
Antimicrobial Potential of the Genera Geobacillus and Parageobacillus, as Well as Endolysins Biosynthesized by Their Bacteriophages

 and
and
Abstract
:1. Introduction
2. Antimicrobial Potential of Geobacillus and Parageobacillus
2.1. Geobacillus and Parageobacillus as a Source of Novel Antimicrobial Compounds
2.1.1. Geobacillus-Derived Volatile Organic Substances and Antibiotic Pigments
2.1.2. Geobacillus and Parageobacillus Derived Bacteriocins
2.1.3. Geobacillus and Parageobacillus Derived Bacteriocin-like Inhibitory Substances
2.1.4. Challenges in the Use of Geobacillus/Parageobacillus Bacteriocins and BLISes
2.2. The Potential of Geobacillus and Parageobacillus as Probiotics
2.3. A Fusion and Display System Based on Proteins Derived from Geobacillus
3. Antimicrobial Potential of Endolysins Derived from Bacteriophages Infecting Geobacillus and Parageobacillus
3.1. Historical Perspectives
3.2. Endolysins as Novel Antimicrobials
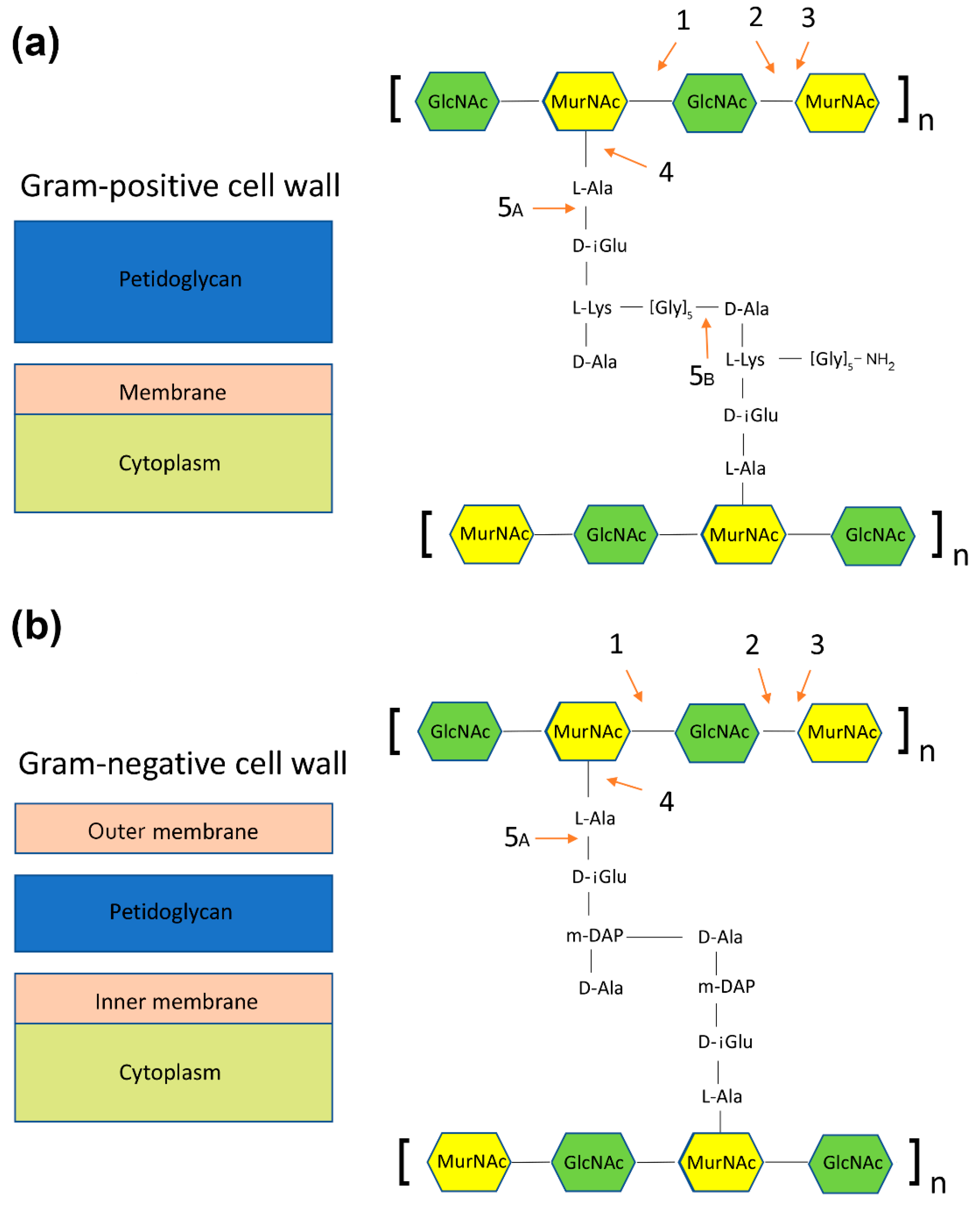
- N-acetylmuramidases (bond between N-acetylglucosamine and N-acetylmuramic acid) (Figure 3, panels a and b),
- N-acetylglucosaminidases (bond between N-acetylmuramic acid and N-acetylglucosamine) (Figure 3, panels a and b),
- Transglycosylases (bond between N-acetylmuramic acid and N-acetylglucosamine, a different mechanism than in N-acetylglucosaminidases) (Figure 3, panels a and b),
- Amidases (bond between N-acetylomuramic acid and L-alanine) (Figure 3, panels a and b)
- Endopeptidases:
3.3. Thermophilic Endolysins from Bacteriophage Infected Geobacillus Species
3.3.1. Thermostable GVE2 Endolysin
3.3.2. Thermostable G05 Endolysin
3.3.3. Thermostable GVE3 Endolysin
3.3.4. Thermostable TP-84_28 Endolysin
3.4. Challenges in the Application of the Geobacillus Bacteriophage Derived Endolysins
4. Outlook for Using Antimicrobials Derived from Geobacillus, Parageobacillus, and Their Bacteriophages
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Najar, I.N.; Thakur, N. A systematic review of the genera Geobacillus and Parageobacillus: Their evolution, current taxonomic status and major applications. Microbiology 2020, 166, 800–816. [Google Scholar] [CrossRef] [PubMed]
- Jeżewska-Frąckowiak, J.; Seroczyńska, K.; Banaszczyk, J.; Jedrzejczak, G.; Żylicz-Stachula, A.; Skowron, P.M. The promises and risks of probiotic Bacillus species. Acta Biochim. Pol. 2018, 65, 509–519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stein, T. Bacillus subtilis antibiotics: Structures, syntheses and specific functions. Mol. Microbiol. 2005, 56, 845–857. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Yi, H. Potential antitumor and anti-inflammatory activities of an extracellular polymeric substance (EPS) from Bacillus subtilis isolated from a housefly. Sci. Rep. 2022, 12, 1383. [Google Scholar] [CrossRef] [PubMed]
- Wada, K.; Suzuki, H. Chapter 15—Biotechnological platforms of the moderate thermophiles, Geobacillus species: Notable properties and genetic tools. In Physiological and Biotechnological Aspects of Extremophiles; Salwan, R., Sharma, V., Eds.; Academic Press: Cambridge, MA, USA, 2020; pp. 195–218. [Google Scholar] [CrossRef]
- Zeigler, D.R. The Geobacillus paradox: Why is a thermophilic bacterial genus so prevalent on a mesophilic planet? Microbiology 2014, 160 Pt 1, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nazina, T.N.; Tourova, T.P.; Poltaraus, A.B.; Novikova, E.V.; Grigoryan, A.A.; Ivanova, A.E.; Lysenko, A.M.; Petrunyaka, V.V.; Osipov, G.A.; Belyaev, S.S.; et al. Taxonomic study of aerobic thermophilic bacilli: Descriptions of Geobacillus subterraneus gen. nov., sp. nov. and Geobacillus uzenensis sp. nov. from petroleum reservoirs and transfer of Bacillus stearothermophilus, Bacillus thermocatenulatus, Bacillus thermoleovorans, Bacillus kaustophilus, Bacillus thermodenitrificans to Geobacillus as the new combinations G. stearothermophilus, G. th. Int. J. Syst. Evol. Microbiol. 2001, 51 Pt 2, 433–446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ash, C.; Farrow, J.A.E.; Wallbanks, S.; Collins, M.D. Phylogenetic heterogeneity of the genus Bacillus revealed by comparative analysis of small-subunit-ribosomal RNA sequences. Lett. Appl. Microbiol. 1991, 13, 202–206. [Google Scholar] [CrossRef]
- Aliyu, H.; Lebre, P.; Blom, J.; Cowan, D.; De Maayer, P. Phylogenomic re-assessment of the thermophilic genus Geobacillus. Syst. Appl. Microbiol. 2016, 39, 527–533, Erratum in Syst. Appl. Microbiol. 2018, 41, 529–530. [Google Scholar] [CrossRef] [Green Version]
- Najar, I.N.; Das, S.; Thakur, N. Reclassification of Geobacillus galactosidasius and Geobacillus yumthangensis as Parageobacillus galactosidasius comb. nov. and Parageobacillus yumthangensis comb. nov., respectively. Int. J. Syst. Evol. Microbiol. 2020, 70, 6518–6523. [Google Scholar] [CrossRef]
- Lebre, P.H.; Aliyu, H.; De Maayer, P.; Cowan, D.A. In silico characterization of the global Geobacillus and Parageobacillus secretome. Microb. Cell Fact. 2018, 17, 156. [Google Scholar] [CrossRef]
- Aliyu, H.; Mohr, T.; Cowan, D.; de Maayer, P.; Neumann, A. Time-Course transcriptome of Parageobacillus thermoglucosidasius DSM 6285 grown in the presence of carbon monoxide and air. Int. J. Mol. Sci. 2020, 21, 3870. [Google Scholar] [CrossRef] [PubMed]
- Studholme, D.J. Some (bacilli) like it hot: Genomics of Geobacillus species. Microb. Biotechnol. 2015, 8, 40–48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hussein, A.H.; Lisowska, B.K.; Leak, D.J. The genus Geobacillus and their biotechnological potential. Adv. Appl. Microbiol. 2015, 92, 1–48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nadeem, M.S.; Al-Ghamdi, M.A.; Khan, J.A. Studies on the recombinant production in E. coli and characterization of pharmaceutically important thermostable L-asparaginase from Geobacillus thermodenitrificans. Pak. J. Zool. 2019, 51, 1235–1241. [Google Scholar] [CrossRef]
- Alkhalili, R.N.; Bernfur, K.; Dishisha, T.; Mamo, G.; Schelin, J.; Canbäck, B.; Emanuelsson, C.; Hatti-Kaul, R. Antimicrobial protein candidates from the thermophilic Geobacillus sp. strain ZGt-1: Production, proteomics, and bioinformatics analysis. Int. J. Mol. Sci. 2016, 17, 1363. [Google Scholar] [CrossRef] [Green Version]
- Pokusaeva, K.; Kuisiene, N.; Jasinskyte, D.; Rutiene, K.; Saleikiene, J.; Chitavichius, D. Novel bacteriocins produced by Geobacillus stearothermophilus. Open Life Sci. 2009, 4, 196–203. [Google Scholar] [CrossRef]
- Ren, Y.; Strobel, G.; Sears, J.; Park, M. Geobacillus sp., a thermophilic soil bacterium producing volatile antibiotics. Microb. Ecol. 2010, 60, 130–136. [Google Scholar] [CrossRef] [Green Version]
- Gurumurthy, D.M.; Charanraj, T.P.; Faniband, B.; Tallur, P.N.; Bagewadi, Z.K.; Neelagund, S.E.; Mulla, S.I. Cyanoxanthomycin, a bacterial antimicrobial compound extracted from thermophilic Geobacillus sp. Iso5. Jordan J. Biol. Sci. 2020, 13, 725–729. [Google Scholar]
- Schultz, J.; Kallies, R.; Nunes da Rocha, U.; Rosado, A.S. Draft genome sequence of Geobacillus sp. strain LEMMJ02, a thermophile isolated from Deception Island, an active volcano in Antarctica. Microbiol. Resour. Announc. 2019, 8, e00920-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chikindas, M.L.; Weeks, R.; Drider, D.; Chistyakov, V.A.; Dicks, L.M. Functions and emerging applications of bacteriocins. Curr. Opin. Biotechnol. 2018, 49, 23–28. [Google Scholar] [CrossRef]
- Ge, J.; Kang, J.; Ping, W. Effect of acetic acid on bacteriocin production by gram-positive bacteria. J. Microbiol. Biotechnol. 2019, 29, 1341–1348. [Google Scholar] [CrossRef]
- Meade, E.; Slattery, M.A.; Garvery, M. Bacteriocins, potent antimicrobial peptides and the fight against multi drug resistant species: Resistance is futile? Antibiotics 2020, 9, 32. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez, D.; Mavridou, D.A.I. Making the best of aggression: The many dimensions of bacterial toxin regulation. Trends Microbiol. 2019, 27, 897–905. [Google Scholar] [CrossRef] [Green Version]
- Zimina, M.; Babich, O.; Prosekov, A.; Sukhikh, S.; Ivanova, S.; Shevchenko, M.; Noskova, S. Overview of global trends in classification, methods of preparation and application of bacteriocins. Antibiotics 2020, 9, 553. [Google Scholar] [CrossRef]
- Hammami, R.; Zouhir, A.; Naghmouchi, K.; Ben Hamida, J.; Fliss, I. SciDBMaker: New software for computer-aided design of specialized biological databases. BMC Bioinform. 2008, 9, 121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morton, J.T.; Freed, S.D.; Lee, S.W.; Friedberg, I. A large scale prediction of bacteriocin gene blocks suggests a wide functional spectrum for bacteriocins. BMC Bioinform. 2015, 16, 381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, G.; Li, X.; Wang, Z. APD3: The antimicrobial peptide database as a tool for research and education. Nucleic Acids Res. 2016, 44, D1087–D1093. [Google Scholar] [CrossRef] [Green Version]
- Yount, N.Y.; Weaver, D.C.; de Anda, J.; Lee, E.Y.; Lee, M.W.; Wong, G.C.L.; Yeaman, M.R. Discovery of novel Type II bacteriocins using a new high-dimensional bioinformatic algorithm. Front. Immunol. 2020, 11, 1873. [Google Scholar] [CrossRef] [PubMed]
- Cui, Y.; Luo, L.; Wang, X.; Lu, Y.; Yi, Y.; Shan, Y.; Liu, B.; Zhou, Y.; Lü, X. Mining, Heterologous Expression, Purification, Antibactericidal Mechanism, and Application of Bacteriocins: A Review. Compr. Rev. Food Sci. Food Saf. 2021, 20, 863–899. [Google Scholar] [CrossRef]
- Cheigh, C.-I.; Pyun, Y.-R. Nisin biosynthesis and its properties. Biotechnol. Lett. 2005, 27, 1641–1648. [Google Scholar] [CrossRef]
- Egan, K.; Field, D.; Rea, M.C.; Ross, R.P.; Hill, C.; Cotter, P.D. Bacteriocins: Novel solutions to age old spore-related problems? Front. Microbiol. 2016, 7, 461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smaoui, S.; Elleuch, L.; Bejar, W.; Karray-Rebai, I.; Ayadi, I.; Jaouadi, B.; Mathieu, F.; Chouayekh, H.; Bejar, S.; Mellouli, L. Inhibition of fungi and gram-negative bacteria by bacteriocin BacTN635 produced by Lactobacillus plantarum sp. TN635. Appl. Biochem. Biotechnol. 2010, 162, 1132–1146. [Google Scholar] [CrossRef] [Green Version]
- Sang, Y.; Blecha, F. Antimicrobial peptides and bacteriocins: Alternatives to traditional antibiotics. Anim. Health Res. Rev. 2008, 9, 227–235. [Google Scholar] [CrossRef] [Green Version]
- Cotter, P.D.; Ross, R.P.; Hill, C. Bacteriocins—A viable alternative to antibiotics? Nat. Rev. Microbiol. 2013, 11, 95–105. [Google Scholar] [CrossRef]
- Newstead, L.L.; Varjonen, K.; Nuttall, T.; Paterson, G.K. Staphylococcal-produced bacteriocins and antimicrobial peptides: Their potential as alternative treatments for Staphylococcus aureus infections. Antibiotics 2020, 9, 40. [Google Scholar] [CrossRef] [Green Version]
- Ovchinnikov, K.V.; Kranjec, C.; Thorstensen, T.; Carlsen, H.; Diep, D.B. Successful development of bacteriocins into therapeutic formulation for treatment of MRSA skin infection in a murine model. Antimicrob. Agents Chemother. 2020, 64, e00829-20. [Google Scholar] [CrossRef]
- Simons, A.; Alhanout, K.; Duval, R.E. Bacteriocins, antimicrobial peptides from bacterial origin: Overview of their biology and their impact against multidrug-resistant bacteria. Microorganisms 2020, 8, 639. [Google Scholar] [CrossRef] [PubMed]
- Lopetuso, L.R.; Giorgio, M.E.; Saviano, A.; Scaldaferri, F.; Gasbarrini, A.; Cammarota, G. Bacteriocins and bacteriophages: Therapeutic weapons for gastrointestinal diseases? Int. J. Mol. Sci. 2019, 20, 183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heilbronner, S.; Krismer, B.; Brötz-Oesterhelt, H.; Peschel, A. The microbiome-shaping roles of bacteriocins. Nat. Rev. Microbiol. 2021, 19, 726–739. [Google Scholar] [CrossRef]
- Garg, N.; Oman, T.J.; Andrew Wang, T.-S.; De Gonzalo, C.V.G.; Walker, S.; van der Donk, W.A. Mode of action and structure–activity relationship studies of geobacillin I. J. Antibiot. 2014, 67, 133–136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garg, N.; Tang, W.; Goto, Y.; Nair, S.K.; van der Donk, W.A. Lantibiotics from Geobacillus thermodenitrificans. Proc. Natl. Acad. Sci. USA 2012, 109, 5241–5246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garg, N.; Goto, Y.; Chen, T.; van der Donk, W.A. Characterization of the stereochemical configuration of lanthionines formed by the lanthipeptide synthetase GeoM. Pept. Sci. 2016, 106, 834–842. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ceylan, A.; Başbülbül, G. The use of bacteriocin produced by Geobacillus toebii HBB 218 to prevent the growth of Bacillus coagulans and Geobacillus thermophilus in canned food. Türk Mikrobiyol. Cemiy. Derg. 2019, 49, 140–146. [Google Scholar] [CrossRef] [Green Version]
- Novotny, J.F.; Perry, J.J. Characterization of bacteriocins from two strains of Bacillus thermoleovorans, a thermophilic hydrocarbon-utilizing species. Appl. Environ. Microbiol. 1992, 58, 2393–2396. [Google Scholar] [CrossRef] [Green Version]
- Vaičikauskaitė, M.; Ger, M.; Valius, M.; Maneikis, A.; Lastauskienė, E.; Kalėdienė, L.; Kaunietis, A. Geobacillin 26—High molecular weight bacteriocin from a thermophilic bacterium. Int. J. Biol. Macromol. 2019, 141, 333–344. [Google Scholar] [CrossRef] [PubMed]
- Sharp, R.J.; Bingham, A.H.A.; Comer, M.J.; Atkinson, A.Y. 1979 Partial characterization of a bacteriocin (thermocin) from Bacillus stearothermophilus RS93. Microbiology 1979, 111, 449–451. [Google Scholar] [CrossRef] [Green Version]
- Yule, R.; Barridge, B.D. Isolation and characterization of a bacteriocin produced by Bacillus stearothermophilus strain NU-10. Can. J. Microbiol. 1976, 22, 1743–1750. [Google Scholar] [CrossRef] [PubMed]
- Fikes, J.D.; Crabtree, B.L.; Barridge, B.D. Studies on the mode of action of a bacteriocin produced by Bacillus stearothermophilus. Can. J. Microbiol. 1983, 29, 1576–1582. [Google Scholar] [CrossRef] [PubMed]
- Pranckute, R.; Kaunietis, A.; Kananavičiute, R.; Lebedeva, J.; Kuisiene, N.; Šaleikiene, J.; Čitavičius, D. Differences of antibacterial activity spectra and properties of bacteriocins, produced by Geobacillus sp. bacteria isolated from different environments. J. Microbiol. Biotechnol. Food Sci. 2015, 5, 155–161. [Google Scholar] [CrossRef] [Green Version]
- Kaunietis, A.; Pranckute, R.; Lastauskienė, E.; Čitavičius, D. Medium optimization for bacteriocin production and bacterial cell growth of Geobacillus sp. 15 strain. J. Antimicrob. Agents 2017, 3, 1. [Google Scholar] [CrossRef] [Green Version]
- Shafia, F. Thermocins of Bacillus stearothermophilus. J. Bacteriol. 1966, 92, 524–525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Egan, K.; Kelleher, P.; Field, D.; Rea, M.C.; Ross, R.P.; Cotter, P.D.; Hill, C. Genome sequence of Geobacillus stearothermophilus DSM 458, an antimicrobial-producing thermophilic bacterium, isolated from a sugar beet factory. Genome Announc. 2017, 5, e01172-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, X.; Kuipers, O.P. Identification and classification of known and putative antimicrobial compounds produced by a wide variety of Bacillales species. BMC Genom. 2016, 17, 882. [Google Scholar] [CrossRef] [Green Version]
- Egan, K.; Field, D.; Ross, R.P.; Cotter, P.D.; Hill, C. In silico prediction and exploration of potential bacteriocin gene clusters within the bacterial genus Geobacillus. Front. Microbiol. 2018, 9, 2116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walker, M.C.; Eslami, S.M.; Hetrick, K.J.; Ackenhusen, S.E.; Mitchell, D.A.; van der Donk, W.A. Precursor peptide-targeted mining of more than one hundred thousand genomes expands the lanthipeptide natural product family. BMC Genom. 2020, 21, 387. [Google Scholar] [CrossRef]
- Alkhalili, R.N.; Canbäck, B. Identification of putative novel class-I lanthipeptides in Firmicutes: A combinatorial in silico analysis approach performed on genome sequenced bacteria and a close inspection of Z-geobacillin lanthipeptide biosynthesis gene cluster of the thermophilic Geobacillus sp. strain ZGt-1. Int. J. Mol. Sci. 2018, 19, 2650. [Google Scholar] [CrossRef] [Green Version]
- Zacharof, M.P.; Lovitt, R.W. Bacteriocins produced by lactic acid bacteria—A review article. APCBEE Procedia 2012, 2, 50–56. [Google Scholar] [CrossRef] [Green Version]
- Gurban oglu Gulahmadov, S.; Batdorj, B.; Dalgalarrondo, M.; Chobert, J.M.; Alekper oglu Kuliev, A.; Haertlé, T. Characterization of bacteriocin-like inhibitory substances (BLIS) from lactic acid bacteria isolated from traditional Azerbaijani cheeses. Eur. Food Res. Technol. 2006, 224, 229–235. [Google Scholar] [CrossRef]
- Başbülbül Özdemir, G.; Biyik, H.H. Isolation and characterization of a bacteriocin-like substance produced by Geobacillus toebii strain HBB-247. Indian J. Microbiol. 2011, 52, 104–108. [Google Scholar] [CrossRef] [Green Version]
- Muhammad, S.A.; Ahmed, S.; Ismail, T.; Hameed, A. Taguchi’s experimental design for optimizing the production of novel thermostable polypeptide antibiotic from Geobacillus pallidus SAT4. Pak. J. Pharm. Sci. 2014, 27, 11–23. [Google Scholar]
- Ołdak, A.; Zielińska, D. Bakteriocyny bakterii fermentacji mlekowej jako alternatywa antybiotyków. Postepy Hig. Med. Dosw. 2017, 71, 328–338. [Google Scholar]
- Van Loveren, H.; Sanz, Y.; Salminen, S. Health claims in Europe: Probiotics and prebiotics as case examples. Annu. Rev. Food Sci. Technol. 2012, 3, 247–261. [Google Scholar] [CrossRef] [PubMed]
- Araya, M.; Gopal, P.; Lindgren, S.E.; Lodi, R.; Oliver, G.; Saxelin, M.L.; Servin, A.L.; Stanton, C.; Gilliland, S.E.; Morelli, L.; et al. Joint FAO/WHO expert consultation on evaluation of health and nutritional properties of probiotics in food including powder milk with live lactic acid bacteria. In Probiotics in Food: Health and Nutritional Properties and Guidelines for Evaluation; FAO Food and Nutrition paper 85; FAO: Rome, Italy, 2001; Available online: http://www.fao.org/3/a-a0512e.pdf (accessed on 11 February 2022).
- Salminen, S.; Ouwehand, A.; Benno, Y.; Lee, Y.-K. Probiotics: How should they be defined? Trends Food Sci. Technol. 1999, 10, 107–110. [Google Scholar] [CrossRef]
- Siragusa, G. Modern Probiology—Direct Fed Microbials and the Avian Gut Microbiota. In Proceedings of the 23rd Annual Australian Poultry Science Symposium, Sydney, NSW, Australia, 19–22 February 2012. [Google Scholar]
- Vargas-Albores, F.; Porchas-Cornejo, M.A.; Martínez-Porchas, M.; Villalpando-Canchola, E.; Gollas-Galván, T.; Martínez-Córdova, L.R. Bacterial biota of shrimp intestine is significantly modified by the use of a probiotic mixture: A high throughput sequencing approach. Helgol. Mar. Res. 2017, 71, 5. [Google Scholar] [CrossRef] [Green Version]
- Jha, R.; Das, R.; Oak, S.; Mishra, P. Probiotics (Direct-Fed Microbials) in poultry nutrition and their effects on nutrient utilization, growth and laying performance, and gut health: A systematic review. Animals 2020, 10, 1863. [Google Scholar] [CrossRef]
- Susanti, D.; Volland, A.; Tawari, N.; Baxter, N.; Gangaiah, D.; Plata, G.; Nagireddy, A.; Hawkins, T.; Mane, S.P.; Kumar, A. Multi-Omics characterization of host-derived Bacillus spp. probiotics for improved growth performance in poultry. Front. Microbiol. 2021, 12, 747845. [Google Scholar] [CrossRef]
- Łubkowska, B.; Jeżewska-Frąckowiak, J.; Sobolewski, I.; Skowron, P.M. Bacteriophages of thermophilic ‘bacillus group’ bacteria—A review. Microorganisms 2021, 9, 1522. [Google Scholar] [CrossRef]
- Mahdhi, A.; Hmila, Z.; Behi, A.; Bakhrouf, A. Preliminary characterization of the probiotic properties of Candida famata and Geobacillus thermoleovorans. Iran. J. Microbiol. 2011, 3, 129–134. [Google Scholar] [PubMed]
- McMullan, G.; Christie, J.M.; Rahman, T.J.; Banat, I.M.; Ternan, N.G.; Marchant, R. Habitat, applications and genomics of the aerobic, thermophilic genus Geobacillus. Biochem. Soc. Trans. 2004, 32, 214–217. [Google Scholar] [CrossRef] [Green Version]
- Luo, Y.; Korza, G.; De Marco, A.M.; Kuipers, O.P.; Li, Y.Q.; Setlow, P. Properties of spores of Bacillus subtilis with or without a transposon that decreases spore germination and increases spore wet heat resistance. J. Appl. Microbiol. 2021, 131, 2918–2928. [Google Scholar] [CrossRef]
- Sing, D.; Sing, C.F. Impact of direct soil exposures from airborne dust and geophagy on human health. Int. J. Environ. Res. Public Health 2010, 7, 1205–1223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nicholson, W.L. Using thermal inactivation kinetics to calculate the probability of extreme spore longevity: Implications for paleomicrobiology and lithopanspermia. Orig. Life Evol. Biosph. 2003, 33, 621–631. [Google Scholar] [CrossRef]
- Ketch, L.A.; Malloch, D.; Mahaney, W.C.; Huffman, M.A. Comparative microbial analysis and clay mineralogy of soils eaten by chimpanzees (Pan troglodytes schweinfurthii) in Tanzania. Soil Biol. Biochem. 2001, 33, 199–203. [Google Scholar] [CrossRef]
- Bisi-Johnson, M.A.; Obi, C.L.; Ekosse, G.E. Microbiological and health related perspectives of geophagia: An overview. Afr. J. Biotechnol. 2010, 9, 5784–5791. Available online: http://www.academicjournals.org/AJB (accessed on 11 February 2022).
- Nyanza, E.C.; Joseph, M.; Premji, S.S.; Thomas, D.S.; Mannion, C. Geophagy practices and the content of chemical elements in the soil eaten by pregnant women in artisanal and small scale gold mining communities in Tanzania. BMC Pregnancy Childbirth 2014, 14, 144. [Google Scholar] [CrossRef] [Green Version]
- Huebl, L.; Leick, S.; Guettl, L.; Akello, G.; Kutalek, R. Geophagy in Northern Uganda: Perspectives from consumers and clinicians. Am. J. Trop. Med. Hyg. 2016, 95, 1440–1449. [Google Scholar] [CrossRef]
- Golokhvast, K.; Sergievich, A.; Grigoriev, N. Geophagy (rock eating), experimental stress and cognitive idiosyncrasy. Asian Pac. J. Trop. Biomed. 2014, 4, 362–366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blum, W.E.H.; Zechmeister-Boltenstern, S.; Keiblinger, K.M. Does Soil Contribute to the Human Gut Microbiome? Microorganisms 2019, 7, 287. [Google Scholar] [CrossRef] [Green Version]
- Hirt, H. Healthy soils for healthy plants for healthy humans: How beneficial microbes in the soil, food and gut are interconnected and how agriculture can contribute to human health. EMBO Rep. 2020, 21, e51069. [Google Scholar] [CrossRef]
- Abbasi, A.; Sheykhsaran, E.; Kafil, H.S. Obstacles and challenges in the use of probiotics. In Postbiotics: Science, Technology and Applications; Bentham Science Publishers: Al Sharjah, United Arab Emirates, 2021. [Google Scholar] [CrossRef]
- Żółkiewicz, J.; Marzec, A.; Ruszczyński, M.; Feleszko, W. Postbiotics-A step beyond pre- and probiotics. Nutrients 2020, 12, 2189. [Google Scholar] [CrossRef]
- Mande, S.S.; Sarfaty, S.; Allen, M.D.; Perham, R.N.; Hol, W.G.J. Protein-protein interactions in the pyruvate dehydrogenase multienzyme complex: Dihydrolipoamide dehydrogenase complexed with the binding domain of dihydrolipoamide acetyltransferase. Structure 1996, 4, 277–286. [Google Scholar] [CrossRef] [Green Version]
- Henderson, C.E.; Perham, R.N. Purification of the pyruvate dehydrogenase multienzyme complex of Bacillus stearothermophilus and resolution of its four component polypeptides. Biochem. J. 1980, 189, 161–172. [Google Scholar] [CrossRef] [Green Version]
- Domingo, G.J.; Caivano, A.; Sartorius, R.; Barba, P.; Bäckström, M.; Piatier-Tonneau, D.; Guardiola, J.; De Berardinis, P.; Perham, R.N. Induction of specific T-helper and cytolytic responses to epitopes displayed on a virus-like protein scaffold derived from the pyruvate dehydrogenase multienzyme complex. Vaccine 2003, 21, 1502–1509. [Google Scholar] [CrossRef]
- Domingo, G.J.; Orru’, S.; Perham, R.N. Multiple display of peptides and proteins on a macromolecular scaffold derived from a multienzyme complex. J. Mol. Biol. 2001, 305, 259–267. [Google Scholar] [CrossRef] [PubMed]
- D’Apice, L.; Sartorius, R.; Caivano, A.; Mascolo, D.; Del Pozzo, G.; Di Mase, D.S.; Ricca, E.; Li Pira, G.; Manca, F.; Malanga, D.; et al. Comparative analysis of new innovative vaccine formulations based on the use of procaryotic display systems. Vaccine 2007, 25, 1993–2000. [Google Scholar] [CrossRef] [PubMed]
- Allen, M.D.; Perham, R.N. The catalytic domain of dihydrolipoyl acetyltransferase from the pyruvate dehydrogenase multienzyme complex of Bacillus stearothermophilus. Expression, purification and reversible denaturation. FEBS Lett. 1997, 413, 339–343. [Google Scholar] [CrossRef] [Green Version]
- Howard, M.J.; Chauhan, H.J.; Domingo, G.J.; Fuller, C.; Perham, R.N. Protein-protein interaction revealed by NMR T2 relaxation experiments: The lipoyl domain and E1 component of the pyruvate dehydrogenase multienzyme complex of Bacillus stearothermophilus. J. Mol. Biol. 2000, 295, 1023–1037. [Google Scholar] [CrossRef]
- Trovato, M. Novel antigen delivery systems. World J. Virol. 2015, 4, 156. [Google Scholar] [CrossRef]
- Trovato, M. Delivery strategies for novel vaccine formulations. World J. Virol. 2012, 1, 4. [Google Scholar] [CrossRef] [PubMed]
- Peng, T.; Lee, H.; Lim, S. Isolating a trimer intermediate in the self-assembly of E2 protein cage. Biomacromolecules 2012, 13, 699–705. [Google Scholar] [CrossRef]
- Ren, D.; Dalmau, M.; Randall, A.; Shindel, M.M.; Baldi, P.; Wang, S.-W. Biomimetic design of protein nanomaterials for hydrophobic molecular transport. Adv. Funct. Mater. 2012, 15, 3170–3180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krebs, S.J.; McBurney, S.P.; Kovarik, D.N.; Waddell, C.D.; Jaworski, J.P.; Sutton, W.F.; Gomes, M.M.; Trovato, M.; Waagmeester, G.; Barnett, S.J.; et al. Multimeric scaffolds displaying the HIV-1 envelope MPER induce MPER-specific antibodies and cross-neutralizing antibodies when co-immunized with gp160 DNA. PLoS ONE 2014, 9, e113463, Erratum in PLoS ONE 2015, 10, e0120027. [Google Scholar] [CrossRef] [PubMed]
- Jaworski, J.P.; Krebs, S.J.; Trovato, M.; Kovarik, D.N.; Brower, Z.; Sutton, W.F.; Waagmeester, G.; Sartorius, R.; D’Apice, L.; Caivano, A.; et al. Co-immunization with multimeric scaffolds and DNA rapidly induces potent autologous HIV-1 neutralizing antibodies and CD8+ T cells. PLoS ONE 2012, 7, e31464. [Google Scholar] [CrossRef] [Green Version]
- Caivano, A.; Doria-Rose, N.A.; Buelow, B.; Sartorius, R.; Trovato, M.; D’Apice, L.; Domingo, G.J.; Sutton, W.F.; Haigwood, N.L.; De Berardinis, P. HIV-1 Gag p17 presented as virus-like particles on the E2 scaffold from Geobacillus stearothermophilus induces sustained humoral and cellular immune responses in the absence of IFNγ production by CD4+ T cells. Virology 2010, 407, 296–305. [Google Scholar] [CrossRef] [Green Version]
- Sun, Q.; Chen, Q.; Blackstock, D.; Chen, W. Post-translational modification of bionanoparticles as a modular platform for biosensor assembly. ACS Nano 2015, 9, 8554–8561. [Google Scholar] [CrossRef]
- Wichgers Schreur, P.J.; Tacken, M.; Gutjahr, B.; Keller, M.; van Keulen, L.; Kant, J.; van de Water, S.; Lin, Y.; Eiden, M.; Rissmann, M.; et al. Vaccine efficacy of self-assembled multimeric protein scaffold particles displaying the glycoprotein Gn Head Domain of Rift Valley Fever Virus. Vaccines 2021, 9, 301. [Google Scholar] [CrossRef] [PubMed]
- Hankin, E.H. L’action bactericide des eaux de la Jumna et du Gange sur le vibrion du cholera. Ann. Inst. Pasteur 1896, 10, 511. [Google Scholar]
- Dublanchet, A.; Bourne, S. The epic of phage therapy. Can. J. Infect. Dis. Med. Microbiol. 2007, 18, 15–18. [Google Scholar] [CrossRef] [Green Version]
- Abedon, S.T.; Kuhl, S.J.; Blasdel, B.G.; Kutter, E.M. Phage treatment of human infections. Bacteriophage 2011, 1, 66–85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reuter, M.; Kruger, D.H. Approaches to optimize therapeutic bacteriophage and bacteriophage-derived products to combat bacterial infections. Virus Genes 2020, 56, 136–149. [Google Scholar] [CrossRef]
- Jun, S.Y.; Jang, I.J.; Yoon, S.; Jang, K.; Yu, K.S.; Cho, J.Y.; Seong, M.W.; Jung, G.M.; Yoon, S.J.; Kang, S.H. Pharmacokinetics and tolerance of the hage endolysin-based candidate drug SAL200 after a single intravenous administration among healthy volunteers. Antimicrob. Agents Chemother. 2017, 61, e02629-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- González-Mora, A.; Hernández-Pérez, J.; Iqbal, H.M.N.; Rito-Palomares, M.; Benavides, J. Bacteriophage-based vaccines: A potent approach for antigen delivery. Vaccines 2020, 8, 504. [Google Scholar] [CrossRef] [PubMed]
- Woźnica, M.; Bigos, J.; Łobocka, M. Lysis of bacterial cells in the process of bacteriophage release—Canonical and newly discovered mechanisms. Postepy Hig. Med. Dosw. 2015, 69, 114–126. [Google Scholar]
- Oliveira, H.; Azeredo, J.; Lavigne, R.; Kluskens, L.D. Bacteriophage endolysins as a response to emerging foodborne pathogens. Trends Food Sci. Technol. 2012, 28, 103–115. [Google Scholar] [CrossRef] [Green Version]
- Borysowski, J.; Weber-Da.browska, B.; Górski, A. Bacteriophage endolysins as a novel class of antibacterial agents. Exp. Biol. Med. 2006, 231, 366–377. [Google Scholar] [CrossRef]
- Young, R. Phage lysis: Three steps, three choices, one outcome. J. Microbiol. 2014, 52, 243–258. [Google Scholar] [CrossRef]
- Maciejewska, B.; Olszak, T.; Drulis-Kawa, Z. Applications of bacteriophages versus phage enzymes to combat and cure bacterial infections: An ambitious and also a realistic application? Appl. Microbiol. Biotechnol. 2018, 102, 2563–2581. [Google Scholar] [CrossRef] [Green Version]
- Schmelcher, M.; Donovan, D.M.; Loessner, M.J. Bacteriophage endolysins as novel antimicrobials. Future Microbiol. 2012, 7, 1147–1171. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, H.; Melo, L.D.R.; Santos, S.B.; Nobrega, F.L.; Ferreira, E.C.; Cerca, N.; Azeredo, J.; Kluskens, L.D. Molecular aspects and comparative genomics of bacteriophage endolysins. J. Virol. 2013, 87, 4558–4570. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, H.; São-José, C.; Azeredo, J. Phage-derived peptidoglycan degrading enzymes: Challenges and future prospects for in vivo therapy. Viruses 2018, 10, 292. [Google Scholar] [CrossRef] [Green Version]
- Mirski, T.; Mizak, L.; Nakonieczna, A.; Gryko, R. Bacteriophages, phage endolysins and antimicrobial peptides—The possibilities for their common use to combat infections and in the design of new drugs. Ann. Agric. Environ. Med. 2019, 26, 203–209. [Google Scholar] [CrossRef] [PubMed]
- Nelson, D.C.; Schmelcher, M.; Rodriguez-Rubio, L.; Klumpp, J.; Pritchard, D.G.; Dong, S.; Donovan, D.M. Endolysins as antimicrobials. Adv. Virus Res. 2012, 83, 299–365. [Google Scholar] [CrossRef] [Green Version]
- Rahman, M.; Wang, W.; Sun, Q.; Shah, J.A.; Li, C.; Sun, Y.; Li, Y.; Zhang, B.; Chen, W.; Wang, S. Endolysin, a promising solution against antimicrobial resistance. Antibiotics 2021, 10, 1277. [Google Scholar] [CrossRef] [PubMed]
- Jin, M.; Ye, T.; Zhang, X. Roles of bacteriophage GVE2 endolysin in host lysis at high temperatures. Microbiology 2013, 159, 1597–1605. [Google Scholar] [CrossRef] [Green Version]
- Swift, S.M.; Reid, K.P.; Donovan, D.M.; Ramsay, T.G. Thermophile lytic enzyme fusion proteins that target Clostridium perfringens. Antibiotics 2019, 8, 214. [Google Scholar] [CrossRef] [Green Version]
- Miller, E.; Warek, U.; Xu, D. Endolysin from Bacteriophage against Geobacillus and Methods of Using. International Patent Application WIPO/PCT WO 2016/123425 Al, PCT/US2016/015516, 2016. [Google Scholar]
- Van Zyl, L.J.; Sunda, F.; Taylor, M.P.; Cowan, D.A.; Trindade, M.I. Identification and characterization of a novel Geobacillus thermoglucosidasius bacteriophage, GVE3. Arch. Virol. 2015, 160, 2269–2282. [Google Scholar] [CrossRef] [Green Version]
- Skowron, P.M.; Kropinski, A.M.; Zebrowska, J.; Janus, L.; Szemiako, K.; Czajkowska, E.; Maciejewska, N.; Skowron, M.; Łoś, J.; Łoś, M.; et al. Sequence, genome organization, annotation and proteomics of the thermophilic, 47.7-kb Geobacillus stearothermophilus bacteriophage TP-84 and its classification in the new Tp84virus genus. PLoS ONE 2018, 13, e0195449, Erratum in PLoS ONE 2018, 13, e0196798. [Google Scholar] [CrossRef]
- Plotka, M.; Kaczorowska, A.K.; Stefanska, A.; Morzywolek, A.; Fridjonsson, O.H.; Dunin-Horkawicz, S.; Kozlowski, L.; Hreggvidsson, G.O.; Kristjansson, J.K.; Dabrowski, S.; et al. Novel highly thermostable endolysin from Thermus scotoductus MAT2119 bacteriophage Ph2119 with amino acid sequence similarity to eukaryotic peptidoglycan recognition proteins. Appl. Environ. Microbiol. 2014, 80, 886–895. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murray, E.; Draper, L.A.; Ross, R.P.; Hill, C. The advantages and challenges of using endolysins in a clinical setting. Viruses 2021, 13, 680. [Google Scholar] [CrossRef]
- Miner-Williams, W.M.; Stevens, B.R.; Moughan, P.J. Are intact peptides absorbed from the healthy gut in the adult human? Nutr. Res. Rev. 2014, 27, 308–329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fallingborg, J. Intraluminal pH of the human gastrointestinal tract. Dan. Med. Bull. 1999, 46, 183–196. [Google Scholar] [PubMed]
- Jun, S.Y.; Jung, G.M.; Yoon, S.J.; Youm, S.Y.; Han, H.Y.; Lee, J.H.; Kang, S.H. Pharmacokinetics of the phage endolysin-based candidate drug SAL200 in monkeys and its appropriate intravenous dosing period. Clin. Exp. Pharmacol. Physiol. 2016, 43, 1013–1016. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
| Bacteriocin | Parent Strain | Determined Antimicrobial Bioactivity against Mesophilic Strains | Properties | Reference |
|---|---|---|---|---|
| geobacillin I | G. thermodenitrificans NG-80-2 G. thermodenitrificans DSM465 G. thermodenitrificans OHT-1 Geobacillus sp. M10EXG G. thermodenitrificans OH2-1 G. thermodenitrificans OH5-2 | Streptococcus dysgalactiae Vancomycin-resistant Enterococcus faecium Methicillin-resistant S. aureus B. anthracis Sterne 7702 | nisin analog; the N-terminal structure resembles nisin rings; the mode of action includes the formation of the pores in the cell membrane; comparable bioactivity range to nisin; higher pH and temperature stability than nisin; | [42] |
| geobacillin II | G. thermodenitrificans NG-80-2 Geobacillus sp. G11MC16 | B. cereus Z4222 B. subtilis | nisin analog; the N-terminal structure resembles nisin rings; antimicrobial activity towards Bacillus spp. only; | [42] |
| toebicin 218 | P. toebii strain HBB-218 | B. coagulans DSM 1 | ND | [44] |
| thermoleovorin-S2 | G. thermoleovorans S-II | Streptococcus faecalis (S. faecalis) S. typhimurium Branhamella catarrhalis | high molecular weight bacteriocin: 42 kDa; stability over a wide pH (pH 3–10); activity at 60 °C and at 70 °C; partial activity at 80 °C; antimicrobial activity towards G. thermoleovorans (except the host strain) | [45] |
| thermoleovorin-N9 | G. thermoleovorans NR-9 | S. faecalis S. typhimurium Branhamella catarrhalis | high molecular weight bacteriocin: 36 kDa; stability over a wide pH range (pH 3–10); activity at 60 °C; antimicrobial activity towards G. thermoleovorans (except the host strain); | [45] |
| geobacillin 26 | G. stearothermophilus 15 | ND | stable at 50 °C only for 2 h; probable mesophilic origin or HGT; no effect on the tested Candida and Staphylococcus strains; may belong to non-lytic bacteriocins and its mode of action may require the binding to external cell receptors and dissipation of the membrane receptors; antimicrobial activity towards Geobacillus and Parageobacillus strains only; | [46] |
| thermocin 32A | G. stearothermophilus 32A | B. cereus DSM 12001 S. haemolyticus P903 | low molecular weight bacteriocins: 5.6–7.2 kDa; activity towards a closely related strain of Geobacillus; extremely high thermostability (no activity loss observed after incubation at 100 °C); activity at the pH range 4–10; | [17] |
| thermocin 17 | G. stearothermophilus 17 | B. cereus DSM 12001 | ||
| thermocin 30 | G. stearothermophilus 30 | S. haemolyticus P903 P. aeruginosa ATCC 27853 | ||
| thermocin 93 | G. stearothermophilus RS93 | B. subtilis ATCC 10783 | molecular weight: 13.5 kDa; antimicrobial activity towards ten Geobacillus strains; optimal activity at pH 7; no activity loss observed after incubation at 70 °C; the mode of action could not be elucidated due to variable results; | [47] |
| thermocin 10 | G. stearothermophilus NU-10 | B. circulans 4516 | low molecular weight: 20 kDa; remains active in the pH range 2–12 at 70 °C; the mode of action probably linked with RNA inactivation or cell membrane structure impairing; | [48,49] |
| unnamed bacteriocines 1 | 101 strains of Geobacillus spp. from the culture collection of the Department of Microbiology and Biotechnology of Vilnius University | Streptococcus pyogenes ATCC 19615 Streptococcus pneumoniae ATCC 6305 E. faecalis ATCC 2912 Enterococcus faecium 402-3/03 Haemophilus influenzae ATCC 10211 S. aureus ATCC 25923 S. haemolyticus P903 S. epidermidis ATCC 12228 P. aeruginosa ATCC 27853 E. coli ATCC 25922 Klebsiella pneumoniae DSM 30104 Yersinia enterocolitica ATCC 9610 S. typhimurium ATCC 14028 S. enteritidis ATCC 13076 Listeria monocytogenes ATCC 19117 Listeria innocua ATCC 33090 B. cereus DSM 12001 B. subtilis ATCC 6633 Clostridium perfringens ATCC 13124 | ND | [50] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zebrowska, J.; Witkowska, M.; Struck, A.; Laszuk, P.E.; Raczuk, E.; Ponikowska, M.; Skowron, P.M.; Zylicz-Stachula, A. Antimicrobial Potential of the Genera Geobacillus and Parageobacillus, as Well as Endolysins Biosynthesized by Their Bacteriophages. Antibiotics 2022, 11, 242. https://doi.org/10.3390/antibiotics11020242
Zebrowska J, Witkowska M, Struck A, Laszuk PE, Raczuk E, Ponikowska M, Skowron PM, Zylicz-Stachula A. Antimicrobial Potential of the Genera Geobacillus and Parageobacillus, as Well as Endolysins Biosynthesized by Their Bacteriophages. Antibiotics. 2022; 11(2):242. https://doi.org/10.3390/antibiotics11020242
Chicago/Turabian StyleZebrowska, Joanna, Małgorzata Witkowska, Anna Struck, Patrycja E. Laszuk, Edyta Raczuk, Małgorzata Ponikowska, Piotr M. Skowron, and Agnieszka Zylicz-Stachula. 2022. "Antimicrobial Potential of the Genera Geobacillus and Parageobacillus, as Well as Endolysins Biosynthesized by Their Bacteriophages" Antibiotics 11, no. 2: 242. https://doi.org/10.3390/antibiotics11020242
APA StyleZebrowska, J., Witkowska, M., Struck, A., Laszuk, P. E., Raczuk, E., Ponikowska, M., Skowron, P. M., & Zylicz-Stachula, A. (2022). Antimicrobial Potential of the Genera Geobacillus and Parageobacillus, as Well as Endolysins Biosynthesized by Their Bacteriophages. Antibiotics, 11(2), 242. https://doi.org/10.3390/antibiotics11020242