
Effects of a Specific Pre- and Probiotic Combination and Parent Stock Vaccination on Performance and Bacterial Communities in Broilers Challenged with a Multidrug-Resistant Escherichia coli

Abstract
:1. Introduction
2. Results
2.1. Performance
2.2. Microbial Metabolites and pH in Crop and Cecal Contents
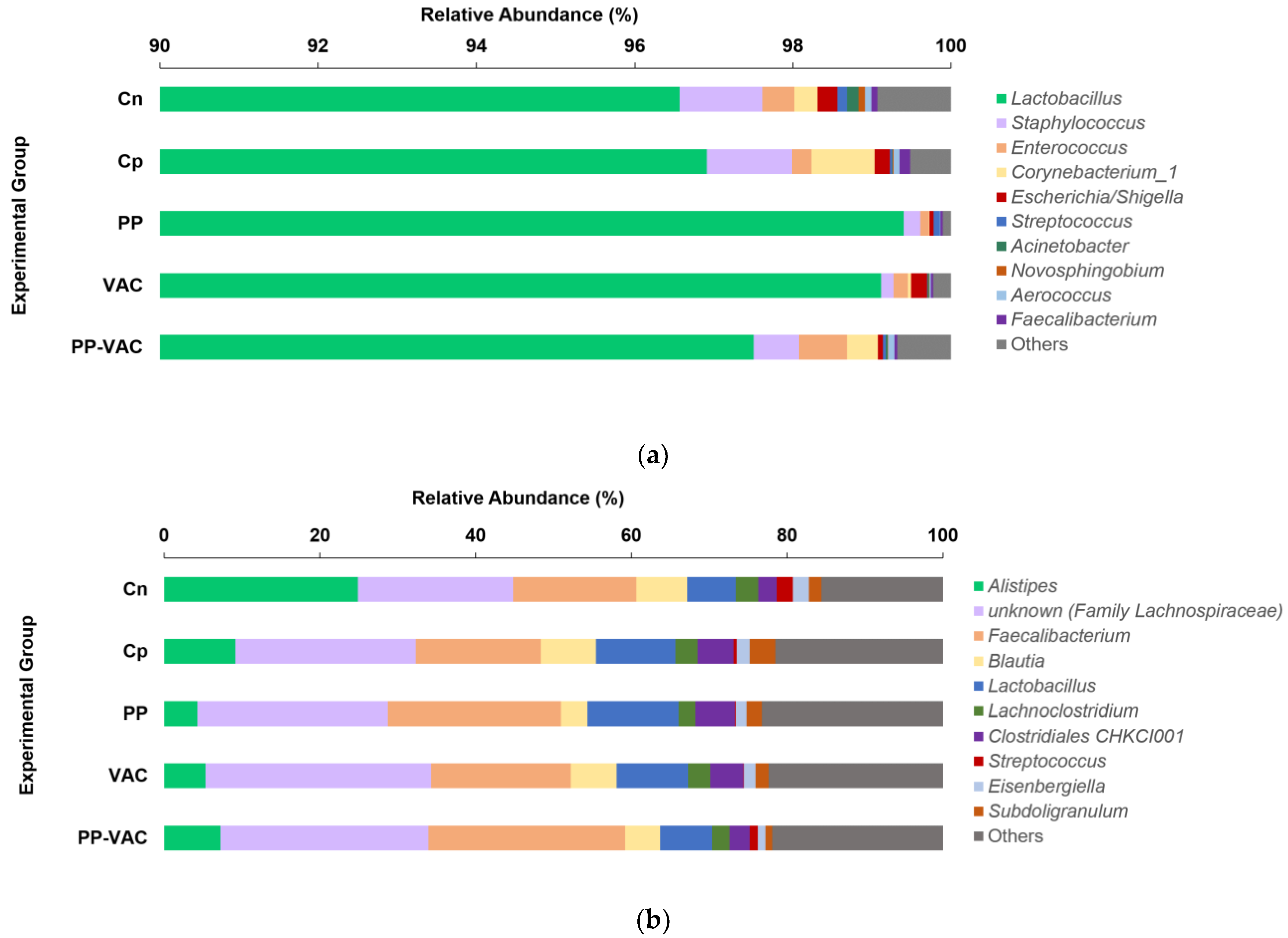
2.3. Relative Abundances of Bacteria and Microbial Ecological Indices in Crop and Cecal Contents
2.4. Quantitative Determination of Enterobacterial Antibiotic Resistance Genes and the Corresponding Escherichia coli/Hafnia/Shigella Group
3. Discussion
4. Materials and Methods
4.1. Broiler Breeders and Vaccination
4.2. Broiler Chicks and Experimental Groups
4.3. Challenge Strain
4.4. Experimental Diets, Prebiotic and Probiotic
4.5. Sampling
4.6. Analysis of Microbial Metabolites and pH
4.7. 16S rDNA Sequencing and qPCR
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Huemer, M.; Mairpady Shambat, S.; Brugger, S.D.; Zinkernagel, A.S. Antibiotic resistance and persistence-Implications for human health and treatment perspectives. EMBO Rep. 2020, 21, e51034. [Google Scholar] [CrossRef] [PubMed]
- No Time to Wait: Securing the Future from Drug-Resistant Infections, Report to the Secretary-General of the United Nations. Available online: https://www.who.int/docs/default-source/documents/no-time-to-wait-securing-the-future-from-drug-resistant-infections-en.pdf (accessed on 10 August 2022).
- Pomba, C.; Rantala, M.; Greko, C.; Baptiste, K.E.; Catry, B.; van Duijkeren, E.; Mateus, A.; Moreno, M.A.; Pyörälä, S.; Ružauskas, M.; et al. Public health risk of antimicrobial resistance transfer from companion animals. J. Antimicrob. Chemother. 2017, 72, 957–968. [Google Scholar] [CrossRef]
- Saliu, E.M.; Vahjen, W.; Zentek, J. Types and prevalence of extended-spectrum beta-lactamase producing Enterobacteriaceae in poultry. Anim. Health Res. Rev. 2017, 18, 46–57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- European Parliament and the Council of the European Union. Regulation (EC) No. 1831/2003 of the European Parliament and of the Council of 22 September 2003 on Additives for Use in Animal Nutrition, Chapter II, Article 11, Paragraph 2; European Parliament and the Council of the European Union: Brussels, Belgium, 2003; Volume L 268, pp. 29–43. [Google Scholar]
- European Parliament and the Council of the European Union. Regulation (EU) 2019/6 of the European Parliament and of the Council of 11 December 2018 on Veterinary Medicinal Products and Repealing Directive 2001/82/EC; European Parliament and the Council of the European Union: Brussels, Belgium, 2019; Volume L 4, pp. 43–167. [Google Scholar]
- World Organisation for Animal Health (OIE). OIE Annual Report on Antimicrobial Agents Intended for Use in Animals, Fifth Report. Available online: https://rr-americas.woah.org/en/news/new-annual-report-amu/ (accessed on 11 August 2022).
- Gadde, U.; Kim, W.H.; Oh, S.T.; Lillehoj, H.S. Alternatives to antibiotics for maximizing growth performance and feed efficiency in poultry: A review. Anim. Health Res. Rev. 2017, 18, 26–45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clavijo, V.; Flórez, M.J.V. The gastrointestinal microbiome and its association with the control of pathogens in broiler chicken production: A review. Poult. Sci. 2018, 97, 1006–1021. [Google Scholar] [CrossRef]
- Gregersen, R.H.; Christensen, H.; Ewers, C.; Bisgaard, M. Impact of Escherichia coli vaccine on parent stock mortality, first week mortality of broilers and population diversity of E. coli in vaccinated flocks. Avian Pathol. J. WVPA 2010, 39, 287–295. [Google Scholar] [CrossRef]
- Cauwerts, K.; De Herdt, P.; Haesebrouck, F.; Vervloesem, J.; Ducatelle, R. The effect of Ornithobacterium rhinotracheale vaccination of broiler breeder chickens on the performance of their progeny. Avian Pathol. J. WVPA 2002, 31, 619–624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crouch, C.F.; Withanage, G.S.; de Haas, V.; Etore, F.; Francis, M.J. Safety and efficacy of a maternal vaccine for the passive protection of broiler chicks against necrotic enteritis. Avian Pathol. J. WVPA 2010, 39, 489–497. [Google Scholar] [CrossRef]
- Gharaibeh, S.; Mahmoud, K. Decay of maternal antibodies in broiler chickens. Poult. Sci. 2013, 92, 2333–2336. [Google Scholar] [CrossRef]
- Larsson, A.; Sjöquist, J. Chicken IgY: Utilizing the evolutionary difference. Comp. Immunol. Microbiol. Infect. Dis. 1990, 13, 199–201. [Google Scholar] [CrossRef]
- Williams, J.; Mallet, S.; Leconte, M.; Lessire, M.; Gabriel, I. The effects of fructo-oligosaccharides or whole wheat on the performance and digestive tract of broiler chickens. Br. Poult. Sci. 2008, 49, 329–339. [Google Scholar] [CrossRef] [PubMed]
- Kim, G.B.; Seo, Y.M.; Kim, C.H.; Paik, I.K. Effect of dietary prebiotic supplementation on the performance, intestinal microflora, and immune response of broilers. Poult. Sci. 2011, 90, 75–82. [Google Scholar] [CrossRef] [PubMed]
- Samli, H.E.; Senkoylu, N.; Koc, F.; Kanter, M.; Agma, A. Effects of Enterococcus faecium and dried whey on broiler performance, gut histomorphology and intestinal microbiota. Arch. Anim. Nutr. 2007, 61, 42–49. [Google Scholar] [CrossRef]
- Cengiz, Ö.; Köksal, B.H.; Tatlı, O.; Sevim, Ö.; Ahsan, U.; Üner, A.G.; Ulutaş, P.A.; Beyaz, D.; Büyükyörük, S.; Yakan, A.; et al. Effect of dietary probiotic and high stocking density on the performance, carcass yield, gut microflora, and stress indicators of broilers. Poult. Sci. 2015, 94, 2395–2403. [Google Scholar] [CrossRef] [PubMed]
- Kariyawasam, S.; Wilkie, B.N.; Gyles, C.L. Resistance of broiler chickens to Escherichia coli respiratory tract infection induced by passively transferred egg-yolk antibodies. Vet. Microbiol. 2004, 98, 273–284. [Google Scholar] [CrossRef] [PubMed]
- Borst, L.B.; Suyemoto, M.M.; Chen, L.R.; Barnes, H.J. Vaccination of breeder hens with a polyvalent killed vaccine for pathogenic Enterococcus cecorum does not protect offspring from enterococcal spondylitis. Avian Pathol. J. WVPA 2019, 48, 17–24. [Google Scholar] [CrossRef]
- Zenner, C.; Hitch, T.C.A.; Riedel, T.; Wortmann, E.; Tiede, S.; Buhl, E.M.; Abt, B.; Neuhaus, K.; Velge, P.; Overmann, J.; et al. Early-life immune system maturation in chickens using a synthetic community of cultured gut bacteria. mSystems 2021, 6, e01300-20. [Google Scholar] [CrossRef]
- Massacci, F.R.; Lovito, C.; Tofani, S.; Tentellini, M.; Genovese, D.A.; De Leo, A.A.P.; Papa, P.; Magistrali, C.F.; Manuali, E.; Trabalza-Marinucci, M.; et al. Dietary Saccharomyces cerevisiae boulardii CNCM I-1079 positively affects performance and intestinal ecosystem in broilers during a Campylobacter jejuni Infection. Microorganisms 2019, 7, 596. [Google Scholar] [CrossRef] [Green Version]
- Żbikowski, A.; Pawłowski, K.; Śliżewska, K.; Dolka, B.; Nerc, J.; Szeleszczuk, P. Comparative effects of using new multi-strain synbiotics on chicken growth performance, hematology, serum biochemistry and immunity. Animals 2020, 10, 1555. [Google Scholar] [CrossRef]
- De Herdt, P.; Broeckx, M.; Vankeirsbilck, W.; Van Den Abeele, G.; Van Gorp, S. Improved broiler performance associated with Ornithobacterium rhinotracheale vaccination in breeders. Avian Dis. 2012, 56, 365–368. [Google Scholar] [CrossRef]
- Awad, W.; Ghareeb, K.; Böhm, J. Intestinal structure and function of broiler chickens on diets supplemented with a synbiotic containing Enterococcus faecium and oligosaccharides. Int. J. Mol. Sci. 2008, 9, 2205–2216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Awad, W.A.; Ghareeb, K.; Abdel-Raheem, S.; Böhm, J. Effects of dietary inclusion of probiotic and synbiotic on growth performance, organ weights, and intestinal histomorphology of broiler chickens. Poult. Sci. 2009, 88, 49–56. [Google Scholar] [CrossRef]
- Talebi, A.; Amani, A.; Pourmahmod, M.; Saghaei, P.; Rezaie, R. Synbiotic enhances immune responses against infectious bronchitis, infectious bursal disease, Newcastle disease and avian influenza in broiler chickens. Vet. Res. Forum Int. Q. J. 2015, 6, 191–197. [Google Scholar]
- Ren, H.; Vahjen, W.; Dadi, T.; Saliu, E.M.; Boroojeni, F.G.; Zentek, J. Synergistic effects of probiotics and phytobiotics on the intestinal microbiota in young broiler chicken. Microorganisms 2019, 7, 684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rehman, H.U.; Vahjen, W.; Awad, W.A.; Zentek, J. Indigenous bacteria and bacterial metabolic products in the gastrointestinal tract of broiler chickens. Arch. Anim. Nutr. 2007, 61, 319–335. [Google Scholar] [CrossRef]
- Józefiak, D.; Rutkowski, A.; Jensen, B.B.; Engberg, R.M. Effects of dietary inclusion of triticale, rye and wheat and xylanase supplementation on growth performance of broiler chickens and fermentation in the gastrointestinal tract. Anim. Feed Sci. Technol. 2007, 132, 79–93. [Google Scholar] [CrossRef]
- Goodarzi Boroojeni, F.; Vahjen, W.; Mader, A.; Knorr, F.; Ruhnke, I.; Röhe, I.; Hafeez, A.; Villodre, C.; Männer, K.; Zentek, J. The effects of different thermal treatments and organic acid levels in feed on microbial composition and activity in gastrointestinal tract of broilers. Poult. Sci. 2014, 93, 1440–1452. [Google Scholar] [CrossRef]
- Tejero-Sariñena, S.; Barlow, J.; Costabile, A.; Gibson, G.R.; Rowland, I. In vitro evaluation of the antimicrobial activity of a range of probiotics against pathogens: Evidence for the effects of organic acids. Anaerobe 2012, 18, 530–538. [Google Scholar] [CrossRef]
- Oyarzabal, O.A.; Conner, D.E.; Blevins, W.T. Fructooligosaccharide utilization by Salmonellae (†) and potential direct-fed-microbial bacteria for poultry. J. Food Prot. 1995, 58, 1192–1196. [Google Scholar] [CrossRef]
- Bar-Shira, E.; Cohen, I.; Elad, O.; Friedman, A. Role of goblet cells and mucin layer in protecting maternal IgA in precocious birds. Dev. Comp. Immunol. 2014, 44, 186–194. [Google Scholar] [CrossRef]
- Kogut, M.H.; Lee, A.; Santin, E. Microbiome and pathogen interaction with the immune system. Poult. Sci. 2020, 99, 1906–1913. [Google Scholar] [CrossRef] [PubMed]
- Hartog, G.; Vries-Reilingh, G.; Wehrmaker, A.M.; Savelkoul, H.; Parmentier, H.; Lammers, A. Intestinal immune maturation is accompanied by temporal changes in the composition of the microbiota. Benef. Microbes 2016, 7, 677–685. [Google Scholar] [CrossRef] [PubMed]
- Borey, M.; Bed’Hom, B.; Bruneau, N.; Estellé, J.; Larsen, F.; Blanc, F.; der Laan, M.H.; Dalgaard, T.; Calenge, F. Caecal microbiota composition of experimental inbred MHC-B lines infected with IBV differs according to genetics and vaccination. Sci. Rep. 2022, 12, 9995. [Google Scholar] [CrossRef] [PubMed]
- Beirão, B.C.B.; Ingberman, M.; Mesa, D.; Salles, G.B.C.; Muniz, E.C.; Caron, L.F. Effects of aroA deleted E. coli vaccine on intestinal microbiota and mucosal immunity. Comp. Immunol. Microbiol. Infect. Dis. 2021, 75, 101612. [Google Scholar] [CrossRef]
- Orso, C.; Stefanello, T.B.; Franceschi, C.H.; Mann, M.B.; Varela, A.P.M.; Castro, I.M.S.; Frazzon, J.; Frazzon, A.P.; Andretta, I.; Ribeiro, A.M. Changes in the ceca microbiota of broilers vaccinated for coccidiosis or supplemented with salinomycin. Poult. Sci. 2021, 100, 100969. [Google Scholar] [CrossRef]
- Beirão, B.C.B.; Ingberman, M.; Fávaro, C., Jr.; Mesa, D.; Bittencourt, L.C.; Fascina, V.B.; Caron, L.F. Effect of an Enterococcus faecium probiotic on specific IgA following live Salmonella Enteritidis vaccination of layer chickens. Avian Pathol. J. WVPA 2018, 47, 325–333. [Google Scholar] [CrossRef]
- Abd El-Ghany, W.A. Pseudomonas aeruginosa infection of avian origin: Zoonosis and one health implications. Vet. World 2021, 14, 2155–2159. [Google Scholar] [CrossRef]
- Oakley, B.B.; Kogut, M.H. Spatial and temporal changes in the broiler chicken cecal and fecal microbiomes and correlations of bacterial taxa with cytokine gene expression. Front. Vet. Sci. 2016, 3, 11. [Google Scholar] [CrossRef] [Green Version]
- Gong, J.; Si, W.; Forster, R.J.; Huang, R.; Yu, H.; Yin, Y.; Yang, C.; Han, Y. 16S rRNA gene-based analysis of mucosa-associated bacterial community and phylogeny in the chicken gastrointestinal tracts: From crops to ceca. FEMS Microbiol. Ecol. 2007, 59, 147–157. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Q.; Lan, F.; Li, X.; Yan, W.; Sun, C.; Li, J.; Yang, N.; Wen, C. The spatial and temporal characterization of gut microbiota in broilers. Front. Vet. Sci. 2021, 8, 712226. [Google Scholar] [CrossRef]
- Grześkowiak, Ł.; Dadi, T.H.; Zentek, J.; Vahjen, W. Developing gut microbiota exerts colonisation resistance to Clostridium (syn. Clostridioides) difficile in piglets. Microorganisms 2019, 7, 218. [Google Scholar] [CrossRef]
- Yang, W.Y.; Lee, Y.; Lu, H.; Chou, C.H.; Wang, C. Analysis of gut microbiota and the effect of lauric acid against necrotic enteritis in Clostridium perfringens and Eimeria side-by-side challenge model. PLoS ONE 2019, 14, e0205784. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rutebemberwa, A.; Stevens, M.J.; Perez, M.J.; Smith, L.P.; Sanders, L.; Cosgrove, G.; Robertson, C.E.; Tuder, R.M.; Harris, J.K. Novosphingobium and its potential role in chronic obstructive pulmonary diseases: Insights from microbiome studies. PLoS ONE 2014, 9, e111150. [Google Scholar] [CrossRef] [PubMed]
- Mohammed, J.P.; Mattner, J. Autoimmune disease triggered by infection with alphaproteobacteria. Expert Rev. Clin. Immunol. 2009, 5, 369–379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hedegaard, L.; Christensen, H.; Chadfield, M.S.; Christensen, J.P.; Bisgaard, M. Association of Streptococcus pluranimalium with valvular endocarditis and septicaemia in adult broiler parents. Avian Pathol. 2009, 38, 155–160. [Google Scholar] [CrossRef] [Green Version]
- Chadfield, M.S.; Christensen, J.P.; Decostere, A.; Christensen, H.; Bisgaard, M. Geno-and phenotypic diversity of avian isolates of Streptococcus gallolyticus subsp. gallolyticus (Streptococcus bovis) and associated diagnostic problems. J. Clin. Microbiol. 2007, 45, 822–827. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Devriese, L.A.; Haesebrouck, F.; de Herdt, P.; Dom, P.; Ducatelle, R.; Desmidt, M.; Messier, S.; Higgins, R. Streptococcus suis infections in birds. Avian Pathol. J. WVPA 1994, 23, 721–724. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Stewart, S.N.; Robinson, K.; Yang, Q.; Lyu, W.; Whitmore, M.A.; Zhang, G. Linkage between the intestinal microbiota and residual feed intake in broiler chickens. J. Anim. Sci. Biotechnol. 2021, 12, 22. [Google Scholar] [CrossRef]
- Polansky, O.; Sekelova, Z.; Faldynova, M.; Sebkova, A.; Sisak, F.; Rychlik, I. Important metabolic pathways and biological processes expressed by chicken cecal microbiota. Appl. Environ. Microbiol. 2015, 82, 1569–1576. [Google Scholar] [CrossRef] [Green Version]
- Fleming, S.E.; Fitch, M.D.; DeVries, S.; Liu, M.L.; Kight, C. Nutrient utilization by cells isolated from rat jejunum, cecum and colon. J. Nutr. 1991, 121, 869–878. [Google Scholar] [CrossRef]
- Xiang, H.; Gan, J.; Zeng, D.; Li, J.; Yu, H.; Zhao, H.; Yang, Y.; Tan, S.; Li, G.; Luo, C.; et al. Specific microbial taxa and functional capacity contribute to chicken abdominal fat deposition. Front. Microbiol. 2021, 12, 643025. [Google Scholar] [CrossRef]
- Tang, S.; Xin, Y.; Ma, Y.; Xu, X.; Zhao, S.; Cao, J. Screening of microbes associated with swine growth and fat deposition traits across the intestinal tract. Front. Microbiol. 2020, 11, 586776. [Google Scholar] [CrossRef] [PubMed]
- Gong, L.; Xiao, G.; Zheng, L.; Yan, X.; Qi, Q.; Zhu, C.; Feng, X.; Huang, W.; Zhang, H. Effects of dietary tributyrin on growth performance, biochemical indices, and intestinal microbiota of yellow-feathered broilers. Animals 2021, 11, 3425. [Google Scholar] [CrossRef]
- Mishra, R.P.; Oviedo-Orta, E.; Prachi, P.; Rappuoli, R.; Bagnoli, F. Vaccines and antibiotic resistance. Curr. Opin. Microbiol. 2012, 15, 596–602. [Google Scholar] [CrossRef]
- Jansen, K.U.; Knirsch, C.; Anderson, A.S. The role of vaccines in preventing bacterial antimicrobial resistance. Nat. Med. 2018, 24, 10–19. [Google Scholar] [CrossRef] [PubMed]
- ROSS 308 AP Parent Stock Performance Objectives. Available online: https://en.aviagen.com/assets/Tech_Center/Ross_PS/Ross308AP-ParentStock-PerformanceObjectives-2021-EN.pdf (accessed on 15 March 2021).
- Leeson, S.; Summers, J.D. Commercial Poultry Nutrition, 3rd ed.; Nottingham University Press: Nottingham, UK, 2005; p. 398. [Google Scholar]
- ROSS 308 Parent Stock Nutrition Specifications. Available online: https://en.aviagen.com/assets/Tech_Center/Ross_PS/Ross308-PS-NS-2016-EN.pdf (accessed on 10 December 2020).
- Spitzer, F.; Vahjen, W.; Pieper, R.; Martinez-Vallespin, B.; Zentek, J. A standardised challenge model with an enterotoxigenic F4+ Escherichia coli strain in piglets assessing clinical traits and faecal shedding of fae and est-II toxin genes. Arch. Anim. Nutr. 2014, 68, 448–459. [Google Scholar] [CrossRef] [PubMed]
- GfE. Empfehlungen zur Energie- und Nährstoffversorgung der Legehennen und Masthühner (Broiler); DLG: Frankfurt, Germany, 1999; p. 145. [Google Scholar]
- Fuhrmann, L.; Vahjen, W.; Zentek, J.; Günther, R.; Saliu, E.-M. The impact of pre-and probiotic product combinations on ex vivo growth of avian pathogenic Escherichia coli and Salmonella Enteritidis. Microorganisms 2022, 10, 121. [Google Scholar] [CrossRef]
- Kröger, S.; Vahjen, W.; Zentek, J. Influence of lignocellulose and low or high levels of sugar beet pulp on nutrient digestibility and the fecal microbiota in dogs. J. Anim. Sci. 2017, 95, 1598–1605. [Google Scholar] [CrossRef]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef]
- Yilmaz, P.; Parfrey, L.W.; Yarza, P.; Gerken, J.; Pruesse, E.; Quast, C.; Schweer, T.; Peplies, J.; Ludwig, W.; Glöckner, F.O. The SILVA and “All-species Living Tree Project (LTP)” taxonomic frameworks. Nucleic Acids Res. 2014, 42, D643–D648. [Google Scholar] [CrossRef] [Green Version]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [PubMed]
- Vahjen, W.; Pietruszyńska, D.; Starke, I.C.; Zentek, J. High dietary zinc supplementation increases the occurrence of tetracycline and sulfonamide resistance genes in the intestine of weaned pigs. Gut Pathog. 2015, 7, 23. [Google Scholar] [CrossRef] [Green Version]
- Toro, C.S.; Farfán, M.; Contreras, I.; Flores, O.; Navarro, N.; Mora, G.C.; Prado, V. Genetic analysis of antibiotic-resistance determinants in multidrug-resistant Shigella strains isolated from Chilean children. Epidemiol. Infect. 2005, 133, 81–86. [Google Scholar] [CrossRef] [PubMed]
- Yagi, T.; Kurokawa, H.; Shibata, N.; Shibayama, K.; Arakawa, Y. A preliminary survey of extended-spectrum beta-lactamases (ESBLs) in clinical isolates of Klebsiella pneumoniae and Escherichia coli in Japan. FEMS Microbiol. Lett. 2000, 184, 53–56. [Google Scholar] [PubMed] [Green Version]
- Malinen, E.; Kassinen, A.; Rinttilä, T.; Palva, A. Comparison of real-time PCR with SYBR Green I or 5′-nuclease assays and dot-blot hybridization with rDNA-targeted oligonucleotide probes in quantification of selected faecal bacteria. Microbiology 2003, 149, 269–277. [Google Scholar] [CrossRef]

{kind=link}
| Experimental Groups 2 | ||||||
|---|---|---|---|---|---|---|
| Week | Cn | Cp | PP | VAC | PP-VAC | p-Value |
| Average bodyweight (g)/animal | ||||||
| 0 | 39.0 ± 0.70 | 39.8 ± 0.56 | 39.3 ± 1.04 | 38.6 ± 0.84 | 38.7 ± 1.36 | 0.262 |
| 1 | 92.4 ± 10.9 a | 86.6 ± 9.23 a | 92.0 ± 6.97 a | 104 ± 14.5 a,b | 123 ± 13.6 b | <0.001 |
| 2 | 224 ± 27.3 a | 223 ± 23.62 a | 265 ± 17.0 a,b | 277 ± 25.6 b | 319 ± 20.6 c | <0.001 |
| 3 | 520 ± 75.3 a | 543 ± 52.8 a,b | 580 ± 37.1 a,b,c | 613 ± 40.5 b,c | 680 ± 31.0 c | <0.001 |
| 4 | 929 ± 119 a | 985 ± 70.9 a,b | 1017 ± 64.3 a,b | 1091 ± 31.2 b,c | 1144 ± 45.5 c | <0.001 |
| Average bodyweight gain (g)/animal | ||||||
| 1 | 53.5 ± 10.8 a,b | 46.8 ± 8.74 a | 52.7 ± 7.78 a,b | 65.7 ± 15.1 b,c | 83.8 ± 13.7 c | <0.001 |
| 2 | 132 ± 17.1 a | 147 ± 14.9 a | 169 ± 14.6 b | 172 ± 12.9 b | 197 ± 9.10 c | <0.001 |
| 3 | 296 ± 48.4 a | 310 ± 30.1 a,b | 316 ± 21.8 a,b | 336 ± 20.2 a,b | 361 ± 20.5 b | 0.026 |
| 4 | 410 ± 44.7 | 442 ± 26.7 | 437 ± 29.7 | 479 ± 49.5 | 464 ± 21.0 | 0.086 |
| 1–4 | 890 ± 119 a | 945 ± 70.6 a,b | 974 ± 64.4 a,b | 1052 ± 31.3 b,c | 1106 ± 44.2 c | <0.001 |
| Average feed intake (g)/animal | ||||||
| 1 | 68.4 ± 12.7 a | 71.4 ± 10.9 a | 68.7 ± 4.41 a | 91.5 ± 18.9 a,b | 96.8 ± 14.0 b | 0.002 |
| 2 | 203 ± 12.84 a | 233 ± 16.2 b | 253 ± 6.11 b,c | 277 ± 29.5 c,d | 287 ± 17.5 d | <0.001 |
| 3 | 368 ± 59.0 a | 387 ± 41.4 a | 412 ± 34.0 a,b | 421 ± 29.0 a,b | 477 ± 19.6 b | 0.003 |
| 4 | 557 ± 62.0 a | 599 ± 41.0 a,b | 612 ± 47.2 a,b | 633 ± 33.9 a,b | 672 ± 33.4 b | 0.018 |
| 1–4 | 1197 ± 141 a | 1289 ± 88.2 a,b | 1347 ± 82.6 a,b | 1422 ± 76.2 b,c | 1532 ± 64.4 c | <0.001 |
| Feed conversion ratio 3 | ||||||
| 1 | 1.39 ± 0.14 a,b | 1.54 ± 0.14 b | 1.47 ± 0.21 b | 1.48 ± 0.16 b | 1.17 ± 0.03 a | 0.007 |
| 2 | 1.55 ± 0.17 | 1.60 ± 0.18 | 1.51 ± 0.12 | 1.61 ± 0.14 | 1.47 ± 0.04 | 0.253 |
| 3 | 1.25 ± 0.02 a | 1.25 ± 0.02 a | 1.30 ± 0.03 b | 1.25 ± 0.04 a | 1.32 ± 0.04 b | 0.001 |
| 4 | 1.36 ± 0.02 a | 1.36 ± 0.04 a | 1.40 ± 0.03 a,b | 1.33 ± 0.14 a | 1.45 ± 0.02 b | <0.001 |
| 1–4 | 1.35 ± 0.04 | 1.38 ± 0.01 | 1.39 ± 0.03 | 1.35 ± 0.07 | 1.39 ± 0.01 | 0.449 |
| Experimental Groups 2 | |||||||
|---|---|---|---|---|---|---|---|
| Cn | Cp | PP | VAC | PP-VAC | SEM | p-Value | |
| Crop | |||||||
| pH | 5.17 * | 5.16 * | 4.86 | 4.88 | 4.74 # | 0.052 | 0.022 |
| L-lactate | 20.4 a | 22.0 a,b,# | 49.6 b,c,* | 41.5 a,b,c | 52.5 c | 3.34 | 0.002 |
| D-lactate | 7.80 | 12.1 | 21.8 | 15.5 | 34.1 | 3.07 | 0.100 |
| acetate | 8.31 | 7.92 | 10.9 | 6.47 | 11.8 | 0.946 | 0.396 |
| propionate | 0.120 b | 0.083 a,b | 0.071 a | 0.077 a,b | 0.070 a | 0.005 | 0.013 |
| i-butyrate | 0.087 | 0.137 | 0.104 | 0.101 | 0.118 | 0.006 | 0.176 |
| n-butyrate | 0.026 | 0.025 | 0.021 | 0.017 | 0.016 | 0.004 | 0.661 |
| i-valerate | 0.010 a,b | 0.008 a | 0.013 a,b | 0.013 a,b | 0.019 b | 0.001 | 0.021 |
| n-valerate | 0.043 | 0.033 | 0.030 | 0.032 | 0.024 | 0.002 | 0.168 |
| total BCFA | 0.097 | 0.145 | 0.118 | 0.114 | 0.136 | 0.007 | 0.181 |
| total SCFA | 6.45 | 8.26 | 11.1 | 6.72 | 12.0 | 0.896 | 0.263 |
| Cecum | |||||||
| pH | 7.24 c | 7.10 b,c | 6.75 a | 6.88 a,b | 6.77 a | 0.042 | <0.001 |
| acetate | 62.3 a | 69.6 a,b | 81.1 a,b | 90.6 b | 88.4 b | 2.48 | <0.001 |
| propionate | 8.87 | 7.82 | 8.76 | 10.1 | 8.95 | 0.451 | 0.655 |
| i-butyrate | 1.58 | 1.37 | 1.22 | 1.86 | 1.84 | 0.092 | 0.117 |
| n-butyrate | 7.80 a,# | 12.7 a,b | 16.6 a,b,* | 20.2 b | 19.4 b | 1.07 | <0.001 |
| i-valerate | 1.34 | 1.41 | 1.29 | 1.75 | 1.72 | 0.084 | 0.179 |
| n-valerate | 1.24 a,b | 1.22 a,b | 1.11 a | 1.54 b | 1.35 a,b | 0.059 | 0.021 |
| total BCFA | 2.92 | 2.78 | 2.51 | 3.62 | 3.56 | 0.155 | 0.113 |
| total SCFA | 83.3 a | 94.1 a,b,# | 110 a,b | 126 b,* | 122 b | 3.44 | <0.001 |
| Experimental Groups 2 | |||||||
|---|---|---|---|---|---|---|---|
| Cn | Cp | PP | VAC | PP-VAC | SEM | p-Value | |
| Crop | |||||||
| Firmicutes | 91.3 a,b,# | 91.3 a | 97.6 b,* | 96.9 a,b | 96.7 b,* | 0.710 | 0.002 |
| Proteobacteria | 8.22 b | 7.83 b | 2.40 a,b | 3.06 a,b | 1.92 a | 0.711 | 0.003 |
| Actinobacteria | 0.202 | 0.033 | 0.013 | 0.052 | 0.653 | 0.078 | 0.152 |
| Bacteroidetes | 0.059 b | n.d. a | n.d. a | 0.003 a | n.d. a | 0.006 | <0.001 |
| Cecum | |||||||
| Firmicutes | 73.8 a | 88.5 b | 92.5 b | 92.1 b | 91.7 b | 1.36 | <0.001 |
| Bacteroidetes | 25.9 b,* | 11.1 a,b,# | 7.32 a | 7.68 a | 8.02 a | 1.36 | <0.001 |
| Proteobacteria | 0.001 # | 0.275 * | 0.138 | 0.202 | 0.005 # | 0.026 | 0.010 |
| Actinobacteria | 0.047 | 0.069 | 0.047 | 0.044 | 0.046 | 0.006 | 0.990 |
| Tenericutes | 0.067 | 0.006 | 0.001 | 0.034 | 0.012 | 0.008 | 0.061 |
| Experimental Groups 2 | ||||||||
|---|---|---|---|---|---|---|---|---|
| Target Gene 3 | Day | Cn | Cp | PP | VAC | PP-VAC | SEM | p-Value |
| sul1-3 | ||||||||
| 10 | 4.09 a,# | 7.68 b | 7.39 a,b,* | 6.75 a,b | 7.19 a,b | 0.330 | 0.015 | |
| 14 | 6.32 # | 7.92 | 7.33 | 7.73 | 8.24 * | 0.219 | 0.053 | |
| 21 | 6.26 a | 7.57 a,b | 8.35 b | 7.60 a,b | 6.95 a,b | 0.196 | 0.012 | |
| 28 | 7.81 | 8.55 | 8.14 | 8.03 | 7.72 | 0.128 | 0.354 | |
| dhfr1a | ||||||||
| 10 | 2.52 a | 4.44 a,b | 5.65 b | 2.37 a | 3.33 a,b | 0.517 | 0.010 | |
| 14 | 1.43 | 4.48 | 2.37 | 3.76 | 3.77 | 0.523 | 0.222 | |
| 21 | 2.55 a | 5.63 a,b | 6.44 b | 1.43 a | 2.47 a | 0.574 | <0.001 | |
| 28 | 3.38 | 4.76 | 2.76 | 1.43 | 2.70 | 0.525 | 0.080 | |
| SHV-12 | ||||||||
| 10 | 3.29 | 2.80 | 4.58 | 3.51 | 4.81 | 0.457 | 0.217 | |
| 14 | 3.86 | 4.08 | 3.85 | 1.36 | 3.08 | 0.438 | 0.295 | |
| 21 | 3.63 | 3.64 | 3.20 | 1.36 | 1.36 | 0.384 | 0.124 | |
| 28 | 3.99 | 3.12 | 2.93 | 1.36 | 1.36 | 0.385 | 0.097 | |
| E. coli/Hafnia/Shigella | ||||||||
| 10 | 8.76 a,b,# | 9.51 b,* | 9.34 a,b | 8.57 a | 8.88 a,b | 0.135 | 0.020 | |
| 14 | 8.75 | 9.05 | 8.57 | 8.84 | 8.81 | 0.126 | 0.845 | |
| 21 | 8.87 | 8.82 | 9.01 | 8.82 | 9.31 | 0.113 | 0.396 | |
| 28 | 8.84 | 9.03 | 9.18 | 8.75 | 9.08 | 0.109 | 0.703 | |
| Experimental Group 1 | Parent Stock Vaccination 2 | Pre-/Probiotic Combination 3 | E. coli- Challenge 4 |
|---|---|---|---|
| Cn | No | No | No |
| Cp | No | No | Yes |
| PP | No | Yes | Yes |
| VAC | Yes | No | Yes |
| PP-VAC | Yes | Yes | Yes |
| Starter | Grower | |||
|---|---|---|---|---|
| Basal diet | Pre-/Probiotic Diet 1 | Basal Diet | Pre-/Probiotic Diet 1 | |
| Ingredients [%] | ||||
| Soybean Meal | 33.55 | 33.55 | 30.18 | 30.18 |
| Maize | 32.31 | 32.31 | 34.93 | 34.93 |
| Wheat | 24.85 | 23.85 | 25.00 | 24.00 |
| Soybean oil | 4.40 | 4.40 | 5.48 | 5.48 |
| Limestone | 1.68 | 1.68 | 1.54 | 1.54 |
| Monocalcium phosphate | 1.28 | 1.28 | 1.08 | 1.08 |
| Trace mineral and vitamin premix 2 | 1.20 | 1.20 | 1.20 | 1.20 |
| DL-Methionine | 0.35 | 0.35 | 0.30 | 0.30 |
| L-Lysine HCL | 0.28 | 0.28 | 0.22 | 0.22 |
| Threonine | 0.10 | 0.10 | 0.07 | 0.07 |
| FOS 3 | 0.00 | 1.00 | 0.00 | 1.00 |
| Analyzed Nutrient Composition [g/kg] | ||||
| Dry matter | 910 | 910 | 914 | 913 |
| Crude ash | 52.9 | 53.9 | 49.4 | 48.7 |
| Crude protein | 229 | 233 | 213 | 212 |
| Ether extract | 69.9 | 61.4 | 73.1 | 71.4 |
| Crude fiber | 28.2 | 24.9 | 25.0 | 27.6 |
| Methionine | 7.05 | 7.62 | 6.41 | 5.74 |
| Lysine | 13.8 | 13.5 | 12.4 | 10.5 |
| Calcium | 8.63 | 8.94 | 7.86 | 7.89 |
| Total phosphorus | 6.25 | 6.44 | 5.77 | 5.78 |
| AMEN 4 [MJ/kg] | 12.6 | 12.6 | 12.9 | 12.9 |
| Target Gene | Primer | Sequence (5′ to 3′) | Annealing Temperature | Reference |
|---|---|---|---|---|
| sul1-3 | sul1-3-f | CGATCCGGGGATGGGATTTT | 60 °C | [70] |
| sul1-3-r | CACCGAGACCAATAGCGGAA | |||
| dhfr1a | dhfr1a-f | GGAGTGCCAAAGGTGAACAGC | 50 °C | [71] |
| dhfr1a-r | GAGGCGAAGTCTTGGGTAAAAAC | |||
| SHV-12 | SHV-12-f | ATTTGTCGCTTCTTTACTCGC | 55 °C | [72] |
| SHV-12-r | TTTATGGCGTTACCTTTGACC | |||
| E. coli/Hafnia alvei/ Shigella spp. group | Entero-f | GTTAATACCTTTGCTCATTGA | 55 °C | [73] |
| Entero-r | ACCAGGGTATCTAATCCTGTT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fuhrmann, L.; Zentek, J.; Vahjen, W.; Günther, R.; Saliu, E.-M. Effects of a Specific Pre- and Probiotic Combination and Parent Stock Vaccination on Performance and Bacterial Communities in Broilers Challenged with a Multidrug-Resistant Escherichia coli. Antibiotics 2022, 11, 1703. https://doi.org/10.3390/antibiotics11121703
Fuhrmann L, Zentek J, Vahjen W, Günther R, Saliu E-M. Effects of a Specific Pre- and Probiotic Combination and Parent Stock Vaccination on Performance and Bacterial Communities in Broilers Challenged with a Multidrug-Resistant Escherichia coli. Antibiotics. 2022; 11(12):1703. https://doi.org/10.3390/antibiotics11121703
Chicago/Turabian StyleFuhrmann, Laura, Jürgen Zentek, Wilfried Vahjen, Ronald Günther, and Eva-Maria Saliu. 2022. "Effects of a Specific Pre- and Probiotic Combination and Parent Stock Vaccination on Performance and Bacterial Communities in Broilers Challenged with a Multidrug-Resistant Escherichia coli" Antibiotics 11, no. 12: 1703. https://doi.org/10.3390/antibiotics11121703