
Soil Component: A Potential Factor Affecting the Occurrence and Spread of Antibiotic Resistance Genes
Abstract
:1. Introduction
2. The Occurrence and Spread of ARGs in Soil
2.1. The Pollution Status of ARB and ARGs in Soil
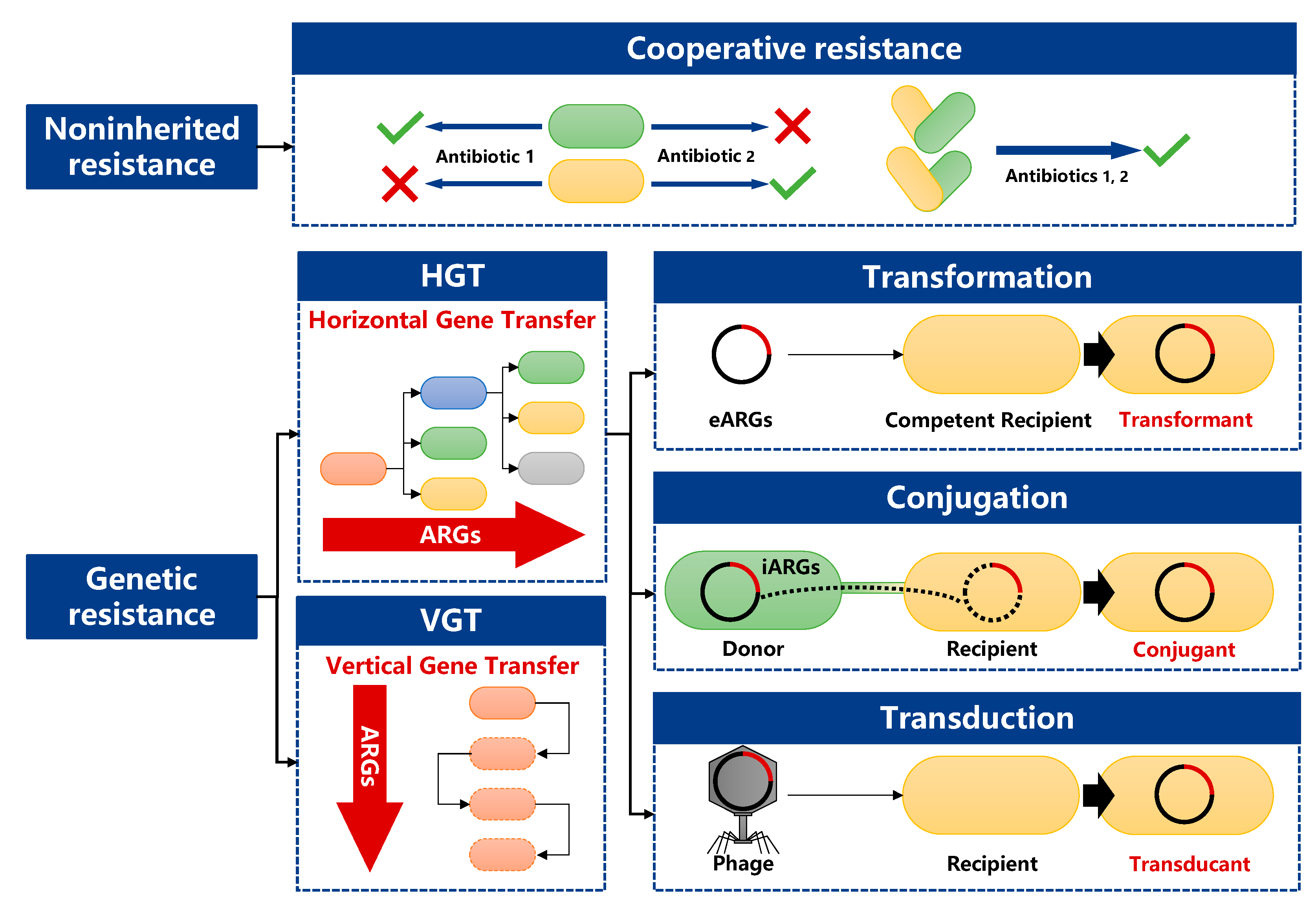
2.2. Transmission Routes of ARGs
3. Effects of Soil Types and Soil Components on the Occurrence of ARGs from the Macro Perspective
3.1. The Effects of Soil Types on the Occurrence of ARGs
3.2. The Control of Exogenously Added Soil Components on ARGs

{kind=link}
{kind=link}
| Soil Types | Research Process | Important Conclusions |
|---|---|---|
| Red, yellow, and black soils | Corpse decomposition [82] | Soil types have few impacts on ARGs; treatment, microbiome, NH4+ concentrates and pH are primary determinants of ARGs. |
| Loamy-sand, loam, and clay | Wastewater irrigation [8] | Soil type was the key factor in ARGs distribution; soil ARGs relative abundances were independent of the irrigation water quality |
| Fluvo-aquic, saline-alkali, and cinnamon soils | Long-term manure application [5] | Soil types influenced the ARGs distribution; higher diversity and abundance of ARGs occurred in fluvo-aquic and saline-alkali soil than cinnamon soil; Sand, pH and Zn contributed more to the pattern of ARGs in the cinnamon soils; sand and Cd, clay and Pb contributed the most in the fluvo-aquic and saline-alkali soils, respectively. |
| Acidic, near-neutral, and alkaline soils | Long-term manure application [6] | Soil types indirectly affected ARGs, while bacterial abundance and mobile genetic elements directly impacted ARG profiles; the effect of manure fertilization on the ARG profile in acidic and near-neutral soils was stronger than that in alkaline soil. |
| Humic acrisol, calcaric cambisols, and histosols | Interval fertilization [93] | Soil types affected ARGs. |
| Red soil, loess, and black soil | Fertilization [9] | The main contributor to the evolution of ARGs varied from soil types; no significant difference of antibiotic resistant bacteria and ARGs was observed among compost types. |
| Grassland soils | Long-term grazing [7] | ARGs shaped by the initial plant, soil environmental parameters (NO3−-N, TN, TP, pH) and microbiomes in grassland; long-term historic grazing had no effect on ARGs in grassland soils. |
| Sediments | Interannual variation [94] | TOC and clay were the major environmental factors regulating the variations in ARGs in sediments |
| Environmental Medium | Exogenously Added Soil Components | Important Conclusions | |
|---|---|---|---|
| Results | Reasons | ||
| Soil [87] | Biochar and pyroligneous acid | Both single and combined application of pyroligneous acid and biochar reduced the absolute abundance of ARGs in the rhizosphere and non-rhizosphere soils of leafy vegetables. | Pyroligneous acid and biochar reduced the bioavailability of heavy metal and improved soil properties. |
| Soil [10] | Biochar | Biochar impeded the vertical transport of ARGs. | Biochar addition enhanced dissolved organic matter export from soil, changed its composition. |
| Soil [92] | Biochar | Biochar amendment significantly decreased the abundance of ARGs in non-planted soil, but was not sufficient enough to alleviate ARGs level in planted-soil and plants. | Biochar was not sufficient enough to alleviate ARGs level; Increasing soil microbial diversity is more useful in mitigating ARG spread and accumulation. |
| Soil [91] | Biochar | Lolium multiflorum exhibited significantly stronger abatement of ARGs when combined with biochar than used alone; Soil pH and trace elements exerted weaker effects on ARGs after the application of biochar. | Lolium multiflorum and biochar improved soil physical structure, directly promoted the abatement of antibiotics and ARGs. |
| Soil and lettuce [88] | Biochar | Biochar reduced the relative abundance of ARGs in lettuce leaves, roots and soil. | The increased adsorption due to biochar and microbial degradation significantly alleviated environmental pressure; Bacteria were adsorbed, thus hindering their transport. |
| Soil and lettuce [89] | Biochar | Biochar can prevent soil antibiotics from accumulating in lettuce tissues; The enrichment of antibiotic resistant bacteria and the abundance of ARGs in lettuce was reduced by biochar treatment. | |
| Anaerobic digestion of swine manure [90] | Biochar | Biochar contributed to ARGs removal. | Biochar indirectly affected ARGs by changing intI1 and microbial structure. |
| Sludge composting [11] | Natural zeolite | Natural zeolite only controlled over some ARGs and had limited effect on bacterial community changes. | Porous structure of natural zeolite hindered microbial exposure and reduced heavy metal selection pressure. |
| Chicken manure composting [86] | Zeolite and biochar | Biochar and zeolite reduced the relative abundance of ARGs. | Biochar and zeolite had a suppressing effect on the abundance of intI1, and a reducing effect of horizontal gene transfer through conjugation and transformation. |
4. Effects of Soil Components on the HGT of ARGs from the Micro Perspective
4.1. The Effects of Soil Components on the HGT of ARGs in Pure Bacterial System
| Medium | Results | Reasons |
|---|---|---|
| Kaolinite, goethite, birnessite, and montmorillonite [13] | Birnessite promoted conjugation. The effects of kaolinite and montmorillonite were irregular. Goethite promoted conjugation at low concentration (0–0.5 g/L) and inhibited it at high concentration (5 g/L). | Birnessite promoted the production of intracellular reactive oxygen species (ROS); increased the expression levels of oxidative stress-regulated genes (rpoS) and outer membrane protein genes (ompA, ompF, ompC). Birnessite altered the expression levels of conjugation-related genes (globally regulation genes (korA, korB, trbA); mating pair formation (MPF) system genes (trbBp, traF); DNA transfer and replication (DTR) system genes (trfAp, traJ)). |
| Dissolved biochar [105] | The effects on conjugation were related to the concentration and source of biochar. | Humic acid-like substance in dissolved biochar improved the conjugative efficiency. The inhibitory effects of small-molecule matters dominated, decreasing conjugative transfer frequency. |
| Pyroligneous acid and its three fractions [103] | Reduced the abundance of ARGs and MGEs in soil. | High content of organic acids inhibited the bacterial growth. |
| Dissolved biochar [102] | Attenuated the promotion effect of Cu (Ⅱ) to conjugation. | Dissolved biochar affected intracellular ROS production level, cell membrane permeability, and the expression level of global regulatory genes (korA, korB, trbA), pore formation and membrane trafficking genes (ompA, ompC), MPF system gene (trbB), DTR system gene (trfA), etc. |
| CeO2 nanoparticle [104] (soil pollutant) | Inhibited conjugation at low concentration (1, 5 mg/L), while promoted it at high concentration (25, 50 mg/L). | CeO2 nanoparticle affected many aspects, such as intracellular ROS production, polysaccharide synthesis in EPS, cell-to-cell contact, ATP supply, and the expression level of conjugation-related genes (MPF system gene (trbBp), DTR system gene (trfAp), putative transmembrane ATPase gene (traG)), etc. |
| Gut of C. elegans [106] (soil animal) | The conjugation efficiency in gut was higher than soil, and increased with time and temperature. | The abundance of MPF system gene (trbBp) and DTR system gene (trfAp) was increased. |
4.2. Influence Mechanisms of Soil Components on HGT of ARGs
4.2.1. Intracellular Changes and Responses
Intracellular ROS Production
SOS Response
Cell Membrane Permeability
ATP Synthesis Capacity
Conjugation Activity of Intracellular Plasmids
4.2.2. Cell-Cell Contact and Quorum Sensing
4.2.3. Bacterial Uptake of Extracellular ARGs
The Competent State of Bacteria
Availability of Extracellular ARGs
4.2.4. Bacterial Concentration
5. Conclusions
6. Methods
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Van Boeckel, T.P.; Gandra, S.; Ashok, A.; Caudron, Q.; Grenfell, B.T.; Levin, S.A.; Laxminarayan, R. Global antibiotic consumption 2000 to 2010: An analysis of cross mark 742 national pharmaceutical sales data. Lancet Infect. Dis. 2014, 14, 742–750. [Google Scholar] [CrossRef]
- Sarmah, A.K.; Meyer, M.T.; Boxall, A.B.A. A global perspective on the use, sales, exposure pathways, occurrence, fate and effects of veterinary antibiotics (VAs) in the environment. Chemosphere 2006, 65, 725–759. [Google Scholar] [CrossRef]
- Jutkina, J.; Marathe, N.P.; Flach, C.F.; Larsson, D.G.J. Antibiotics and common antibacterial biocides stimulate horizontal transfer of resistance at low concentrations. Sci. Total Environ. 2018, 616, 172–178. [Google Scholar] [CrossRef]
- Li, B.; Qiu, Y.; Song, Y.Q.; Lin, H.; Yin, H. Dissecting horizontal and vertical gene transfer of antibiotic resistance plasmid in bacterial community using microfluidics. Environ. Int. 2019, 131, 105007. [Google Scholar] [CrossRef]
- Wang, L.J.; Wang, J.H.; Wang, J.; Zhu, L.S.; Conkle, J.L.; Yang, R. Soil types influence the characteristic of antibiotic resistance genes in greenhouse soil with long-term manure application. J. Hazard. Mater. 2020, 392, 11. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Cheng, D.M.; Zhang, Y.T.; Xie, J.; Xiong, H.; Wan, Y.; Zhang, Y.; Chen, X.; Shi, X. Soil type shapes the antibiotic resistome profiles of long-term manured soil. Sci. Total Environ. 2021, 786, 147361. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Z.Z.; Li, L.F.; Makhalanyane, T.P.; Xu, C.; Li, K.; Xue, K.; Xu, C.; Qian, R.; Zhang, B.; Du, J.; et al. The composition of antibiotic resistance genes is not affected by grazing but is determined by microorganisms in grassland soils. Sci. Total Environ. 2021, 761, 143205. [Google Scholar] [CrossRef]
- Seyoum, M.M.; Obayomi, O.; Bernstein, N.; Williams, C.F.; Gillor, O. Occurrence and distribution of antibiotics and corresponding antibiotic resistance genes in different soil types irrigated with treated wastewater. Sci. Total Environ. 2021, 782, 146835. [Google Scholar] [CrossRef]
- Zhang, J.Y.; Sui, Q.W.; Tong, J.; Zhong, H.; Wang, Y.W.; Chen, M.X.; Wei, Y.S. Soil types influence the fate of antibiotic-resistant bacteria and antibiotic resistance genes following the land application of sludge composts. Environ. Int. 2018, 118, 34–43. [Google Scholar] [CrossRef] [PubMed]
- Qiu, L.L.; Wu, J.J.; Qian, Y.; Nafees, M.; Zhang, J.; Du, W.; Yin, Y.; Guo, H. Impact of biochar-induced vertical mobilization of dissolved organic matter, sulfamethazine and antibiotic resistance genes variation in a soil-plant system. J. Hazard. Mater. 2021, 417, 126022. [Google Scholar] [CrossRef]
- Zhang, J.Y.; Chen, M.X.; Sui, Q.W.; Tong, J.; Jiang, C.; Lu, X.; Zhang, Y.; Wei, Y. Impacts of addition of natural zeolite or a nitrification inhibitor on antibiotic resistance genes during sludge composting. Water Res. 2016, 91, 339–349. [Google Scholar] [CrossRef] [PubMed]
- Demaneche, S.; Jocteur-Monrozier, L.; Quiquampoix, H.; Simonet, P. Evaluation of biological and physical protection against nuclease degradation of clay-bound plasmid DNA. Appl. Environ. Microbiol. 2001, 67, 293–299. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Wu, Y.C.; Huang, Q.Y.; Cai, P. Insights into conjugative transfer of antibiotic resistance genes affected by soil minerals. Eur. J. Soil Sci. 2020, 72, 1143–1153. [Google Scholar] [CrossRef]
- Nesme, J.; Simonet, P. The soil resistome: A critical review on antibiotic resistance origins, ecology and dissemination potential in telluric bacteria. Environ. Microbiol. 2015, 17, 913–930. [Google Scholar] [CrossRef]
- Ondon, B.S.; Li, S.N.; Zhou, Q.X.; Li, F. Sources of antibiotic resistant bacteria (ARB) and antibiotic resistance genes (ARGs) in the soil: A review of the spreading mechanism and human health risks. In Reviews of Environmental Contamination and Toxicology; de Voogt, P., Ed.; Springer International Publishing: Cham, Switzerland, 2021; Volume 256, pp. 121–153. [Google Scholar]
- Knapp, C.W.; McCluskey, S.M.; Singh, B.K.; Campbell, C.D.; Hudson, G.; Graham, D.W. Antibiotic resistance gene abundances correlate with metal and geochemical conditions in archived Scottish soils. PLoS ONE 2011, 6, e27300. [Google Scholar] [CrossRef]
- Knapp, C.W.; Zhang, W.; Sturm, B.S.M.; Graham, D.W. Differential fate of erythromycin and beta-lactam resistance genes from swine lagoon waste under different aquatic conditions. Environ. Pollut. 2010, 158, 1506–1512. [Google Scholar] [CrossRef]
- He, J.Z.; Yan, Z.Z.; Chen, Q.L. Transmission of antibiotic resistance genes in agroecosystems: An overview. Front. Agric. Sci. Eng. 2020, 7, 329–332. [Google Scholar] [CrossRef]
- D’Costa, V.M.; King, C.E.; Kalan, L.; Morar, M.; Sung, W.W.L.; Schwarz, C.; Froese, D.; Zazula, G.; Calmels, F.; Debruyne, R.; et al. Antibiotic resistance is ancient. Nature 2011, 477, 457–461. [Google Scholar] [CrossRef]
- Xu, H.; Chen, Z.Y.; Huang, R.Y.; Cui, Y.; Li, Q.; Zhao, Y.; Wang, X.; Mao, D.; Luo, Y.; Ren, H.Q. Antibiotic resistance gene-carrying plasmid spreads into the plant endophytic bacteria using soil bacteria as carriers. Environ. Sci. Technol. 2021, 55, 10462–10470. [Google Scholar] [CrossRef]
- Liu, J.X.; Zhao, Z.; Avillan, J.J.; Call, D.R.; Davis, M.; Sischo, W.M.; Zhang, A. Dairy farm soil presents distinct microbiota and varied prevalence of antibiotic resistance across housing areas. Environ. Pollut. 2019, 254, 113058. [Google Scholar] [CrossRef]
- Furlan, J.P.R.; Stehling, E.G. Multiple sequence types, virulence determinants and antimicrobial resistance genes in multidrug- and colistin-resistant Escherichia coli from agricultural and non-agricultural soils. Environ. Pollut. 2021, 288, 117804. [Google Scholar] [CrossRef]
- Graves, A.K.; Liwimbi, L.; Israel, D.W.; van Heugten, E.; Robinson, B.; Cahoon, C.W.; Lubbers, J.F. Distribution of ten antibiotic resistance genes in E. coli isolates from swine manure, lagoon effluent and soil collected from a lagoon waste application field. Folia Microbiol. 2011, 56, 131–137. [Google Scholar] [CrossRef] [PubMed]
- Knapp, C.W.; Dolfing, J.; Ehlert, P.A.I.; Graham, D.W. Evidence of increasing antibiotic resistance gene abundances in archived soils since 1940. Environ. Sci. Technol. 2010, 44, 580–587. [Google Scholar] [CrossRef]
- Zhang, Y.; Hu, X.; Wang, X. Recent advances on antibiotic resistance genes encoded by bacteriophages. China Environ. Sci. 2022, 42, 2315–2320. [Google Scholar]
- Wu, N.; Qiao, M.; Zhang, B.; Cheng, W.-D.; Zhu, Y.-G. Abundance and diversity of tetracycline resistance genes in soils adjacent to representative swine feedlots in China. Environ. Sci. Technol. 2010, 44, 6933–6939. [Google Scholar] [CrossRef] [PubMed]
- Ji, X.L.; Shen, Q.H.; Liu, F.; Ma, J.; Xu, G.; Wang, Y.; Wu, M. Antibiotic resistance gene abundances associated with antibiotics and heavy metals in animal manures and agricultural soils adjacent to feedlots in Shanghai; China. J. Hazard. Mater. 2012, 235, 178–185. [Google Scholar] [CrossRef]
- Wang, F.H.; Qiao, M.; Chen, Z.; Su, J.-Q.; Zhu, Y.-G. Antibiotic resistance genes in manure-amended soil and vegetables at harvest. J. Hazard. Mater. 2015, 299, 215–221. [Google Scholar] [CrossRef]
- Liu, Z.B.; Klümper, U.; Shi, L.; Ye, L.; Li, M. From pig breeding environment to subsequently produced pork: Comparative analysis of antibiotic resistance genes and bacterial community composition. Front. Microbiol. 2019, 10, 43. [Google Scholar] [CrossRef]
- Zhou, Y.; Niu, L.; Zhu, S.; Lu, H.; Liu, W. Occurrence, abundance, and distribution of sulfonamide and tetracycline resistance genes in agricultural soils across China. Sci. Total Environ. 2017, 599, 1977–1983. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Huang, W.; Lyu, X.; He, X.; Chen, L.; Song, Y. Effect of Long-term Biogas Slurry Fertilization on Distribution of Tetracycline and Sulfonamide Resistance Genes in Soil. Fujian J. Agric. Sci. 2021, 36, 699–705. [Google Scholar]
- Peng, S.; Wang, Y.; Lin, X. Abundance of the tetracycline resistance genes in a paddy soil after continuous application of composted swine manure for 6 years. China Environ. Sci. 2015, 35, 1173–1180. [Google Scholar]
- Chessa, L.; Jechalke, S.; Ding, G.C.; Pusino, A.; Mangia, N.P.; Smalla, K. The presence of tetracycline in cow manure changes the impact of repeated manure application on soil bacterial communities. Biol. Fertil. Soils 2016, 52, 1121–1134. [Google Scholar] [CrossRef]
- Anand, T.; Bera, B.C.; Vaid, R.K.; Barua, S.; Riyesh, T.; Virmani, N.; Hussain, M.; Singh, R.K.; Tripathi, B.N. Abundance of antibiotic resistance genes in environmental bacteriophages. J. Gen. Virol. 2016, 97, 3458–3466. [Google Scholar] [CrossRef] [PubMed]
- Munir, M.; Xagoraraki, I. Levels of antibiotic resistance genes in manure, biosolids, and fertilized soil. J. Environ. Qual. 2011, 40, 248–255. [Google Scholar] [CrossRef] [PubMed]
- Radu, E.; Woegerbauer, M.; Rab, G.; Oismüller, M.; Strauss, P.; Hufnagl, P.; Gottsberger, R.A.; Krampe, J.; Weyermair, K.; Kreuzinger, N. Resilience of agricultural soils to antibiotic resistance genes introduced by agricultural management practices. Sci. Total Environ. 2021, 756, 143699. [Google Scholar] [CrossRef]
- Lin, H.; Chapman, S.J.; Freitag, T.E.; Kyle, C.; Ma, J.; Yang, Y.; Zhang, Z. Fate of tetracycline and sulfonamide resistance genes in a grassland soil amended with different organic fertilizers. Ecotoxicol. Environ. Saf. 2019, 170, 39–46. [Google Scholar] [CrossRef]
- Knapp, C.W.; Callan, A.C.; Aitken, B.; Shearn, R.; Koenders, A.; Hinwood, A. Relationship between antibiotic resistance genes and metals in residential soil samples from western Australia. Environ. Sci. Pollut. Res. 2017, 24, 2484–2494. [Google Scholar] [CrossRef] [PubMed]
- Corona, F.; Martinez, J.L. Phenotypic Resistance to Antibiotics. Antibiotics 2013, 2, 237–255. [Google Scholar] [CrossRef]
- Frost, I.; Smith, W.P.J.; Mitri, S.; San Millan, A.; Davit, Y.; Osborne, J.M.; Pitt-Francis, J.M.; MacLean, R.C.; Foster, K.R. Cooperation, competition and antibiotic resistance in bacterial colonies. ISME J. 2018, 12, 1582–1593. [Google Scholar] [CrossRef]
- Sorg, R.A.; Lin, L.; van Doorn, G.S.; Sorg, M.; Olson, J.; Nizet, V.; Veening, J.-W. Collective resistance in microbial communities by intracellular antibiotic deactivation. PLoS Biol. 2016, 14, e2000631. [Google Scholar] [CrossRef]
- Brook, I. The role of beta-lactamase-producing-bacteria in mixed infections. BMC Infect. Dis. 2009, 9, 202. [Google Scholar] [CrossRef] [PubMed]
- Davies, J.; Davies, D. Origins and evolution of antibiotic resistance. Microbiol. Mol. Biol. Rev. 2010, 74, 417–433. [Google Scholar] [CrossRef] [PubMed]
- Perron, G.G.; Whyte, L.; Turnbaugh, P.J.; Goordial, J.; Hanage, W.P.; Dantas, G.; Desai, M.M. Functional characterization of bacteria isolated from ancient Arctic soil exposes diverse resistance mechanisms to modern antibiotics. PLoS ONE 2015, 10, e0069533. [Google Scholar] [CrossRef]
- Pontes, D.S.; Aquino de Araujo, R.S.; Dantas, N.; Scotti, L.; Scotti, M.T.; de Moura, R.O.; Bezerra Mendonca-Junior, F.J. Genetic mechanisms of antibiotic resistance and the role of antibiotic adjuvants. Curr. Top. Med. Chem. 2018, 18, 42–74. [Google Scholar] [CrossRef]
- von Wintersdorff, C.J.H.; Penders, J.; van Niekerk, J.M.; Mills, N.D.; Majumder, S.; van Alphen, L.B.; Savelkoul, P.H.M.; Wolffs, P.F.G. Dissemination of antimicrobial resistance in microbial ecosystems through horizontal gene transfer. Front. Microbiol. 2016, 7, 173. [Google Scholar] [CrossRef]
- Seoane, J.; Yankelevich, T.; Dechesne, A.; Merkey, B.; Sternberg, C.; Smets, B.F. An individual-based approach to explain plasmid invasion in bacterial populations. FEMS Microbiol. Ecol. 2011, 75, 17–27. [Google Scholar] [CrossRef] [PubMed]
- Meng, M.; Li, Y.; Yao, H. Plasmid-mediated transfer of antibiotic resistance genes in soil. Antibiotics 2022, 11, 525. [Google Scholar] [CrossRef]
- Virolle, C.; Goldlust, K.; Djermoun, S.; Bigot, S.; Lesterlin, C. Plasmid transfer by conjugation in gram-negative bacteria: From the cellular to the community Level. Genes 2020, 11, 1239. [Google Scholar] [CrossRef]
- Chen, M.-l.; An, X.-l.; Yang, K.; Zhu, Y.-g. Soil phage and their mediation on the horizontal transfer of antibiotic resistance genes: A review. Yingyong Shengtai Xuebao 2021, 32, 2267–2274. [Google Scholar] [CrossRef]
- Abe, K.; Nomura, N.; Suzuki, S. Biofilms: Hot spots of horizontal gene transfer (HGT) in aquatic environments, with a focus on a new HGT mechanism. FEMS Microbiol. Ecol. 2020, 96, fiaa031. [Google Scholar] [CrossRef]
- Getino, M.; Cruz, F.D.L.; Baquero, F.; Bouza, E.; Gutiérrez-Fuentes, J.A.; Coque, T.M. Natural and artificial strategies to control the conjugative transmission of plasmids. Microbiol. Spectr. 2018, 6, MTBP-0015-2016. [Google Scholar] [CrossRef] [PubMed]
- Thomas, C.M.; Nielsen, K.M. Mechanisms of, and barriers to, horizontal gene transfer between bacteria. Nat. Rev. Microbiol. 2005, 3, 711–721. [Google Scholar] [CrossRef] [PubMed]
- de Vries, J.; Wackernagel, W. Microbial horizontal gene transfer and the DNA release from transgenic crop plants. Plant Soil 2004, 266, 91–104. [Google Scholar] [CrossRef]
- Li, S.N.; Zhang, C.F.; Li, F.X.; Hua, T.; Zhou, Q.X.; Ho, S.H. Technologies towards antibiotic resistance genes (ARGs) removal from aquatic environment: A critical review. J. Hazard. Mater. 2021, 411, 125148. [Google Scholar] [CrossRef]
- Chen, P.; Chen, C.; Li, X. Transport of antibiotic resistance plasmids in porous media and the influence of surfactants. Front. Environ. Sci. Eng. 2018, 12, 5. [Google Scholar] [CrossRef]
- Ochman, H.; Lawrence, J.G.; Groisman, E.A. Lateral gene transfer and the nature of bacterial innovation. Nature 2000, 405, 299–304. [Google Scholar] [CrossRef]
- Partridge, S.R.; Kwong, S.M.; Firth, N.; Jensen, S.O. Mobile genetic elements associated with antimicrobial resistance. Clin. Microbiol. Rev. 2018, 31, e00088-00017. [Google Scholar] [CrossRef] [PubMed]
- Smillie, C.; Pilar Garcillan-Barcia, M.; Victoria Francia, M.; Rocha, E.P.C.; de la Cruz, F. Mobility of plasmids. Microbiol. Mol. Biol. Rev. 2010, 74, 434–452. [Google Scholar] [CrossRef] [PubMed]
- Waters, V.L. Conjugative transfer in the dissemination of beta-lactam and aminoglycoside resistance. Front. Biosci. 1999, 4, 433–456. [Google Scholar] [CrossRef]
- Norman, A.; Hansen, L.H.; Sorensen, S.J. Conjugative plasmids: Vessels of the communal gene pool. Philos. Trans. R. Soc. B-Biol. Sci. 2009, 364, 2275–2289. [Google Scholar] [CrossRef]
- Dionisio, F.; Conceicao, I.C.; Marques, A.C.R.; Fernandes, L.; Gordo, I. The evolution of a conjugative plasmid and its ability to increase bacterial fitness. Biol. Lett. 2005, 1, 250–252. [Google Scholar] [CrossRef] [PubMed]
- Hall, J.P.J.; Williams, D.; Paterson, S.; Harrison, E.; Brockhurst, M.A. Positive selection inhibits gene mobilization and transfer in soil bacterial communities. Nat. Ecol. Evol. 2017, 1, 1348–1353. [Google Scholar] [CrossRef] [PubMed]
- Clark, A.J.; Adelberg, E.A. Bacterial conjugation. Annu. Rev. Microbiol. 1962, 16, 289–319. [Google Scholar] [CrossRef]
- Llosa, M.; Gomis-Ruth, F.X.; Coll, M.; de la Cruz, F. Bacterial conjugation: A two-step mechanism for DNA transport. Plasmid 2002, 48, 243. [Google Scholar] [CrossRef] [PubMed]
- Fox, R.E.; Zhong, X.; Krone, S.M.; Top, E.M. Spatial structure and nutrients promote invasion of IncP-1 plasmids in bacterial populations. ISME J. 2008, 2, 1024–1039. [Google Scholar] [CrossRef]
- Nielsen, K.M.; van Elsas, J.D. Stimulatory effects of compounds present in the rhizosphere on natural transformation of Acinetobacter sp BD413 in soil. Soil Biol. Biochem. 2001, 33, 345–357. [Google Scholar] [CrossRef]
- Klumper, U.; Dechesne, A.; Riber, L.; Brandt, K.K.; Gulay, A.; Sorensen, S.J.; Smets, B.F. Metal stressors consistently modulate bacterial conjugal plasmid uptake potential in a phylogenetically conserved manner. Isme J. 2017, 11, 152–165. [Google Scholar] [CrossRef]
- Fan, X.-T.; Li, H.; Chen, Q.-L.; Zhang, Y.-S.; Ye, J.; Zhu, Y.-G.; Su, J.-Q. Fate of Antibiotic Resistant Pseudomonas putida and Broad Host Range Plasmid in Natural Soil Microcosms. Front. Microbiol. 2019, 10, 194. [Google Scholar] [CrossRef]
- Colombi, E.; Straub, C.; Kuenzel, S.; Templeton, M.D.; McCann, H.C.; Rainey, P.B. Evolution of copper resistance in the kiwifruit pathogen Pseudomonas syringae pv. actinidiae through acquisition of integrative conjugative elements and plasmids. Environ. Microbiol. 2017, 19, 819–832. [Google Scholar] [CrossRef]
- Goncalves, O.S.; Santana, M.F. The coexistence of monopartite integrative and conjugative elements in the genomes of Acidobacteria. Gene 2021, 777, 145476. [Google Scholar] [CrossRef]
- Furuya, E.Y.; Lowy, F.D. Antimicrobial-resistant bacteria in the community setting. Nat. Rev. Microbiol. 2006, 4, 36–45. [Google Scholar] [CrossRef]
- Muniesa, M.; Colomer-Lluch, M.; Jofre, J. Potential impact of environmental bacteriophages in spreading antibiotic resistance genes. Future Microbiol. 2013, 8, 739–751. [Google Scholar] [CrossRef]
- Pantasticocaldas, M.; Duncan, K.E.; Istock, C.A.; Bell, J.A. Population-Dynamics of Bacteriophage and Bacillus-Subtilis in Soil. Ecology 1992, 73, 1888–1902. [Google Scholar] [CrossRef]
- Liao, H.; Li, H.; Duan, C.S.; Zhou, X.Y.; An, X.L.; Zhu, Y.G.; Su, J.Q. Metagenomic and viromic analysis reveal the anthropogenic impacts on the plasmid and phage borne transferable resistome in soil. Environ. Int. 2022, 170, 107595. [Google Scholar] [CrossRef] [PubMed]
- Guemes, A.G.C.; Youle, M.; Cantu, V.A.; Felts, B.; Nulton, J.; Rohwer, F. Viruses as Winners in the Game of Life. Annu. Rev. Virol. 2016, 3, 197–214. [Google Scholar] [CrossRef]
- Wilhelm, S.W.; Weinbauer, M.G.; Suttle, C.A.; Jeffrey, W.H. The role of sunlight in the removal and repair of viruses in the sea. Limnol. Oceanogr. 1998, 43, 586–592. [Google Scholar] [CrossRef]
- Williamson, K.E.; Fuhrmann, J.J.; Wommack, K.E.; Radosevich, M. Viruses in soil ecosystems: An unknown quantity within an unexplored territory. Annu. Rev. Virol. 2017, 4, 201–219. [Google Scholar] [CrossRef]
- Kenzaka, T.; Tani, K.; Sakotani, A.; Yamaguchi, N.; Nasu, M. High-frequency phage-mediated gene transfer among Escherichia coli cells, determined at the single-cell level. Appl. Environ. Microbiol. 2007, 73, 3291–3299. [Google Scholar] [CrossRef] [PubMed]
- Subirats, J.; Sanchez-Melsio, A.; Borrego, C.M.; Luis Balcazar, J.; Simonet, P. Metagenomic analysis reveals that bacteriophages are reservoirs of antibiotic resistance genes. Int. J. Antimicrob. Agents 2016, 48, 163–167. [Google Scholar] [CrossRef]
- Larranaga, O.; Brown-Jaque, M.; Quiros, P.; Gomez-Gomez, C.; Blanch, A.R.; Rodriguez-Rubio, L.; Muniesa, M. Phage particles harboring antibiotic resistance genes in fresh-cut vegetables and agricultural soil. Environ. Int. 2018, 115, 133–141. [Google Scholar] [CrossRef] [PubMed]
- Feng, T.S.; Su, W.H.; Zhu, J.X.; Yang, J.W.; Wang, Y.; Zhou, R.; Yu, Q.; Li, H. Corpse decomposition increases the diversity and abundance of antibiotic resistance genes in different soil types in a fish model. Environ. Pollut. 2021, 286, 117560. [Google Scholar] [CrossRef] [PubMed]
- Obayomi, O.; Seyoum, M.M.; Ghazaryan, L.; Tebbe, C.C.; Murase, J.; Bernstein, N.; Gillor, O. Soil texture and properties rather than irrigation water type shape the diversity and composition of soil microbial communities. Appl. Soil Ecol. 2021, 161, 103834. [Google Scholar] [CrossRef]
- Goldberg, E.D. Black Carbon in the Environment; John Wiley: New York, NY, USA, 1985; p. 198. [Google Scholar]
- Belviso, C. Zeolite for Potential Toxic Metal Uptake from Contaminated Soil: A Brief Review. Processes 2020, 8, 820. [Google Scholar] [CrossRef]
- Zhou, G.X.; Qiu, X.W.; Wu, X.Y.; Lu, S.B. Horizontal gene transfer is a key determinant of antibiotic resistance genes profiles during chicken manure composting with the addition of biochar and zeolite. J. Hazard. Mater. 2021, 408, 124883. [Google Scholar] [CrossRef]
- Zheng, H.; Feng, N.L.; Yang, T.N.; Shi, M.; Wang, X.; Zhang, Q.; Zhao, J.; Li, F.; Sun, K.; Xing, B. Individual and combined applications of biochar and pyroligneous acid mitigate dissemination of antibiotic resistance genes in agricultural soil. Sci. Total Environ. 2021, 796, 148962. [Google Scholar] [CrossRef] [PubMed]
- Duan, M.L.; Li, H.C.; Gu, J.; Tuo, X.X.; Sun, W.; Qian, X.; Wang, X.J. Effects of biochar on reducing the abundance of oxytetracycline, antibiotic resistance genes, and human pathogenic bacteria in soil and lettuce. Environ. Pollut. 2017, 224, 787–795. [Google Scholar] [CrossRef]
- Ye, M.; Sun, M.M.; Feng, Y.F.; Wan, J.Z.; Xin, J. Effect of biochar amendment on the control of soil sulfonamides, antibiotic-resistant bacteria, and gene enrichment in lettuce tissues. J. Hazard. Mater. 2015, 309, 219–227. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Chen, Z.Q.; Wen, Q.X. Impacts of biochar on anaerobic digestion of swine manure: Methanogenesis and antibiotic resistance genes dissemination. Bioresour. Technol. 2021, 324, 124679. [Google Scholar] [CrossRef]
- Liang, Y.T.; Pei, M.; Wang, D.D.; Cao, S.N.; Xiao, X.; Sun, B. Improvement of soil ecosystem multifunctionality by dissipating manure-induced antibiotics and resistance genes. Environ. Sci. Technol. 2017, 51, 4988–4998. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.L.; Fan, X.T.; Zhu, D.; An, X.L.; Su, J.Q.; Cui, L. Effect of biochar amendment on the alleviation of antibiotic resistance in soil and phyllosphere of Brassica chinensis L. Soil Biol. Biochem. 2018, 119, 74–82. [Google Scholar] [CrossRef]
- Sui, Q.W.; Zhang, J.Y.; Chen, M.X.; Wang, R.; Wang, Y.; Wei, Y. Fate of microbial pollutants and evolution of antibiotic resistance in three types of soil amended with swine slurry. Environ. Pollut. 2019, 245, 353–362. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.P.; Zhao, S.; Chen, Y.R.; Yang, J.; Hou, L.-j.; Liu, M.; Yang, Y. Antibiotic resistance genes in sediments of the Yangtze Estuary: From 2007 to 2019. Sci. Total Environ. 2020, 744, 140713. [Google Scholar] [CrossRef] [PubMed]
- Huang, Q.; Chen, J.X.; Zhu, J.J.; Hao, X.; Dao, G.; Chen, W.; Cai, P.; Huang, Q. Divergent bacterial transformation exerted by soil minerals. Sci. Total Environ. 2021, 784, 147173. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.J.; Sheng, X.; Zhang, W.; Lin, Z.; Gao, Y. Nonmonotonic effect of montmorillonites on the horizontal transfer of antibiotic resistance genes to bacteria. Environ. Sci. Technol. Lett. 2020, 7, 421–427. [Google Scholar] [CrossRef]
- Fang, J.; Jin, L.; Meng, Q.K.; Shan, S.; Wang, D.; Lin, D.H. Biochar effectively inhibits the horizontal transfer of antibiotic resistance genes via transformation. J. Hazard. Mater. 2022, 423, 127150. [Google Scholar] [CrossRef]
- Paget, E.; Simonet, P. Development of engineered genomic DNA to monitor the natural transformation of Pseudomonas stutzeri in soil-like microcosms. Can. J. Microbiol. 1997, 43, 78–84. [Google Scholar] [CrossRef]
- Chamier, B.; Lorenz, M.G.; Wackernagel, W. Natural transformation of Acinetobacter calcoaceticus by plasmid DNA adsorbed on sand and groundwater aquifer material. Appl. Environ. Microbiol. 1993, 59, 1662–1667. [Google Scholar] [CrossRef]
- Wang, L.; Yuan, L.; Li, Z.H.; Zhang, X.; Sheng, G.-P. Quantifying the occurrence and transformation potential of extracellular polymeric substances (EPS)-associated antibiotic resistance genes in activated sludge. J. Hazard. Mater. 2021, 408, 124428. [Google Scholar] [CrossRef]
- Dong, P.Y.; Wang, H.; Fang, T.T.; Wang, Y.; Ye, Q.H. Assessment of extracellular antibiotic resistance genes (eARGs) in typical environmental samples and the transforming ability of eARG. Environ. Int. 2019, 125, 90–96. [Google Scholar] [CrossRef]
- Liu, X.M.; Wang, D.; Wang, L.; Tang, J.C. Dissolved biochar eliminates the effect of Cu(II) on the transfer of antibiotic resistance genes between bacteria. J. Hazard. Mater. 2022, 424, 127251. [Google Scholar] [CrossRef]
- Zheng, H.; Wang, R.R.; Zhang, Q.; Zhao, J.; Li, F.; Luo, X.; Xing, B.S. Pyroligneous acid mitigated dissemination of antibiotic resistance genes in soil. Environ. Int. 2020, 145, 106158. [Google Scholar] [CrossRef] [PubMed]
- Yu, K.Q.; Chen, F.R.; Yue, L.; Luo, Y.; Wang, Z.; Xing, B. CeO2 nanoparticles regulate the propagation of antibiotic resistance genes by altering cellular contact and plasmid transfer. Environ. Sci. Technol. 2020, 54, 10012–10021. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.M.; Wang, D.; Tang, J.C.; Liu, F.; Wang, L. Effect of dissolved biochar on the transfer of antibiotic resistance genes between bacteria. Environ. Pollut. 2021, 288, 117718. [Google Scholar] [CrossRef]
- Zhou, G.W.; Zheng, F.; Fan, X.T.; Li, M.-J.; Sun, Q.-Y.; Zhu, Y.-G.; Yang, X.R. Host age increased conjugal plasmid transfer in gut microbiota of the soil invertebrate Caenorhabditis elegans. J. Hazard. Mater. 2022, 424, 127525. [Google Scholar] [CrossRef] [PubMed]
- Beaber, J.W.; Hochhut, B.; Waldor, M.K. SOS response promotes horizontal dissemination of antibiotic resistance genes. Nature 2004, 427, 72–74. [Google Scholar] [CrossRef]
- Lu, Y.; Zeng, J.M.; Wu, B.N.; Shunmei, E.; Wang, L.; Cai, R.; Zhang, N.; Li, Y.; Huang, X.; Huang, B.; et al. Quorum sensing N-acyl homoserine lactones-SdiA suppresses Escherichia coli-Pseudomonas aeruginosa conjugation through inhibiting tral expression. Front. Cell. Infect. Microbiol. 2017, 7, 7. [Google Scholar] [CrossRef]
- Zheng, J.; Chen, T.; Chen, H. Antibiotic resistome promotion in drinking water during biological activated carbon treatment: Is it influenced by quorum sensing? Sci. Total Environ. 2018, 612, 1–8. [Google Scholar] [CrossRef]
- Lu, W.W.; Wang, M.; Wu, J.Q.; Jiang, Q.Y.; Jin, J.R.; Jin, Q.; Yang, W.W.; Chen, J.; Wang, Y.J.; Xiao, M. Spread of chloramphenicol and tetracycline resistance genes by plasmid mobilization in agricultural soil. Environ. Pollut. 2020, 260, 113998. [Google Scholar] [CrossRef]
- van Elsas, J.D.; Chiurazzi, M.; Mallon, C.A.; Elhottova, D.; Kristufek, V.; Salles, J.F. Microbial diversity determines the invasion of soil by a bacterial pathogen. Proc. Natl. Acad. Sci. USA 2012, 109, 1159–1164. [Google Scholar] [CrossRef]
- Chen, Q.L.; An, X.L.; Li, H.; Zhu, Y.G.; Su, J.Q.; Cui, L. Do manure-borne or indigenous soil microorganisms influence the spread of antibiotic resistance genes in manured soil? Soil Biol. Biochem. 2017, 114, 229–237. [Google Scholar] [CrossRef]
- Van Acker, H.; Coenye, T. The role of reactive oxygen species in antibiotic-mediated killing of bacteria. Trends Microbiol. 2017, 25, 456–466. [Google Scholar] [CrossRef] [PubMed]
- Cabiscol, E.; Tamarit, J.; Ros, J. Oxidative stress in bacteria and protein damage by reactive oxygen species. Int. Microbiol. 2000, 3, 3–8. [Google Scholar] [PubMed]
- Reniere, M.L. Reduce, Induce, Thrive: Bacterial Redox Sensing during Pathogenesis. J. Bacteriol. 2018, 200, e00128-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, X.F.; Yin, H.L.; Li, G.Y.; Wang, W.; Wong, P.K.; Zhao, H.; An, T. Antibiotic-resistance gene transfer in antibiotic-resistance bacteria under different light irradiation: Implications from oxidative stress and gene expression. Water Res. 2019, 149, 282–291. [Google Scholar] [CrossRef]
- Han, X.; Lv, P.; Wang, L.G.; Long, F.; Ma, X.-L.; Liu, C.; Feng, Y.-J.; Yang, M.-F.; Xiao, X. Impact of nano-TiO2 on horizontal transfer of resistance genes mediated by filamentous phage transduction. Environ. Sci.-Nano 2020, 7, 1214–1224. [Google Scholar] [CrossRef]
- Sagar, S.; Kumar, R. Role of SOS response in bacterial drug resistance. Int. J. Pharm. Sci. Rev. Res. 2014, 25, 102–105. [Google Scholar]
- Ding, C.S.; Pan, J.; Jin, M.; Yang, D.; Shen, Z.; Wang, J.; Zhang, B.; Liu, W.; Fu, J.; Guo, X.; et al. Enhanced uptake of antibiotic resistance genes in the presence of nanoalumina. Nanotoxicology 2016, 10, 1051–1060. [Google Scholar] [CrossRef]
- Tan, H.D.; Fu, L.; Seno, M. Optimization of bacterial plasmid transformation using nanomaterials based on the yoshida effect. Int. J. Mol. Sci. 2010, 11, 4962–4973. [Google Scholar] [CrossRef]
- Hernandez, G.; Bollini, A.; Huarte, M.; Bazzoni, G.; Piehl, L.; Chiarotto, M.; de Celis, E.R.; Rasia, M. In vitro effect of aluminium upon erythrocyte membrane properties. Clin. Hemorheol. Microcirc. 2008, 40, 191–205. [Google Scholar] [CrossRef]
- Yamamoto, Y.; Hachiya, A.; Matsumoto, H. Oxidative damage to membranes by a combination of aluminum and iron in suspension-cultured tobacco cells. Plant Cell Physiol. 1997, 38, 1333–1339. [Google Scholar] [CrossRef]
- Ma, W.T.; Peng, D.H.; Walker, S.L.; Cao, B.; Gao, C.-H.; Huang, Q.; Cai, P. Bacillus subtilis biofilm development in the presence of soil clay minerals and iron oxides. Npj Biofilms Microbiomes 2017, 3, 4. [Google Scholar] [CrossRef]
- Ouyang, K.; Walker, S.L.; Yu, X.Y.; Gao, C.-H.; Huang, Q.; Cai, P. Metabolism, survival, and gene expression of Pseudomonas putida to hematite nanoparticles mediated by surface-bound humic acid. Environ. Sci.-Nano 2018, 5, 682–695. [Google Scholar] [CrossRef]
- Wang, Q.; Liu, L.; Hou, Z.L.; Wang, L.; Ma, D.; Yang, G.; Guo, S.; Luo, J.; Qi, L.; Luo, Y. Heavy metal copper accelerates the conjugative transfer of antibiotic resistance genes in freshwater microcosms. Sci. Total Environ. 2020, 717, 137055. [Google Scholar] [CrossRef] [PubMed]
- Li, M.Z.; He, Y.N.; Sun, J.; Li, J.; Bai, J.; Zhang, C. Chronic exposure to an environmentally relevant triclosan concentration induces persistent triclosan resistance but reversible antibiotic tolerance in Escherichia coli. Environ. Sci. Technol. 2019, 53, 3277–3286. [Google Scholar] [CrossRef] [PubMed]
- Chen, I.; Christie, P.J.; Dubnau, D. The ins and outs of DNA transfer in bacteria. Science 2005, 310, 1456–1460. [Google Scholar] [CrossRef] [PubMed]
- Samuels, A.L.; Lanka, E.; Davies, J.E. Conjugative junctions in RP4-mediated mating of Escherichia coli. J. Bacteriol. 2000, 182, 2709–2715. [Google Scholar] [CrossRef] [PubMed]
- Zhong, X.; Krol, J.E.; Top, E.M.; Krone, S.M. Accounting for mating pair formation in plasmid population dynamics. J. Theor. Biol. 2010, 262, 711–719. [Google Scholar] [CrossRef] [PubMed]
- Diethmaier, C.; Chawla, R.; Canzoneri, A.; Kearns, D.B.; Lele, P.P.; Dubnau, D. Viscous drag on the flagellum activates Bacillus subtilis entry into the K-state. Mol. Microbiol. 2017, 106, 367–380. [Google Scholar] [CrossRef]
- Karygianni, L.; Ren, Z.; Koo, H.; Thurnheer, T. Biofilm matrixome: Extracellular components in structured microbial communities. Trends Microbiol. 2020, 28, 668–681. [Google Scholar] [CrossRef]
- Tsuneda, S.; Aikawa, H.; Hayashi, H.; Yuasa, A.; Hirata, A. Extracellular polymeric substances responsible for bacterial adhesion onto solid surface. FEMS Microbiol. Lett. 2003, 223, 287–292. [Google Scholar] [CrossRef]
- Hu, X.J.; Kang, F.X.; Yang, B.; Zhang, W.; Qin, C.; Gao, Y. Extracellular polymeric substances acting as a permeable barrier hinder the lateral transfer of antibiotic resistance genes. Front. Microbiol. 2019, 10, 736. [Google Scholar] [CrossRef]
- Xayarath, B.; Freitag, N.E. When being alone is enough: Noncanonical functions of canonical bacterial quorum-sensing systems. Future Microbiol. 2016, 11, 1447–1459. [Google Scholar] [CrossRef]
- Raju Maddela, N.; Sheng, B.; Yuan, S.; Zhou, Z.; Villamar-Torres, R.; Meng, F. Roles of quorum sensing in biological wastewater treatment: A critical review. Chemosphere 2019, 221, 616–629. [Google Scholar] [CrossRef] [PubMed]
- Shank, E.A.; Kolter, R. Extracellular signaling and multicellularity in Bacillus subtilis. Curr. Opin. Microbiol. 2011, 14, 741–747. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inaoka, T.; Ochi, K. Re1A protein is involved in induction of genetic competence in certain Bacillus subtilis strains by moderating the level of intracellular GTP. J. Bacteriol. 2002, 184, 3923–3930. [Google Scholar] [CrossRef] [PubMed]
- Lopez, D.; Vlamakis, H.; Kolter, R. Generation of multiple cell types in Bacillus subtilis. FEMS Microbiol. Rev. 2009, 33, 152–163. [Google Scholar] [CrossRef] [PubMed]
- Seitz, P.; Blokesch, M. Cues and regulatory pathways involved in natural competence and transformation in pathogenic and environmental gram-negative bacteria. FEMS Microbiol. Rev. 2013, 37, 336–363. [Google Scholar] [CrossRef]
- Vestergard, M.; Ekelund, F.; Winding, A.; Jacobsen, C.S.; Christensen, S. Starved bacteria retain their size but lose culturability-Lessons from a 5000 years old undisturbed A-horizon. Soil Biol. Biochem. 2011, 43, 1379–1382. [Google Scholar] [CrossRef]
- Johnston, C.; Martin, B.; Fichant, G.; Polard, P.; Claverys, J.-P. Bacterial transformation: Distribution, shared mechanisms and divergent control. Nat. Rev. Microbiol. 2014, 12, 181–196. [Google Scholar] [CrossRef]
- Lu, N.; Zilles, J.L.; Nguyen, T.H. Adsorption of extracellular chromosomal DNA and its effects on natural transformation of Azotobacter vinelandii. Appl. Environ. Microbiol. 2010, 76, 4179. [Google Scholar] [CrossRef]
- Pei, R.T.; Gunsch, C.K. Plasmid conjugation in an activated sludge microbial community. Environ. Eng. Sci. 2009, 26, 825–831. [Google Scholar] [CrossRef]
- Dahlberg, C.; Bergstrom, M.; Hermansson, M. In situ detection of high levels of horizontal plasmid transfer in marine bacterial communities. Appl. Environ. Microbiol. 1998, 64, 2670–2675. [Google Scholar] [CrossRef] [PubMed]
- Bandyopadhyay, A.; O’Brien, S.; Frank, K.L.; Dunny, G.M.; Hu, W.S. Antagonistic donor density effect conserved in multiple Enterococcal conjugative plasmids. Appl. Environ. Microbiol. 2016, 82, 4537–4545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
| Place | Soil Type | ARGs | Relative Abundance |
|---|---|---|---|
| China [26] | Feedlot vicinity | tetM, tetO, tetQ, tetW | 10−5–10−2 |
| China [27] | Feedlot vicinity | tetB(P), tetM, tetO, tetW | 10−3–10−6 |
| China [28] | Feedlots | tetA(P), tetG, tetC, tetL, tetX, tetM, tetA | 10−2–10−4 |
| China [29] | Feedlots | tetA, tetB, tetM | 10−6–10−1 |
| China [30] | Farmland | tetB(P), tetM, tetO, tetQ, tetT, tetW | 10−8–10−2 |
| China [31] | Farmland | tetG, tetY, tetZ | 10−7–10−4 |
| China [32] | Farmland | tetB(P), tetC, tetG, tetL, tetO, tetS, tetW, tetZ | 10−6–10−1 |
| Italy [33] | Feedlots | tetQ, tetW | 10−9–10−5 |
| India [34] | Feedlots | tetA, tetW | 10−1 a |
| America [35] | Farmland | tetO, tetW | 10−7–10−4 |
| Austria [36] | Farmland | tetW | 10−5–10−4 |
| The Netherlands [24] | Typical sites | tetM, tetO, tetQ, tetW | 10−4–10−2 |
| Scotland [16] | Typical sites | tetM, tetQ, tetW | 10−5–10−2 |
| Scotland [37] | Farm | tetA, tetB, tetC, tetG, tetW | 10−6–10−5 |
| Australia [38] | Residential area | tetM, tetW | 10−9–10−2 |
| Medium | Important Conclusions | |
|---|---|---|
| Results | Reasons | |
| Kaolinite, illite, and montmorillonite [12] | Plasmids adsorbed on minerals could resist higher concentrations of nucleases and form more transformants than free plasmids. | The adsorption of the nuclease on minerals protected the plasmids, but it can still be involved in transformation. |
| Kaolinite, Goethite, and montmorillonite [95] | Low concentrations (1–2 g/L) have little effect; high concentration (10 g/L) of kaolinite and montmorillonite inhibited transformation; high concentration (10 g/L) of goethite promoted transformation. | Kaolinite and montmorillonite: strong adsorption to competence stimulating factor, decrease the expression level of competent genes (phrC, comS); goethite: increase cell membrane damage. |
| Montmorillonite [96] | Low concentration (about 0–0.025 g/L) promoted transformation; high concentration (about 0.025–2 g/L) inhibited transformation. | Low concentration: increase the contact between plasmids and cells; forming holes on cell membrane; High concentration: plasmids were adsorbed; heavy metals released from montmorillonite cause the aggregation of the plasmids. |
| Biochar [97] | Significantly inhibited the transformation of extracellular antibiotic resistance genes (eARGs) | Biochar dissolutions: Induce intramolecular condensation and agglomeration of plasmids; decrease the cell membrane permeability; biochar solids: Adsorb plasmids and deactivate E. coli. |
| Soil microcosm [98] | DNA adsorbed on soil particles still transformed competent cells | Minerals did not inhibit the transformation, but blocked DNA contact with the recipient. |
| Soil microcosm [99] | Plasmid adsorbed on sand transformed significantly less efficient than did plasmid in solution; the transformation by sand-adsorbed chromosomal was as high as that by plasmid in solution. | Transformation occurred by direct uptake of DNA from the mineral surfaces; transformation requires multiple plasmids, and the probability of multiple free plasmids meeting bacteria at the same time is higher than that on mineral surfaces; the chances of bacteria taking up DNA on the mineral surface are proportional to the size of the DNA, and chromosomes of the same mass are larger and easier to take up. |
| Activated sludge EPS [100] | The transformation ability of free ARGs was higher than that in activated sludge extracellular polymeric substances (EPS) when calculated per ng DNA, and lower when calculated per g volatile suspended solids. | Activated sludge EPS is rich in ARGs. |
| Sediment [101] | The transformation efficiency of adsorbed eARG was higher than that of free eARGs. | Sand adsorbed bacteria and plasmids at the same time, facilitating contact between the two, and was related to the conformation of the plasmid. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shi, H.; Hu, X.; Li, W.; Zhang, J.; Hu, B.; Lou, L. Soil Component: A Potential Factor Affecting the Occurrence and Spread of Antibiotic Resistance Genes. Antibiotics 2023, 12, 333. https://doi.org/10.3390/antibiotics12020333
Shi H, Hu X, Li W, Zhang J, Hu B, Lou L. Soil Component: A Potential Factor Affecting the Occurrence and Spread of Antibiotic Resistance Genes. Antibiotics. 2023; 12(2):333. https://doi.org/10.3390/antibiotics12020333
Chicago/Turabian StyleShi, Hongyu, Xinyi Hu, Wenxuan Li, Jin Zhang, Baolan Hu, and Liping Lou. 2023. "Soil Component: A Potential Factor Affecting the Occurrence and Spread of Antibiotic Resistance Genes" Antibiotics 12, no. 2: 333. https://doi.org/10.3390/antibiotics12020333