
Transcriptome Analysis of the Response of Mature Helicobacter pylori Biofilm to Different Doses of Lactobacillus salivarius LN12 with Amoxicillin and Clarithromycin


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Effects of CFSs of Different Doses of L. salivarus LN12 in Combination with AMX and CLR on the Biomass of H. pylori Biofilms
2.2. Effects of CFSs of Different Doses of L. salivarus LN12 in Combination with AMX and CLR on the Viability of H. pylori Biofilms
2.3. Effects of CFSs of Different Doses of L. salivarus LN12 in Combination with AMX and CLR on the Structure of H. pylori Biofilms
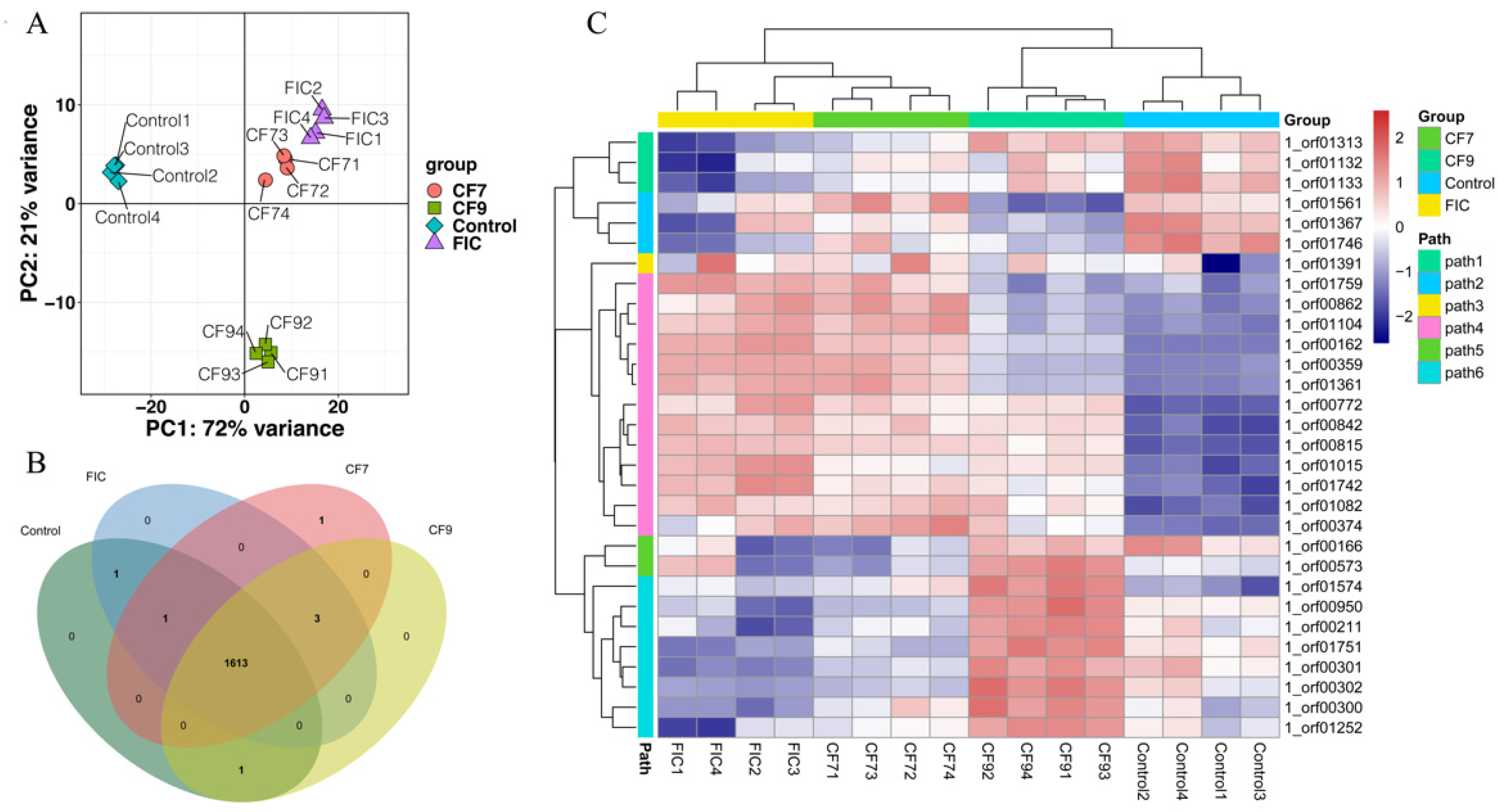
2.4. Transcriptome Analysis
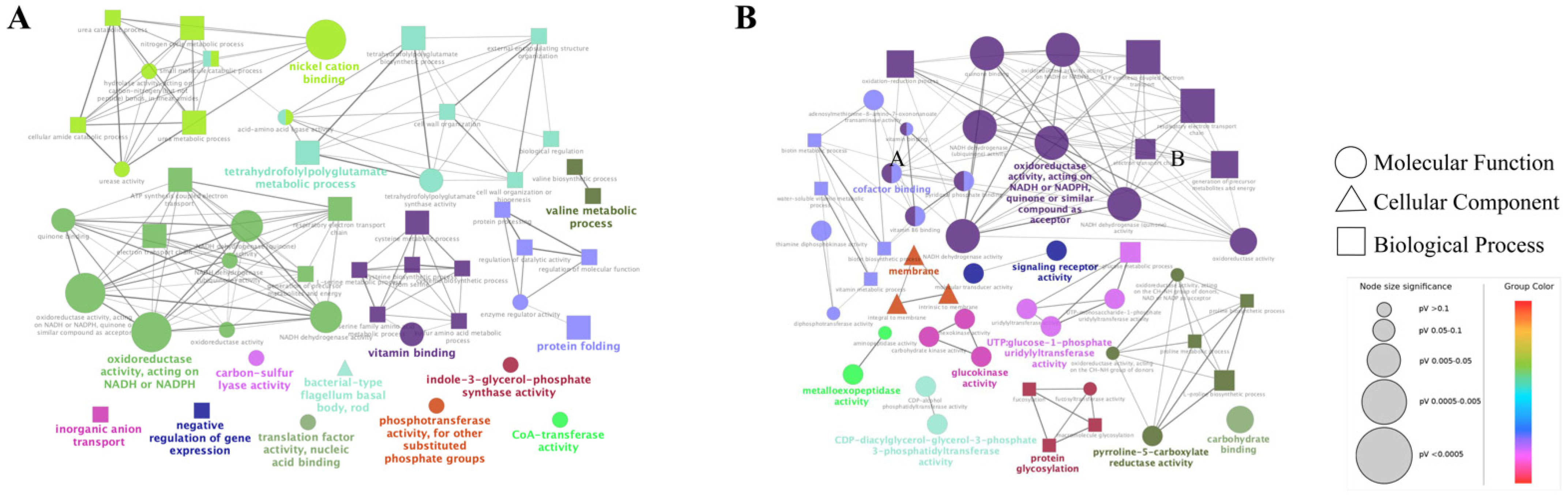
2.4.1. GO Enrichment Analysis
2.4.2. KEGG Enrichment Analysis
2.4.3. RT-qPCR Validation
3. Discussion
4. Materials and Methods
4.1. Strains and Culture Conditions
4.2. H. pylori Biofilm Culture
4.3. Preparation of the Combination of Lactobacillus Salivarius LN12 Cell-Free Supernatant (CFS) and Antibiotics
4.4. Effects of Different Treatments on the Viability of H. pylori Biofilms
4.5. Effects of Different Treatments on the Biomass of H. pylori Biofilms
4.6. Effects of Different Treatments on the Structure of H. pylori Biofilm
4.7. Preparation of RNA-Seq Sequencing Library
4.8. Bioinformatics Analysis
4.9. RT-qPCR Validation
4.10. Data Analysis and Availability
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Chmiela, M.; Kupcinskas, J. Review: Pathogenesis of Helicobacter pylori infection. Helicobacter 2019, 24 (Suppl. 1), e12638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, Y.; Zhu, Y.; Lu, N.H. Recent progress in Helicobacter pylori treatment. Chin. Med. J.-Peking 2020, 133, 335–343. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Meng, X.; Li, Y.; Zhao, C.N.; Tang, G.Y.; Li, S.; Gan, R.Y.; Li, H.B. Natural Products for the Prevention and Management of Helicobacter pylori Infection. Compr. Rev. Food Sci. Food Saf. 2018, 17, 937–952. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salama, N.R.; Hartung, M.L.; Müller, A. Life in the human stomach persistence strategies of the bacterial pathogen Helicobacter pylori. Nat. Rev. 2013, 11, 385–398. [Google Scholar] [CrossRef] [PubMed]
- Hathroubi, S.; Servetas, S.L.; Windham, I.; Merrell, D.S.; Ottemann, K.M. Helicobacter pylori Biofilm Formation and Its Potential Role in Pathogenesis. Microbiol. Mol. Biol. Rev. 2018, 82, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Hathroubi, S.; Zerebinski, J.; Ottemann, K.M. Helicobacter pylori Biofilm Involves a Multigene Stress-Biased Response, Including a Structural Role for Flagella. mBio 2018, 9, e01973-18. [Google Scholar] [CrossRef] [Green Version]
- Xia, M.; Chen, H.; Liu, S. The synergy of resveratrol and alcohol against Helicobacter pylori and underlying anti-Helicobacter pylori mechanism of resveratrol. J. Appl. Microbiol. 2020, 128, 1179–1190. [Google Scholar] [CrossRef]
- Hathroubi, S.; Hu, S.; Ottemann, K.M. Genetic requirements and transcriptomics of Helicobacter pylori biofilm formation on abiotic and biotic surfaces. NPJ Biofilms Microbioms 2020, 6, 56. [Google Scholar] [CrossRef]
- Zhao, Y.; Cai, Y.; Chen, Z.; Li, H.; Xu, Z.; Li, W.; Jia, J.; Sun, Y. SpoT-mediated NapA upregulation promotes oxidative stress-induced Helicobacter pylori biofilm formation and confers multidrug resistance. Antimicrob. Agents Chemother. 2021, 65, e00152-21. [Google Scholar] [CrossRef]
- Ye, Q.; Shao, X.; Shen, R.; Chen, D.; Shen, J. Changes in the human gut microbiota composition caused by Helicobacter pylori eradication therapy: A systematic review and meta-analysis. Helicobacter 2020, 25, e12713. [Google Scholar] [CrossRef]
- Suthisamphat, N.; Dechayont, B.; Phuaklee, P.; Prajuabjinda, O.; Vilaichone, R.K.; Itharat, A.; Mokmued, K.; Prommee, N. Anti-Helicobacter pylori, Anti-Inflammatory, Cytotoxic, and Antioxidant Activities of Mace Extracts from Myristica fragrans. Evid.-Based Complement Altern. Med. 2020, 2020, 7576818. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tran Trung, H.; Truong Thi Huynh, H.; Nguyen Thi Thuy, L.; Nguyen Van Minh, H.; Thi Nguyen, M.N.; Luong Thi, M.N. Growth-Inhibiting, Bactericidal, Antibiofilm, and Urease Inhibitory Activities of Hibiscus rosa sinensis L. Flower Constituents toward Antibiotic Sensitive- and Resistant-Strains of Helicobacter pylori. ACS Omega 2020, 5, 20080–20089. [Google Scholar] [CrossRef] [PubMed]
- Cardoso, O.; Donato, M.M.; Luxo, C.; Almeida, N.; Liberal, J.; Figueirinha, A.; Batista, M.T. Anti-Helicobacter pylori potential of Agrimonia eupatoria L. and Fragaria vesca. J. Funct. Foods 2018, 44, 299–303. [Google Scholar] [CrossRef]
- Prasad, A.; Devi, A.T.; Prasad, M.N.N.; Zameer, F.; Shruthi, G.; Shivamallu, C. Phyto anti-biofilm elicitors as potential inhibitors of Helicobacter pylori. 3 Biotech 2019, 9, 53. [Google Scholar] [CrossRef]
- Zhang, J.; Chen, Z.; Kong, J.; Liang, Y.; Chen, K.; Chang, Y.; Yuan, H.; Wang, Y.; Liang, H.; Li, J.; et al. Fullerenol Nanoparticles Eradicate Helicobacter pylori via pH-Responsive Peroxidase Activity. ACS Appl. Mater. Interfaces 2020, 12, 29013–29023. [Google Scholar] [CrossRef]
- Yang, S.J.; Huang, C.H.; Yang, J.C.; Wang, C.H.; Shieh, M.J. Residence Time-Extended Nanoparticles by Magnetic Field Improve the Eradication Efficiency of Helicobacter pylori. ACS Appl. Mater. Interfaces 2020, 12, 54316–54327. [Google Scholar] [CrossRef]
- Arif, M.; Sharaf, M.; Samreen; Khan, S.; Chi, Z.; Liu, C.G. Chitosan-based nanoparticles as delivery-carrier for promising antimicrobial glycolipid biosurfactant to improve the eradication rate of Helicobacter pylori biofilm. J. Biomater. Sci. Polym. Ed. 2021, 32, 813–832. [Google Scholar] [CrossRef]
- Qureshi, N.; Li, P.; Gu, Q. Probiotic therapy in Helicobacter pylori infection: A potential strategy against a serious pathogen? Appl. Microbiol. Biotechnol. 2019, 103, 1573–1588. [Google Scholar] [CrossRef]
- Ji, J.; Yang, H. In Vitro Effects of Lactobacillus plantarum LN66 and Antibiotics Used Alone or in Combination on Helicobacter pylori Mature Biofilm. Microorganisms 2021, 9, 424. [Google Scholar] [CrossRef]
- Graham, D.Y.; Fischbach, L. Helicobacter pylori treatment in the era of increasing antbiotic resistance. Gut 2010, 59, 1143–1153. [Google Scholar] [CrossRef]
- Wang, F.; Feng, J.R.; Chen, P.F.; Liu, X.P.; Ma, M.X.; Zhou, R.; Chang, Y.; Liu, J.; Li, J.; Zhao, Q. Probiotics in Helicobacter pylori eradication therapy: Systematic review and network meta-analysis. Clin. Res. Hepatol. Gastroenterol. 2017, 41, 466–475. [Google Scholar] [CrossRef] [PubMed]
- Lv, Z.; Wang, B.; Zhou, X.; Wang, F.; Xie, Y.; Zheng, H.; Lv, N. Efficacy and safety of probiotics as adjuvant agents for Helicobacter pylori infection: A meta-analysis. Exp. Ther. Med. 2015, 9, 707–716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, F.; Yang, H. Effects of Lactobacillus salivarius LN12 in Combination with Amoxicillin and Clarithromycin on Helicobacter pylori Biofilm In Vitro. Microorganisms 2021, 9, 1611. [Google Scholar] [CrossRef] [PubMed]
- Krzyzek, P.; Grande, R.; Migdal, P.; Paluch, E.; Gosciniak, G. Biofilm Formation as a Complex Result of Virulence and Adaptive Responses of Helicobacter pylori. Pathogens 2020, 9, 1062. [Google Scholar] [CrossRef] [PubMed]
- Franklin, M.J.; Chang, C.; Akiyama, T.; Bothner, B. New Technologies for Studying Biofilms. Microbiol. Spectr. 2015, 8, 3. [Google Scholar] [CrossRef] [Green Version]
- Ge, X.R.; Cai, Y.Y.; Chen, Z.H.; Gao, S.Z.; Geng, X.W.; Li, Y.; Li, Y.; Jia, J.H.; Sun, Y.D. Bifunctional Enzyme SpoT Is Involved in Biofilm Formation of Helicobacter pylori with Multidrug Resistance by Upregulating Efflux Pump Hp1174 (gluP). Antimicrob. Agents Chemother. 2018, 62, e00957-18. [Google Scholar] [CrossRef] [Green Version]
- Yu, M.; Wang, X.; Ling, F.; Wang, H.; Zhang, P.; Shao, S. Atractylodes lancea volatile oils attenuated Helicobacter pylori NCTC11637 growth and biofilm. Microb. Pathog. 2019, 135, 103641. [Google Scholar] [CrossRef]
- Li, P.Y.; Chen, X.N.; Shen, Y.N.; Li, H.T.; Zou, Y.Q.; Yuan, G.; Hu, P.; Hu, H.Y. Mucus penetration enhanced lipid polymer nanoparticles improve the eradication rate of Helicobacter pylori biofilm. J. Control. Release 2019, 300, 52–63. [Google Scholar] [CrossRef]
- Chen, X.N.; Li, P.Y.; Shen, Y.N.; Zou, Y.Q.; Yuan, G.; Hu, H.Y. Rhamnolipid-involved antibiotics combinations improve the eradication of Helicobacter pylori biofilm in vitro: A comparison with conventional triple therapy. Microb. Pathog. 2019, 131, 112–119. [Google Scholar] [CrossRef]
- Smith, M.A.; Finel, M.; Korolik, V.; Mendz, G.L. Characteristics of the aerobic respiratory chains of the microaerophiles Campylobacter jejuni and Helicobacter pylori. Arch. Microbiol. 2000, 174, 1–10. [Google Scholar] [CrossRef]
- Flemming, D.; Schlitt, A.; Spehr, V.; Bischof, T.; Friedrich, T. Iron-sulfur cluster N2 of the Escherichia coli NADH:ubiquinone oxidoreductase (complex I) is located on subunit NuoB. J. Biol. Chem. 2003, 278, 47602–47609. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, W.; Xiao, Y.; Long, Y.; Li, Y.; Peng, F.; Zhu, C.; He, T.; Lou, D.; Zhu, Y. Sodium fluoride causes oxidative damage to silkworm (Bombyx mori) testis by affecting the oxidative phosphorylation pathway. Ecotoxicol. Environ. Saf. 2021, 218, 112229. [Google Scholar] [CrossRef] [PubMed]
- Bai, H.; Zhou, D.G.; Zhang, X.W.; Cao, Y.F.; Xiao, X.L.; Zhang, Y.; Yu, Y.G. The responses of Salmonella enterica serovar Typhimurium to vanillin in apple juice through global transcriptomics. Int. J. Food Microbiol. 2021, 347, 109189. [Google Scholar] [CrossRef] [PubMed]
- Rader, B.A.; Wreden, C.; Hicks, K.G.; Sweeney, E.G.; Ottemann, K.M.; Guillemin, K. Helicobacter pylori perceives the quorum-sensing molecule Al-2 as a chemorepellent via the chemoreceptor TIpB. Microbiol.-Sgm 2011, 157, 2445–2455. [Google Scholar] [CrossRef] [Green Version]
- Anderson, J.; Huang, J.; Wreden, C.; Sweeney, E.; Goers, J.; Remington, S.J.; Guillemin, K. Chemorepulsion from the Quorum Signal Autoinducer-2 Promotes Helicobacter pylori Biofilm Dispersal. MBio 2015, 6, e00379-15. [Google Scholar] [CrossRef] [Green Version]
- Allard-Massicotte, R.; Tessier, L.; Lecuyer, F.; Lakshmanan, V.; Lucier, J.F.; Garneau, D.; Caudwell, L.; Vlamakis, H.; Bais, H.P.; Beauregard, P.B. Bacillus subtilis Early Colonization of Arabidopsis thaliana Roots Involves Multiple Chemotaxis Receptors. mBio 2016, 7, e01664-16. [Google Scholar] [CrossRef] [Green Version]
- Wistrand-Yuen, E.; Knopp, M.; Hjort, K.; Koskiniemi, S.; Berg, O.G.; Andersson, D.I. Evolution of high-level resistance during low-level antibiotic exposure. Nat. Commun. 2018, 9, 1599. [Google Scholar] [CrossRef] [Green Version]
- Jutkina, J.; Marathe, N.P.; Flach, C.F.; Larsson, D.G.J. Antibiotics and common antibacterial biocides stimulate horizontal transfer of resistance at low concentrations. Sci. Total. Environ. 2018, 616–617, 172–178. [Google Scholar] [CrossRef]
- Sato, M.; Sinha, P.K.; Torres-Bacete, J.; Matsuno-Yagi, A.; Yagi, T. Energy transducing roles of antiporter-like subunits in Escherichia coli NDH-1 with main focus on subunit NuoN (ND2). J. Biol. Chem. 2013, 288, 24705–24716. [Google Scholar] [CrossRef] [Green Version]
- Lane, M.C.; O’Toole, P.W.; Moore, S.A. Molecular basis of the interaction between the flagellar export proteins FliI and FliH from Helicobacter pylori. J. Biol. Chem. 2006, 281, 508–517. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.J.; Sheu, B.S.; Huang, A.H.; Lin, S.T.; Yang, H.B. Characterization of flgK gene and FlgK protein required for H. pylori colonization-from cloning to clinical relevance. World J. Gastroenterol. 2006, 12, 3989–3993. [Google Scholar] [CrossRef] [PubMed]
- Chu, J.K.; Zhu, S.; Herrera, C.M.; Henderson, J.C.; Liu, J.; Trent, M.S.; Hoover, T.R. Loss of a Cardiolipin Synthase in Helicobacter pylori G27 Blocks Flagellum Assembly. J. Bacteriol. 2019, 201, e00372-19. [Google Scholar] [CrossRef] [PubMed]
- Kim, N.; Marcus, E.A.; Wen, Y.; Weeks, D.L.; Scott, D.R.; Jung, H.C.; Song, I.S.; Sachs, G. Genes of Helicobacter pylori regulated by attachment to AGS cells. Infect. Immun. 2004, 72, 2358–2368. [Google Scholar] [CrossRef] [Green Version]
- O’Neill, J.; Roujeinikova, A. Cloning, purification and crystallization of MotB, a stator component of the proton-driven bacterial flagellar motor. Acta Crystallogr. Sect. F Struct. Biol. Cryst. Commun. 2008, 64, 561–563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, A.; Hodgson, N.; Yan, M.; Joo, J.; Gu, L.; Sang, H.; Gregory-Bryson, E.; Wood, W.G.; Ni, Y.; Smith, K.; et al. Screening Helicobacter pylori genes induced during infection of mouse stomachs. World J. Gastroenterol. 2012, 18, 4323–4334. [Google Scholar] [CrossRef]
- Homuth, G.; Domm, S.; Kleiner, D.; Schumann, W. Transcriptional analysis of major heat shock genes of Helicobacter pylori. J. Bacteriol. 2000, 182, 4257–4263. [Google Scholar] [CrossRef] [Green Version]
- Keikha, M. Is there a relationship between Helicobacter pylori vacA i1 or i2 alleles and development into peptic ulcer and gastric cancer? A meta-analysis study on an Iranian population. New Microbes New Infect. 2020, 36, 100726. [Google Scholar] [CrossRef]
- Yang, J.; Yang, H. Transcriptome Analysis of the Clostridioides difficile Response to Different Doses of Bifidobacterium breve. Front. Microbiol. 2020, 11, 1863. [Google Scholar] [CrossRef]
- Wang, N.; Hang, X.M.; Zhang, M.; Peng, X.Y.; Yang, H. New genetic environments of the macrolide-lincosamide-streptogramin resistance determinant erm(X) and their influence on potential horizontal transferability in Bifidobacteria. Int. J. Antimicrob. Agents 2017, 50, 572–580. [Google Scholar] [CrossRef]
- Bindea, G.; Mlecnik, B.; Hackl, H.; Charoentong, P.; Tosolini, M.; Kirilovsky, A.; Fridman, W.H.; Pages, F.; Trajanoski, Z.; Galon, J. ClueGO: A Cytoscape plug-in to decipher functionally grouped gene ontology and pathway annotation networks. Bioinformatics 2009, 25, 1091–1093. [Google Scholar] [CrossRef] [Green Version]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jin, F.; Yang, H. Transcriptome Analysis of the Response of Mature Helicobacter pylori Biofilm to Different Doses of Lactobacillus salivarius LN12 with Amoxicillin and Clarithromycin. Antibiotics 2022, 11, 262. https://doi.org/10.3390/antibiotics11020262
Jin F, Yang H. Transcriptome Analysis of the Response of Mature Helicobacter pylori Biofilm to Different Doses of Lactobacillus salivarius LN12 with Amoxicillin and Clarithromycin. Antibiotics. 2022; 11(2):262. https://doi.org/10.3390/antibiotics11020262
Chicago/Turabian StyleJin, Fang, and Hong Yang. 2022. "Transcriptome Analysis of the Response of Mature Helicobacter pylori Biofilm to Different Doses of Lactobacillus salivarius LN12 with Amoxicillin and Clarithromycin" Antibiotics 11, no. 2: 262. https://doi.org/10.3390/antibiotics11020262
APA StyleJin, F., & Yang, H. (2022). Transcriptome Analysis of the Response of Mature Helicobacter pylori Biofilm to Different Doses of Lactobacillus salivarius LN12 with Amoxicillin and Clarithromycin. Antibiotics, 11(2), 262. https://doi.org/10.3390/antibiotics11020262