Scholar] [CrossRef] [PubMed]
- WHO Regional Office for Europe/European Centre for Disease Prevention and Control. Antimicrobial Resistance Surveillance in Europe 2022–2020 Data; WHO Regional Office for Europe: Copenhagen, Denmark, 2022. [Google Scholar]
- van Bunnik, B.A.D.; Woolhouse, M.E.J. Modelling the Impact of Curtailing Antibiotic Usage in Food Animals on Antibiotic Resistance in Humans. R. Soc. Open Sci. 2017, 4, 161067. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saltelli, A.; Tarantola, S.; Chan, K.P.-S. A Quantitative Model-Independent Method for Global Sensitivity Analysis of Model Output. Technometrics 1999, 41, 39–56. [Google Scholar] [CrossRef]
- Reusser, D. fast: Implementation of the Fourier Amplitude Sensitivity Test (FAST). Available online: https://github.com/cran/fast (accessed on 1 October 2018).
- Muloi, D.M.; Wee, B.A.; McClean, D.M.H.; Ward, M.J.; Pankhurst, L.; Phan, H.; Ivens, A.C.; Kivali, V.; Kiyong’a, A.; Ndinda, C.; et al. Population Genomics of Escherichia Coli in Livestock-Keeping Households across a Rapidly Developing Urban Landscape. Nat. Microbiol. 2022, 7, 581–589. [Google Scholar] [CrossRef] [PubMed]
- Thorpe, H.; Booton, R.; Kallonen, T.; Gibbon, M.J.; Couto, N.; Passet, V.; Sebastian Lopez Fernandez, J.; Rodrigues, C.; Matthews, L.; Mitchell, S.; et al. One Health or Three? Transmission Modelling of Klebsiella Isolates Reveals Ecological Barriers to Transmission between Humans, Animals and the Environment. bioRxiv 2021. [Google Scholar] [CrossRef]
- Laxminarayan, R.; Van Boeckel, T.; Teillant, A. The Economic Costs of Withdrawing Antimicrobial Growth Promoters from the Livestock Sector. OECD Food Agric. Fish. Pap. 2015. [Google Scholar] [CrossRef]
- Kennedy, K.; Collignon, P. Colonisation with Escherichia Coli Resistant to “Critically Important” Antibiotics: A High Risk for International Travellers. Eur. J. Clin. Microbiol. Infect. Dis. 2010, 29, 1501–1506. [Google Scholar] [CrossRef] [PubMed]
- Jørgensen, S.B.; Søraas, A.; Sundsfjord, A.; Liestøl, K.; Leegaard, T.M.; Jenum, P.A. Fecal Carriage of Extended Spectrum β- Lactamase Producing Escherichia Coli & Klebsiella Pneumoniae after Urinary Tract Infection ± A Three Year Prospective Cohort Study. PLoS ONE 2017, 12, e0173510. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez, T.d.B.; Marcato, F.; Costa, E.d.F.; van den Brand, H.; Hoorweg, F.A.; Wolthuis-Fillerup, M.; Engel, B.; Schnabel, S.K.; van Reenen, C.G. Longitudinal Study on the Prevalence of Extended Spectrum Cephalosporins-Resistant Escherichia Coli Colonization in Dutch Veal Farms. Vet. Microbiol. 2022, 273, 109520. [Google Scholar] [CrossRef]
- Moor, J.; Aebi, S.; Rickli, S.; Mostacci, N.; Overesch, G.; Oppliger, A.; Hilty, M. Dynamics of Extended-Spectrum Cephalosporin-Resistant Escherichia Coli in Pig Farms: A Longitudinal Study. Int. J. Antimicrob. Agents 2021, 58, 106382. [Google Scholar] [CrossRef] [PubMed]
- Widiasih, D.A.; Ido, N.; Omoe, K.; Sugii, S.; Shinagawa, K. Duration and Magnitude of Faecal Shedding of Shiga Toxin-Producing Escherichia Coli from Naturally Infected Cattle. Epidemiol. Infect. 2004, 132, 67–75. [Google Scholar] [CrossRef] [PubMed]
- Sharma, R.; Larney, F.J.; Chen, J.; Yanke, L.J.; Morrison, M.; Topp, E.; McAllister, T.A.; Yu, Z. Selected Antimicrobial Resistance during Composting of Manure from Cattle Administered Sub-Therapeutic Antimicrobials. J. Environ. Qual. 2009, 38, 567–575. [Google Scholar] [CrossRef]
- Mathematica. Version 11.3. Wolfram Research, Inc.: Champaign, IL, USA, 2018.
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022. [Google Scholar]
- Bezanson, J.; Edelman, A.; Karpinski, S.; Shah, V.B. Julia: A Fresh Approach to Numerical Computing. SIAM Rev. 2017, 59, 65–98. [Google Scholar] [CrossRef] [Green Version]
- Scott, A.M.; Beller, E.; Glasziou, P.; Clark, J.; Ranakusuma, R.W.; Byambasuren, O.; Bakhit, M.; Page, S.W.; Trott, D.; Mar, C.D. Is Antimicrobial Administration to Food Animals a Direct Threat to Human Health? A Rapid Systematic Review. Int. J. Antimicrob. Agents 2018, 52, 316–323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dutil, L.; Irwin, R.; Finley, R.; Ng, L.K.; Avery, B.; Boerlin, P.; Bourgault, A.M.; Cole, L.; Daignault, D.; Desruisseau, A.; et al. Ceftiofur Resistance in Salmonella Enterica Serovar Heidelberg from Chicken Meat and Humans, Canada. Emerg. Infect. Dis. 2010, 16, 48–54. [Google Scholar] [CrossRef]
- Mughini-Gras, L.; Dorado-García, A.; van Duijkeren, E.; van den Bunt, G.; Dierikx, C.M.; Bonten, M.J.M.; Bootsma, M.C.J.; Schmitt, H.; Hald, T.; Evers, E.G.; et al. Attributable Sources of Community-Acquired Carriage of Escherichia Coli Containing β-Lactam Antibiotic Resistance Genes: A Population-Based Modelling Study. Lancet Planet. Health 2019, 3, e357–e369. [Google Scholar] [CrossRef] [Green Version]
- Muloi, D.; Ward, M.J.; Pedersen, A.B.; Fèvre, E.M.; Woolhouse, M.E.J.; van Bunnik, B.A.D. Are Food Animals Responsible for Transfer of Antimicrobial-Resistant Escherichia Coli or Their Resistance Determinants to Human Populations? A Systematic Review. Foodborne Pathog. Dis. 2018, 15, 467–474. [Google Scholar] [CrossRef] [Green Version]
- Wee, B.A.; Muloi, D.M.; van Bunnik, B.A.D. Quantifying the Transmission of Antimicrobial Resistance at the Human and Livestock Interface with Genomics. Clin. Microbiol. Infect. 2020, 26, 1612–1616. [Google Scholar] [CrossRef]
- Pehrsson, E.C.; Tsukayama, P.; Patel, S.; Mejia-Bautista, M.; Sosa-Soto, G.; Navarrete, K.M.; Calderon, M.; Cabrera, L.; Hoyos-Arango, W.; Bertoli, M.T.; et al. Interconnected Microbiomes and Resistomes in Low-Income Human Habitats. Nature 2016, 533, 212–216. [Google Scholar] [CrossRef]
- Capaldi, A.; Behrend, S.; Berman, B.; Smith, J.; Wright, J.; Lloyd, A.L. Parameter Estimation and Uncertainty Quantification for an Epidemic Model. Math. Biosci. Eng. 2012, 9, 553–576. [Google Scholar] [CrossRef] [PubMed]
- Collignon, P.; Beggs, J.J.; Walsh, T.R.; Gandra, S.; Laxminarayan, R. Anthropological and Socioeconomic Factors Contributing to Global Antimicrobial Resistance: A Univariate and Multivariable Analysis. Lancet Planet. Health 2018, 2, e398–e405. [Google Scholar] [CrossRef]
- Nadimpalli, M.L.; Marks, S.J.; Montealegre, M.C.; Gilman, R.H.; Pajuelo, M.J.; Saito, M.; Tsukayama, P.; Njenga, S.M.; Kiiru, J.; Swarthout, J.; et al. Urban Informal Settlements as Hotspots of Antimicrobial Resistance and the Need to Curb Environmental Transmission. Nat. Microbiol. 2020, 5, 787–795. [Google Scholar] [CrossRef] [PubMed]
- Van Bunnik, B.A.D.; Ssematimba, A.; Hagenaars, T.J.; Nodelijk, G.; Haverkate, M.R.; Marc, M.J.; Hayden, M.K.; Weinstein, R.A.; Bootsma, M.C.J.; De Jong, M.C.M. Small Distances Can Keep Bacteria at Bay for Days. Proc. Natl. Acad. Sci. USA 2014, 111, 3556–3560. [Google Scholar] [CrossRef] [Green Version]
- van der Zwet, W.C.; Nijsen, I.E.J.; Jamin, C.; van Alphen, L.B.; von Wintersdorff, C.J.H.; Demandt, A.M.P.; Savelkoul, P.H.M. Role of the Environment in Transmission of Gram-Negative Bacteria in Two Consecutive Outbreaks in a Haematology-Oncology Department. Infect. Prev. Pract. 2022, 4, 100209. [Google Scholar] [CrossRef] [PubMed]
- Berendonk, T.U.; Manaia, C.M.; Merlin, C.; Fatta-Kassinos, D.; Cytryn, E.; Walsh, F.; Buergmann, H.; Sorum, H.; Norstrom, M.; Pons, M.-N.; et al. Tackling Antibiotic Resistance: The Environmental Framework. Nat. Rev. Microbiol. 2015, 13, 310–317. [Google Scholar] [CrossRef]
- Shenoy, E.S.; Paras, M.L.; Noubary, F.; Walensky, R.P.; Hooper, D.C. Natural History of Colonization with Methicillin-Resistant Staphylococcus Aureus (MRSA) and Vancomycin-Resistant Enterococcus (VRE): A Systematic Review. BMC Infect. Dis. 2014, 14, 177. [Google Scholar] [CrossRef]
- Bengtsson-Palme, J.; Kristiansson, E.; Larsson, D.G.J. Environmental Factors Influencing the Development and Spread of Antibiotic Resistance. FEMS Microbiol. Rev. 2018, 42, 68–80. [Google Scholar] [CrossRef]
- Zainab, S.M.; Junaid, M.; Xu, N.; Malik, R.N. Antibiotics and Antibiotic Resistant Genes (ARGs) in Groundwater: A Global Review on Dissemination, Sources, Interactions, Environmental and Human Health Risks. Water Res. 2020, 187, 116455. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lepper, H.C.; Woolhouse, M.E.J.; van Bunnik, B.A.D. The Role of the Environment in Dynamics of Antibiotic Resistance in Humans and Animals: A Modelling Study. Antibiotics 2022, 11, 1361. https://doi.org/10.3390/antibiotics11101361
Lepper HC, Woolhouse MEJ, van Bunnik BAD. The Role of the Environment in Dynamics of Antibiotic Resistance in Humans and Animals: A Modelling Study. Antibiotics. 2022; 11(10):1361. https://doi.org/10.3390/antibiotics11101361
Chicago/Turabian StyleLepper, Hannah C., Mark E. J. Woolhouse, and Bram A. D. van Bunnik. 2022. "The Role of the Environment in Dynamics of Antibiotic Resistance in Humans and Animals: A Modelling Study" Antibiotics 11, no. 10: 1361. https://doi.org/10.3390/antibiotics11101361