Abstract
The “One Health” approach increasingly demonstrates the global spread of pathogenic microorganisms and their antimicrobial resistance in the environment, both in animals and humans. Salmonella enterica subsp. diarizonae is nowadays very often isolated from cold-blooded reptiles to a lesser extent from sheep, but unfortunately more and more often from humans. However, there are a few studies describing the isolation of Salmonella enterica subsp. diarizonae from migratory wild birds. The mallard duck (Anas platyrhynchos), a wild animal that traverses the continent of Eurasia, can be an excellent indicator of the spread of intestinal microbes as well as their resistance to antibiotics. This is the first report of the Salmonella enterica subsp. diarizonae detection in Poland in a migrating mallard duck. This research presented the identification difficulties associated with the isolation of Salmonella enterica subsp. diarizonae using three different biochemical tests and advanced serology tests. At the same time, we detected very high antimicrobial resistance in the isolated strain. By using the minimum inhibitory concentration (MIC) method, it was found that the isolated strain of S. enterica subsp. diarizonae has high antibiotic resistance against 14 of the 33 tested antimicrobials agents. The resistance genes that have been identified in S. enterica subsp. diarizonae include aadA, strA/strB, and blaTEM.
1. Introduction
Salmonellosis is an important cause of water-borne [1] and food-borne epidemics in humans. In the European Union, salmonellosis is the second most frequently reported human gastrointestinal infection right after campylobacteriosis. In 2021 a total of 90,105 human salmonellosis cases were reported by EU EFSA [2]. People contract Salmonella not only from animal origin foods but also from eating contaminated vegetables and fruits or from their pets. Now there are more and more reports of human diseases caused by Salmonella enterica subsp. diarizonae [3]. Salmonella enterica subsp. diarizonae is a subspecies IIIb of six within Salmonella enterica spp. It is most often described as a Gram-negative rod infecting cold-blooded animals, including reptiles kept as pets [4,5]. However, Salmonella enterica subsp. diarizonae was also isolated in Norway and Switzerland from sheep [6,7] and Greece [8]. Chatzopoulos et al. showed that Salmonella enterica subsp. diarizonae is an opportunistic gastrointestinal pathogen in lamb [9].
Thus, the spread of Salmonella enterica subsp. diarizonae in the environment is only a matter of time.
Taking into account the well-known concept of caring for “One Health” approach, which relates to both the protection of human and animal health and considering the impact of the environment on humans and animals, the WHO poses the problem of antibiotic-resistant pathogens as one of the main challenges the health care of the 21st century is facing. The level of antibiotic resistance of bacteria in the environment can be indirectly tested by pathogens isolated from wild animals, such as migratory birds. The mallard duck (Anas platyrhynchos), also known as the wild duck, is commonly found in Poland and Europe. It is a migrating bird; therefore, it is also found in North America and Asia. It lives in areas of wild nature and urban agglomerations in the vicinity of water reservoirs [10]. Mallards can lead a sedentary or wandering/migratory lifestyle; therefore, it is an excellent bioindicator of the occurrence of pathogens and drug-resistant bacteria in the local and cross-border environment.
The aim of this study was to serologically and biochemically identify and determine the antibiotic resistance of Salmonella spp. isolated from mallard ducks.
2. Results
In the examined 17 mallard ducks (nine male and eight female), no pathological changes were observed in the muscle, liver, and intestine. From all the examined internal organs, only seven strains of Salmonella spp. from the intestines have been isolated. All strains belonged to Salmonella enterica spp. Six were classified as Salmonella enterica subsp. enterica (two strains of S. Derby and four strains of S. Enteritidis) and one strain presumptive Salmonella enterica subsp. diarizonae. A strain suspected of being Salmonella enterica subsp. diarizonae was checked for further analysis because, so far, it has not been isolated from mallard ducks in Poland. The isolated strain of Salmonella spp. in the applied biochemical tests showed diversified results. Common positive reactions from the three tests include H2S production, D-glucose, D-mannitol, and lysine decarboxylase for VITEK and Lab-made beta-galactosidase, fermentation/glucose, D-maltose, and D-trehalose, and for API 20 E and Lab-made tests—arabinose (other results were presented in Supplementary Materials Table S1).
Therefore, we obtained the classification of various Salmonella enterica spp. In the API 20E test, the strain was designated as 89.6% Salmonella spp., 10.3% Salmonella enterica subsp. arizonae and VITEK in 99% of Salmonella enterica subsp. diarizona (detailed reactions in individual tests are presented in Supplementary Materials Table S1).
2.1. Serological Identification
As a result of the serotyping, the tested isolate was identified as S. enterica subsp. diarizonae O:58 group strain. This result requires confirmation by the WHO Collaborating Centre for Reference and Research on Salmonella, Institut Pasteur (Paris, France).
2.2. Antimicrobial Susceptibility Testing
The following chemotherapeutic agents (MERLIN Diagnostika GmbH, Bremen, Niemcy) were included in our study: Ampicillin (AM), Amoxicillin/Clavulanic acid (AMC), Cefalexin (CN), Cefalotin (CF), Ceftriaxone (CFP), Cefequinome (CEQ), Ceftiofur (CFT), Enrofloxacin (ENR), Florfenicol (FFC), Flumequine (UB), Gentamicin (GM), Imipenem (IPM), Marbofloxacin (MRB), Neomycin (NEO), Polymixin B (PB), Tetracycline (TE), and Trimethoprim/Sulfamethoxazole (SXT) and additional antibiotics, Cefalotin (CF), cefoperazone (CFP), Imipenem (IPM), Flumequine (UB), Marbofloxacin (MRB), Tetracycline (TE), Polymixin B (PB), and Trimethoprim/Sulfamethoxazole (SXT).
The S. enterica subsp. diarizonae O: 58 group strain was resistant to 14 antimicrobials (CFX-CPH-CLO-PG-NAF-GEN-NEO-STR-UB-ERY-TYL-LIN-TIA-TYLV) belonging to five classes of antimicrobials agents (β-lactams, aminoglycosides, fluoroquinolones, macrolides, and lincosamides) and florfenicol, tiamulin, tylvalosin. Resistance to individual antibiotics is shown in Table 1.

Table 1.
Antimicrobial resistance of the Salmonella enterica subsp. diarizonae O:58 isolate from mallard ducks.
2.3. Detection of Antimicrobial Resistance Genes (ARGs) by Multiplex PCR
Antimicrobial resistance genes present in Salmonella enterica subsp. diarizonae O:58 isolated from wild ducks (Anas platyrhynchos) (Figure 1). The isolate harboured the aadA, strA/strB, blaTEM genes, while aphA1, aphA2, aadB, tetA, tetB, sul1, sul2, blaSHV, blaCMY-2 were absent.
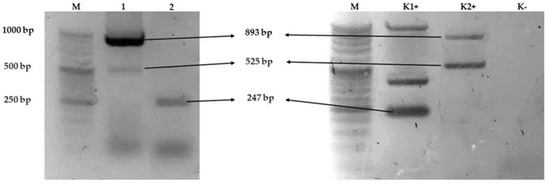
Figure 1.
Agarose gel (2%) electrophoresis showing the amplification of multiplex PCR line 1 strA/strB (893 bp) and aadA (525 bp), multiplex PCR line 2 blaTEM (247 bp) of Salmonella enterica subsp. diarizonae O58. Notes: line M DNA marker, line K1+ blaTEM (247 bp), and line K2+ positive control genes sul2 (721 bp), aadA (525 bp), as positive controls were Klebsiella pneumoniae and E. coli strains isolated from poultry and cattle. Lane K—no template (negative control). Abbreviations: M—DNA marker (DNA Marker 2+, (50–1000 bp, A&A Biotechnology Gdańsk, Poland); bp—base pairs.
3. Discussion
Mallard duck belongs to the order of the Anseriformes during foraging. It filters the surface of the water reservoir, which makes them vulnerable to collecting numerous contaminants with food. They are also often found in large flocks of the same species and groups with other waterfowl, which may additionally predispose them to cross infections. The area where the duck was collected for research is a typical field circuit with a large number of water reservoirs, rivers, and streams. It is a very favourable habitat for waterfowl that frequently use the area, including sedentary as well as migratory populations. The northern duck, as the migratory population, is characterized by a higher body weight compared with the permanent population. Numerous studies on mallard migration indicate that ducks come to Poland from Finland, Russia, Belarus, and Ukraine, and often from Western Europe, countries such as France, Germany, Austria, and the Czech Republic [11,12,13]. It can be said with a high probability that the ducks described in the research came from the eastern part of Europe, including the countries of Belarus, Ukraine, and Russia. Thus, migration may play a role in the transmission of Salmonella spp. and antibiotic-resistant bacteria. Previously, greylag goose was described as the possible vector of Salmonella enterica subsp. diarizonae 14:k:z53 in Norway [14].
Table S1 shows the comparison between VITEK2 and API20E results, respectively. This table shows the different points and similarities of the results in the same samples when applying the other methods. This demonstrates that the commonly used API system does not allow for unambiguous identification of Salmonella spp., other than Salmonella enterica subsp. enterica.
The drawback of the traditional phenotypic method is that it requires the availability of more than 150 specific antisera and well-trained, experienced personnel to correctly interpret the results. Consequently, it is not possible for all laboratories to carry out this method in-house, and often, laboratories have to send the isolates to a national reference laboratory or an expert laboratory. This process can significantly delay the time of obtaining results. In terms of performance, the method may give false positive reactions due to weak or nonspecific agglutination. Autoagglutination or loss of antigen expression, as observed for rough and nonmotile isolates, results in unidentified serotypes [15,16]. Our research clearly shows difficulties in classical biochemical and serological identification of Salmonella ssp., especially between IIIa and IIIb. Data on the isolation of Salmonella enterica subsp. diarizonae strains in Poland are limited. According to the National Salmonella Center from 1995–2007 Salmonella enterica subsp. diarizonae was identified sporadically, much less than 1% [15]. Serological analysis of these Salmonella enterica subsp. diarizonae strains occurring in Poland presented several different serovars. Six serovars that belonged to groups O35, O38, O48, O58, O60, and O61 have been identified, mainly in animals and humans. Amongst them, the serovar Salmonella enterica subsp. diarizonae 58:z52:z35, i.e., S. IIIb (58:z52:z35), was detected twice in 1998 (in the internal organs of the biting adder Bitis arietans and in the faeces of the Canadian beaver Castor canadensis). They were lactose-positive strains. In the same year, single isolation of the strain S. IIIb (48:k:z57) was obtained from the water (wild bathing area, river Raba, near Myślenice). The strain was lactose negative, ONPG positive after 2 h and presented a hitherto unknown antigenic formula. Strains of Salmonella enterica. subsp. diarizonae serovars S. IIIb (48: k:z53) and S. IIIb (61: k:1,5,7) were also isolated from two patients requiring hospitalization (child’s faeces, adult cerebrospinal fluid) [15].
Amongst Salmonella strains originating mainly from the environment, food and isolates from animals which were sent to the National Salmonella Center in Poland in the years 2008–2020, fourteen strains of Salmonella enterica subsp. diarizonae were identified (<1%). They represented different serovars of groups O38, O47, O48, O50, and O61. Strains of serovar S. IIIb (38:r:z) were isolated from food and the environment. Seven serovars S. IIIb (47:k:z35), S. IIIb (48:k:z53), S. IIIb (50:z:z52), S. IIIb (50:z52:z35), S. IIIb (50:r:z35), S. IIIb (61:c:z35), and S. IIIb (61:k:1,5) were found in human (unpublished study, Tokarska-Pietrzak E. and Piechowicz L.). Four serovars: S. IIIb (48:i:z), S. IIIb (61:r:z), S. IIIb (61:c:z35), and S. IIIb (65:k:z), were identified in animals. In 2021, one Salmonella enterica subsp. diarizonae strains were isolated from a duck. It belonged to group O58 and represented a serovar that was isolated in wild mallard ducks in Poland for the first time.
Because of Salmonella spp. significance to the poultry industry and the risk of alimentary infections in humans, a large number of studies have been carried out on its epidemiology in wild birds such as mallard ducks. Thus, there are guidelines within the poultry industry to avoid or minimize contact between wild birds and domestic poultry, especially with ducks and geese bred in an open system. Wild birds are not considered a direct source of infection for livestock but rather a source of feed contamination amongst farm animals. The resistance of bacteria to one or more antibiotics poses a risk for human and animal health. Ducks and other wild birds may be colonized by antibiotic-resistant bacteria present in their environment. It can be hypothesized that wild birds can act as vectors or amplifiers, which carry resistant bacteria to livestock.
The results of the phenotypic resistance test indicated that Salmonella enterica subsp. diarizonae O:58 isolated from mallard ducks could be categorized as MDR, that is, bacteria exhibiting resistance to one or more antibiotics from three or more classes of antibiotics [17,18].
The isolate harboured the aadA, strA/strB, major genes for resistance to streptomycin, but not for gentamycin (aadB) and neomycin (aphA1, aphA2), which belong to aminoglycosides. In addition, we found the blaTEM gene that belongs to ESBL-encoding genes associated with extended-spectrum resistance to β-lactams in multidrug-resistant strains of Gram-negative bacteria such as Klebsiella pneumoniae. However, the strains were phenotypically sensitive to ampicillin, probably because of the gene being silenced. ESBL-encoding genes are usually localized on large plasmids that simplify their spread amongst gram-negative rods via conjugation. What is more, these plasmids may carry other genes associated with resistance to other classes of antibiotics. Because of the plasticity of Salmonella, its adaptability, and the development of mechanisms of resistance to antibiotics using genetic strategies such as gene mutations or horizontal transfer of resistance genes [19], the presence of the blaTEM gene seems to be alarming.
The aadB gene coding resistance to gentamycin, and aphA1, aphA2 to neomycin, were not found in the present study. However, the strain was resistant to this antibiotic. This resistance to gentamycin may be mediated by other resistance genes such as grm [20], which was not evaluated in this study.
The next step of the research will be the serological analysis of the isolated strain Salmonella enterica subsp. diarizonae O:58 at the Pasteur Institute in Paris and NGS Whole Genome Sequencing
4. Materials and Methods
The material for the research was fragments of liver, muscles, and intestines obtained from 17 shot mallard ducks (Anas platyrhynchos) in nature from hunting grounds in Lubelszczyzna in Poland. The ducks were harvested in accordance with the Polish hunting law [10] by shooting during the hunting season, which lasts from 1 September to the end of December 2021. The mallard (Anas platyrhynchos) is on the list of game species in accordance with the Regulation of the Minister of the Environment on establishing the list of game species [21]. After harvesting, all uneviscerated birds were packed in a foamed polystyrene refrigerated box and transported to the laboratory at 4 °C.
4.1. Salmonella spp. Isolation and Identification
Salmonella spp. from all samples were isolated in accordance with PN-EN ISO 6579-1:2017-04 Microbiology of the food chain—Horizontal method for the detection, enumeration, and serotyping of Salmonella—Part 1: Detection of Salmonella spp. [22].
Samples were suspended in a ninefold volume of buffered peptone water (BPW GRASO, Starogard, Poland) for each 1 g of sample (5 g of liver, 5 g of intestines and 25 g of muscle) in sterile stomacher bags (Whirl-Pak, NASco, Madison, WI, USA). Then they were placed in a stomacher and crushed for 2 min. The selective proliferation of Salmonella spp. was carried out using modified semisolid Rappaport-Vassiliadis (MSRV) agar (GRASO) and Muller–Kauffmann tetrathionate-novobiocin (MKTTn) broth (GRASO). Two selective enrichment media, xylose lysine deoxycholate agar (XLD; GRASO) and Salmonella chromagar agar (CHROMagar Salmonella PLUS; GRASO), were used as described in Pławińska-Czarnak et al. (2021) [23]. Salmonella suspect colonies were transferred to nonselective nutrient agar (GRASO) to obtain the pure culture for further biochemical and to a semisolid medium by Garda (GRASO) for further serological tests.
4.1.1. Salmonella spp. Identification with Molecular Biology Methods
A real-time PCR method based on the detection of genes specific for Salmonella spp. was used to confirm presumptive identification. DNA for real-time PCR was extracted from bacterial cells using a commercial Kylt® DNA Extraction-Mix II (AniCon, Hoeltinghausen, Germany). Real-time PCR to detect Salmonella was performed according to the manufacturer’s instructions (AniCon, Hoeltinghausen, Germany) using Applied Biosystems 7500 Fast Real-Time PCR System (Thermo, Waltham, MA, USA).
4.1.2. Biochemical Strain Identification
Biochemical tests were also conducted with the use of homemade media inoculated with bacteria grown on nutrient agar, incubated for 24 h (with the exception of dulcitol, inositol, and rhamnose fermentation tests, gelatine hydrolysis test, and d-tartrate test—in case the results were negative after 24 h, the incubation was prolonged up to 5 days). The tests with negative results were verified and repeated.
For biochemical identification of the strains, the VITEK® 2 GN cards and Api20E (BioMérieux, Craponne, France) were used according to the manufacturer’s instructions. Biochemical tests were also done with the use of lab-made media, inoculated with bacteria grown on nutrient agar, incubated for 24 h (with the exception of dulcitol, inositol, and rhamnose fermentation tests, gelatine hydrolysis test, and d-tartrate test—in case while the results were negative after 24 h, the incubation was prolonged up to 5 days) according to Załęska et al. (1969) [24]. The tests with negative results were verified and repeated.
4.1.3. Serological Typing
Serological typing of Salmonella for the detection of somatic and flagellar antigens was performed using the slide agglutination test. The polyvalent and monovalent Salmonella anti-O and anti-H diagnostic sera were used (Statens Serum Institut, Copenhagen, Denmark; BIOMED, Kraków, Poland; Immunolab, Gdynia, Poland). The results of the antigen determination were used for the final serological characterization using the White–Kauffmann–Le Minor scheme [25,26]. A 24 h culture of the tested strain on nutrient agar medium was used for the tests (BioMaxima, Lublin, Poland).
4.2. Antimicrobial Susceptibility Testing
The 8 classes of antimicrobials agents (β-lactams, aminoglycosides, polymyxins, fluoroquinolones, tetracyclines macrolides, lincosamides, and sulfonamide) as well as florfenicol, tiamulin, and tylvalosin were used for the antimicrobial susceptibility test.
Antimicrobial susceptibility was assessed by determining the MIC values using a 96 well MICRONAUT Special Plates with antimicrobials: AMX/CL—amoxicillin and clavulanic acid; AMX—amoksycylina; CFQ—cefquinome; CFTI—ceftiofur; CFX—cephalexin, CLO—cloxacillin; COL—colistin; CPH—cefapiryna; DOX—docycycline; ENR—enrofloxacin; ERY—erythromycin; FLR—florfenicol; GEN—gentamicin; LIN—lincomycin, LIN/SP—lincomycin/specinicin; NAF—nafcillin; NEO—neomycin; NOR—norfloxacin; OXY—oxytetracycline; PG—benzylpenicillin; STR—streptomycin; TR/SMX—trimethoprim-sulfamethoxazole; TIA—tiamulin; TYL—tylosin; TYLV—tylvalosin (MERLIN Diagnostika GmbH, Bremen, Niemcy). To analyze MIC patterns of S. enterica subsp. diarizonae Merlin MICRONAUT (MERLIN Diagnostika GmbH, Bremen, Niemcy) was used. The MICs were interpreted according to the Clinical and Laboratory Standards Institute (CLSI) and FDA breakpoints [27].
Minimum Inhibitory Concentrations (MICs) of Amoxicillin/Clavulanic acid (AMC), Ampicillin (AM), Cefalexin (CN), Cefalotin (CF), Ceftriaxone (CFP), Cefequinome (CEQ), Ceftiofur (CFT), Enrofloxacin (ENR), Florfenicol (FFC), Flumequine (UB), Gentamicin (GM), Imipenem (IPM), Marbofloxacin (MRB), Neomycin (N), Polymixin B (PB), Tetracycline (TE), and Trimethoprim/Sulfamethoxazole (SXT) and additional antibiotics, Ampicillin (AM), Cefalotin (CF), cefoperazone (CFP), Imipenem (IPM), Flumequine (UB), Marbofloxacin (MRB), Tetracycline (TE), Polymixin B (PB), Trimethoprim/Sulfamethoxazole (SXT) were assessed by AST-GN 96 CARD and VITEK2 system. The AST card is essentially a miniaturized and abbreviated version of the doubling dilution technique for MICs determined by the microdilution method [28].
4.3. Detection of Antimicrobial Resistance Genes (ARGs) by Multiplex PCR
Mueller–Hinton agar was used to culture the bacterial isolates overnight at 35 °C. Bacterial DNA isolation was performed using a standard bacterial DNA isolation Kylt® DNA Extraction-Mix II (Anicon, Emstek, Germany).
Twelve resistance genes (aadA, aadB, strA/strB, aphA1, aphA2, tetA, tetB, sul1, sul2, bla TEM, blaSHV, and blaCMY-2) were analyzed by conventional PCR using specific primer pairs). The primer sequences predicted PCR product sizes and references shown in Table 2.
Table 2.
Primers sequences for detection of antimicrobial resistance genes in the Salmonella spp. isolate and multiplex PCR annealing temperature [29,30].
Multiplex PCR protocols were established to screen for genes linked with antibiotic resistance of S. enterica subsp. diarizonae. Each PCR reaction contained 3 μL of DNA (20 ng), 12.5 μL of DreamTaq Green 2 × Master Mix (Thermo Fisher Scientific, Waltham, MA, USA), 20 pmol of each of the forward and reverse primers (Genomed, Warsaw, Poland), and nuclease-free water to a final volume of 25 µL. PCR conditions in all Multiplex were as follows: 1 cycle at 95 °C for 3 min for an initial denaturation, followed by 35 cycles of denaturation for 30 s at 95 °C, primer annealing for 30 s at temperature described in Table 2, primer extension for 1 min at 72 °C, and a final extension for 7 min at 72 °C. All reactions were performed on GeneAmp PCR System 2700 Thermocycler (Applied Biosystems, Waltham, MA, USA). PCR primers, product size and sources are described in Table 2. The PCR products were visualized in 2% agarose gels containing Ethidium Bromide (Invitrogen, Waltham, MA, USA) under UV radiation. PCR followed thermal cycling according to Matsui et al. (2021) [30].
5. Conclusions
Our research showed that Salmonella enterica subsp. diarizonae does not cause pathological changes in the internal organs of mallard ducks (Anas platyrhynchos), so there is a very high probability of transmitting the pathogen over long distances in the intestinal trackt. On the other hand, studies have shown very serious difficulties in the routine diagnosis of Salmonella enterica subsp. diarizonae. The use of standard biochemical tests (such as API20E) and commercially available sera carries a high risk of confusing the strain with closely related Salmonella enterica subsp. arizonae. The fact of the occurrence of multidrug resistance in the studied strain is very disturbing.
The results of this study suggest that wild waterfowl may serve as a significant vector of antibiotic-resistant bacteria and may have a crucial impact on public health. To our knowledge, this is the first case of detection of S. enterica subsp. diarizonae in wild duck and its biochemical and antibiotic resistance pattern analysis. A better understanding of the factors that potentially contribute to the spreading of resistant zoonotic pathogens can improve the tools used by public health specialists to control antimicrobial resistance. More studies with larger numbers of samples, and DNA sequencing of bacterial strains, are needed to prove the hypothesis about the exact routes of transmission of antibiotic-resistance genes and S. enterica subsp. diarizonae itself. The prevalence of migrated mallard ducks around the world indicates that they may be a vector that transfers antimicrobial resistance genes from countries with high antibiotic use to countries with programs to reduce the use of chemotherapeutic agents. This can have serious implications for the health of wildlife, livestock, and humans.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/antibiotics11040530/s1, Table S1: a summary of the results of biochemical reactions for the isolated strain of Salmonella spp. in the VITEK test, API-20E, and individual biochemical lab-made tests.
Author Contributions
Conceptualization, J.P.-C. and K.A.; methodology, J.P.-C. and K.W.; Methodology—biochemical and serological typing and molecular typing, J.P.-C., K.W. and E.T.-P.; data curation, J.P.-C., K.W., E.T.-P. and L.P.; software, J.P.-C., K.W. and P.K.; validation, J.P.-C., K.W. and A.K.; formal analysis, J.P.-C., K.W. and Z.B.; investigation, J.P.-C., K.W. and Z.B.; resources, J.B., J.W., P.K. and Z.B.; data curation, P.K. and J.B.; writing—original draft preparation, J.P.-C., K.W., J.B., J.W., P.K., E.T.-P. and L.P.; writing—review and editing, A.K. and K.A.; visualization, K.W. and J.W.; supervision, J.P.-C. and K.W., project administration, J.P.-C.; funding acquisition, K.A., A.K. and Z.B. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
The study complied with Directive 2010/63/EU and the Act of the Polish Parliament dated 15 January 2015 on the protection of animals used for scientific purposes (Journal of Laws 2015, item 266). The wild ducks were not killed for the purposes of the study. The hunt took place in accordance with Polish hunting law (Act of the Polish Parliament dated 13 October 1995, item 713, the Hunting law, Chapter 3, Art. 8 Hunt) during the hunting season 2021.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available in Supplementary Materials.
Acknowledgments
Special thanks to Jolanta Przybylska for help with the laboratory work. Special thanks to Przemysław Walerjan for help with editing.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
References
- Liu, H.; Whitehouse, C.A.; Li, B. Presence and Persistence of Salmonella in Water: The Impact on Microbial Quality of Water and Food Safety. Front. Public Health 2018, 6, 159. [Google Scholar] [CrossRef] [PubMed]
- EFSA (European Food Safety Authority) and ECDC (European Centre for Disease Prevention and Control). The European Union Summary Report on Antimicrobial Resistance in zoonotic and indicator bacteria from humans, animals and food in 2018/2019. EFSA J. 2021, 19, e06490. [Google Scholar] [CrossRef]
- Giner-Lamia, J.; Vinuesa, P.; Betancor, L.; Silva, C.; Bisio, J.; Soleto, L.; Chabalgoity, J.A.; Puente, J.L.; Soncini, F.C.; García-Vescovi, E.; et al. Genome analysis of Salmonella enterica subsp. diarizonae isolates from invasive human infections reveals enrichment of virulence-related functions in lineage ST1256. BMC Genom. 2019, 20, 99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schröter, M.; Roggentin, P.; Hofmann, J.; Speicher, A.; Laufs, R.; Mack, D. Pet Snakes as a Reservoir for Salmonella enterica subsp. diarizonae (Serogroup IIIb): A Prospective Study. Appl. Environ. Microbiol. 2004, 70, 613–615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bjelland, A.M.; Sandvik, L.M.; Skarstein, M.M.; Svendal, L.; Debenham, J.J. Prevalence of Salmonella serovars isolated from reptiles in Norwegian zoos. Acta Vet. Scand. 2020, 62, 3. [Google Scholar] [CrossRef]
- Alvseike, O.; Skjerve, E. Prevalence of a Salmonella subspecies diarizonae in Norwegian sheep herds. Prev. Vet. Med. 2002, 52, 277–285. [Google Scholar] [CrossRef]
- Stokar-Regenscheit, N.; Overesch, G.; Giezendanner, R.; Roos, S.; Gurtner, C. Salmonella enterica subsp. diarizonae serotype 61:k:1,5,(7) associated with chronic proliferative rhinitis and high nasal colonization rates in a flock of Texel sheep in Switzerland. Prev. Vet. Med. 2017, 145, 78–82. [Google Scholar] [CrossRef]
- Chatzopoulos, D.C.; Sarrou, S.; Vasileiou, N.G.C.; Ioannidi, K.S.; Peteinaki, E.; Valiakos, G.; Tsokana, C.N.; Papadopoulos, E.; Spyrou, V.; Mavrogianni, V.S.; et al. Dissemination of intestinal pathogens between lambs and puppies in sheep farms. Small Rumin. Res. 2016, 141, 5–10. [Google Scholar] [CrossRef]
- Chatzopoulos, D.C.; Vasileiou, N.G.C.; Ioannidi, K.S.; Katsafadou, A.I.; Mavrogianni, V.S.; Michael, C.K.; Katsarou, E.I.; Karavanis, E.; Papadopoulos, N.; Sbiraki, A.; et al. Experimental study of the potential role of Salmonella enterica subsp. Diarizonae in the diarrhoeic syndrome of lambs. Pathogens 2021, 10, 113. [Google Scholar] [CrossRef]
- Gawin, P. Wild game in Poland. In Hunting Traditions, Law, Game; Gawin, P., Ed.; SBM Sp. z o.o.: Warsaw, Poland, 2015; pp. 129–131. ISBN 9788378458661. [Google Scholar]
- David Black Modelling the Potential Spatiotemporal Spread of Avian Influenza. 2006. Available online: https://www.geos.ed.ac.uk/~mscgis/05-06/s0566972/index.html#Introduction (accessed on 1 June 2021).
- Redlisiak, M.; Wardecki, Ł.; Karpińska, O.; Hayatli, F.; Grzębkowski, M.K.K. Report on the project of ringing of the Mallards Anas platyrhynchos in the Warsaw Metropolitan Area in 2015–2018: Sprawozdanie z projektu znakowania krzy ż ówki Anas platyr. Kulon 2018, 23, 177–243. [Google Scholar]
- Van Toor, M.L.; Hedenström, A.; Waldenström, J.; Fiedler, W.; Holland, R.A.; Thorup, K.; Wikelski, M. Flexibility of Continental Navigation and Migration in European Mallards. PLoS ONE 2013, 8, e72629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lillehaug, A.; Jonassen, C.M.; Bergsjø, B.; Hofshagen, M.; Tharaldsen, J.; Nesse, L.L.; Handeland, K. Screening of feral pigeon (Colomba livia), mallard (Anas platyrhynchos) and graylag goose (Anser anser) populations for Campylobacter spp., Salmonella spp., avian influenza virus and avian paramyxovirus. Acta Vet. Scand. 2005, 46, 193–202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dera-Tomaszewska, B. Epidemiology and Pathology of Salmonella Enteritidis and Salmonella Serovars First Isolated in Poland; Medical University of Gdańsk: Gdańsk, Poland, 2013; Volume 43. [Google Scholar]
- Wattiau, P.; Boland, C.; Bertrand, S. Methodologies for Salmonella enterica subsp. Enterica Subtyping: Gold Standards and Alternatives. Appl. Environ. Microbiol. 2011, 77, 7877–7885. [Google Scholar] [CrossRef] [Green Version]
- Pintado, V. Old and new antibiotics for therapy of multidrug resistant bacteria/Fármacos antiguos y nuevos en el tratamiento de la infección por bacterias multirresistentes. Rev. Esp Quim. 2016, 29, 39–42. [Google Scholar]
- Goodlet, K.J.; Benhalima, F.Z.; Nailor, M.D. A systematic review of single-dose aminoglycoside therapy for urinary tract infection: Is it time to resurrect an old strategy? Antimicrob. Agents Chemother. 2019, 63, e02165-18. [Google Scholar] [CrossRef] [Green Version]
- Kapoor, G.; Saigal, S.A.E. Action and resistance mechanisms of antibiotics: A guide for clinicians. J. Anaesthesiol. Clin. Pharmacol. 2017, 33, 300–305. [Google Scholar] [CrossRef]
- Gebreyes, W.A.; Altier, C. Molecular characterization of multidrug-resistant Salmonella enterica subsp. enterica serovar Typhimurium isolates from swine. J. Clin. Microbiol. 2002, 40, 2813–2822. [Google Scholar] [CrossRef] [Green Version]
- Regulation of the Minister of the Environment of 11 March 2005 on the List of Game Species Dz.U. 2005 nr 45 poz. 433. 2005. Available online: https://isap.sejm.gov.pl/isap.nsf/DocDetails.xsp?id=WDU20050450433 (accessed on 1 June 2021).
- EN ISO 6579-1:2017; Microbiology of the Food Chain—Horizontal Method for the Detection, Enumeration and Serotyping of Salmonella—Part 1: Detection of Salmonella spp. ISO: Geneva, Switzerland, 2017.
- Pławińska-Czarnak, J.; Wódz, K.; Kizerwetter-Świda, M.; Nowak, T.; Bogdan, J.; Kwieciński, P.; Kwieciński, A.; Anusz, K. Citrobacter braakii yield false-positive identification as salmonella, a note of caution. Foods 2021, 10, 2177. [Google Scholar] [CrossRef]
- Załęska, H.; Teisseyre, T.; Janczura, E. Media and nutrients for microorganisms of the Enterobacteriaceae family. In Bacteriological Media. Composition and Preparation; Załęska, H., Ed.; National Institute of Hygiene, Department of Bacteriology, Methodological Publishers of the National Institute of Hygiene: Warsaw, Poland, 2015; Volume 5, pp. 82–131. ISBN 978-83-7845-866-1. [Google Scholar]
- Issenhuth-Jeanjean, S.; Roggentin, P.; Mikoleit, M.; De Pinna, E.; Nair, S.; Fields, P.I.; Weill, F.X. Supplement 2008–2010 (no. 48) to the White-Kauffmann-Le Minor Scheme. Res. Microbiol. 2014, 165, 526–530. [Google Scholar] [CrossRef] [Green Version]
- Grimont, P.A.D.; Weill, F.-X. Antigenic Formulae of the Salmonella Serovars, 9th ed.; WHO Collaborating Centre for Reference and Research on Salmonella: Paris, France, 2007; pp. 1–167. [Google Scholar]
- CLSI. Performance Standards for Antimicrobial Susceptibility Testing, 28th ed.; CLSI Supplement M100; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2018. [Google Scholar]
- Ramtahal, M.A.; Somboro, A.M.; Amoako, D.G.; Abia, A.L.K.; Perrett, K.; Bester, L.A.; Essack, S.Y. Molecular Epidemiology of Salmonella enterica in Poultry in South Africa Using the Farm-to-Fork Approach. Int. J. Microbiol. 2022, 2022, 5121273. [Google Scholar] [CrossRef]
- Kozak, G.K.; Boerlin, P.; Janecko, N.; Reid-Smith, R.J.; Jardine, C. Antimicrobial resistance in Escherichia coli isolates from Swine and wild small mammals in the proximity of swine farms and in natural environments in Ontario, Canada. Appl. Environ. Microbiol. 2009, 75, 559–566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsui, K.; Nakazawa, C.; Khin, S.T.M.M.; Iwabuchi, E.; Asai, T.; Ishihara, K. Molecular characteristics and antimicrobial resistance of Salmonella enterica serovar schwarzengrund from chicken meat in japan. Antibiotics 2021, 10, 1336. [Google Scholar] [CrossRef] [PubMed]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).