
Pseudomonas aeruginosa and Staphylococcus aureus Display Differential Proteomic Responses to the Silver(I) Compound, SBC3


Abstract
:1. Introduction
2. Results
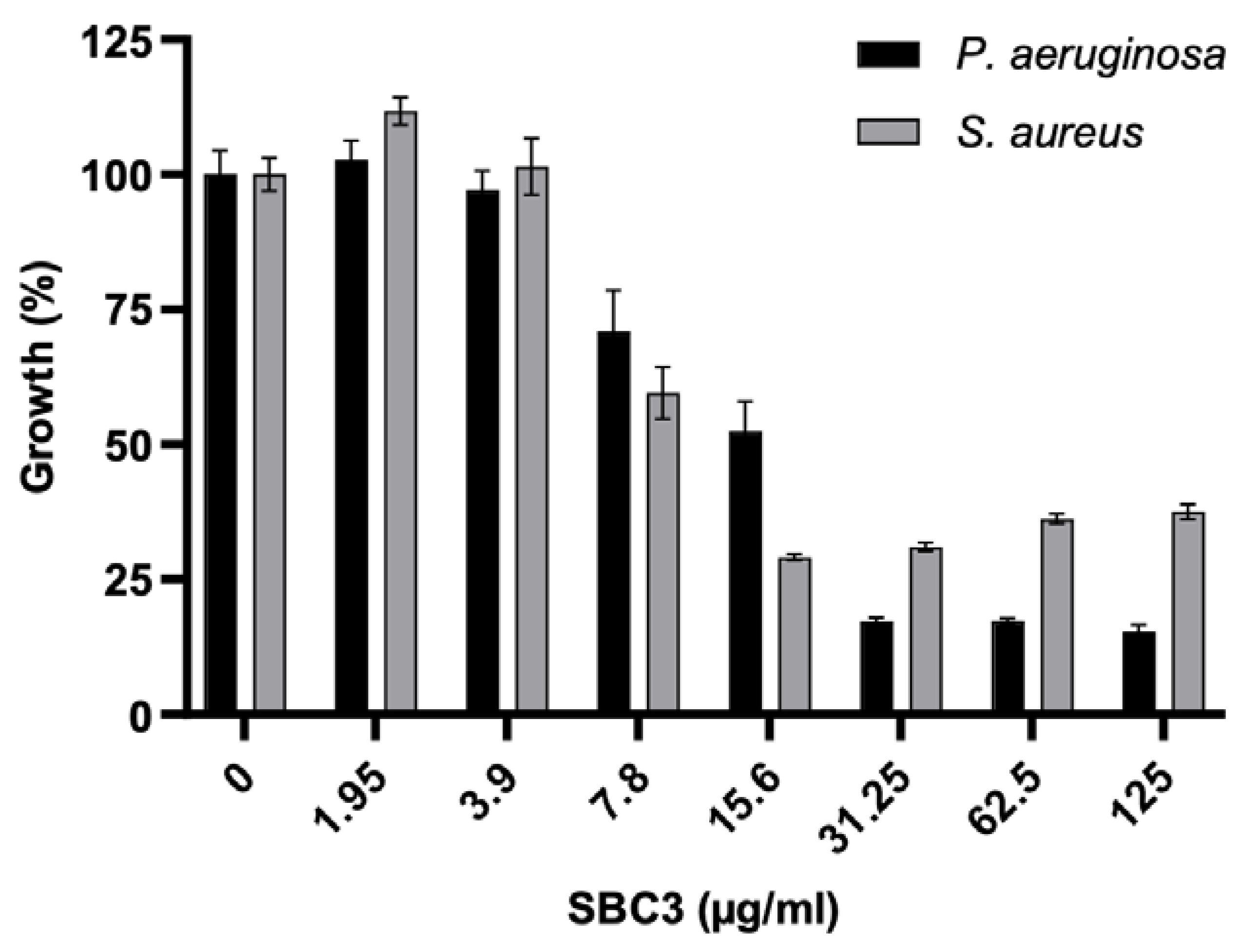
2.1. The Antimicrobial Effect of SBC3
2.2. Characterisation of the Proteomic Response of Bacteria to SBC3
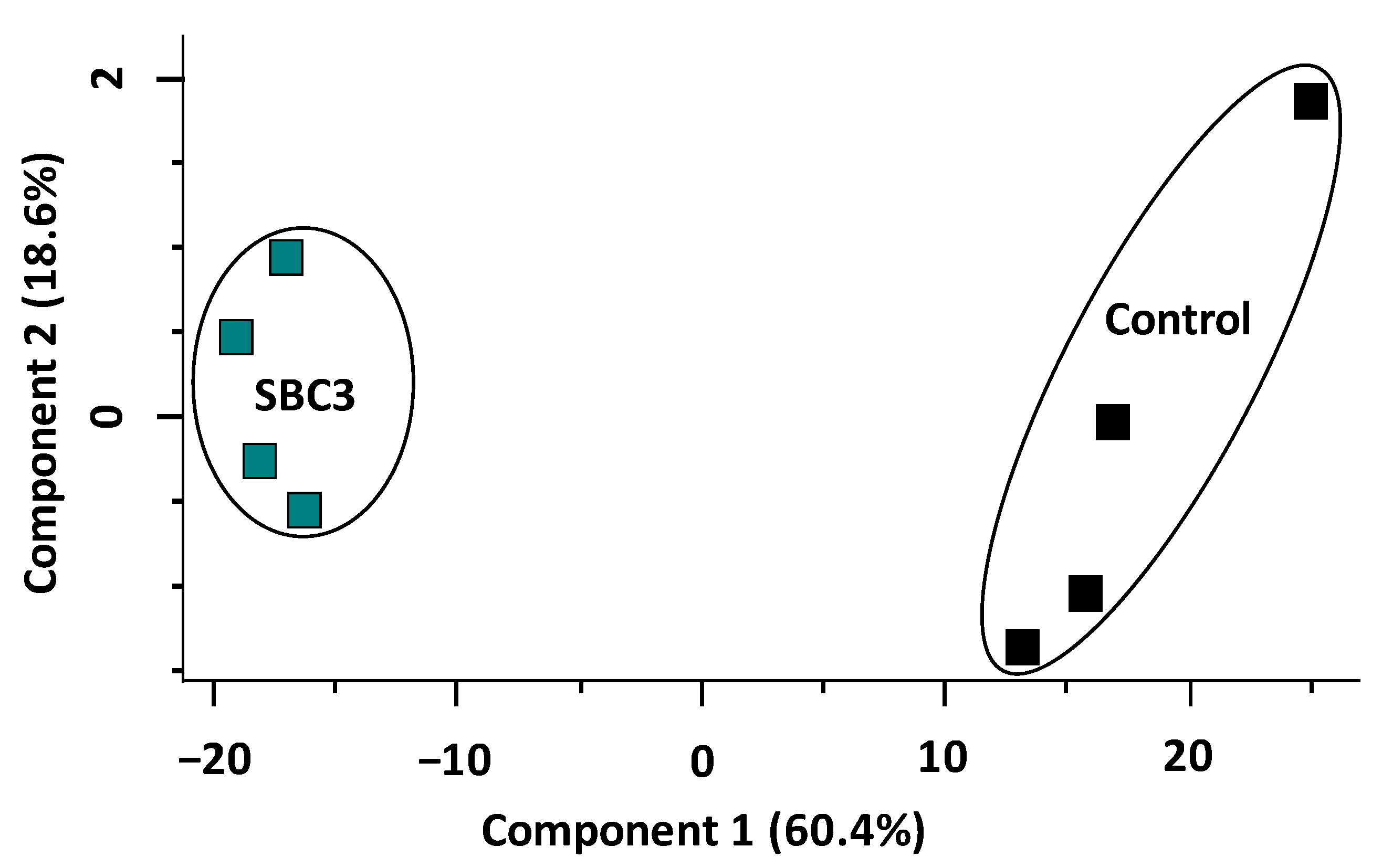
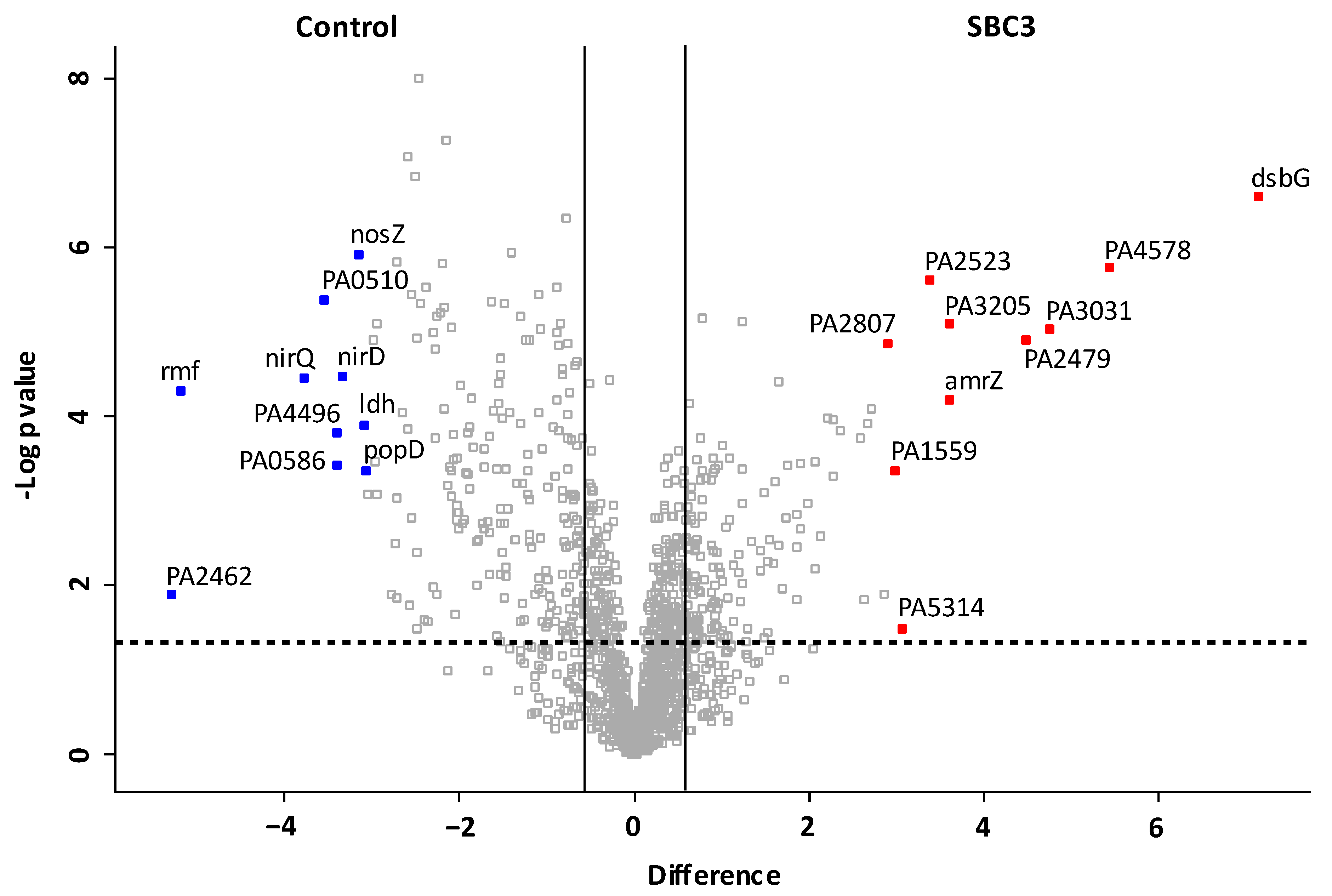
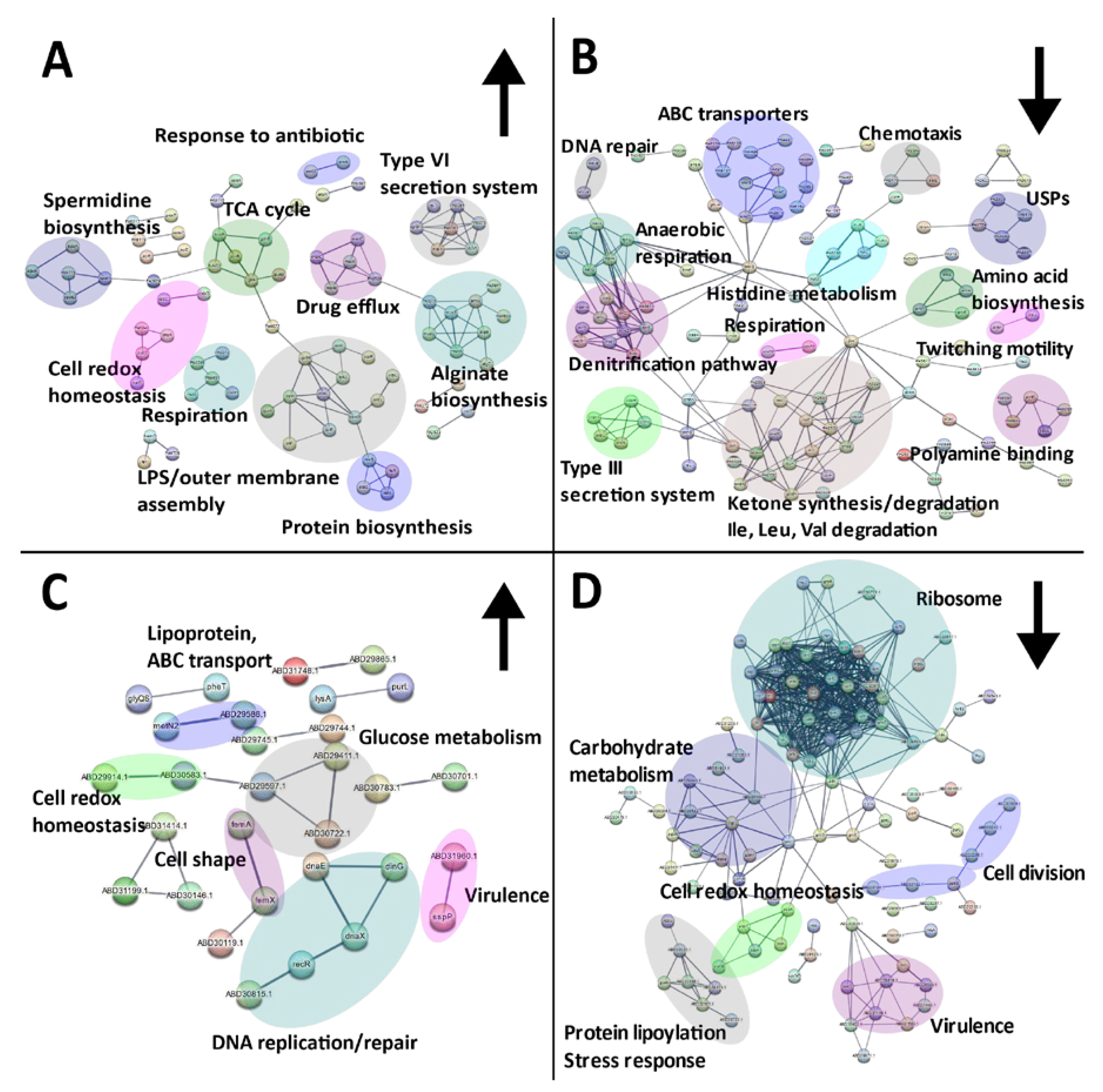
2.3. Characterisation of the Effect of SBC3 on the Proteome of P. aeruginosa
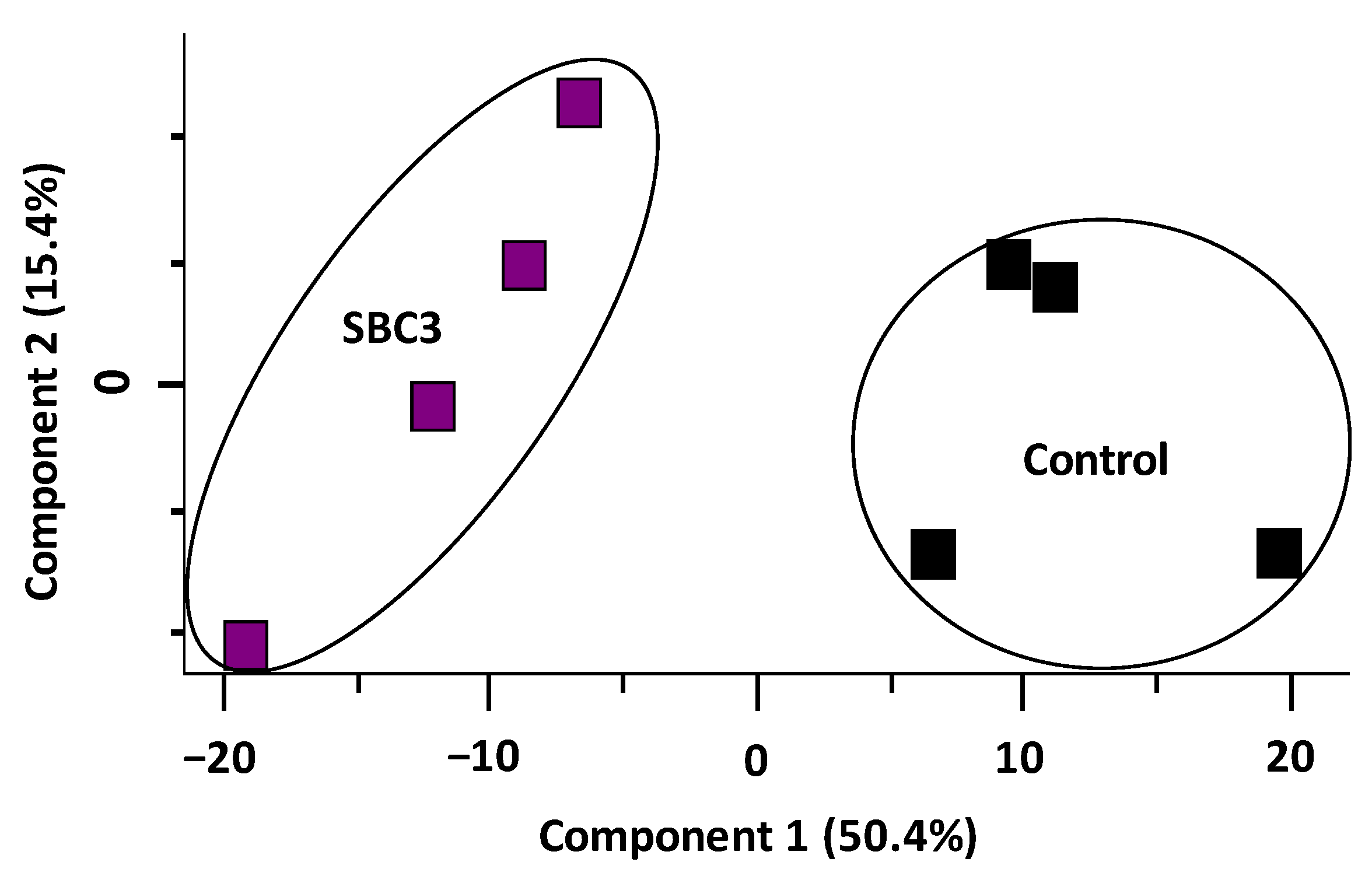
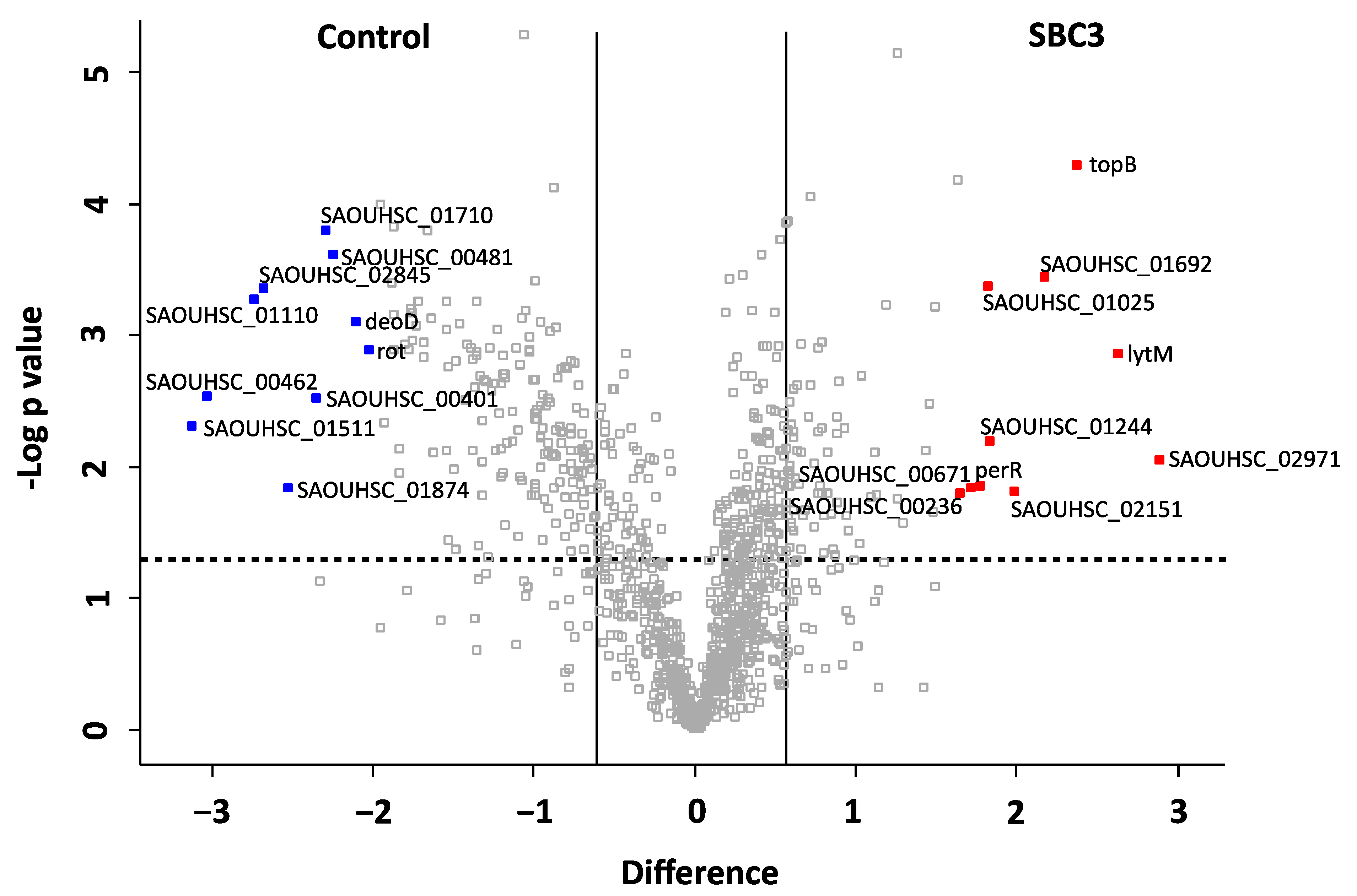
2.4. Characterisation of the Effect of SBC3 on the Proteome of S. aureus
2.5. Interaction Network Analysis on the Response of P. aeruginosa and S. aureus to SBC3
3. Discussion
3.1. SBC3 Compromises Cell Structural Integrity in Both S. aureus and P. aeruginosa
3.2. The Proteomic Effects of SBC3 on P. aeruginosa
3.2.1. Translocation and Biofilm Formation
3.2.2. Virulence
3.2.3. Anaerobic Respiration/Stress Response
3.2.4. Detoxification Mechanisms
3.2.5. Aerobic Respiration
3.3. The Proteomic Response of S. aureus to SBC3
3.3.1. Protein Synthesis
3.3.2. Protein Lipoylation
3.3.3. Glucose Metabolism
3.3.4. Cell Redox Homeostasis
4. Materials and Methods
4.1. Bacterial Culture Conditions
4.2. Antibacterial Susceptibility Assays
4.3. Proteomic Analysis of P. aeruginosa Treated with SBC3
4.4. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Kumar, M.; Sarma, D.K.; Shubham, S.; Kumawat, M.; Verma, V.; Nina, P.B.; JP, D.; Kumar, S.; Singh, B.; Tiwari, R.R. Futuristic Non-antibiotic Therapies to Combat Antibiotic Resistance: A Review. Front. Microbiol. 2021, 12, 609459. [Google Scholar] [CrossRef] [PubMed]
- Lee Ventola, M.C. The Antibiotics Resistance Crisis Part 1: Causes and Threats. Pharm. Ther. 2015, 40, 277–283. [Google Scholar]
- Worthington, R.J.; Melander, C. Overcoming resistance to β-Lactam antibiotics. J. Org. Chem. 2013, 78, 4207–4213. [Google Scholar] [CrossRef] [PubMed]
- Bush, K.; Bradford, P.A. β-lactams and β-lactamase inhibitors: An overview. Cold Spring Harb. Perspect. Med. 2016, 6, a025247. [Google Scholar] [CrossRef]
- Romaniuk, J.A.H.; Cegelski, L. Peptidoglycan and Teichoic Acid Levels and Alterations in Staphylococcus aureus by Cell-Wall and Whole-Cell Nuclear Magnetic Resonance. Biochemistry 2018, 57, 3966–3975. [Google Scholar] [CrossRef]
- Delcour, A.H. Outer membrane permeability and antibiotic resistance. Biochim. Biophys. Acta-Proteins Proteom. 2009, 1794, 808–816. [Google Scholar] [CrossRef]
- Li, H.; Luo, Y.F.; Williams, B.J.; Blackwell, T.S.; Xie, C.M. Structure and function of OprD protein in Pseudomonas aeruginosa: From antibiotic resistance to novel therapies. Int. J. Med. Microbiol. 2012, 302, 63–68. [Google Scholar] [CrossRef]
- Jubeh, B.; Breijyeh, Z.; Karaman, R. Resistance of gram-positive bacteria to current antibacterial agents and overcoming approaches. Molecules 2020, 25, 2888. [Google Scholar] [CrossRef]
- Aminov, R.I. A brief history of the antibiotic era: Lessons learned and challenges for the future. Front. Microbiol. 2010, 1, 134. [Google Scholar] [CrossRef]
- Özdemir, I.; Özcan, E.Ö.; Günal, S.; Gürbüz, N. Synthesis and antimicrobial activity of novel Ag-N-Heterocyclic carbene complexes. Molecules 2010, 15, 2499–2508. [Google Scholar] [CrossRef]
- Prencipe, F.; Zanfardino, A.; Di Napoli, M.; Rossi, F.; D’errico, S.; Piccialli, G.; Mangiatordi, G.F.; Saviano, M.; Ronga, L.; Varcamonti, M.; et al. Silver (I) n-heterocyclic carbene complexes: A winning and broad spectrum of antimicrobial properties. Int. J. Mol. Sci. 2021, 22, 2497. [Google Scholar] [CrossRef]
- Hindi, K.M.; Panzner, M.J.; Tessier, C.A.; Cannon, C.L.; Youngs, W.J. The medicinal applications of imidazolium carbene-metal complexes. Chem. Rev. 2009, 109, 3859–3884. [Google Scholar] [CrossRef]
- O’Beirne, C.; Piatek, M.E.; Fossen, J.; Müller-Bunz, H.; Andes, D.R.; Kavanagh, K.; Patil, S.A.; Baumann, M.; Tacke, M. Continuous flow synthesis and antimicrobial evaluation of NHC* silver carboxylate derivatives of SBC3 in vitro and in vivo. Metallomics 2021, 13, mfaa011. [Google Scholar] [CrossRef]
- O’Loughlin, J.; Napolitano, S.; Alkhathami, F.; O’Beirne, C.; Marhöfer, D.; O’Shaughnessy, M.; Howe, O.; Tacke, M.; Rubini, M. The Antibacterial Drug Candidate SBC3 is a Potent Inhibitor of Bacterial Thioredoxin Reductase. ChemBioChem 2021, 22, 1093–1098. [Google Scholar] [CrossRef]
- Alexander, J.W. History of the medical use of silver. Surg. Infect. 2009, 10, 289–292. [Google Scholar] [CrossRef]
- Politano, A.D.; Campbell, K.T.; Rosenberger, L.H.; Sawyer, R.G. Use of silver in the prevention and treatment of infections: Silver review. Surg. Infect. 2013, 14, 8–20. [Google Scholar] [CrossRef]
- Amiri-Dashatan, N.; Koushki, M.; Abbaszadeh, H.A.; Rostami-Nejad, M.; Rezaei-Tavirani, M. Proteomics applications in health: Biomarker and drug discovery and food industry. Iran. J. Pharm. Res. 2018, 17, 1523–1536. [Google Scholar]
- van Duin, D.; Paterson, D.L. Multidrug-Resistant Bacteria in the Community: Trends and Lessons Learned. Infect. Dis. Clin. N. Am. 2016, 30, 377–390. [Google Scholar] [CrossRef]
- Goldsworthy, M.J.H. Gene expression of Pseudomonas aeruginosa and MRSA within a catheter-associated urinary tract infection biofilm model. Biosci. Horiz. 2008, 1, 28–37. [Google Scholar] [CrossRef]
- Trizna, E.Y.; Yarullina, M.N.; Baidamshina, D.R.; Mironova, A.V.; Akhatova, F.S.; Rozhina, E.V.; Fakhrullin, R.F.; Khabibrakhmanova, A.M.; Kurbangalieva, A.R.; Bogachev, M.I.; et al. Bidirectional alterations in antibiotics susceptibility in Staphylococcus aureus—Pseudomonas aeruginosa dual-species biofilm. Sci. Rep. 2020, 10, 14849. [Google Scholar] [CrossRef]
- Reigada, I.; San-Martin-Galindo, P.; Gilbert-Girard, S.; Chiaro, J.; Cerullo, V.; Savijoki, K.; Nyman, T.A.; Fallarero, A.; Miettinen, I. Surfaceome and Exoproteome Dynamics in Dual-Species Pseudomonas aeruginosa and Staphylococcus aureus Biofilms. Front. Microbiol. 2021, 12, 672975. [Google Scholar] [CrossRef]
- Turner, R.J. Is silver the ultimate antimicrobial bullet? Antibiotics 2018, 7, 112. [Google Scholar] [CrossRef] [Green Version]
- Dakal, T.C.; Kumar, A.; Majumdar, R.S.; Yadav, V. Mechanistic basis of antimicrobial actions of silver nanoparticles. Front. Microbiol. 2016, 7, 1831. [Google Scholar] [CrossRef]
- Sharif, S.; Kim, S.J.; Labischinski, H.; Schaefer, J. Characterization of peptidoglycan in fem-deletion mutants of methicillin-resistant staphylococcus aureus by solid-state NMR. Biochemistry 2009, 48, 3100–3108. [Google Scholar] [CrossRef]
- Klein, K.; Sonnabend, M.S.; Frank, L.; Leibiger, K.; Franz-Wachtel, M.; Macek, B.; Trunk, T.; Leo, J.C.; Autenrieth, I.B.; Schütz, M.; et al. Deprivation of the Periplasmic Chaperone SurA Reduces Virulence and Restores Antibiotic Susceptibility of Multidrug-Resistant Pseudomonas aeruginosa. Front. Microbiol. 2019, 10, 100. [Google Scholar] [CrossRef]
- Ebbensgaard, A.; Mordhorst, H.; Aarestrup, F.M.; Hansen, E.B. The role of outer membrane proteins and lipopolysaccharides for the sensitivity of escherichia coli to antimicrobial peptides. Front. Microbiol. 2018, 9, 2153. [Google Scholar] [CrossRef]
- Farhana, A.; Khan, Y.S. Biochemistry, Lipopolysaccharide; StatPearls Publishing: Tampa, FL, USA, 2020. [Google Scholar]
- Johnson, L.; Mulcahy, H.; Kanevets, U.; Shi, Y.; Lewenza, S. Surface-localized spermidine protects the Pseudomonas aeruginosa: Outer membrane from antibiotic treatment and oxidative stress. J. Bacteriol. 2012, 194, 813–826. [Google Scholar] [CrossRef]
- Hentzer, M.; Teitzel, G.M.; Balzer, G.J.; Heydorn, A.; Molin, S.; Givskov, M.; Parsek, M.R. Alginate overproduction affects pseudomonas aeruginosa biofilm structure and function. J. Bacteriol. 2001, 183, 5395–5401. [Google Scholar] [CrossRef]
- Jones, J.C.; Grotewold, N.; Wozniak, D.; Gloag, E. Pseudomonas aeruginosa Initiates a Rapid and Specific Transcriptional Response during Surface Attachment. J. Bacteriol. 2022, 204, e00086-22. [Google Scholar] [CrossRef]
- Persat, A.; Inclan, Y.F.; Engel, J.N.; Stone, H.A.; Gitai, Z. Type IV pili mechanochemically regulate virulence factors in Pseudomonas aeruginosa. Proc. Natl. Acad. Sci. USA 2015, 112, 7563–7568. [Google Scholar] [CrossRef]
- Shi, W.; Sun, H. Type IV pilus-dependent motility and its possible role in bacterial pathogenesis. Infect. Immun. 2002, 70, 1–4. [Google Scholar] [CrossRef]
- Beaussart, A.; Baker, A.E.; Kuchma, S.L.; El-Kirat-Chatel, S.; Otoole, G.A.; Dufrêne, Y.F. Nanoscale adhesion forces of Pseudomonas aeruginosa type IV pili. ACS Nano 2014, 8, 10723–10733. [Google Scholar] [CrossRef] [Green Version]
- Tammam, S.; Sampaleanu, L.M.; Koo, J.; Manoharan, K.; Daubaras, M.; Burrows, L.L.; Howell, P.L. PilMNOPQ from the Pseudomonas aeruginosa type IV pilus system form a transenvelope protein interaction network that interacts with PilA. J. Bacteriol. 2013, 195, 2126–2135. [Google Scholar] [CrossRef]
- Nguyen, Y.; Sugiman-Marangos, S.; Harvey, H.; Bell, S.D.; Charlton, C.L.; Junop, M.S.; Burrows, L.L. Pseudomonas aeruginosa minor pilins prime type IVa pilus assembly and promote surface display of the PilY1 adhesin. J. Biol. Chem. 2015, 290, 601–611. [Google Scholar] [CrossRef]
- Marko, V.A.; Kilmury, S.L.N.; MacNeil, L.T.; Burrows, L.L. Pseudomonas aeruginosa type IV minor pilins and PilY1 regulate virulence by modulating FimS-AlgR activity. PLoS Pathog. 2018, 14, e1007074. [Google Scholar] [CrossRef]
- Hauser, A.R. The type III secretion system of Pseudomonas aeruginosa: Infection by injection. Nat. Rev. Microbiol. 2009, 7, 654–665. [Google Scholar] [CrossRef]
- Coburn, B.; Sekirov, I.; Finlay, B.B. Type III secretion systems and disease. Clin. Microbiol. Rev. 2007, 20, 535–549. [Google Scholar] [CrossRef]
- Dillon, M.M.; Almeida, R.N.D.; Laflamme, B.; Martel, A.; Weir, B.S.; Desveaux, D.; Guttman, D.S. Molecular evolution of Pseudomonas syringae type iii secreted effector proteins. Front. Plant Sci. 2019, 10, 148. [Google Scholar] [CrossRef]
- Rangel, S.M.; Diaz, M.H.; Knoten, C.A.; Zhang, A.; Hauser, A.R. The Role of ExoS in Dissemination of Pseudomonas aeruginosa during Pneumonia. PLoS Pathog. 2015, 11, e1004945. [Google Scholar] [CrossRef]
- Emma, K.; Josephine, T.; José, D.A.; Alicia, R.; Arne, R. PopB-PcrV Interactions Are Essential for Pore Formation in the Pseudomonas aeruginosa Type III Secretion System Translocon. MBio 2022, 13, e0238122. [Google Scholar]
- McCaw, M.L.; Lykken, G.L.; Singh, P.K.; Yahr, T.L. ExsD is a negative regulator of the Pseudomonas aeruginosa type III secretion regulon. Mol. Microbiol. 2002, 46, 1123–1133. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Ye, M.; Li, X.; Li, J.; Deng, Z.; Yao, Y.-F.; Ou, H.Y. Identification and characterization of an antibacterial type VI secretion System in the carbapenem-resistant strain Klebsiella pneumoniae HS11286. Front. Cell. Infect. Microbiol. 2017, 7, 442. [Google Scholar] [CrossRef]
- Chen, L.; Zou, Y.; Kronfl, A.A.; Wu, Y. Type VI secretion system of Pseudomonas aeruginosa is associated with biofilm formation but not environmental adaptation. Microbiologyopen 2020, 9, e991. [Google Scholar] [CrossRef] [Green Version]
- Yoon, S.S.; Hennigan, R.F.; Hilliard, G.M.; Ochsner, U.A.; Parvatiyar, K.; Kamani, M.C.; Allen, H.L.; DeKievit, T.R.; Gardner, P.R.; Schwab, U.; et al. Pseudomonas aeruginosa anaerobic respiration in biofilms: Relationships to cystic fibrosis pathogenesis. Dev. Cell 2002, 3, 593–603. [Google Scholar] [CrossRef]
- Masamba, P.; Kappo, A.P. Parasite survival and disease persistence in cystic fibrosis, schistosomiasis and pathogenic bacterial diseases: A role for universal stress proteins? Int. J. Mol. Sci. 2021, 22, 10878. [Google Scholar] [CrossRef]
- Boes, N.; Schreiber, K.; Schobert, M. SpoT-triggered stringent response controls usp gene expression in Pseudomonas aeruginosa. J. Bacteriol. 2008, 190, 7189–7199. [Google Scholar] [CrossRef]
- Trunk, K.; Benkert, B.; Quäck, N.; Münch, R.; Scheer, M.; Garbe, J.; Jänsch, L.; Trost, M.; Wehland, J.; Buer, J.; et al. Anaerobic adaptation in Pseudomonas aeruginosa: Definition of the Anr and Dnr regulons. Environ. Microbiol. 2010, 12, 1719–1733. [Google Scholar] [CrossRef]
- Arat, S.; Bullerjahn, G.S.; Laubenbacher, R. A network biology approach to denitrification in Pseudomonas aeruginosa. PLoS ONE 2015, 10, e0118235. [Google Scholar] [CrossRef]
- Klünemann, T.; Preuß, A.; Adamczack, J.; Rosa, L.F.M.; Harnisch, F.; Layer, G.; Blankenfeldt, W. Crystal Structure of Dihydro-Heme d1 Dehydrogenase NirN from Pseudomonas aeruginosa Reveals Amino Acid Residues Essential for Catalysis. J. Mol. Biol. 2019, 431, 3246–3260. [Google Scholar] [CrossRef]
- Storbeck, S.; Saha, S.; Krausze, J.; Klink, B.U.; Heinz, D.W.; Layer, G. Crystal structure of the heme d 1 biosynthesis enzyme NirE in complex with its substrate reveals new insights into the catalytic mechanism of S-adenosyl-L-methionine-dependent uroporphyrinogen III methyltransferases. J. Biol. Chem. 2011, 286, 26754–26767. [Google Scholar] [CrossRef]
- Adamczack, J.; Hoffmann, M.; Papke, U.; Haufschildt, K.; Nicke, T.; Bröring, M.; Sezer, M.; Weimar, R.; Kuhlmann, U.; Hildebrandt, P.; et al. NirN protein from Pseudomonas aeruginosa is a novel electron-bifurcating dehydrogenase catalyzing the last step of Heme d1 biosynthesis. J. Biol. Chem. 2014, 289, 30753–30762. [Google Scholar] [CrossRef]
- Kawasaki, S.; Arai, H.; Kodama, T.; Igarashi, Y. Gene cluster for dissimilatory nitrite reductase (nir) from Pseudomonas aeruginosa: Sequencing and identification of a locus for heme d1 biosynthesis. J. Bacteriol. 1997, 179, 235–242. [Google Scholar] [CrossRef]
- Schreiber, K.; Krieger, R.; Benkert, B.; Eschbach, M.; Arai, H.; Schobert, M.; Jahn, D. The anaerobic regulatory network required for Pseudomonas aeruginosa nitrate respiration. J. Bacteriol. 2007, 189, 4310–4314. [Google Scholar] [CrossRef] [Green Version]
- Lee, Y.; Song, S.; Sheng, L.; Zhu, L.; Kim, J.S.; Wood, T.K. Substrate binding protein DppA1 of ABC transporter DppBCDF increases biofilm formation in Pseudomonas aeruginosa by inhibiting Pf5 prophage lysis. Front. Microbiol. 2018, 9, 30. [Google Scholar] [CrossRef]
- Kaiser, J.C.; Heinrichs, D.E. Branching out: Alterations in bacterial physiology and virulence due to branched-chain amino acid deprivation. MBio 2018, 9, e01188-18. [Google Scholar] [CrossRef]
- Puchalska, P.; Crawford, P.A. Multi-dimensional Roles of Ketone Bodies in Fuel Metabolism, Signaling, and Therapeutics. Cell Metab. 2017, 25, 262–284. [Google Scholar] [CrossRef]
- Wang, Z.; Bie, P.; Cheng, J.; Lu, L.; Cui, B.; Wu, Q. The ABC transporter YejABEF is required for resistance to antimicrobial peptides and the virulence of Brucella melitensis. Sci. Rep. 2016, 6, 31876. [Google Scholar] [CrossRef]
- López, C.A.; Travers, T.; Pos, K.M.; Zgurskaya, H.I.; Gnanakaran, S. Dynamics of Intact MexAB-OprM Efflux Pump: Focusing on the MexA-OprM Interface. Sci. Rep. 2017, 7, 16521. [Google Scholar] [CrossRef]
- Gotoh, N.; Tsujimoto, H.; Nomura, A.; Okamoto, K.; Tsuda, M.; Nishino, T. Functional replacement of OprJ by OprM in the MexCD-OprJ multidrug efflux system of Pseudomonas aeruginosa. FEMS Microbiol. Lett. 1998, 165, 21–27. [Google Scholar] [CrossRef]
- Wright, B.W.; Kamath, K.S.; Krisp, C.; Molloy, M.P. Proteome profiling of Pseudomonas aeruginosa PAO1 identifies novel responders to copper stress. BMC Microbiol. 2019, 19, 69. [Google Scholar] [CrossRef]
- Kwong, W.K.; Zheng, H.; Moran, N.A. Convergent evolution of a modified, acetate-driven TCA cycle in bacteria. Nat. Microbiol. 2017, 2, 17067. [Google Scholar] [CrossRef]
- Serbanescu, D.; Ojkic, N.; Banerjee, S. Cellular resource allocation strategies for cell size and shape control in bacteria. FEBS J. 2021, 289, 7891–7906. [Google Scholar] [CrossRef]
- Arenz, S.; Wilson, D.N. Bacterial protein synthesis as a target for antibiotic inhibition. Cold Spring Harb. Perspect. Med. 2016, 6, a025361. [Google Scholar] [CrossRef] [Green Version]
- Piatek, M.; O’Beirne, C.; Beato, Z.; Tacke, M.; Kavanagh, K. Exposure of Candida parapsilosis to the silver(I) compound SBC3 induces alterations in the proteome and reduced virulence. Metallomics 2022, 14, mfac046. [Google Scholar] [CrossRef]
- Rowland, E.A.; Snowden, C.K.; Cristea, I.M. Protein lipoylation: An evolutionarily conserved metabolic regulator of health and disease. Curr. Opin. Chem. Biol. 2018, 42, 75–85. [Google Scholar] [CrossRef]
- Teoh, W.P.; Chen, X.; Laczkovich, I.; Alonzo, F. Staphylococcus aureus adapts to the host nutritional landscape to overcome tissue-specific branched-chain fatty acid requirement. Proc. Natl. Acad. Sci. USA 2021, 118, e2022720118. [Google Scholar] [CrossRef]
- Grayczyk, J.P.; Harvey, C.J.; Laczkovich, I.; Alonzo, F. A Lipoylated Metabolic Protein Released by Staphylococcus aureus Suppresses Macrophage Activation. Cell Host Microbe 2017, 2, 678–687.e9. [Google Scholar] [CrossRef]
- Laczkovich, I.; Teoh, W.P.; Flury, S.; Grayczyk, J.P.; Zorzoli, A.; Alonzo, F. Increased flexibility in the use of exogenous lipoic acid by Staphylococcus aureus. Mol. Microbiol. 2018, 109, 150–168. [Google Scholar] [CrossRef]
- Cui, W.Q.; Qu, Q.W.; Wang, J.P.; Bai, J.W.; Bello-Onaghise, G.; Li, Y.A.; Zhou, Y.H.; Chen, X.R.; Liu, X.; Zheng, S.D.; et al. Discovery of potential anti-infective therapy targeting glutamine synthetase in Staphylococcus xylosus. Front. Chem. 2019, 7, 381. [Google Scholar] [CrossRef]
- Wang, H.; Wang, M.; Yang, X.; Xu, X.; Hao, Q.; Yan, A.; Hu, M.; Lobinski, R.; Li, H.; Sun, H. Antimicrobial silver targets glyceraldehyde-3-phosphate dehydrogenase in glycolysis of: E. coli. Chem. Sci. 2019, 10, 7193–7199. [Google Scholar] [CrossRef]
- Wang, H.; Wang, M.; Xu, X.; Gao, P.; Xu, Z.; Zhang, Q.; Li, H.; Yan, A.; Kao, R.Y.T.; Sun, H. Multi-target mode of action of silver against Staphylococcus aureus endows it with capability to combat antibiotic resistance. Nat. Commun. 2021, 12, 3331. [Google Scholar] [CrossRef]
- Tomlinson, K.L.; Lung, T.W.F.; Dach, F.; Annavajhala, M.K.; Gabryszewski, S.J.; Groves, R.A.; Drikic, M.; Francoeur, N.J.; Sridhar, S.H.; Smith, M.L.; et al. Staphylococcus aureus induces an itaconate-dominated immunometabolic response that drives biofilm formation. Nat. Commun. 2021, 12, 1399. [Google Scholar] [CrossRef]
- Margalit, A.; Carolan, J.C.; Sheehan, D.; Kavanagh, K. The Aspergillus fumigatus secretome alters the proteome of Pseudomonas aeruginosa to stimulate bacterial growth: Implications for co-infection. Mol. Cell. Proteom. 2020, 19, 1346–1359. [Google Scholar] [CrossRef]
- Perez-Riverol, Y.; Bai, J.; Bandla, C.; García-Seisdedos, D.; Hewapathirana, S.; Kamatchinathan, S.; Kundu, D.J.; Prakash, A.; Frericks-Zipper, A.; Eisenacher, M.; et al. The PRIDE database resources in 2022: A hub for mass spectrometry-based proteomics evidences. Nucleic Acids Res. 2022, 50, D543–D552. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Protein Name | Protein ID | Function | Fold Change |
|---|---|---|---|---|
| dsbG | Thiol:disulfide interchange protein DsbG | Q9I106 | Disulfide bond formation | +142.1 |
| PA4578 | Uncharacterised | Q9HVK6 | Unknown | +43.7 |
| PA3031 | Uncharacterised | G3XCW5 | Unknown | +27.1 |
| PA2479 | Probable two-component response regulator | Q9I103 | DNA binding | +22.5 |
| PA3205 | Uncharacterised | Q9HZ35 | Unfolded protein binding | +12.2 |
| amrZ | Transcription factor AmrZ | G3XCY4 | Transcription; transcription regulation | +12.2 |
| PA2523 | Probable two-component response regulator | G3XD16 | Positive regulation of proteolysis; positive regulation of single-species biofilm formation on inanimate substrate | +10.4 |
| PA5314 | Cupin_3 domain-containing protein | Q9HTP0 | Unknown | +8.4 |
| PA1559 | NAD_binding_4 domain-containing protein | Q9I3F8 | Unknown | +7.9 |
| PA2807 | Azurin | Q9I036 | Copper binding; electron transfer activity | +7.5 |
| popD | Translocator outer membrane protein PopD | Q9I323 | Chaperone binding; type III secretion system | −8.4 |
| ldh | Leucine dehydrogenase | Q9HYI7 | Cellular amino acid metabolic process; oxidoreductase | −8.6 |
| nosZ | Nitrous-oxide reductase | Q9HYL2 | Oxidoreductase; denitrification pathway | −8.9 |
| nirD | Siroheme decarboxylase NirD subunit | P95412 | Lyase activity; denitrification pathway | −10.1 |
| PA0586 | Uncharacterised | Q9I5V1 | Unknown | −10.5 |
| PA4496 | Probable binding protein component of ABC transporter | Q9HVS5 | Peptide transmembrane transporter activity | −10.6 |
| PA0510 | Uroporphyrinogen-III C-methyltransferase | G3XD80 | Methyltransferase; cobalamin biosynthesis; porphyrin biosynthesis | −11.7 |
| nirQ | Denitrification regulatory protein NirQ | Q51481 | Transcription; transcription regulation | −13.8 |
| rmf | Ribosome modulation factor | Q9HZF9 | Translation regulation | −36.7 |
| PA2462 | Haemagg_act domain-containing protein | Q9I120 | Macromolecule metabolic process; primary metabolic process; nitrogen compound metabolic process | −39.4 |
| Gene Name | Protein Name | Protein ID | Function | Fold Change |
|---|---|---|---|---|
| SAOUHSC_02971 | Neutral metalloproteinase | Q2FUX4 | Metal ion binding; protease | +7.4 |
| lytM | Glycyl-glycine endopeptidase LytM | O33599 | Cell wall biogenesis/degradation; virulence | +6.2 |
| topB | DNA topoisomerase 3 | Q2FW03 | DNA topological change | +5.2 |
| SAOUHSC_01692 | CMP/dCMP-type deaminase domain-containing protein | Q2G2A1 | Metal binding | +4.5 |
| SAOUHSC_02151 | ABC transporter permease | Q2FWX0 | ABC-type transporter activity | +4.0 |
| SAOUHSC_01244 | YlxR domain-containing protein | Q2G2D1 | Unknown | +3.6 |
| SAOUHSC_01025 | Membrane spanning protein | Q2FZH9 | Unknown | +3.5 |
| perR | Peroxide-responsive repressor PerR | Q2G282 | Transcription of antioxidant-encoding proteins (katA, trxB, bcp, ahpCF) | +3.4 |
| SAOUHSC_00671 | Probable autolysin SsaALP | Q2G0D4 | Cell wall biogenesis/degradation | +3.3 |
| SAOUHSC_00236 | 6-phospho-beta-glucosidase, putative | Q2G1A9 | Carbohydrate catabolic process | +3.1 |
| SAOUHSC_02845 | Thioredoxin domain-containing protein | Q2FV89 | Cell redox homeostasis | −4.1 |
| deoD | Purine nucleoside phosphorylase DeoD-type | Q2FWB8 | Glycosyltransferase | −4.3 |
| SAOUHSC_01874 | Rhodanese domain-containing protein | Q2FXH3 | Unknown | −4.7 |
| SAOUHSC_01710 | Acetyl-CoA carboxylase, biotin carboxyl carrier protein, putative | Q2FXX0 | Unknown | −4.9 |
| SAOUHSC_01511 | Uncharacterised protein | Q2FY81 | Unknown | −5.1 |
| SAOUHSC_01110 | Fibrinogen-binding protein | Q2FZC2 | Complement binding | −5.8 |
| SAOUHSC_00481 | S4 domain-containing protein | Q2G0R5 | RNA binding | −6.4 |
| SAOUHSC_00401 | Myeloperoxidase inhibitor SPIN | Q2G0X2 | Host immune evasion | −6.7 |
| SAOUHSC_00462 | Hydrolase TatD | Q2G1S2 | Deoxyribonuclease activity | −8.2 |
| rot | HTH-type transcriptional regulator rot | Q9RFJ6 | Negatively regulates the transcription of virulence factors (lipase, haemolysins, proteases). Positive regulation of cell surface adhesins. | −8.8 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Piatek, M.; O’Beirne, C.; Beato, Z.; Tacke, M.; Kavanagh, K. Pseudomonas aeruginosa and Staphylococcus aureus Display Differential Proteomic Responses to the Silver(I) Compound, SBC3. Antibiotics 2023, 12, 348. https://doi.org/10.3390/antibiotics12020348
Piatek M, O’Beirne C, Beato Z, Tacke M, Kavanagh K. Pseudomonas aeruginosa and Staphylococcus aureus Display Differential Proteomic Responses to the Silver(I) Compound, SBC3. Antibiotics. 2023; 12(2):348. https://doi.org/10.3390/antibiotics12020348
Chicago/Turabian StylePiatek, Magdalena, Cillian O’Beirne, Zoe Beato, Matthias Tacke, and Kevin Kavanagh. 2023. "Pseudomonas aeruginosa and Staphylococcus aureus Display Differential Proteomic Responses to the Silver(I) Compound, SBC3" Antibiotics 12, no. 2: 348. https://doi.org/10.3390/antibiotics12020348
APA StylePiatek, M., O’Beirne, C., Beato, Z., Tacke, M., & Kavanagh, K. (2023). Pseudomonas aeruginosa and Staphylococcus aureus Display Differential Proteomic Responses to the Silver(I) Compound, SBC3. Antibiotics, 12(2), 348. https://doi.org/10.3390/antibiotics12020348