Abstract
Carbapenem-resistant Enterobacterales (CRE) is a critical public health problem in South America, where the prevalence of NDM metallo-betalactamases has increased substantially in recent years. In this study, we used whole genome sequencing to characterize a multidrug-resistant (MDR) Klebsiella pneumoniae (UCO-361 strain) clinical isolate from a teaching hospital in Chile. Using long-read (Nanopore) and short-read (Illumina) sequence data, we identified a novel un-typeable megaplasmid (314,976 kb, pNDM-1_UCO-361) carrying the blaNDM-1 carbapenem resistance gene within a Tn3000 transposon. Strikingly, conjugal transfer of pNDM-1_UCO-361 plasmid only occurs at low temperatures with a high frequency of 4.3 × 10−6 transconjugants/receptors at 27 °C. UCO-361 belonged to the ST1588 clone, previously identified in Latin America, and harbored aminoglycoside, extended-spectrum β-lactamases (ESBLs), carbapenem, and quinolone-resistance determinants. These findings suggest that blaNDM-1-bearing megaplasmids can be adapted to carriage by some K. pneumoniae lineages, whereas its conjugation at low temperatures could contribute to rapid dissemination at the human–environmental interface.
1. Introduction
Carbapenem-resistant Enterobacterales (CRE) is a global concern for public health due to the limited therapeutic options to treat serious infections caused by these microorganisms [1], which are also associated with higher economic costs. For this reason, the World Health Organization (WHO) listed CRE and ESBL-producing Enterobacterales as top critical-priority pathogens for which new therapeutic options and surveillance are urgently needed [2].
Carbapenem-resistance is normally mediated by the production of carbapenemases, which are commonly associated with highly prevalent international clonal groups [3]. Among carbapenemases, New Delhi metallo-β-lactamases (NDM) have been identified worldwide, and their prevalence has increased substantially in recent years in South America [4,5]. Worryingly, infections produced by NDM-producers are associated with high mortality rates, representing an important challenge for public health [5]. Among NDM carbapenemases, NDM-1 has been identified in several Gram-negative pathogens, including Enterobacterales and non-fermenting rods recovered in different geographical areas, from human and animal hosts and environmental sources [5,6].
Due to the emergence of carbapenem-resistant bacterial pathogens mediated by NDM-enzymes, Wu et al. published a comprehensive review on NDM enzymes that included the epidemiological data of NDM-positive Enterobacterales deposited in GenBank in 2018. Accordingly, the authors demonstrated that most countries from Latin America (LA) showed a prevalence of ≤5%, whereas the authors did not find data in some countries, such as Chile and Bolivia [5]. In this sense, the Reference Laboratory of Healthcare-Associated Infections (HAIs) of the Instituto de Salud Pública de Chile (ISP, Chile) confirmed the first isolation of the K. pneumoniae producer of NDM-1 in Chile in May 2014 (https://www.ispch.cl/sites/default/files/BoletinRam-30112015A_0.pdf, accessed on 5 March 2022). Importantly, we previously determined that this isolate, named UCO-361, was genetically related to an NDM-producer K. pneumoniae collected in Brazil, revealing the importance of genomic surveillance [7]. After their first identification, 84 NDM-producing Gram-negative isolates were reported during the 2014–2017 period in Chile (https://www.ispch.cl/sites/default/files/BoletinCarbapenemasas-02042019A%20(1).pdf, accessed on 5 March 2022), which demonstrates that these enzymes are circulating in the country at an unknown frequency. Because of this phenomenon, it is highly important to determine the role of plasmids, such as megaplasmids, in the rapid dissemination of carbapenemases. In this sense, blaNDM gene is contained in plasmids of different replicon types, where IncX3 and IncC (IncA/C2) have been identified as the most prevalent type of plasmids carrying these genes in Enterobacterales worldwide [5,8]. Recently, diverse megaplasmids have been associated with the dissemination of multidrug resistance in clinically relevant pathogens [9,10,11,12]. Megaplasmids are referred to large plasmids of more than 100-kp that are characterized by their mosaic structure containing regions from diverse origins, including metals- and/or antibiotic-resistance genes (ARGs) [12,13]. In this study, we used a genomic approach to characterize a carbapenem-resistant Klebsiella pneumoniae (UCO-361 strain) clinical isolate carrying a novel transferable NDM-1-encoding megaplasmid.
2. Materials and Methods
A carbapenem-resistant K. pneumoniae strain, UCO-361, that corresponds to the first NDM-1-carrying strain detected in Chile, was analyzed. UCO-361 was recovered from a rectal swab of a colonized inpatient in a public hospital in Santiago (Capital of Chile) and is genetically related to a Brazilian clone [7]. This isolate was recovered from a hospitalized patient that shared a room with a colonized patient who was previously hospitalized in Brazil [7]. Moreover, Escherichia coli J53 (sodium-azide-resistant strain, SAZR) was used as a recipient strain for mating experiments. E. coli ATCC 25922 and Serratia marcescens UCO-143 (blaIMP+) strains were used as controls for susceptibility studies and as a positive control for carbapenemase production in phenotypic tests, respectively. The strains Acinetobacter baumannii UCO-323 and A. baumannii UCO-324, which were previously characterized in our laboratory by the detection of blaNDM-1 by conventional PCR and Sanger sequencing, were used as positive controls for the blaNDM-1 gene.
Susceptibility studies were carried out by disk diffusion and broth dilution methods, according to the recommendations of Clinical & Laboratory Standards Institute (CLSI) [14]. The following antimicrobial disks (Oxoid Ltd., Basingstoke, UK) were included: imipenem (IPM, 10 μg), meropenem (MEM, 10 μg), ertapenem (ETP, 10 μg), cefotaxime (CTX, 30 μg), ceftazidime (CAZ, 30 μg), cefepime (FEP, 30 μg), amoxicillin/clavulanate (AMC, 20/10 μg), piperacillin/tazobactam (TZP, 100/10 μg), ceftazidime/avibactam (30/20 μg), aztreonam (ATM, 30 μg), amikacin (AMK, 30 μg), gentamicin (GEN, 10 μg), levofloxacin (LEV, 5 μg), ciprofloxacin (CIP, 5 μg), tetracycline (TET, 30 μg) and sulfamethoxazole/trimethoprim (SXT, 23.75/1.25 μg). Moreover, susceptibility to colistin (CST) was determined by the colistin-agar test (CLSI, 2021). Additionally, Carba NP method was used to test the carbapenemase activity [14], whereas metallo-betalactamase (MβL) production was investigated by the imipenem-EDTA disk assay [15].
Initial detection of blaNDM-1 was carried out by conventional PCR, using primers and conditions described by Poirel et al., 2011 [16]. Next, UCO-361 was subjected to whole genome sequencing (WGS) through short- (NextSeq, Illumina Paired-end platform) and long-reads (MinION; Oxford Nanopore Technologies). For the transconjugant strain (E. coli Tc-01), short-read (Illumina) sequencing was utilized to confirm the antibiotic-resistance genes (ARGs) transferred after mating assays. De novo assemblies were achieved using Unicycler v0.4.8 and SPades v3.15.4 assemblers, depending on the sequencing platforms utilized [17]. The resulting contigs were visualized with Bandage v0.8.1, whereas genomic annotation was performed using Prokka [18].
From the assembled genome, multilocus sequence type (MLST), plasmid replicons, antimicrobial resistance genes, virulence genes, capsule (K) and lipopolysaccharide (O) loci were identified using the platforms MLSTFinder 2.0 (https://cge.food.dtu.dk/services/MLST/, accessed in 5 March 2022), PlasmidFinder 2.1 (https://cge.food.dtu.dk/services/PlasmidFinder/, accessed in 5 March 2022), ResFinder 4.1 (https://cge.food.dtu.dk/services/ResFinder/, accessed in 5 March 2022), and PathogenWatch (https://pathogen.watch/, accessed in 5 March 2022), respectively. Assembly, annotation, and visualization of plasmids were carried out by Proksee (https://proksee.ca/, accessed in 5 March 2022). Plasmid sequences were compared against the PLSDB plasmid database [19].
Conjugation assays were conducted as described by Potron et al. [20]. For transconjugant selection, we utilized tryptic soy agar (Oxoid Ltd., Basingstoke, UK) plates containing SAZ (300 μg/mL) and supplemented with different antibiotics combinations per plate: SAZ-AMP (16 μg/mL), SAZ-ETP (0.125 μg/mL), SAZ-AMK (4 μg/mL) and SAZ-CTX (4 μg/mL). Mating experiments were carried out at 27 and 37 °C, and conjugation frequency (R) was estimated as R = number of transconjugants/number of recipients [21]. To verify the isogenicity between the recipient and transconjugant strains, they were subjected to Enterobacterial Repetitive Intergenic Consensus PCR (ERIC-PCR) according to Aydin et al. [22].
3. Results and Discussion
UCO-361 was isolated from a rectal swab sample from a colonized inpatient. It displayed resistance to IPM, MEM, ETP, CTX, CAZ, FEP, AMC, TZP, CZA, ATM, GEN, TET, SXT and CIP, intermediate to LEV and susceptible to AMK and CST (MIC < 3.5 µg/mL). According to the definition proposed by Magiorakos et al. [23], UCO-361 corresponds to an extensively drug-resistant (XDR) strain. Moreover, Carba NP and imipenem-EDTA tests were positive for the donor (UCO-361) and transconjugant (Tc-01) strains, evidencing the presence of a transferable MβL. Moreover, PCR results revealed the presence of blaNDM in UCO-361.
To characterize the genetic features of UCO-361, we performed a hybrid analysis using short-read (Illumina, San Diego, CA, USA) and long-read (Oxford Nanopore Technologies, Oxford, UK) sequencing platforms. As a result, MLSTFinder analysis revealed that UCO-361 belonged to the ST1588, which has been previously identified in 3 hospitals in Rio de Janeiro, Brazil [24]. Moreover, UCO-361 presented the capsular serotype KL108/O1 that has been associated with hypervirulent clones [25]. Specifically, multidrug-resistant and hypervirulent strains associated with KL108 have been detected in different regions [25]. Furthermore, virulence genes associated with the hyperproduction of capsule (hypermucoid phenotype), such as rmpADC and rmpA2 [25], were not detected in UCO-361.
On the other hand, ResFinder was used to detect the ARGs contained in UCO-361, which encompassed several β-lactamases, including ESBLs (blaCTX-M-15 and blaSHV-106) and carbapenemase (blaNDM-1), aminoglycosides-modifying enzymes (aph(3″)-lb, aac(3)-lla, aph(6)-ld, aac(6′)lb-cr), and quinolones- and trimethoprim/sulfamethoxazole-resistance genes (Table 1). Moreover, blaNDM-1 was detected in Tc-01 in addition to other β-lactamases, aminoglycosides-modifying enzymes, and quinolones-resistance genes (Table 1).

Table 1.
Susceptibility profile and resistome of K. pneumoniae UCO-361 and transconjugant E. coli Tc-01 strains.
From the hybrid-sequencing approach, we identified a megaplasmid of 314,976 bp in UCO-361, named pNDM-1_UCO361, that carries the blaNDM-1 and oqxB genes, as the ARG detected (Figure 1A). Furthermore, we determined that blaNDM-1 was in the Tn3000 transposon that includes a copy of IS3000 and truncated ΔISAba125 upstream blaNDM-1, whereas the bleomycin-resistance gene bleMBL, in addition to trpF, dsdD, ΔgroES, groEL and a copy of IS3000, were located downstream (Figure 1B). In this sense, blaNDM genes are frequently described surrounded by the insertion sequence (IS) ISAba125 upstream bleMBL downstream [5,8], which is congruent with our results. Interestingly, blaNDM-1 has been previously detected in a plasmid within the transposon Tn3000 [24], remarking the relevance of this transposon in the spread of NDM-1.
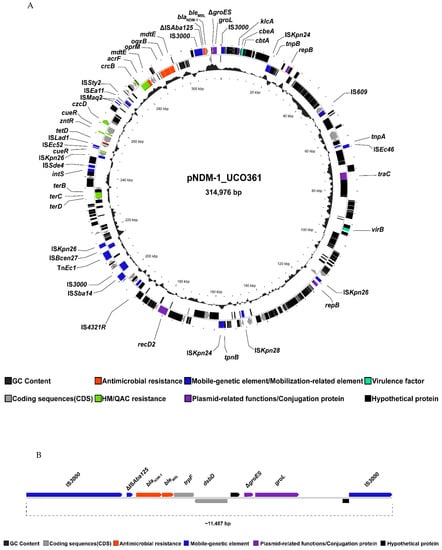
Figure 1.
Circular representation of plasmid harboring blaNDM-1 in K. pneumoniae UCO-361. (A) General representation of genes encoded by pNDM-1_UCO361 showing coding sequences (CDS) in black, mobile-genetic elements in green, antibiotic-resistance genes in red and plasmid replication/conjugation proteins in blue. The genetic context of blaNDM-1 is highlighted inside the red box. (B) Linear view of Tn3000 containing the blaNDM-1 gene. Figures were created with Proksee (Proksee.ca).
Remarkably, pNDM-1_UCO361 does not match with any Inc group deposited in the PlasmidFinder database. Moreover, comparisons of pNDM-1_UCO361 using the PLSDB platform (https://ccb-microbe.cs.uni-saarland.de/plsdb/, accessed on 5 March 2022) showed no matches with complete plasmid sequences retrieved from the NCBI nucleotide database. Furthermore, using the mash dist function of PLSDB, we determined that the closest plasmid related to pNDM-1_UCO361 corresponded to the pNDM-1-EC12 plasmid isolated in Enterobacter cloacae strain EC12 (accession number NZ_MN598004.1), in which blaNDM-1 was identified in a common region of 2488 bp. Moreover, blaNDM-1 has been previously identified in a megaplasmid (ca. 350 kb) in Raoultella ornithinicola collected from water samples in rural China [11]. This plasmid (accession number CP041388) contained the blaNDM-1 in a different genetic environment in comparison to pNDM-1_UCO361 since it was bracketed by two copies of IS91, conforming to an unusual integron class 1 [11].
In terms of conjugation-related proteins, pNDM-1_UCO361 contained the traC gene (Figure 1A), which encodes for a type-IV secretion system involved in the assembly of the F conjugative pilus [26]. Furthermore, pNDM-1_UCO361 presented several mobile-genetic elements, such as insertion sequences (IS) (Figure 1A), which remarks its potential to acquire exogenous genes. In addition to traC, pNDM-1_UCO361 carries the hns gene that encodes for the H-NS protein, which regulates the expression of T4SS during plasmid conjugation [27].
Interestingly, although pNDM-1_UCO361 harbors the traC gene, we detected an additional plasmid of 197,209 bp, which belongs to the IncFIB(K) containing the complete tra locus, which is involved in plasmid conjugation [28]. Moreover, this IncFIB(K) plasmid does not carry any antibiotic-resistance gene; thus, we hypothesized that it might mediate the transfer of pNDM-1_UCO361 simultaneously with other plasmids carrying additional ARGs. In this sense, we were able to transfer the blaNDM-1 gene through the mating assay between K. pneumoniae UCO-361 and E. coli J53 only at 27 °C in both agar and broth media. Moreover, we determined that the conjugation frequency of the blaNDM-1 corresponded to 4.3 × 10−6 transconjugants/recipient cell. As a result, the transconjugant E. coli strain Tc-01 displayed the same ERIC-PCR banding pattern with strain J53 (recipient), demonstrating their isogenicity.
Phenotypically, Tc-01 exhibited resistance to IPM, MEM, ETP, CTX, CAZ, FEP, AMC, TZP, ATM, AMC, CZA, GEN, TET and non-susceptibility to CIP, LEV, AMK and CST. From WGS data, we determined that Tc-01 obtained several ARGs (Table 1), which explains the resulting multidrug-resistant resistant phenotype displayed by this strain. As expected, the blaSHV-1 gene was not transferred from UCO-361 due to its chromosomal location in K. pneumoniae [29]. These results suggest that additional genes and/or plasmids were co-transferred with pNDM-1_UCO361, conferring resistance to various antibiotics. Unlike previously published studies [30,31], both donor and transconjugant strains were resistant to CIP, which is congruent to the increasing prevalence of plasmid-borne quinolone-resistance genes reported previously in Chile [32] and is consistent with what was reported in Brazil [24]. Furthermore, we determined that the IncFIB(K) plasmid was not transferred after the conjugation assay since the replicon was not detected in the whole genome of Tc-01.
Importantly, conjugation was effective only at 27 °C, with frequencies higher than that previously reported at 25 °C [31,33]. The successful transfer of the blaNDM-1 gene at this temperature might be related to its environmental origin [5,34]. In this regard, temperature could play a fundamental role in the transfer of blaNDM-1, and even other resistance genes, due to the thermosensibility of the conjugation apparatus in some plasmid families. Accordingly, it has been established that the traG gene, which is present in the IncFIB(K) plasmid in UCO-361, encodes a binding protein between the relaxosome and the mating pair complex in the conjugation process. This gene is transcribed in a temperature-dependent manner, where its expression levels are considerably reduced at 37 °C [35]. Moreover, Gibert et al. demonstrated that the conjugation process R27 plasmid is thermoregulated, being promoted at 25 °C and suppressed at 37 °C [36]. However, the hns gene, which is present in pNDM-1_UCO361, has been described as inhibiting the conjugation of IncX3 plasmids at 30 and 42 °C and not at 37 °C [27]. Hence, we hypothesize that the IncFIB(K) plasmid present in UCO-361 plays a relevant role in the transfer of pNDM-1_UCO361, independently of the inhibition activity of H-NS. Therefore, it is important to elucidate the role of temperature in the regulation of proteins involved in the transfer of resistant megaplasmids since the likelihood of resistance gene transfer may be more frequent in the environment than in the human body.
In conclusion, it is important to evaluate the role of resistance plasmids that can contribute to the emergence of carbapenem-resistance Gram-negative bacteria. Although it is well known that the spread of NDM-1 occurs in the environment, it is necessary to understand the effect of temperature and plasmid interactions on this phenomenon.
Author Contributions
M.Q.-A., A.O.-C., F.E., B.F. and B.B. participated in the methodology and conceptualization of this work. A.O.-C., N.L. and G.G.-R. participated in the writing preparation. N.L., S.M.-M., G.R., C.A.L., H.B.-T., M.C., F.S.-O., J.C.H. and G.G.-R. contributed to the conceptualization of this work. A.O.-C., G.G.-R. and N.L. were awarded with grants to support this work. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by The National Fund for Scientific and Technological Development Grants FONDECYT N°1130838 and FONDECYT-Iniciación N°11190602, and by the ANID Millennium Science Initiative/Millennium Initiative for Collaborative Research on Bacterial Resistance, code NCN17_081.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
This Whole Genome Shotgun project has been deposited at DDBJ/ENA/GenBank under the accession JAMJQY000000000 (WGS) and JAMJQY010000002.1 (plasmid pNDM-1_UCO361). The version described in this paper is version JAMJQY010000000.
Acknowledgments
The authors acknowledge the microbiologists of the included hospitals who provided the isolate to the Programa de Vigilancia de Carbapenemasas en Enterobacterias, Instituto de Salud Pública de Chile (ISPCH). N.L. is a research fellow of CNPq (grant 314336/2021-4).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Paterson, D.L. Resistance in Gram-Negative Bacteria: Enterobacteriaceae. Am. J. Med. 2006, 119, S20–S28. [Google Scholar] [CrossRef] [PubMed]
- Tacconelli, E.; Carrara, E.; Savoldi, A.; Harbarth, S.; Mendelson, M.; Monnet, D.L.; Pulcini, C.; Kahlmeter, G.; Kluytmans, J.; Carmeli, Y.; et al. Discovery, Research, and Development of New Antibiotics: The WHO Priority List of Antibiotic-Resistant Bacteria and Tuberculosis. Lancet Infect. Dis. 2018, 18, 318–327. [Google Scholar] [CrossRef]
- Chen, L.; Mathema, B.; Chavda, K.D.; DeLeo, F.R.; Bonomo, R.A.; Kreiswirth, B.N. Carbapenemase-Producing Klebsiella pneumoniae: Molecular and Genetic Decoding. Trends Microbiol. 2014, 22, 686–696. [Google Scholar] [CrossRef]
- Safavi, M.; Bostanshirin, N.; Hajikhani, B.; Yaslianifard, S.; van Belkum, A.; Goudarzi, M.; Hashemi, A.; Darban-Sarokhalil, D.; Dadashi, M. Global Genotype Distribution of Human Clinical Isolates of New Delhi Metallo-β-Lactamase-Producing Klebsiella pneumoniae; A Systematic Review. J. Glob. Antimicrob. Resist. 2020, 23, 420–429. [Google Scholar] [CrossRef]
- Wu, W.; Feng, Y.; Tang, G.; Qiao, F.; Mcnally, A.; Zong, Z. NDM Metallo-Lactamases and Their Bacterial Producers in Health Care Settings. Clin. Microbiol. Rev. 2019, 32, e00115-18. [Google Scholar] [CrossRef]
- Zhai, Y.; Lee, S.; Teng, L.; Ma, Z.; Hilliard, N.B.; May, R.J.; Brown, S.A.; Yu, F.; Desear, K.E.; Cherabuddi, K.; et al. Dissemination Mechanisms of NDM Genes in Hospitalized Patients. JAC-Antimicrob. Resist. 2021, 3, dlab032. [Google Scholar] [CrossRef] [PubMed]
- Carrasco-Anabalón, S.; Conceição Neto, C.O.; D’Alincourt Carvalho-Assef, A.P.; Lima, C.A.; Cifuentes, M.; Silva, F.; Barrera, B.; Domínguez, M.; González-Rocha, G.; Bello-Toledo, H. Introduction of NDM-1 and OXA-370 from Brazil into Chile in Strains of Klebsiella pneumoniae Isolated from a Single Patient. Int. J. Infect. Dis. 2019, 81, 28–30. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.U.; Maryam, L.; Zarrilli, R. Structure, Genetics and Worldwide Spread of New Delhi Metallo-β-Lactamase (NDM): A Threat to Public Health. BMC Microbiol. 2017, 17, 101. [Google Scholar] [CrossRef]
- Yao, H.; Li, A.; Yu, R.; Schwarz, S.; Dong, H.; Du, X.D. Multiple Copies of Bla NDM-5 Located on Conjugative Megaplasmids from Porcine Escherichia coli Sequence Type 218 Isolates. Antimicrob. Agents Chemother. 2020, 64, e02134-19. [Google Scholar] [CrossRef]
- Li, M.; Guan, C.; Song, G.; Gao, X.; Yang, W.; Wang, T.; Zhang, Y. Characterization of a Conjugative Multidrug Resistance IncP-2 Megaplasmid, PPAG5, from a Clinical Pseudomonas aeruginosa Isolate. Microbiol. Spectr. 2022, 10, e0199221. [Google Scholar] [CrossRef]
- Zou, H.; Berglund, B.; Xu, H.; Chi, X.; Zhao, Q.; Zhou, Z.; Xia, H.; Li, X.; Zheng, B. Genetic Characterization and Virulence of a Carbapenem-Resistant Raoultella ornithinolytica Isolated from Well Water Carrying a Novel Megaplasmid Containing bla NDM-1. Environ. Pollut. 2020, 260, 114041. [Google Scholar] [CrossRef] [PubMed]
- Cazares, A.; Moore, M.P.; Hall, J.P.J.; Wright, L.L.; Grimes, M.; Emond-Rhéault, J.G.; Pongchaikul, P.; Santanirand, P.; Levesque, R.C.; Fothergill, J.L.; et al. A Megaplasmid Family Driving Dissemination of Multidrug Resistance in Pseudomonas. Nat. Commun. 2020, 11, 1370. [Google Scholar] [CrossRef] [PubMed]
- Ghaly, T.M.; Paulsen, I.T.; Sajjad, A.; Tetu, S.G.; Gillings, M.R. A Novel Family of Acinetobacter Mega-Plasmids Are Disseminating Multi-Drug Resistance Across the Globe While Acquiring Location-Specific Accessory Genes. Front. Microbiol. 2020, 11, 3058. [Google Scholar] [CrossRef] [PubMed]
- Clinical and Laboratory Standards Institute (CLSI). Performance Standards for Antimicrobial Susceptibility Testing, 31st ed.; CLSI: Wayne, PA, USA, 2021; ISBN 9781684401048. [Google Scholar]
- Lee, K.; Chong, Y.; Shin, H.B.; Kim, Y.A.; Yong, D.; Yum, J.H. Modified Hodge and EDTA-Disk Synergy Tests to Screen Metallo-Beta-Lactamase-Producing Strains of Pseudomonas and Acinetobacter Species. Clin. Microbiol. Infect. 2001, 7, 88–91. [Google Scholar] [CrossRef]
- Poirel, L.; Walsh, T.R.; Cuvillier, V.; Nordmann, P. Multiplex PCR for Detection of Acquired Carbapenemase Genes. Diagn. Microbiol. Infect. Dis. 2011, 70, 119–123. [Google Scholar] [CrossRef]
- Antonopoulos, D.A.; Assaf, R.; Aziz, R.K.; Brettin, T.; Bun, C.; Conrad, N.; Davis, J.J.; Dietrich, E.M.; Disz, T.; Gerdes, S.; et al. PATRIC as a Unique Resource for Studying Antimicrobial Resistance. Brief. Bioinform. 2019, 20, 1094–1102. [Google Scholar] [CrossRef]
- Seemann, T. Prokka: Rapid Prokaryotic Genome Annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef] [PubMed]
- Schmartz, G.P.; Hartung, A.; Hirsch, P.; Kern, F.; Fehlmann, T.; Müller, R.; Keller, A. PLSDB: Advancing a Comprehensive Database of Bacterial Plasmids. Nucleic Acids Res. 2022, 50, D273–D278. [Google Scholar] [CrossRef] [PubMed]
- Potron, A.; Poirel, L.; Rondinaud, E.; Nordmann, P. Intercontinental Spread of OXA-48 Beta-Lactamase-Producing Enterobacteriaceae over a 11-Year Period, 2001 to 2011. Euro Surveill. 2013, 18, 20549. [Google Scholar] [CrossRef]
- Rumi, M.V.; Mas, J.; Elena, A.; Cerdeira, L.; Muñoz, M.E.; Lincopan, N.; Gentilini, É.R.; di Conza, J.; Gutkind, G. Co-Occurrence of Clinically Relevant β-Lactamases and MCR-1 Encoding Genes in Escherichia coli from Companion Animals in Argentina. Vet. Microbiol. 2019, 230, 228–234. [Google Scholar] [CrossRef]
- Aydin, F.; Gümüşsoy, K.S.; Atabay, H.I.; Iça, T.; Abay, S. Prevalence and Distribution of Arcobacter Species in Various Sources in Turkey and Molecular Analysis of Isolated Strains by ERIC-PCR. J. Appl. Microbiol. 2007, 103, 27–35. [Google Scholar] [CrossRef] [PubMed]
- Magiorakos, A.P.; Srinivasan, A.; Carey, R.B.; Carmeli, Y.; Falagas, M.E.; Giske, C.G.; Harbarth, S.; Hindler, J.F.; Kahlmeter, G.; Olsson-Liljequist, B.; et al. Multidrug-Resistant, Extensively Drug-Resistant and Pandrug-Resistant Bacteria: An International Expert Proposal for Interim Standard Definitions for Acquired Resistance. Clin. Microbiol. Infect. 2012, 18, 268–281. [Google Scholar] [CrossRef]
- Aires, C.A.M.; Pereira, P.S.; de Araujo, C.F.M.; Chagas, T.P.G.; Oliveira, J.C.R.; Buonora, S.N.; Albano, R.M.; Carvalho-Assef, A.P.D.A.; Asensi, M.D. Multiclonal Expansion of Klebsiella pneumoniae Isolates Producing NDM-1 in Rio de Janeiro, Brazil. Antimicrob. Agents Chemother. 2017, 61, e01048-16. [Google Scholar] [CrossRef] [PubMed]
- Dong, N.; Yang, X.; Chan, E.W.C.; Zhang, R.; Chen, S. Klebsiella Species: Taxonomy, Hypervirulence and Multidrug Resistance. EBioMedicine 2022, 79, 103998. [Google Scholar] [CrossRef]
- Schandel, K.A.; Muller, M.M.; Webster, R.E. Localization of TraC, a Protein Involved in Assembly of the F Conjugative Pilus. J. Bacteriol. 1992, 174, 3800–3806. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Baomo, L.; Lili, S.; Moran, R.A.; van Schaik, W.; Chao, Z. Temperature-Regulated IncX3 Plasmid Characteristics and the Role of Plasmid-Encoded H-NS in Thermoregulation. Front. Microbiol. 2022, 1, 765492. [Google Scholar] [CrossRef]
- Bragagnolo, N.; Rodriguez, C.; Samari-Kermani, N.; Fours, A.; Korouzhdehi, M.; Lysenko, R.; Audette, G.F. Protein Dynamics in F-like Bacterial Conjugation. Biomedicines 2020, 8, 362. [Google Scholar] [CrossRef] [PubMed]
- Newire, E.A.; Ahmed, S.F.; House, B.; Valiente, E.; Pimentel, G. Detection of New SHV-12, SHV-5 and SHV-2a Variants of Extended Spectrum Beta-Lactamase in Klebsiella pneumoniae in Egypt. Ann. Clin. Microbiol. Antimicrob. 2013, 12, 16. [Google Scholar] [CrossRef]
- Kumarasamy, K.K.; Toleman, M.A.; Walsh, T.R.; Bagaria, J.; Butt, F.; Balakrishnan, R.; Chaudhary, U.; Doumith, M.; Giske, C.G.; Irfan, S.; et al. Emergence of a New Antibiotic Resistance Mechanism in India, Pakistan, and the UK: A Molecular, Biological, and Epidemiological Study. Lancet Infect. Dis. 2010, 10, 597–602. [Google Scholar] [CrossRef]
- Walsh, T.R.; Weeks, J.; Livermore, D.M.; Toleman, M.A. Dissemination of NDM-1 Positive Bacteria in the New Delhi Environment and Its Implications for Human Health: An Environmental Point Prevalence Study. Lancet Infect. Dis. 2011, 11, 355–362. [Google Scholar] [CrossRef]
- Elgorriaga-Islas, E.; Guggiana-Nilo, P.; Domínguez-Yévenes, M.; González-Rocha, G.; Mella-Montecinos, S.; Labarca-Labarca, J.; García-Cañete, P.; Bello-Toledo, H. Prevalence of Plasmid-Mediated Quinolone Resistance Determinant Aac(6′)-Ib-Cr among ESBL Producing Enterobacteria Isolates from Chilean Hospitals. Enferm. Infecc. Microbiol. Clin. 2012, 30, 466–468. [Google Scholar] [CrossRef] [PubMed]
- Potron, A.; Poirel, L.; Nordmann, P. Plasmid-Mediated Transfer of the Bla(NDM-1) Gene in Gram-Negative Rods. FEMS Microbiol. Lett. 2011, 324, 111–116. [Google Scholar] [CrossRef] [PubMed]
- Nordmann, P.; Poirel, L.; Walsh, T.R.; Livermore, D.M. The Emerging NDM Carbapenemases. Trends Microbiol. 2011, 19, 588–595. [Google Scholar] [CrossRef] [PubMed]
- Gunton, J.E.; Gilmour, M.W.; Alonso, G.; Taylor, D.E. Subcellular Localization and Functional Domains of the Coupling Protein, TraG, from IncHI1 Plasmid R27. Microbiology 2005, 151, 3549–3561. [Google Scholar] [CrossRef] [PubMed]
- Gibert, M.; Paytubi, S. Temperature Dependent Control of the R27 Conjugative Plasmid Genes. Front. Mol. Biosci. 2020, 7, 124. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).