
Antimicrobial Resistance, Biofilm Formation, and Virulence Genes in Enterococcus Species from Small Backyard Chicken Flocks


 , ,
, ,  , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. Bacterial Isolation and Identification
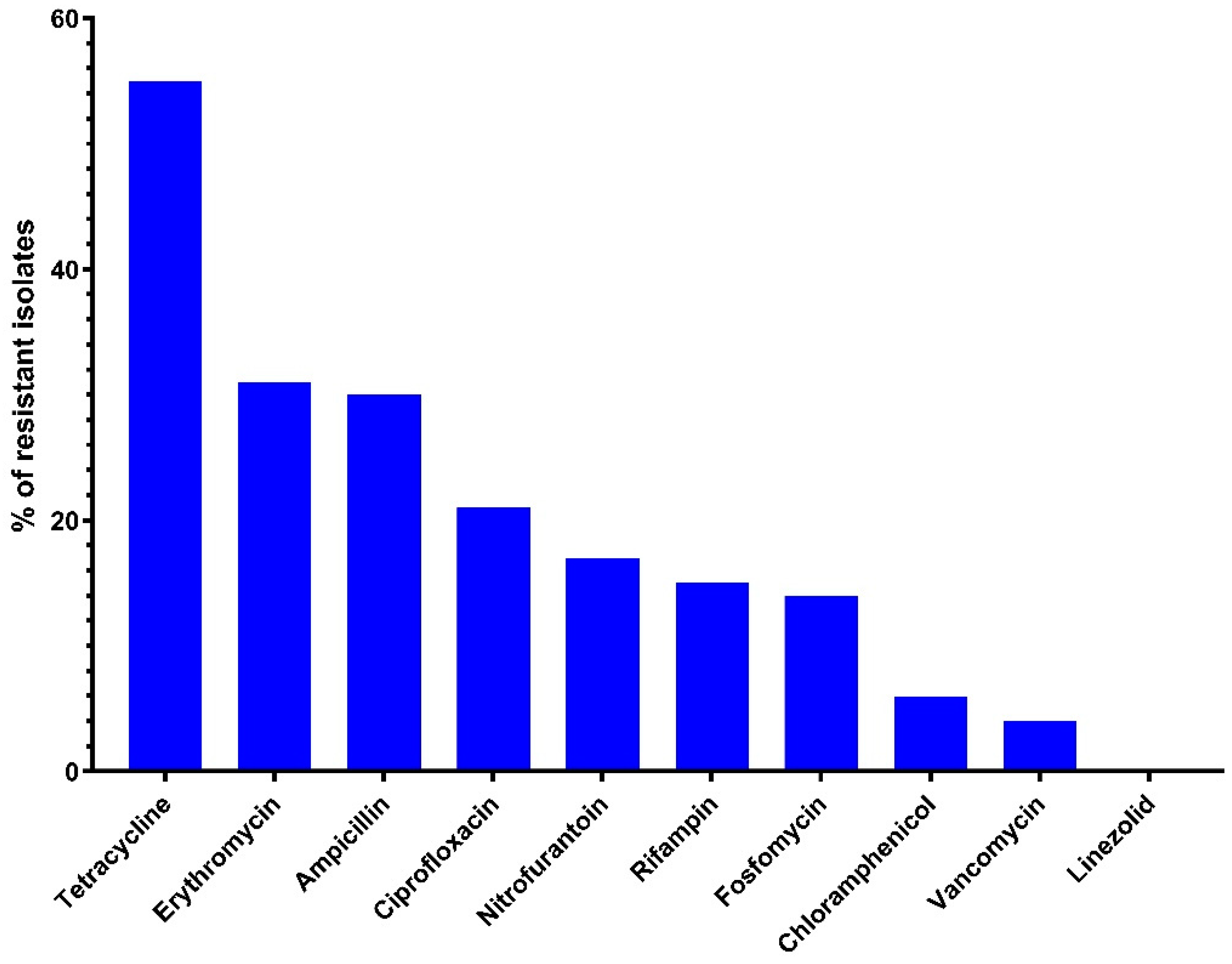
2.2. Antimicrobial Sensitivity
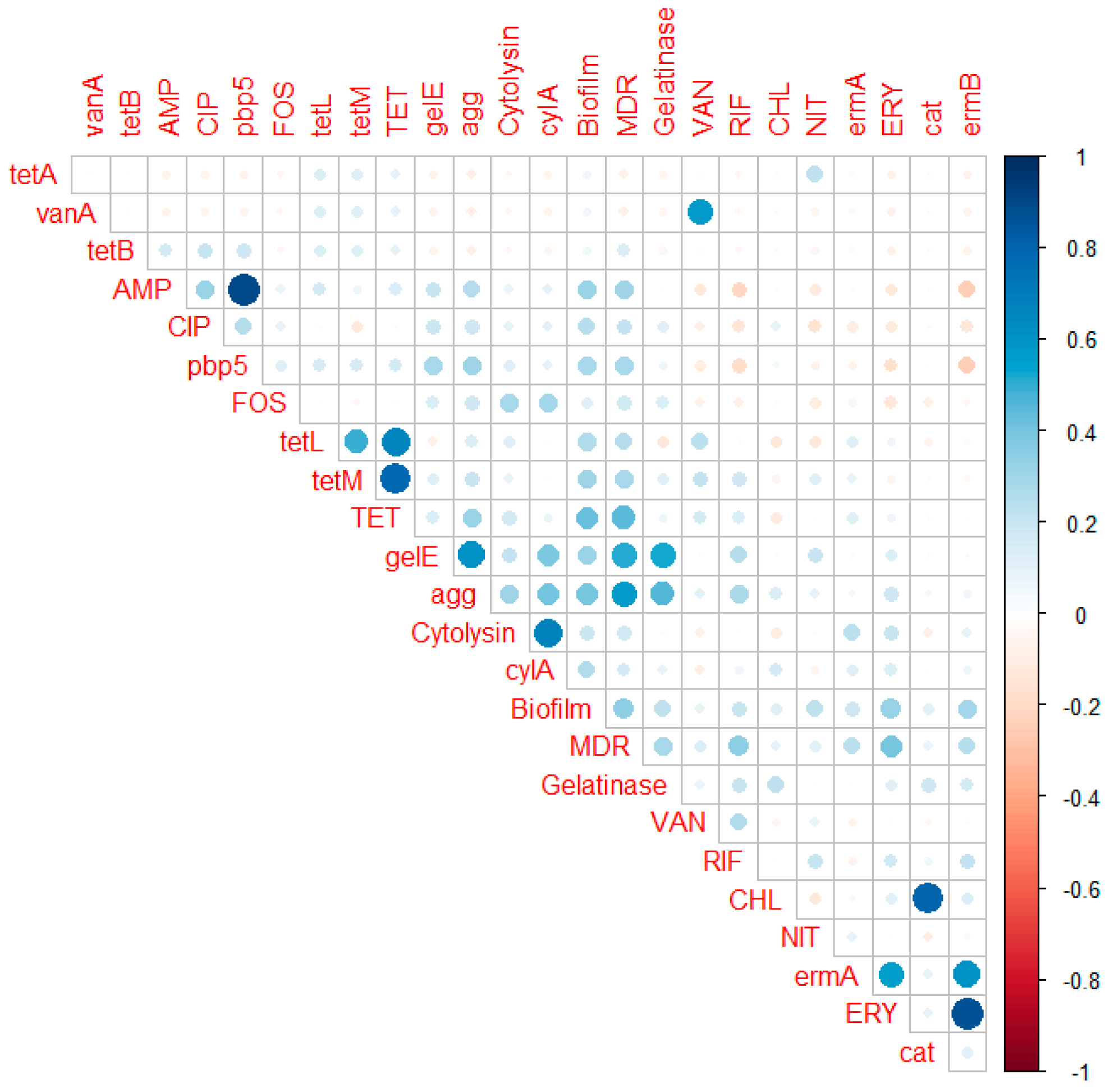
2.3. Antimicrobial-Resistance Genes
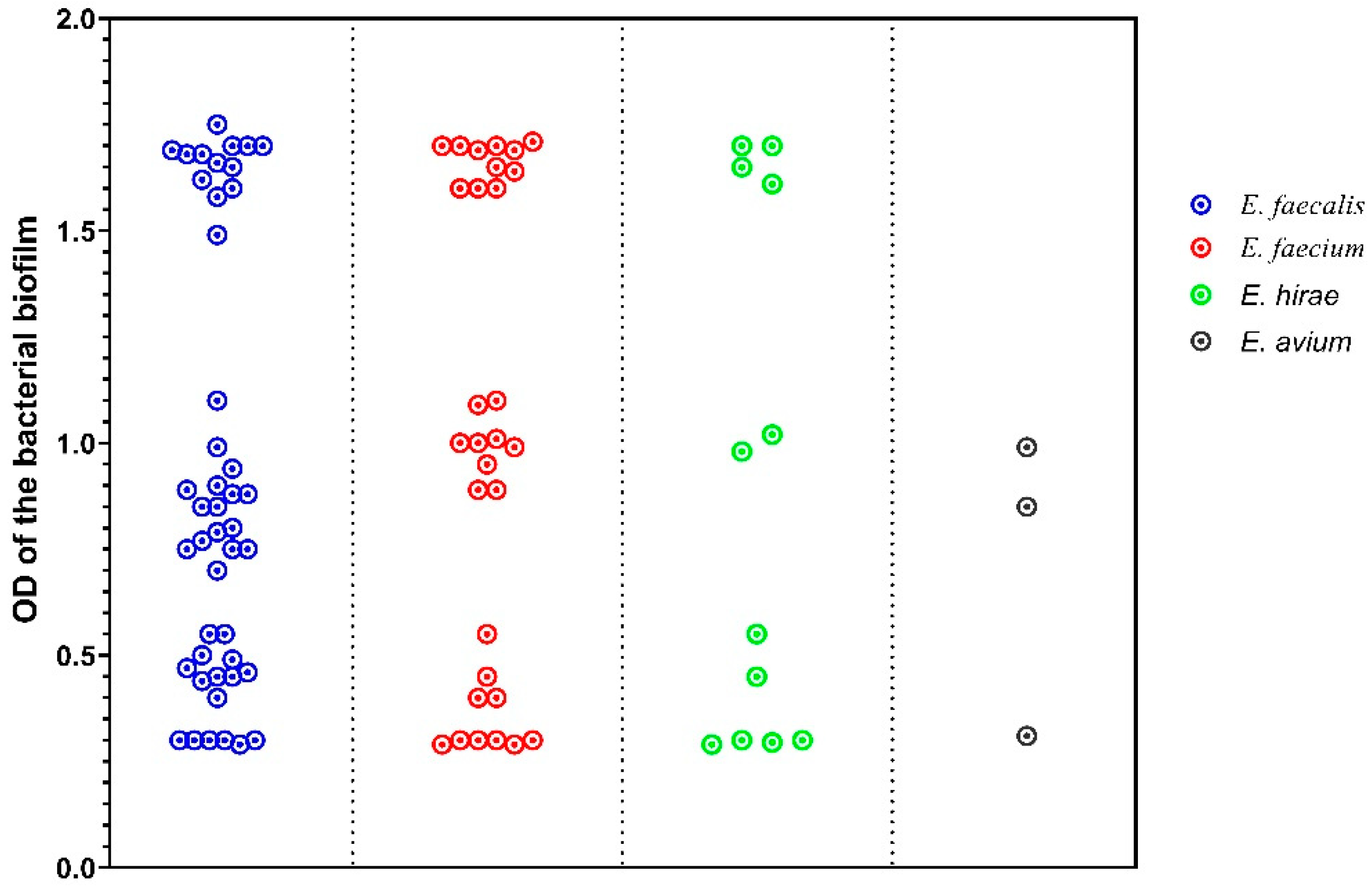
2.4. Biofilm Formation
2.5. Gelatinase and Cytolysin Activity
2.6. Virulence Genes
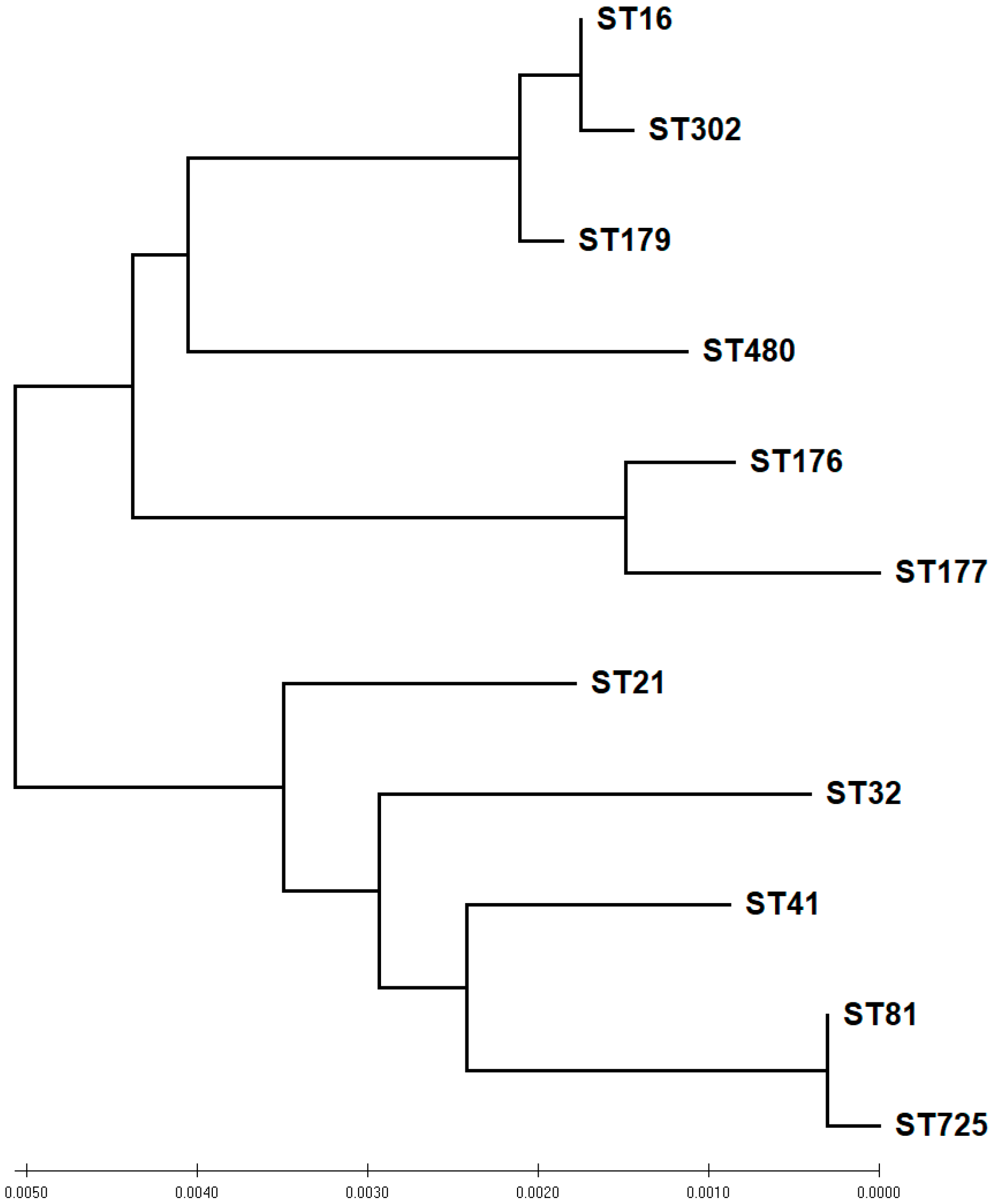
2.7. MLST of E. faecalis and E. faecium
2.8. repA Genes (Plasmid Families)
3. Discussion
4. Materials and Methods
4.1. Study Area
4.2. Bacterial Isolation
4.3. DNA Extraction and 16S rRNA Gene Amplification and Sequencing
4.4. Antimicrobial Sensitivity Test
4.5. Detection of Antimicrobial-Resistance Genes
4.6. Phenotypic Detection of Virulence Factors
4.6.1. Quantitative Biofilm Assay
4.6.2. Gelatinase Activity
4.6.3. Cytolysin Activity
4.7. Molecular Detection of Virulence Factor Genes
4.8. Multi-Locus Sequence Typing
4.9. PCR for repA Genes (Plasmid Families)
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fisher, K.; Phillips, C. The ecology, epidemiology and virulence of Enterococcus. Microbiology 2009, 155, 1749–1757. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Byappanahalli, M.N.; Nevers, M.B.; Korajkic, A.; Staley, Z.R.; Harwood, V.J. Enterococci in the environment. Microbiol. Mol. Biol. Rev. 2012, 76, 685–706. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- García-Solache, M.; Rice, L.B. The Enterococcus: A model of adaptability to its environment. Clin. Microbiol. Rev. 2019, 32, e00058-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sava, I.G.; Heikens, E.; Huebner, J. Pathogenesis and immunity in enterococcal infections. Clin. Microbiol. Infect. 2010, 16, 533–540. [Google Scholar] [CrossRef] [PubMed]
- Ammerlaan, H.S.; Harbarth, S.; Buiting, A.G.; Crook, D.W.; Fitzpatrick, F.; Hanberger, H.; Herwaldt, L.A.; van Keulen, P.H.; Kluytmans, J.A.; Kola, A.; et al. Secular trends in nosocomial bloodstream infections: Antibiotic-resistant bacteria increase the total burden of infection. Clin. Infect. Dis. 2013, 56, 798–805. [Google Scholar] [CrossRef] [PubMed]
- Cheng, W.N.; Han, S.G. Bovine mastitis: Risk factors, therapeutic strategies, and alternative treatments—A review. Asian-Australas. J. Anim. Sci. 2020, 33, 1699–1713. [Google Scholar] [CrossRef] [PubMed]
- Wood, M.W.; Lepold, A.; Tesfamichael, D.; Lasarev, M.R. Risk factors for enterococcal bacteriuria in dogs: A retrospective study. J. Vet. Intern. Med. 2020, 34, 2447–2453. [Google Scholar] [CrossRef] [PubMed]
- Robbins, K.M.; Suyemoto, M.M.; Lyman, R.L.; Martin, M.P.; Barnes, H.J.; Borst, L.B. An outbreak and source investigation of enterococcal spondylitis in broilers caused by Enterococcus cecorum. Avian Dis. 2012, 56, 768–773. [Google Scholar] [CrossRef] [PubMed]
- Seputiene, V.; Bogdaite, A.; Ruzauskas, M.; Suziedeliene, E. Antibiotic resistance genes and virulence factors in Enterococcus faecium and Enterococcus faecalis from diseased farm animals: Pigs, cattle and poultry. Pol. J. Vet. Sci. 2012, 15, 431–438. [Google Scholar]
- Katakweba, A.; Mtambo, M.; Olsen, J.E.; Muhairwa, A.P. Awareness of human health risks associated with the use of antibiotics among livestock keepers and factors that contribute to selection of antibiotic resistance bacteria within livestock in Tanzania. Livest. Res. Rural. Dev. 2012, 24, 170. [Google Scholar]
- Osman, K.M.; Badr, J.; Orabi, A.; Elbehiry, A.; Saad, A.; Ibrahim, M.D.; Hanafy, M.H. Poultry as a vector for emerging multidrug resistant Enterococcus spp.: First report of vancomycin (van) and the chloramphenicol–florfenicol (cat-fex-cfr) resistance genes from pigeon and duck faeces. Microb. Pathog. 2019, 128, 195–205. [Google Scholar] [CrossRef] [PubMed]
- Hollenbeck, B.L.; Rice, L.B. Intrinsic and acquired resistance mechanisms in enterococcus. Virulence 2012, 3, 421–433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asgin, N.; Otlu, B. Antibiotic Resistance and Molecular Epidemiology of Vancomycin-Resistant Enterococci in a Tertiary Care Hospital in Turkey. Infect. Drug Resist. 2020, 13, 191–198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Werner, G.; Coque, T.M.; Franz, C.M.; Grohmann, E.; Hegstad, K.; Jensen, L.; van Schaik, W.; Weaver, K. Antibiotic resistant enterococci—Tales of a drug resistance gene trafficker. Int. J. Med. Microbiol. 2013, 303, 360–379. [Google Scholar] [CrossRef] [PubMed]
- Anderson, A.C.; Jonas, D.; Huber, I.; Karygianni, L.; Wolber, J.; Hellwig, E.; Arweiler, N.; Vach, K.; Wittmer, A.; Al-Ahmad, A. Enterococcus faecalis from Food, Clinical Specimens, and Oral Sites: Prevalence of Virulence Factors in Association with Biofilm Formation. Front. Microbiol. 2015, 6, 1534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sandoe, J.A.; Witherden, I.R.; Cove, J.H.; Heritage, J.; Wilcox, M.H. Correlation between enterococcal biofilm formation in vitro and medical-device-related infection potential in vivo. J. Med. Microbiol. 2003, 52, 547–550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kayaoglu, G.; Orstavik, D. Virulence factors of Enterococcus faecalis: Relationship to endodontic disease. Crit. Rev. Oral Biol. Med. 2004, 15, 308–320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pryce, T.M.; Wilson, R.D.; Kulski, J.K. Identification of enterococci by ribotyping with horseradish-peroxidase-labelled 16S rDNA probes. J. Microbiol. Methods 1999, 36, 147–155. [Google Scholar] [CrossRef]
- Malathum, K.; Singh, K.V.; Weinstock, G.M.; Murray, B.E. Repetitive sequence-based PCR versus pulsed-field gel electrophoresis for typing of Enterococcus faecalis at the subspecies level. J. Clin. Microbiol. 1998, 36, 211–215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seetulsingh, P.S.; Tomayko, J.F.; Coudron, P.E.; Markowitz, S.M.; Skinner, C.; Singh, K.V.; Murray, B.E. Chromosomal DNA restriction endonuclease digestion patterns of beta-lactamase-producing Enterococcus faecalis isolates collected from a single hospital over a 7-year period. J. Clin. Microbiol. 1996, 34, 1892–1896. [Google Scholar] [CrossRef] [Green Version]
- Tomayko, J.F.; Murray, B.E. Analysis of Enterococcus faecalis isolates from intercontinental sources by multilocus enzyme electrophoresis and pulsed-field gel electrophoresis. J. Clin. Microbiol. 1995, 33, 2903–2907. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruiz-Garbajosa, P.; Bonten, M.J.; Robinson, D.A.; Top, J.; Nallapareddy, S.R.; Torres, C.; Coque, T.M.; Cantón, R.; Baquero, F.; Murray, B.E.; et al. Multilocus sequence typing scheme for Enterococcus faecalis reveals hospital-adapted genetic complexes in a background of high rates of recombination. J. Clin. Microbiol. 2006, 44, 2220–2228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Homan, W.L.; Tribe, D.; Poznanski, S.; Li, M.; Hogg, G.; Spalburg, E.; Van Embden, J.D.; Willems, R.J. Multilocus sequence typing scheme for Enterococcus faecium. J. Clin. Microbiol. 2002, 40, 1963–1971. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Achtman, M. Evolution, population structure, and phylogeography of genetically monomorphic bacterial pathogens. Annu. Rev. Microbiol. 2008, 62, 53–70. [Google Scholar] [CrossRef] [PubMed]
- Maâtallah, M.; Bakhrouf, A.; Habeeb, M.A.; Turlej-Rogacka, A.; Iversen, A.; Pourcel, C.; Sioud, O.; Giske, C.G. Four Genotyping Schemes for Phylogenetic Analysis of Pseudomonas aeruginosa: Comparison of Their Congruence with Multi-Locus Sequence Typing. PLoS ONE 2013, 8, e82069. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aslam, M.; Diarra, M.S.; Checkley, S.; Bohaychuk, V.; Masson, L. Characterization of antimicrobial resistance and virulence genes in Enterococcus spp. isolated from retail meats in Alberta, Canada. Int. J. Food Microbiol. 2012, 156, 222–230. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.J.; Koo, M. Occurrence, Antimicrobial Resistance and Molecular Diversity of Enterococcus faecium in Processed Pork Meat Products in Korea. Foods 2020, 9, 1283. [Google Scholar] [CrossRef] [PubMed]
- Holman, D.B.; Klima, C.L.; Gzyl, K.E.; Zaheer, R.; Service, C.; Jones, T.H.; McAllister, T.A. Antimicrobial Resistance in Enterococcus spp. Isolated from a Beef Processing Plant and Retail Ground Beef. Microbiol. Spectr. 2021, 9, e0198021. [Google Scholar] [CrossRef] [PubMed]
- Moreno, M.F.; Sarantinopoulos, P.; Tsakalidou, E.; De Vuyst, L. The role and application of enterococci in food and health. Int. J. Food Microbiol. 2006, 106, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Hegstad, K.; Mikalsen, T.; Coque, T.M.; Werner, G.; Sundsfjord, A. Mobile genetic elements and their contribution to the emergence of antimicrobial resistant Enterococcus faecalis and Enterococcus faecium. Clin. Microbiol. Infect. 2010, 16, 541–554. [Google Scholar] [CrossRef]
- Grohmann, E.; Muth, G.; Espinosa, M. Conjugative plasmid transfer in gram-positive bacteria. Microbiol. Mol. Biol. Rev. MMBR 2003, 67, 277–301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hammerum, A.M.; Flannagan, S.E.; Clewell, D.B.; Jensen, L.B. Indication of transposition of a mobile DNA element containing the vat(D) and erm(B) genes in Enterococcus faecium. Antimicrob. Agents Chemother. 2001, 45, 3223–3225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Osborn, M.; Bron, S.; Firth, N.; Holsappel, S.; Huddleston, A.; Kiewiet, R.; Meijer, W.; Seegers, J.; Skurray, R.; Terpstra, P.J.; et al. The evolution of bacterial plasmids. In The Horizontal Gene Pool: Bacterial Plasmids and Gene Spread; CRC Press: Boca Raton, FL, USA, 2000; pp. 301–363. [Google Scholar]
- Coque, T.M. Evolutionary biology of pathogenic enterococci. In Evolutionary Biology of Bacterial and Fungal Pathogens; ASM Press: Washington, DC, USA, 2007; pp. 501–521. [Google Scholar]
- Gilmore, M.S.; Clewell, D.B.; Courvalin, P.; Dunny, G.M.; Murray, B.E.; Rice, L.B. The Enterococci: Pathogenesis, Molecular Biology, and Antibiotic Resistance; ASM Press: Washington, DC, USA, 2002; Volume 10. [Google Scholar]
- Khan, S.A. Rolling-circle replication of bacterial plasmids. Microbiol. Mol. Biol. Rev. 1997, 61, 442–455. [Google Scholar] [CrossRef] [PubMed]
- Jensen, L.B.; Garcia-Migura, L.; Valenzuela, A.J.; Løhr, M.; Hasman, H.; Aarestrup, F.M. A classification system for plasmids from enterococci and other Gram-positive bacteria. J. Microbiol. Methods 2010, 80, 25–43. [Google Scholar] [CrossRef] [PubMed]
- Kwon, K.H.; Hwang, S.Y.; Moon, B.Y.; Park, Y.K.; Shin, S.; Hwang, C.Y.; Park, Y.H. Occurrence of antimicrobial resistance and virulence genes, and distribution of enterococcal clonal complex 17 from animals and human beings in Korea. J. Vet. Diagn. Investig. 2012, 24, 924–931. [Google Scholar] [CrossRef] [PubMed]
- Salem-Bekhit, M.M.; Moussa, I.M.; Muharram, M.M.; Alanazy, F.K.; Hefni, H.M. Prevalence and antimicrobial resistance pattern of multidrug-resistant enterococci isolated from clinical specimens. Indian J. Med. Microbiol. 2012, 30, 44–51. [Google Scholar] [CrossRef] [PubMed]
- Abdallah, M.; Al-Saafin, M. Overview of Prevalence, Characteristics, Risk Factors, Resistance, and Virulence of Vancomycin-Resistant Enterococci in Saudi Arabia. Microb. Drug Resist. 2019, 25, 350–358. [Google Scholar] [CrossRef] [PubMed]
- Qadri, S.H.; Qunibi, W.Y.; Al-Ballaa, S.R.; Kadhi, Y.; Burdette, J.M. Vancomycin resistant enterococcus: A case report and review of the literature. Ann. Saudi Med. 1993, 13, 289–293. [Google Scholar] [CrossRef] [PubMed]
- Pohjola, L.; Nykasenoja, S.; Kivisto, R.; Soveri, T.; Huovilainen, A.; Hanninen, M.L.; Fredriksson-Ahomaa, M. Zoonotic Public Health Hazards in Backyard Chickens. Zoonoses Public Health 2016, 63, 420–430. [Google Scholar] [CrossRef] [PubMed]
- Semedo-Lemsaddek, T.; Bettencourt Cota, J.; Ribeiro, T.; Pimentel, A.; Tavares, L.; Bernando, F.; Oliveira, M. Resistance and virulence distribution in enterococci isolated from broilers reared in two farming systems. Ir. Vet. J. 2021, 74, 22. [Google Scholar] [CrossRef] [PubMed]
- Al-Marri, T.; Al-Marri, A.; Al-Zanbaqi, R.; Ajmi, A.A.; Fayez, M. Multidrug resistance, biofilm formation, and virulence genes of Escherichia coli from backyard poultry farms. Vet. World 2021, 14, 2869–2877. [Google Scholar] [CrossRef] [PubMed]
- Guzman Prieto, A.M.; van Schaik, W.; Rogers, M.R.; Coque, T.M.; Baquero, F.; Corander, J.; Willems, R.J. Global emergence and dissemination of enterococci as nosocomial pathogens: Attack of the clones? Front. Microbiol. 2016, 7, 788. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aarestrup, F.M.; Agerso, Y.; Gerner-Smidt, P.; Madsen, M.; Jensen, L.B. Comparison of antimicrobial resistance phenotypes and resistance genes in Enterococcus faecalis and Enterococcus faecium from humans in the community, broilers, and pigs in Denmark. Diagn. Microbiol. Infect. Dis. 2000, 37, 127–137. [Google Scholar] [CrossRef]
- Kaukas, A.; Hinton, M.; Linton, A.H. Changes in the faecal enterococcal population of young chickens and its effect on the incidence of resistance to certain antibiotics. Lett. Appl. Microbiol. 1986, 2, 5–8. [Google Scholar] [CrossRef]
- Diarra, M.S.; Rempel, H.; Champagne, J.; Masson, L.; Pritchard, J.; Topp, E. Distribution of antimicrobial resistance and virulence genes in Enterococcus spp. and characterization of isolates from broiler chickens. Appl. Environ. Microbiol. 2010, 76, 8033–8043. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshimura, H.; Ishimaru, M.; Endoh, Y.S.; Kojima, A. Antimicrobial susceptibilities of enterococci isolated from faeces of broiler and layer chickens. Lett. Appl. Microbiol. 2000, 31, 427–432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tyson, G.H.; Nyirabahizi, E.; Crarey, E.; Kabera, C.; Lam, C.; Rice-Trujillo, C.; McDermott, P.F.; Tate, H. Prevalence and Antimicrobial Resistance of Enterococci Isolated from Retail Meats in the United States, 2002 to 2014. Appl. Environ. Microbiol. 2018, 84, e01902-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boulianne, M.; Arsenault, J.; Daignault, D.; Archambault, M.; Letellier, A.; Dutil, L. Drug use and antimicrobial resistance among Escherichia coli and Enterococcus spp. isolates from chicken and turkey flocks slaughtered in Quebec, Canada. Can. J. Vet. Res. 2016, 80, 49–59. [Google Scholar] [PubMed]
- Obeng, A.S.; Rickard, H.; Ndi, O.; Sexton, M.; Barton, M. Comparison of antimicrobial resistance patterns in enterococci from intensive and free range chickens in Australia. Avian Pathol. 2013, 42, 45–54. [Google Scholar] [CrossRef] [PubMed]
- Persoons, D.; Dewulf, J.; Smet, A.; Herman, L.; Heyndrickx, M.; Martel, A.; Catry, B.; Butaye, P.; Haesebrouck, F. Prevalence and persistence of antimicrobial resistance in broiler indicator bacteria. Microb. Drug Resist. 2010, 16, 67–74. [Google Scholar] [CrossRef]
- Sadowy, E.; Luczkiewicz, A. Drug-resistant and hospital-associated Enterococcus faecium from wastewater, riverine estuary and anthropogenically impacted marine catchment basin. BMC Microbiol. 2014, 14, 66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jackson, C.R.; Fedorka-Cray, P.J.; Davis, J.A.; Barrett, J.B.; Brousse, J.H.; Gustafson, J.; Kucher, M. Mechanisms of antimicrobial resistance and genetic relatedness among enterococci isolated from dogs and cats in the United States. J. Appl. Microbiol. 2010, 108, 2171–2179. [Google Scholar] [CrossRef] [PubMed]
- Huys, G.; D’Haene, K.; Collard, J.M.; Swings, J. Prevalence and molecular characterization of tetracycline resistance in Enterococcus isolates from food. Appl. Environ. Microbiol. 2004, 70, 1555–1562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cauwerts, K.; Decostere, A.; De Graef, E.M.; Haesebrouck, F.; Pasmans, F. High prevalence of tetracycline resistance in Enterococcus isolates from broilers carrying the erm(B) gene. Avian Pathol. 2007, 36, 395–399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zaheer, R.; Cook, S.R.; Barbieri, R.; Goji, N.; Cameron, A.; Petkau, A.; Polo, R.O.; Tymensen, L.; Stamm, C.; Song, J.; et al. Surveillance of Enterococcus spp. reveals distinct species and antimicrobial resistance diversity across a One-Health continuum. Sci. Rep. 2020, 10, 3937. [Google Scholar] [CrossRef] [PubMed]
- Mlynarczyk, B.; Mlynarczyk, A.; Kmera-Muszynska, M.; Majewski, S.; Mlynarczyk, G. Mechanisms of resistance to antimicrobial drugs in pathogenic Gram-positive cocci. Mini Rev. Med. Chem. 2010, 10, 928–937. [Google Scholar] [CrossRef] [PubMed]
- Frei, A.; Goldenberger, D.; Teuber, M. Antimicrobial susceptibility of intestinal bacteria from Swiss poultry flocks before the ban of antimicrobial growth promoters. Syst. Appl. Microbiol. 2001, 24, 116–121. [Google Scholar] [CrossRef] [PubMed]
- van den Bogaard, A.E.; Stobberingh, E.E. Epidemiology of resistance to antibiotics: Links between animals and humans. Int. J. Antimicrob. Agents 2000, 14, 327–335. [Google Scholar] [CrossRef]
- Petsaris, O.; Miszczak, F.; Gicquel-Bruneau, M.; Perrin-Guyomard, A.; Humbert, F.; Sanders, P.; Leclercq, R. Combined antimicrobial resistance in Enterococcus faecium isolated from chickens. Appl. Environ. Microbiol. 2005, 71, 2796–2799. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Novais, C.; Coque, T.M.; Costa, M.J.; Sousa, J.C.; Baquero, F.; Peixe, L.V. High occurrence and persistence of antibiotic-resistant enterococci in poultry food samples in Portugal. J. Antimicrob. Chemother. 2005, 56, 1139–1143. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.B.; Seo, K.W.; Jeon, H.Y.; Lim, S.K.; Sung, H.W.; Lee, Y.J. Molecular characterization of erythromycin and tetracycline-resistant Enterococcus faecalis isolated from retail chicken meats. Poult. Sci. 2019, 98, 977–983. [Google Scholar] [CrossRef] [PubMed]
- Al-Mustafa, Z.H.; Al-Ghamdi, M.S. Use of antibiotics in the poultry industry in Saudi Arabia: Implications for public health. Ann. Saudi Med. 2002, 22, 4–7. [Google Scholar] [CrossRef] [PubMed]
- Collignon, P.; Conly, J.; Andremont, A. Members of the World Health Organization Advisory Group, Bogota meeting on Integrated Surveillance of Antimicrobial Resistance (WHO-AGISAR). Clin. Infect. Dis. 2016, 63, 1087–1093. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmed, M.O.; Baptiste, K.E. Vancomycin-Resistant Enterococci: A Review of Antimicrobial Resistance Mechanisms and Perspectives of Human and Animal Health. Microb. Drug Resist. 2018, 24, 590–606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jung, W.K.; Lim, J.Y.; Kwon, N.H.; Kim, J.M.; Hong, S.K.; Koo, H.C.; Kim, S.H.; Park, Y.H. Vancomycin-resistant enterococci from animal sources in Korea. Int. J. Food Microbiol. 2007, 113, 102–107. [Google Scholar] [CrossRef] [PubMed]
- Lim, S.-K.; Kim, T.-S.; Lee, H.-S.; Nam, H.-M.; Joo, Y.-S.; Koh, H.-B. Persistence of van A-type Enterococcus faecium in Korean livestock after ban on avoparcin. Microb. Drug Resist. 2006, 12, 136–139. [Google Scholar] [CrossRef] [PubMed]
- da Costa, P.M.; Oliveira, M.; Bica, A.; Vaz-Pires, P.; Bernardo, F. Antimicrobial resistance in Enterococcus spp. and Escherichia coli isolated from poultry feed and feed ingredients. Vet. Microbiol. 2007, 120, 122–131. [Google Scholar] [CrossRef] [PubMed]
- Cavaco, L.M.; Korsgaard, H.; Kaas, R.S.; Seyfarth, A.M.; Leekitcharoenphon, P.; Hendriksen, R.S. First detection of linezolid resistance due to the optrA gene in enterococci isolated from food products in Denmark. J. Glob. Antimicrob. Resist. 2017, 9, 128–129. [Google Scholar] [CrossRef] [PubMed]
- Yoon, S.; Kim, Y.B.; Seo, K.W.; Ha, J.S.; Noh, E.B.; Lee, Y.J. Characteristics of linezolid-resistant Enterococcus faecalis isolates from broiler breeder farms. Poult. Sci. 2020, 99, 6055–6061. [Google Scholar] [CrossRef] [PubMed]
- Castano-Arriba, A.; Gonzalez-Machado, C.; Igrejas, G.; Poeta, P.; Alonso-Calleja, C.; Capita, R. Antibiotic Resistance and Biofilm-Forming Ability in Enterococcal Isolates from Red Meat and Poultry Preparations. Pathogens 2020, 9, 1021. [Google Scholar] [CrossRef] [PubMed]
- Bekele, B.; Ashenafi, M. Distribution of drug resistance among enterococci and Salmonella from poultry and cattle in Ethiopia. Trop. Anim. Health Prod. 2010, 42, 857–864. [Google Scholar] [CrossRef] [PubMed]
- El-Zamkan, M.A.; Mohamed, H.M.A. Antimicrobial resistance, virulence genes and biofilm formation in Enterococcus species isolated from milk of sheep and goat with subclinical mastitis. PLoS ONE 2021, 16, e0259584. [Google Scholar] [CrossRef] [PubMed]
- Oliva, A.; Stefani, S.; Venditti, M.; Di Domenico, E.G. Biofilm-Related Infections in Gram-Positive Bacteria and the Potential Role of the Long-Acting Agent Dalbavancin. Front. Microbiol. 2021, 12, 749685. [Google Scholar] [CrossRef] [PubMed]
- Chai, W.L.; Hamimah, H.; Cheng, S.C.; Sallam, A.A.; Abdullah, M. Susceptibility of Enterococcus faecalis biofilm to antibiotics and calcium hydroxide. J. Oral Sci. 2007, 49, 161–166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hancock, L.E.; Perego, M. The Enterococcus faecalis fsr two-component system controls biofilm development through production of gelatinase. J. Bacteriol. 2004, 186, 5629–5639. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hashem, Y.A.; Amin, H.M.; Essam, T.M.; Yassin, A.S.; Aziz, R.K. Biofilm formation in enterococci: Genotype-phenotype correlations and inhibition by vancomycin. Sci. Rep. 2017, 7, 5733. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roberts, J.C.; Singh, K.V.; Okhuysen, P.C.; Murray, B.E. Molecular epidemiology of the fsr locus and of gelatinase production among different subsets of Enterococcus faecalis isolates. J. Clin. Microbiol. 2004, 42, 2317–2320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohamed, J.A.; Murray, B.E. Lack of correlation of gelatinase production and biofilm formation in a large collection of Enterococcus faecalis isolates. J. Clin. Microbiol. 2005, 43, 5405–5407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cox, C.R.; Coburn, P.S.; Gilmore, M.S. Enterococcal cytolysin: A novel two component peptide system that serves as a bacterial defense against eukaryotic and prokaryotic cells. Curr. Protein Pept. Sci. 2005, 6, 77–84. [Google Scholar] [CrossRef] [PubMed]
- Gholizadeh, P.; Aghazadeh, M.; Ghotaslou, R.; Ahangarzadeh Rezaee, M.; Pirzadeh, T.; Kose, S.; Ganbarov, K.; Yousefi, M.; Kafil, H.S. CRISPR-cas system in the acquisition of virulence genes in dental-root canal and hospital-acquired isolates of Enterococcus faecalis. Virulence 2020, 11, 1257–1267. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Sundsfjord, A.; Song, X. Enterococcus faecalis from patients with chronic periodontitis: Virulence and antimicrobial resistance traits and determinants. Eur. J. Clin. Microbiol. Infect. Dis. 2012, 31, 267–272. [Google Scholar] [CrossRef] [PubMed]
- Sedgley, C.M.; Molander, A.; Flannagan, S.E.; Nagel, A.C.; Appelbe, O.K.; Clewell, D.B.; Dahlen, G. Virulence, phenotype and genotype characteristics of endodontic Enterococcus spp. Oral Microbiol. Immunol. 2005, 20, 10–19. [Google Scholar] [CrossRef] [PubMed]
- Finlay, B.B.; Falkow, S. Common themes in microbial pathogenicity revisited. Microbiol. Mol. Biol. Rev. MMBR 1997, 61, 136–169. [Google Scholar] [CrossRef] [PubMed]
- Stępień-Pyśniak, D.; Hauschild, T.; Nowaczek, A.; Marek, A.; Dec, M. Wild birds as a potential source of known and novel multilocus sequence types of antibiotic-resistant Enterococcus faecalis. J. Wildl. Dis. 2018, 54, 219–228. [Google Scholar] [CrossRef] [PubMed]
- Gregersen, R.H.; Petersen, A.; Christensen, H.; Bisgaard, M. Multilocus sequence typing of Enterococcus faecalis isolates demonstrating different lesion types in broiler breeders. Avian Pathol. 2010, 39, 435–440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farman, M.; Yasir, M.; Al-Hindi, R.R.; Farraj, S.A.; Jiman-Fatani, A.A.; Alawi, M.; Azhar, E.I. Genomic analysis of multidrug-resistant clinical Enterococcus faecalis isolates for antimicrobial resistance genes and virulence factors from the western region of Saudi Arabia. Antimicrob. Resist. Infect. Control 2019, 8, 55. [Google Scholar] [CrossRef] [PubMed]
- Freitas, A.R.; Novais, C.; Ruiz-Garbajosa, P.; Coque, T.M.; Peixe, L. Clonal expansion within clonal complex 2 and spread of vancomycin-resistant plasmids among different genetic lineages of Enterococcus faecalis from Portugal. J. Antimicrob. Chemother. 2009, 63, 1104–1111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Leener, E.; Martel, A.; De Graef, E.M.; Top, J.; Butaye, P.; Haesebrouck, F.; Willems, R.; Decostere, A. Molecular analysis of human, porcine, and poultry Enterococcus faecium isolates and their erm(B) genes. Appl. Environ. Microbiol. 2005, 71, 2766–2770. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wist, V.; Morach, M.; Schneeberger, M.; Cernela, N.; Stevens, M.J.A.; Zurfluh, K.; Stephan, R.; Nüesch-Inderbinen, M. Phenotypic and Genotypic Traits of Vancomycin-Resistant Enterococci from Healthy Food-Producing Animals. Microorganisms 2020, 8, 261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torres, C.; Alonso, C.A.; Ruiz-Ripa, L.; León-Sampedro, R.; Del Campo, R.; Coque, T.M. Antimicrobial Resistance in Enterococcus spp. of animal origin. Microbiol. Spectr. 2018, 6, 24. [Google Scholar] [CrossRef] [PubMed]
- Lopez, M.; Hormazabal, J.; Maldonado, A.; Saavedra, G.; Baquero, F.; Silva, J.; Torres, C.; Del Campo, R. Clonal dissemination of Enterococcus faecalis ST201 and Enterococcus faecium CC17–ST64 containing Tn5382–vanB2 among 16 hospitals in Chile. Clin. Microbiol. Infect. 2009, 15, 586–588. [Google Scholar] [CrossRef] [Green Version]
- Getachew, Y.; Hassan, L.; Zakaria, Z.; Abdul Aziz, S. Genetic variability of vancomycin-resistant Enterococcus faecium and Enterococcus faecalis isolates from humans, chickens, and pigs in Malaysia. Appl. Environ. Microbiol. 2013, 79, 4528–4533. [Google Scholar] [CrossRef] [Green Version]
- Sakai, Y.; Tsukahara, T.; Ushida, K. Possibility of vancomycin-resistant enterococci transmission from human to broilers, and possibility of using the vancomycin-resistant gram-positive cocci as a model in a screening study of vancomycin-resistant enterococci infection in the broiler chick. Anim. Sci. J. 2006, 77, 538–544. [Google Scholar] [CrossRef]
- Blanch, A.; Caplin, J.; Iversen, A.; Kühn, I.; Manero, A.; Taylor, H.; Vilanova, X. Comparison of enterococcal populations related to urban and hospital wastewater in various climatic and geographic European regions. J. Appl. Microbiol. 2003, 94, 994–1002. [Google Scholar] [CrossRef] [PubMed]
- Novais, C.; Coque, T.M.; Ferreira, H.; Sousa, J.C.; Peixe, L. Environmental contamination with vancomycin-resistant enterococci from hospital sewage in Portugal. Appl. Environ. Microbiol. 2005, 71, 3364–3368. [Google Scholar] [CrossRef] [Green Version]
- Cho, S.; Barrett, J.B.; Frye, J.G.; Jackson, C.R. Antimicrobial Resistance Gene Detection and Plasmid Typing Among Multidrug Resistant Enterococci Isolated from Freshwater Environment. Microorganisms 2020, 8, 1338. [Google Scholar] [CrossRef] [PubMed]
- Wardal, E.; Gawryszewska, I.; Hryniewicz, W.; Sadowy, E. Abundance and diversity of plasmid-associated genes among clinical isolates of Enterococcus faecalis. Plasmid 2013, 70, 329–342. [Google Scholar] [CrossRef] [PubMed]
- Rosvoll, T.C.; Lindstad, B.L.; Lunde, T.M.; Hegstad, K.; Aasnaes, B.; Hammerum, A.M.; Lester, C.H.; Simonsen, G.S.; Sundsfjord, A.; Pedersen, T. Increased high-level gentamicin resistance in invasive Enterococcus faecium is associated with aac(6′)Ie-aph(2″)Ia-encoding transferable megaplasmids hosted by major hospital-adapted lineages. FEMS Immunol. Med. Microbiol. 2012, 66, 166–176. [Google Scholar] [CrossRef] [Green Version]
- Tremblay, C.L.; Charlebois, A.; Masson, L.; Archambault, M. Characterization of hospital-associated lineages of ampicillin-resistant Enterococcus faecium from clinical cases in dogs and humans. Front. Microbiol. 2013, 4, 245. [Google Scholar] [CrossRef] [Green Version]
- Song, X.; Sun, J.; Mikalsen, T.; Roberts, A.P.; Sundsfjord, A. Characterisation of the plasmidome within Enterococcus faecalis isolated from marginal periodontitis patients in Norway. PLoS ONE 2013, 8, e62248. [Google Scholar] [CrossRef] [Green Version]
- Wardal, E.; Kuch, A.; Gawryszewska, I.; Żabicka, D.; Hryniewicz, W.; Sadowy, E. Diversity of plasmids and Tn1546-type transposons among VanA Enterococcus faecium in Poland. Eur. J. Clin. Microbiol. Infect. Dis. 2017, 36, 313–328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ike, Y.; Flannagan, S.E.; Clewell, D.B. Hyperhemolytic phenomena associated with insertions of Tn916 into the hemolysin determinant of Enterococcus faecalis plasmid pAD1. J. Bacteriol. 1992, 174, 1801–1809. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, B.; Tomita, H.; Inoue, T.; Ike, Y. Isolation of VanB-type Enterococcus faecalis strains from nosocomial infections: First report of the isolation and identification of the pheromone-responsive plasmids pMG2200, Encoding VanB-type vancomycin resistance and a Bac41-type bacteriocin, and pMG2201, encoding erythromycin resistance and cytolysin (Hly/Bac). Antimicrob. Agents Chemother. 2009, 53, 735–747. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Freitas, A.R.; Coque, T.M.; Novais, C.; Hammerum, A.M.; Lester, C.H.; Zervos, M.J.; Donabedian, S.; Jensen, L.B.; Francia, M.V.; Baquero, F.; et al. Human and swine hosts share vancomycin-resistant Enterococcus faecium CC17 and CC5 and Enterococcus faecalis CC2 clonal clusters harboring Tn 1546 on indistinguishable plasmids. J. Clin. Microbiol. 2011, 49, 925–931. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayes, J.R.; English, L.L.; Carr, L.E.; Wagner, D.D.; Joseph, S.W. Multiple-antibiotic resistance of Enterococcus spp. isolated from commercial poultry production environments. Appl. Environ. Microbiol. 2004, 70, 6005–6011. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weisburg, W.G.; Barns, S.M.; Pelletier, D.A.; Lane, D.J. 16S ribosomal DNA amplification for phylogenetic study. J. Bacteriol. 1991, 173, 697–703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clinical and Laboratory Standards Institute. Performance Standards for Antimicrobial Susceptibility Testing. CLSI Supplement M100, 31st ed.; Clinical and Laboratory Standards Institute: Malvern, PA, USA, 2021. [Google Scholar]
- Magiorakos, A.P.; Srinivasan, A.; Carey, R.B.; Carmeli, Y.; Falagas, M.E.; Giske, C.G.; Harbarth, S.; Hindler, J.F.; Kahlmeter, G.; Olsson-Liljequist, B.; et al. Multidrug-resistant, extensively drug-resistant and pandrug-resistant bacteria: An international expert proposal for interim standard definitions for acquired resistance. Clin. Microbiol. Infect. 2012, 18, 268–281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krumperman, P.H. Multiple antibiotic resistance indexing of Escherichia coli to identify high-risk sources of fecal contamination of foods. Appl. Environ. Microbiol. 1983, 46, 165–170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwarz, S.; Silley, P.; Simjee, S.; Woodford, N.; van Duijkeren, E.; Johnson, A.P.; Gaastra, W. Editorial: Assessing the antimicrobial susceptibility of bacteria obtained from animals. J. Antimicrob. Chemother. 2010, 65, 601–604. [Google Scholar] [CrossRef] [PubMed]
- Nam, S.; Kim, M.J.; Park, C.; Park, J.G.; Maeng, P.J.; Lee, G.C. Detection and genotyping of vancomycin-resistant Enterococcus spp. by multiplex polymerase chain reaction in Korean aquatic environmental samples. Int. J. Hyg. Environ. Health 2013, 216, 421–427. [Google Scholar] [CrossRef] [PubMed]
- Werckenthin, C.; Schwarz, S. Molecular analysis of the translational attenuator of a constitutively expressed erm(A) gene from Staphylococcus intermedius. J. Antimicrob. Chemother. 2000, 46, 785–788. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jensen, L.B.; Frimodt-Moller, N.; Aarestrup, F.M. Presence of erm gene classes in gram-positive bacteria of animal and human origin in Denmark. FEMS Microbiol. Lett. 1999, 170, 151–158. [Google Scholar] [CrossRef] [PubMed]
- Ng, L.K.; Martin, I.; Alfa, M.; Mulvey, M. Multiplex PCR for the detection of tetracycline resistant genes. Mol. Cell. Probes 2001, 15, 209–215. [Google Scholar] [CrossRef] [PubMed]
- Trzcinski, K.; Cooper, B.S.; Hryniewicz, W.; Dowson, C.G. Expression of resistance to tetracyclines in strains of methicillin-resistant Staphylococcus aureus. J. Antimicrob. Chemother. 2000, 45, 763–770. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.; Zhang, S.; Liu, Z.; Liu, P.; Shi, Z.; Wei, J.; Shao, D.; Li, B.; Ma, Z. Molecular characterization of enterohemorrhagic E. coli O157 isolated from animal fecal and food samples in Eastern China. Sci. World J. 2014, 2014, 946394. [Google Scholar] [CrossRef] [Green Version]
- Jureen, R.; Top, J.; Mohn, S.C.; Harthug, S.; Langeland, N.; Willems, R.J. Molecular characterization of ampicillin-resistant Enterococcus faecium isolates from hospitalized patients in Norway. J. Clin. Microbiol. 2003, 41, 2330–2336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stepanovic, S.; Vukovic, D.; Hola, V.; Di Bonaventura, G.; Djukic, S.; Cirkovic, I.; Ruzicka, F. Quantification of biofilm in microtiter plates: Overview of testing conditions and practical recommendations for assessment of biofilm production by staphylococci. APMIS Acta Pathol. Microbiol. Immunol. Scand. 2007, 115, 891–899. [Google Scholar] [CrossRef] [PubMed]
- Lopes Mde, F.; Simoes, A.P.; Tenreiro, R.; Marques, J.J.; Crespo, M.T. Activity and expression of a virulence factor, gelatinase, in dairy enterococci. Int. J. Food Microbiol. 2006, 112, 208–214. [Google Scholar] [CrossRef] [PubMed]
- De Vuyst, L.; Foulquie Moreno, M.R.; Revets, H. Screening for enterocins and detection of hemolysin and vancomycin resistance in enterococci of different origins. Int. J. Food Microbiol. 2003, 84, 299–318. [Google Scholar] [CrossRef]
- Dupre, I.; Zanetti, S.; Schito, A.M.; Fadda, G.; Sechi, L.A. Incidence of virulence determinants in clinical Enterococcus faecium and Enterococcus faecalis isolates collected in Sardinia (Italy). J. Med. Microbiol. 2003, 52, 491–498. [Google Scholar] [CrossRef] [Green Version]
- Vankerckhoven, V.; Van Autgaerden, T.; Vael, C.; Lammens, C.; Chapelle, S.; Rossi, R.; Jabes, D.; Goossens, H. Development of a multiplex PCR for the detection of asa1, gelE, cylA, esp, and hyl genes in enterococci and survey for virulence determinants among European hospital isolates of Enterococcus faecium. J. Clin. Microbiol. 2004, 42, 4473–4479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huerta-Cepas, J.; Serra, F.; Bork, P. ETE 3: Reconstruction, Analysis, and Visualization of Phylogenomic Data. Mol. Biol. Evol. 2016, 33, 1635–1638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Letunic, I.; Bork, P. Interactive Tree of Life (iTOL) v5: An online tool for phylogenetic tree display and annotation. Nucleic Acids Res. 2021, 49, W293–W296. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antimicrobial Agents | E. faecalis (n = 45) | E. faecium (n = 30) | E. hirae (n = 12) | E. avium (n = 3) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| %R | MIC50 | MIC90 | %R | MIC50 | MIC90 | %R | MIC50 | MIC90 | %R | MIC50 | MIC90 | |
| Ampicillin | 15.6 | 2 | 32 | 33.3 | 4 | 64 | 66.7 | 16 | 32 | 66.7 | 32 | 32 |
| Rifampin | 24.4 | 1 | 16 | 10 | 1 | 1 | 0 | 0.5 | 1 | 0 | 1 | 1 |
| Ciprofloxacin | 13.3 | 1 | 8 | 23.3 | 1 | 16 | 33.3 | 1 | 8 | 66.7 | 4 | 8 |
| Fosfomycin | 17.8 | 32 | 256 | 13.3 | 32 | 256 | 8.3 | 32 | 256 | 0 | 32 | 64 |
| Nitrofurantoin | 22.2 | 16 | 256 | 16.7 | 16 | 256 | 8.3 | 16 | 32 | 0 | 32 | 32 |
| Linezolid | 0 | 1 | 2 | 0 | 1 | 2 | 0 | 1 | 2 | 0 | 1 | 2 |
| Vancomycin | 3.3 | 2 | 4 | 3.3 | 2 | 4 | 0 | 2 | 4 | 0 | 2 | 4 |
| Chloramphenicol | 8.9 | 4 | 8 | 6.7 | 4 | 8 | 0 | 4 | 8 | 0 | 4 | 8 |
| Tetracycline | 62.2 | 32 | 64 | 50 | 4 | 32 | 41.7 | 4 | 32 | 66.7 | 16 | 32 |
| Erythromycin | 31.1 | 0.5 | 32 | 16.7 | 16 | 256 | 33.3 | 0.5 | 16 | 0 | 0.5 | 0.5 |
| Resistance Profile | Number of Isolates | %Isolates |
|---|---|---|
| 16 | 17.8 | |
| FOS | 3 | 3.3 |
| TCY | 4 | 4.4 |
| ERY | 2 | 2.2 |
| CIP FOS | 1 | 1.1 |
| TCY RIF | 2 | 2.2 |
| TCY FOS | 2 | 2.2 |
| TCY CIP | 3 | 3.3 |
| NIT TCY | 5 | 5.6 |
| AMP CIP | 5 | 5.6 |
| AMP CHL | 1 | 1.1 |
| AMP TCY | 4 | 4.4 |
| AMP NIT | 2 | 2.2 |
| ERY RIF | 2 | 2.2 |
| ERY CHL | 2 | 2.2 |
| ERY TCY | 3 | 3.3 |
| ERY NIT | 2 | 2.2 |
| VAN TCY | 1 | 1.1 |
| TCY CHL RIF | 1 | 1.1 |
| NIT TCY RIF | 2 | 2.2 |
| AMP TCY FOS | 3 | 3.3 |
| AMP TCY CIP | 3 | 3.3 |
| ERY TCY RIF | 2 | 2.2 |
| ERY TCY FOS | 1 | 1.1 |
| ERY TCY CIP | 3 | 3.3 |
| ERY NIT RIF | 2 | 2.2 |
| ERY NIT TCY | 1 | 1.1 |
| ERY PEN TCY | 6 | 6.7 |
| AMP TCY CHL CIP | 1 | 1.1 |
| AMP NIT CIP FOS | 1 | 1.1 |
| ERY CHL CIP FOS | 1 | 1.1 |
| VAN NIT TCY RIF | 1 | 1.1 |
| VAN ERY TCY RIF | 1 | 1.1 |
| AMP TCY CIP FOS RIF | 1 | 1.1 |
| E. faecalis (N = 45) | E. faecium (N = 30) | E. hirae (N = 12) | E. avium (N = 3) | Total Enterococci (N = 90) | |
|---|---|---|---|---|---|
| vanA | 1 | 0 | 0 | 0 | 1 |
| vanB | 0 | 0 | 0 | 0 | 0 |
| ermA | 5 | 4 | 2 | 0 | 11 |
| ermB | 13 | 9 | 2 | 0 | 24 |
| pbp5 | 7 | 8 | 6 | 2 | 23 |
| tetA | 1 | 0 | 0 | 0 | 1 |
| tetB | 0 | 0 | 0 | 1 | 1 |
| tetM | 26 | 8 | 3 | 2 | 39 |
| tetL | 15 | 12 | 5 | 2 | 34 |
| optrA | 0 | 0 | 0 | 0 | 0 |
| Cat | 3 | 1 | 0 | 0 | 4 |
| Biofilm Category | Mean Biofilm OD | Mean MAR Index | MDR | Gelatinase | Cytolysin | Agg | gelE |
|---|---|---|---|---|---|---|---|
| Non-biofilm producers | 0.29 | 0.017 | 0 | 0 | 0 | 0 | 0 |
| Weak biofilm producers | 0.47 | 0.13 | 0 | 3 | 0 | 2 | 2 |
| Medium biofilm producers | 0.9 | 0.22 | 8 | 3 | 3 | 8 | 7 |
| Strong biofilm producers | 1.65 | 0.3 | 22 | 9 | 9 | 23 | 16 |
| ST | NO | MLST Allelic Profile | Phenotypic Activities | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| gdh | gyd | pstS | gki | aroE | xpt | yqiL | Biofilm | MDR | Gelatinase | Cytolysin | ||
| 16 | 10 | 5 | 1 | 1 | 3 | 7 | 7 | 6 | 10 | 6 | 2 | 3 |
| 21 | 3 | 1 | 7 | 9 | 1 | 1 | 1 | 1 | 1 | 0 | 0 | 0 |
| 32 | 1 | 8 | 7 | 9 | 5 | 4 | 4 | 1 | 1 | 0 | 0 | 0 |
| 41 | 5 | 1 | 7 | 11 | 21 | 1 | 4 | 1 | 3 | 0 | 0 | 1 |
| 81 | 1 | 27 | 2 | 16 | 28 | 26 | 2 | 1 | 1 | 1 | 1 | 0 |
| 176 | 2 | 15 | 7 | 3 | 37 | 39 | 15 | 11 | 2 | 0 | 1 | 0 |
| 177 | 1 | 15 | 2 | 37 | 37 | 39 | 15 | 11 | 0 | 0 | 0 | 0 |
| 179 | 6 | 5 | 1 | 1 | 3 | 7 | 1 | 6 | 5 | 2 | 3 | 2 |
| 302 | 8 | 5 | 1 | 1 | 3 | 7 | 7 | 60 | 8 | 4 | 5 | 2 |
| 480 | 5 | 1 | 1 | 22 | 22 | 7 | 17 | 6 | 5 | 2 | 0 | 1 |
| 752 | 3 | 27 | 2 | 16 | 28 | 26 | 83 | 1 | 3 | 0 | 0 | 0 |
| ST | NO | MLST Allelic Profile | Phenotypic Activities | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| atpA | ddl | gdh | purK | gyd | pstS | adk | Biofilm | MDR | Gelatinase | Cytolysin | ||
| 12 | 1 | 5 | 2 | 6 | 6 | 1 | 7 | 1 | 0 | 0 | 0 | 0 |
| 360 | 2 | 5 | 2 | 6 | 6 | 1 | 1 | 1 | 0 | 0 | 0 | 0 |
| 16 | 3 | 1 | 2 | 1 | 1 | 1 | 1 | 1 | 2 | 1 | 0 | 0 |
| 194 | 8 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 8 | 4 | 2 | 1 |
| 157 | 5 | 7 | 1 | 1 | 1 | 5 | 1 | 1 | 5 | 2 | 0 | 1 |
| 9 | 4 | 4 | 5 | 1 | 3 | 1 | 1 | 1 | 4 | 0 | 0 | 0 |
| 18 | 2 | 15 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 0 | 1 | 0 |
| 82 | 5 | 1 | 36 | 1 | 1 | 1 | 1 | 1 | 4 | 1 | 0 | 1 |
| Enterococcus spp. | Phenotypic Profile | AMR Genes | Virulence Genes | Gelatinase | Cytolysin | Rep-Family | Plasmid |
|---|---|---|---|---|---|---|---|
| E. faecalis | AMP, ERY, TET | pbp5, tetM, tetL | gelE, agg | − | − | 9 | pCF10 |
| E. faecalis | AMP, TET, FOS | pbp5, tetM | gelE, agg, cylA | − | − | ||
| E. faecalis | ERY, NIT, TET | ermA, ermB, tetM | gelE, agg | + | − | 9 | pCF10 |
| E. faecalis | ERY, CHL, CIP, FOS | ermB | gelE, agg, cylA | + | − | ||
| E. faecalis | VAN, ERY, TET, RIF | vanA, ermB, tetM, tetL | gelE, agg | + | − | 9, 1 | pIP501, pCF10 |
| E. faecalis | ERY, NIT, RIF | ermA | gelE, agg, cylA | − | + | ||
| E. faecalis | ERY, NIT, RIF | ermB | gelE, agg | − | − | ||
| E. faecalis | AMP, TET, CIP, RIF, FOS | pbp5 | gelE, agg, cylA | − | + | 6 | pS86 |
| E. faecalis | TET, CHL, RIF | tetM, cat | − | − | |||
| E. faecalis | AMP, TET, FOS | pbp5, tetM | gelE, agg | + | − | ||
| E. faecalis | AMP, TET, FOS | pbp5, tetM, tetL | gelE, agg, cylA | + | + | 9 | pCF10 |
| E. faecalis | ERY, TET, RIF | ermB, tetM | gelE, agg | + | − | 9 | pCF10 |
| E. faecalis | NIT, TET, RIF | tetM | gelE, agg | + | − | 6 | pS86 |
| E. faecalis | ERY, TET, RIF | ermB, tetL | − | − | 9 | pCF10 | |
| E. faecalis | NIT, TET, RIF | gelE, agg, cylA | − | − | |||
| E. faecium | AMP, TET, CIP | pbp5, tetM, tetL | agg | − | − | 2 | pRE25 |
| E. faecium | VAN, NIT, TET, RIF | vanA, tetM, tetL | agg | − | − | 2, 1 | pIP501, pRE25 |
| E. faecium | AMP, TET, CHL, CIP | pbp5, tetM, tetL, cat | geIE, agg | + | − | 2 | pRE25 |
| E. faecium | ERY, AMP, TET | ermA, ermB, tetL | cylA | − | − | ||
| E. faecium | ERY, TET, CIP | tetL | gelE, agg, cylA | − | + | 2, 6 | pRE25, pS86 |
| E. faecium | ERY, TET, CIP | ermB | agg | − | − | ||
| E. faecium | ERY, TET, CIP | ermA, ermB | geIE, agg | + | − | 6 | pS86 |
| E. faecium | ERY, TET, FOS | ermA, ermB, tetM, tetL | agg, cylA | − | + | 2 | pRE25 |
| E. hirae | ERY, AMP, TET | tetL | agg | − | − | ||
| E. hirae | ERY, AMP, TET | ermA, ermB, pbp5, tetM, tetL | − | − | 5 | pN315 | |
| E. hirae | ERY, AMP, TET | ermA, ermB, pbp5, tetL | gelE, agg, cylA | − | + | 5,3 | pN315, pAW63 |
| E. hirae | AMP, NIT, CIP, FOS | pbp5 | geIE, agg | − | − | 6 | pS86 |
| E. hirae | ERY, AMP, TET | ermA, ermB, pbp5, tetM, tetL | agg | − | + | 5, 11 | pN315, pEF1071 |
| E. avium | AMP, TET, CIP | pbp5, tetB, tetM, tetL | − | − | 6 | pS86 | |
| E. avium | AMP, TET, CIP | pbp5, tetM, tetL | geIE | − | − | 6, 9 | pS86, pCF10 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alzahrani, O.M.; Fayez, M.; Alswat, A.S.; Alkafafy, M.; Mahmoud, S.F.; Al-Marri, T.; Almuslem, A.; Ashfaq, H.; Yusuf, S. Antimicrobial Resistance, Biofilm Formation, and Virulence Genes in Enterococcus Species from Small Backyard Chicken Flocks. Antibiotics 2022, 11, 380. https://doi.org/10.3390/antibiotics11030380
Alzahrani OM, Fayez M, Alswat AS, Alkafafy M, Mahmoud SF, Al-Marri T, Almuslem A, Ashfaq H, Yusuf S. Antimicrobial Resistance, Biofilm Formation, and Virulence Genes in Enterococcus Species from Small Backyard Chicken Flocks. Antibiotics. 2022; 11(3):380. https://doi.org/10.3390/antibiotics11030380
Chicago/Turabian StyleAlzahrani, Othman M., Mahmoud Fayez, Amal S. Alswat, Mohamed Alkafafy, Samy F. Mahmoud, Theeb Al-Marri, Ahmed Almuslem, Hassan Ashfaq, and Shaymaa Yusuf. 2022. "Antimicrobial Resistance, Biofilm Formation, and Virulence Genes in Enterococcus Species from Small Backyard Chicken Flocks" Antibiotics 11, no. 3: 380. https://doi.org/10.3390/antibiotics11030380
APA StyleAlzahrani, O. M., Fayez, M., Alswat, A. S., Alkafafy, M., Mahmoud, S. F., Al-Marri, T., Almuslem, A., Ashfaq, H., & Yusuf, S. (2022). Antimicrobial Resistance, Biofilm Formation, and Virulence Genes in Enterococcus Species from Small Backyard Chicken Flocks. Antibiotics, 11(3), 380. https://doi.org/10.3390/antibiotics11030380