
Novel Arginine- and Proline-Rich Candidacidal Peptides Obtained through a Bioinformatic Approach

 ,
,  ,
,  ,
, 
 and
and 
Abstract
:1. Introduction
2. Results
2.1. Bioinformatic Analysis
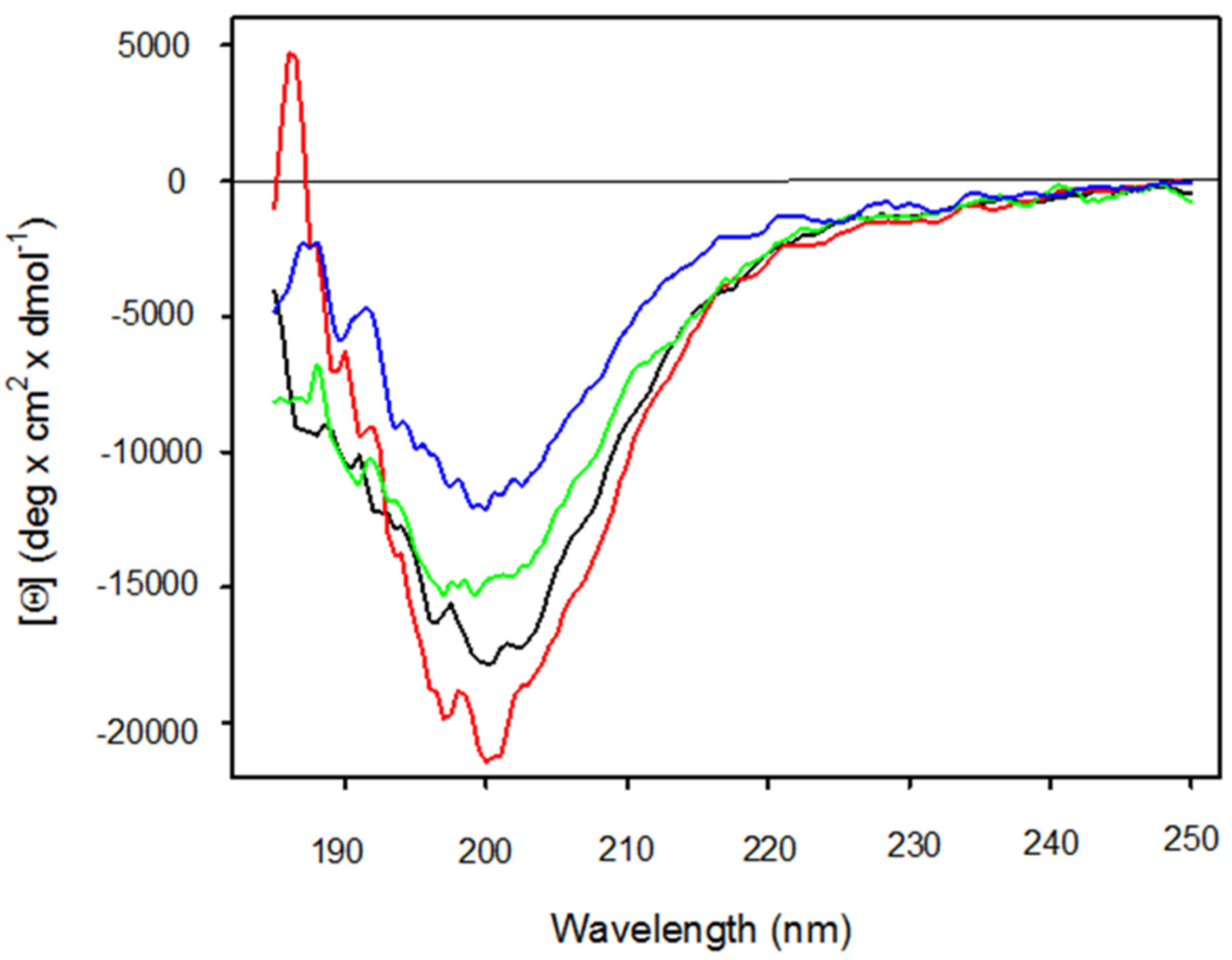
2.2. Structural Studies on Ma1 and Ma2
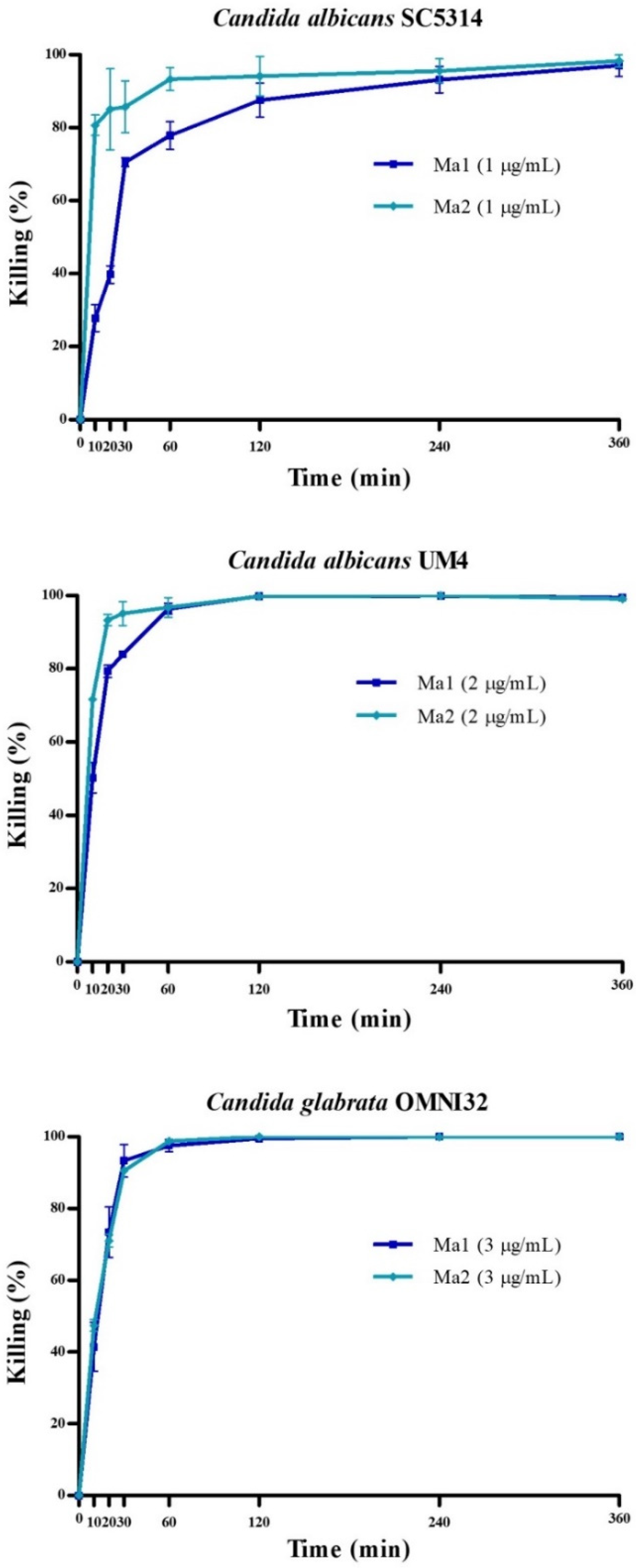
2.3. Activity of Peptides against Planktonic Yeast Cells
2.4. Activity of Peptides against Candida Strains’ Monomicrobial Biofilm
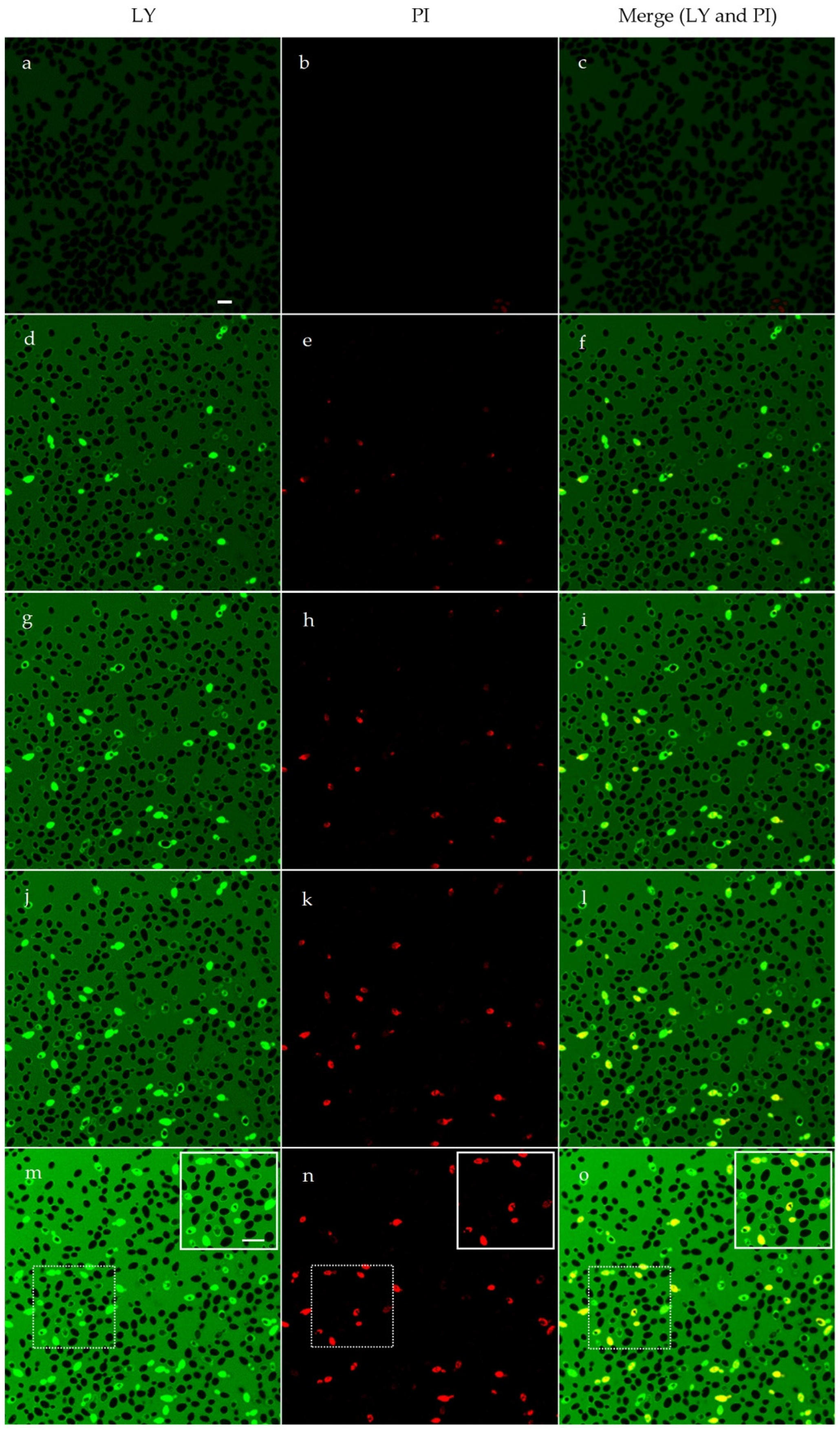
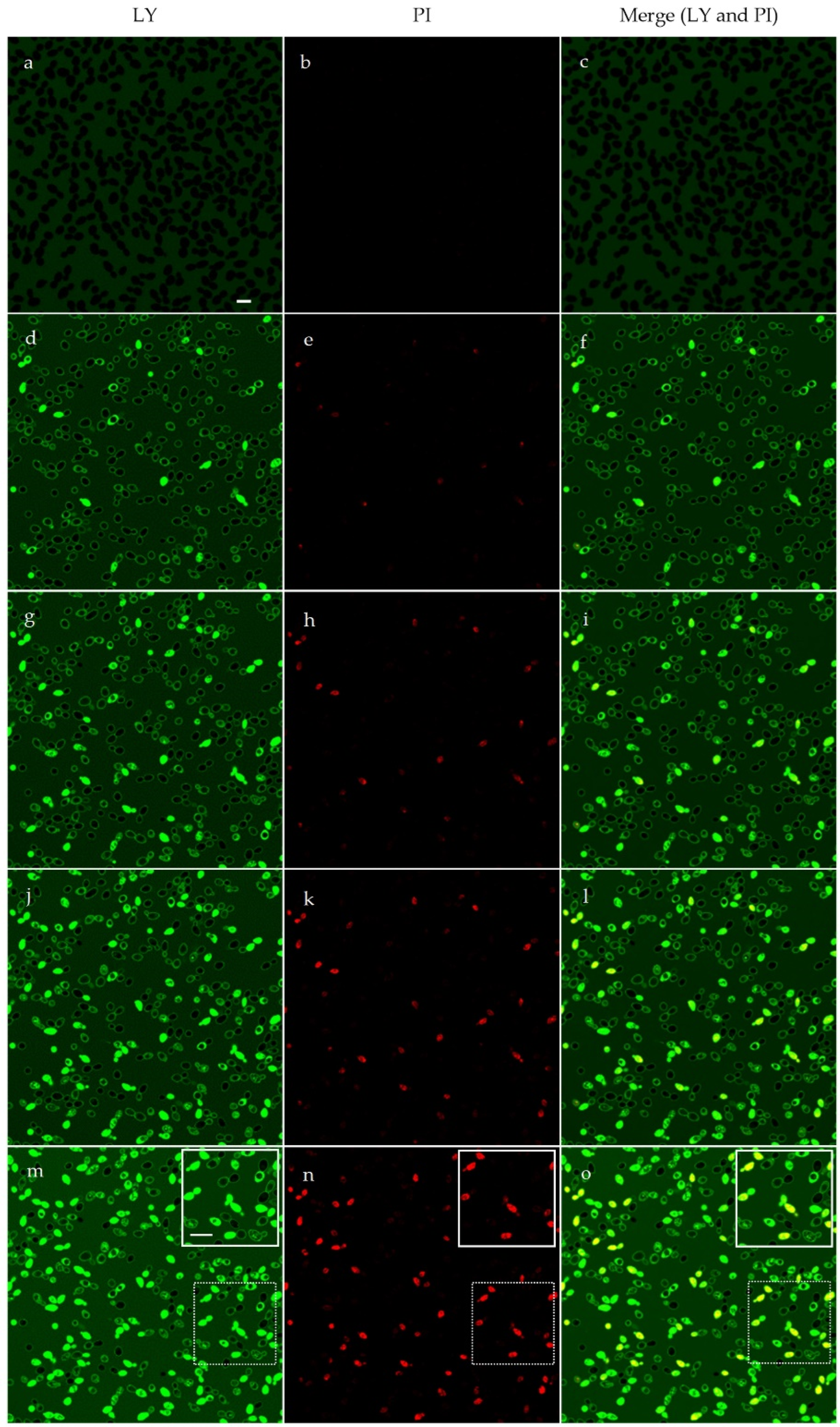
2.5. Confocal Microscopy Studies
2.6. Artificial Vesicle Assay
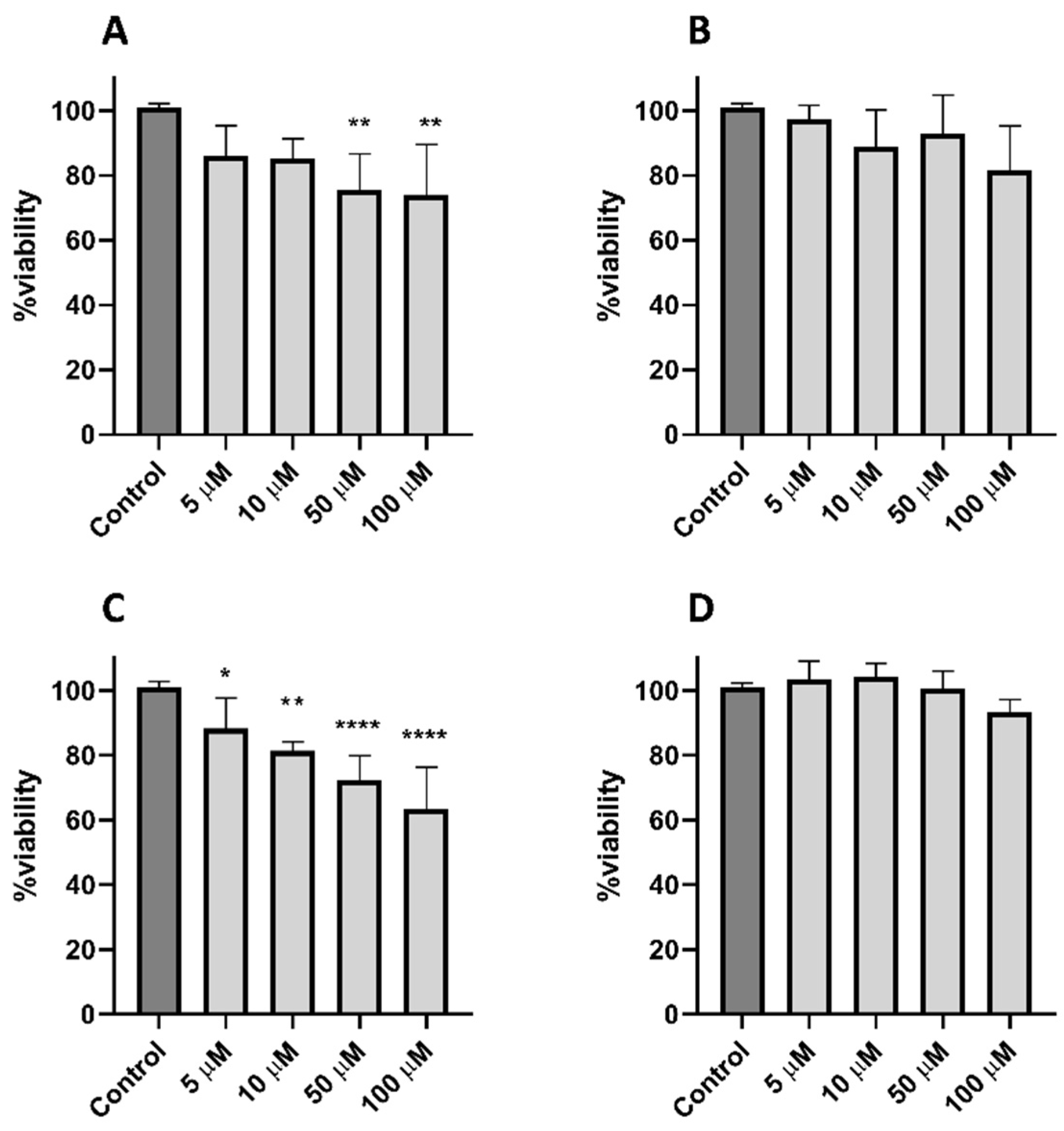
2.7. Cytotoxicity of Peptides
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Bioinformatic Analyses
4.3. Peptide Synthesis and Reversed-Phase High-Performance Liquid Chromatography (RP-HPLC) Analysis and Purification
4.4. Circular Dichroism (CD) Measurement
4.5. Evaluation of Peptides’ Activity against Planktonic Yeast Cells
4.6. Evaluation of Apoptosis Induction in Candida Cells after Treatment with Peptides
4.7. Biofilm Assays
4.7.1. Peptide Treatment on the Early Stage of Biofilm Formation
4.7.2. Peptide Treatment on Mature Biofilm
4.7.3. Biofilm Biomass Quantification
4.8. Confocal Microscopy Studies
4.9. Artificial Vescicles Assay
4.10. Cytotoxicity Tests
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fisher, M.C.; Gurr, S.J.; Cuomo, C.A.; Blehert, D.S.; Jin, H.; Stukenbrock, E.H.; Stajich, J.E.; Kahmann, R.; Boone, C.; Denning, D.W.; et al. Threats Posed by the Fungal Kingdom to Humans, Wildlife, and Agriculture. mBio 2020, 11, e00449-20. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, M.L.; Nosanchuk, J.D. Fungal diseases as neglected pathogens: A wake-up call to public health officials. PLoS Negl. Trop Dis. 2020, 14, e0007964. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jenks, J.D.; Cornely, O.A.; Chen, S.C.; Thompson, G.R., 3rd; Hoenigl, M. Breakthrough invasive fungal infections: Who is at risk? Mycoses 2020, 63, 1021–1032. [Google Scholar] [CrossRef] [PubMed]
- Cornely, O.A.; Lass-Florl, C.; Lagrou, K.; Arsic-Arsenijevic, V.; Hoenigl, M. Improving outcome of fungal diseases—Guiding experts and patients towards excellence. Mycoses 2017, 60, 420–425. [Google Scholar] [CrossRef] [Green Version]
- Fisher, M.C.; Hawkins, N.J.; Sanglard, D.; Gurr, S.J. Worldwide emergence of resistance to antifungal drugs challenges human health and food security. Science 2018, 360, 739–742. [Google Scholar] [CrossRef] [Green Version]
- Revie, N.M.; Iyer, K.R.; Robbins, N.; Cowen, L.E. Antifungal drug resistance: Evolution, mechanisms and impact. Curr. Opin. Microbiol. 2018, 45, 70–76. [Google Scholar] [CrossRef]
- Arastehfar, A.; Gabaldón, T.; Garcia-Rubio, R.; Jenks, J.D.; Hoenigl, M.; Salzer, H.J.F.; Ilkit, M.; Lass-Flörl, C.; Perlin, D.S. Drug-Resistant Fungi: An Emerging Challenge Threatening Our Limited Antifungal Armamentarium. Antibiotics 2020, 9, 877. [Google Scholar] [CrossRef]
- Wu, S.; Wang, Y.; Liu, N.; Dong, G.; Sheng, C. Tackling Fungal Resistance by Biofilm Inhibitors. J. Med. Chem. 2017, 60, 2193–2211. [Google Scholar] [CrossRef]
- Pereira, R.; dos Santos Fontenelle, R.O.; de Brito, E.H.S.; de Morais, S.M. Biofilm of Candida albicans: Formation, regulation and resistance. J. Appl. Microbiol. 2021, 131, 11–22. [Google Scholar] [CrossRef]
- Bhattacharya, S.; Sae-Tia, S.; Fries, B.C. Candidiasis and Mechanisms of Antifungal Resistance. Antibiotics 2020, 9, 312. [Google Scholar] [CrossRef]
- Browne, K.; Chakraborty, S.; Chen, R.; Willcox, M.D.; Black, D.S.; Walsh, W.R.; Kumar, N. A New Era of Antibiotics: The Clinical Potential of Antimicrobial Peptides. Int. J. Mol. Sci. 2020, 21, 7047. [Google Scholar] [CrossRef]
- Luo, X.; Chen, H.; Song, Y.; Qin, Z.; Xu, L.; He, N.; Tan, Y.; Dessie, W. Advancements, challenges and future perspectives on peptide-based drugs: Focus on antimicrobial peptides. Eur. J. Pharm. Sci. 2023, 181, 106363. [Google Scholar] [CrossRef]
- Mookherjee, N.; Anderson, M.A.; Haagsman, H.P.; Davidson, D.J. Antimicrobial host defence peptides: Functions and clinical potential. Nat. Rev. Drug Discov. 2020, 19, 311–332. [Google Scholar] [CrossRef]
- Tornesello, A.L.; Borrelli, A.; Buonaguro, L.; Buonaguro, F.M.; Tornesello, M.L. Antimicrobial Peptides as Anticancer Agents: Functional Properties and Biological Activities. Molecules 2020, 25, 2850. [Google Scholar] [CrossRef]
- Brogden, K.A. Antimicrobial peptides: Pore formers or metabolic inhibitors in bacteria? Nat. Rev. Microbiol. 2005, 3, 238–250. [Google Scholar] [CrossRef]
- Cardoso, M.H.; Meneguetti, B.T.; Costa, B.O.; Buccini, D.F.; Oshiro, K.G.N.; Preza, S.L.E.; Carvalho, C.M.E.; Migliolo, L.; Franco, O.L. Non-Lytic Antibacterial Peptides That Translocate Through Bacterial Membranes to Act on Intracellular Targets. Int. J. Mol. Sci. 2019, 20, 4877. [Google Scholar] [CrossRef] [Green Version]
- Mishra, A.K.; Choi, J.; Moon, E.; Baek, K.-H. Tryptophan-Rich and Proline-Rich Antimicrobial Peptides. Molecules 2018, 23, 815. [Google Scholar] [CrossRef] [Green Version]
- Welch, N.G.; Li, W.; Hossain, M.A.; Separovic, F.; O’Brien-Simpson, N.M.; Wade, J.D. (Re)Defining the Proline-Rich Antimicrobial Peptide Family and the Identification of Putative New Members. Front. Chem. 2020, 8, 607769. [Google Scholar] [CrossRef]
- van der Weerden, N.L.; Bleackley, M.R.; Anderson, M.A. Properties and mechanisms of action of naturally occurring antifungal peptides. Cell Mol. Life Sci. 2013, 70, 3545–3570. [Google Scholar] [CrossRef]
- Ciociola, T.; Giovati, L.; Conti, S.; Magliani, W.; Santinoli, C.; Polonelli, L. Natural and synthetic peptides with antifungal activity. Future Med. Chem. 2016, 8, 1413–1433. [Google Scholar] [CrossRef]
- Mizuhara, N.; Kuroda, M.; Ogita, A.; Tanaka, T.; Usuki, Y.; Fujita, K. Antifungal thiopeptide cyclothiazomycin B1 exhibits growth inhibition accompanying morphological changes via binding to fungal cell wall chitin. Bioorg. Med. Chem. 2011, 19, 5300–5310. [Google Scholar] [CrossRef]
- Delattin, N.; Brucker, K.; Cremer, K.; Cammue, B.P.; Thevissen, K. Antimicrobial Peptides as a Strategy to Combat Fungal Biofilms. Curr. Top. Med. Chem. 2017, 17, 604–612. [Google Scholar] [CrossRef] [PubMed]
- Oshiro, K.G.N.; Rodrigues, G.; Monges, B.E.D.; Cardoso, M.H.; Franco, O.L. Bioactive Peptides Against Fungal Biofilms. Front. Microbiol. 2019, 10, 2169. [Google Scholar] [CrossRef] [PubMed]
- Hancock, R.E.W.; Alford, M.A.; Haney, E.F. Antibiofilm activity of host defence peptides: Complexity provides opportunities. Nat. Rev. Microbiol. 2021, 19, 786–797. [Google Scholar] [CrossRef] [PubMed]
- Parthasarathy, A.; Borrego, E.J.; Savka, M.A.; Dobson, R.C.J.; Hudson, A.O. Amino acid-derived defense metabolites from plants: A potential source to facilitate novel antimicrobial development. J. Biol. Chem. 2021, 296, 100438. [Google Scholar] [CrossRef]
- Lima, A.M.; Azevedo, M.I.G.; Sousa, L.M.; Oliveira, N.S.; Andrade, C.R.; Freitas, C.D.T.; Souza, P.F.N. Plant antimicrobial peptides: An overview about classification, toxicity and clinical applications. Int. J. Biol. Macromol. 2022, 214, 10–21. [Google Scholar] [CrossRef]
- Sharma, P.; Kaur, J.; Sharma, G.; Kashyap, P. Plant derived antimicrobial peptides: Mechanism of target, isolation techniques, sources and pharmaceutical applications. J. Food Biochem. 2022, 46, e14348. [Google Scholar] [CrossRef]
- Srivastava, S.; Dashora, K.; Ameta, K.L.; Singh, N.P.; El-Enshasy, H.A.; Pagano, M.C.; Hesham, A.E.-L.; Sharma, G.D.; Sharma, M.; Bhargava, A. Cysteine-rich antimicrobial peptides from plants: The future of antimicrobial therapy. Phytother. Res. 2021, 35, 256–277. [Google Scholar] [CrossRef]
- Cao, H.; Ke, T.; Liu, R.; Yu, J.; Dong, C.; Cheng, M.; Huang, J.; Liu, S. Identification of a Novel Proline-Rich Antimicrobial Peptide from Brassica napus. PLoS ONE 2015, 10, e0137414. [Google Scholar] [CrossRef] [Green Version]
- Agerberth, B.; Lee, J.Y.; Bergman, T.; Carlquist, M.; Boman, H.G.; Mutt, V.; Jörnvall, H. Amino acid sequence of PR-39. Isolation from pig intestine of a new member of the family of proline-arginine-rich antibacterial peptides. Eur. J. Biochem. 1991, 202, 849–854. [Google Scholar] [CrossRef]
- Sharma, R.; Shrivastava, S.; Kumar Singh, S.; Kumar, A.; Saxena, S.; Kumar Singh, R. Deep-AFPpred: Identifying novel antifungal peptides using pretrained embeddings from seq2vec with 1DCNN-BiLSTM. Brief. Bioinform. 2022, 23, bbab422. [Google Scholar] [CrossRef]
- Ma, K.; Kan, L.; Wang, K. Polyproline II helix is a key structural motif of the elastic PEVK segment of titin. Biochemistry 2001, 40, 3427–3438. [Google Scholar] [CrossRef]
- Perczel, A.; Hollosi, M. Circular Dichroism and the Conformational Analysis of Biomolecules; Fasman, G.D., Ed.; Springer: Boston, MA, USA, 1996. [Google Scholar]
- Guha, S.; Ferrie, R.P.; Ghimire, J.; Ventura, C.R.; Wu, E.; Sun, L.; Kim, S.Y.; Wiedman, G.R.; Hristova, K.; Wimley, W.C. Applications and evolution of melittin, the quintessential membrane active peptide. Biochem. Pharmacol. 2021, 193, 114769. [Google Scholar] [CrossRef]
- Graf, M.; Wilson, D.N. Intracellular Antimicrobial Peptides Targeting the Protein Synthesis Machinery. In Antimicrobial Peptides; Advances in Experimental Medicine and Biology; Matsuzaki, K., Ed.; Springer: Singapore, 2019; Volume 1117, pp. 73–89. [Google Scholar]
- Kopeikin, P.M.; Zharkova, M.S.; Kolobov, A.A.; Smirnova, M.P.; Sukhareva, M.S.; Umnyakova, E.S.; Kokryakov, V.N.; Orlov, D.S.; Milman, B.L.; Balandin, S.V.; et al. Caprine Bactenecins as Promising Tools for Developing New Antimicrobial and Antitumor Drugs. Front. Cell Infect. Microbiol. 2020, 10, 552905. [Google Scholar] [CrossRef]
- Paulsen, V.S.; Blencke, H.-M.; Benincasa, M.; Haug, T.; Eksteen, J.J.; Styrvold, O.B.; Scocchi, M.; Stensvåg, K. Structure-Activity Relationships of the Antimicrobial Peptide Arasin 1—And Mode of Action Studies of the N-Terminal, Proline-Rich Region. PLoS ONE 2013, 8, e53326. [Google Scholar] [CrossRef] [Green Version]
- Podda, E.; Benincasa, M.; Pacor, S.; Micali, F.; Mattiuzzo, M.; Gennaro, R.; Scocchi, M. Dual mode of action of Bac7, a proline-rich antibacterial peptide. Biochim. Biophys Acta 2006, 1760, 1732–1740. [Google Scholar] [CrossRef]
- Cabras, T.; Longhi, R.; Secundo, F.; Nocca, G.; Conti, S.; Polonelli, L.; Fanali, C.; Inzitari, R.; Petruzzelli, R.; Messana, I.; et al. Structural and functional characterization of the porcine proline–rich antifungal peptide SP-B isolated from salivary gland granules. J. Pept. Sci. 2008, 14, 251–260. [Google Scholar] [CrossRef]
- Gioia, M.; Fasciglione, G.F.; Sbardella, D.; Sciandra, F.; Casella, M.; Camerini, S.; Crescenzi, M.; Gori, A.; Tarantino, U.; Cozza, P.; et al. The enzymatic processing of α-dystroglycan by MMP-2 is controlled by two anchoring sites distinct from the active site. PLoS ONE 2018, 13, e0192651. [Google Scholar] [CrossRef] [Green Version]
- Polonelli, L.; Ciociola, T.; Elviri, L.; Zanello, P.P.; Giovati, L.; Arruda, D.C.; Muñoz, J.E.; Mortara, R.A.; Morace, G.; Borghi, E.; et al. A Naturally Occurring Antibody Fragment Neutralizes Infectivity of Diverse Infectious Agents. Sci. Rep. 2016, 6, 35018. [Google Scholar] [CrossRef] [Green Version]
- Polonelli, L.; Ciociola, T.; Magliani, W.; Zanello, P.P.; D’Adda, T.; Galati, S.; De Bernardis, F.; Arancia, S.; Gabrielli, E.; Pericolini, E.; et al. Peptides of the Constant Region of Antibodies Display Fungicidal Activity. PLoS ONE 2012, 7, e34105. [Google Scholar] [CrossRef] [Green Version]
- Ciociola, T.; Pertinhez, T.A.; Giovati, L.; Sperinde, M.; Magliani, W.; Ferrari, E.; Gatti, R.; D’Adda, T.; Spisni, A.; Conti, S.; et al. Dissecting the Structure-Function Relationship of a Fungicidal Peptide Derived from the Constant Region of Human Immunoglobulins. Antimicrob Agents Chemother. 2016, 60, 2435–2442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Fermo, P.; Ciociola, T.; Di Lodovico, S.; D’Ercole, S.; Petrini, M.; Giovati, L.; Conti, S.; Di Giulio, M.; Cellini, L. Antimicrobial Peptide L18R Displays a Modulating Action against Inter-Kingdom Biofilms in the Lubbock Chronic Wound Biofilm Model. Microorganisms 2021, 9, 1779. [Google Scholar] [CrossRef] [PubMed]
- Kolusheva, S.; Shahal, T.; Jelinek, R. Peptide-membrane interactions studied by a new phospholipid/polydiacetylene colorimetric vesicle assay. Biochemistry 2000, 39, 15851–15859. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Candida Strains | EC50 (95% Confidence Intervals) [mol/L] | |
|---|---|---|
| Ma1 | Ma2 | |
| C. albicans SC5314 | 1.241 (1.149–1.341) × 10−7 | 1.079 (0.959–1.214) × 10−7 |
| C. albicans UM4 | 1.532 (1.462–1.605) × 10−7 | 2.038 (1.682–2.468) × 10−7 |
| C. glabrata OMNI32 | 4.160 (2.541–6.812) × 10−7 | 3.907 (3.350–4.558) × 10−7 |
| Candida Strains | EC50 (95% Confidence Intervals) [mol/L] | |
|---|---|---|
| Ma1 | Ma2 | |
| C. albicans SC5314 | 2.842 (2.677–3.018) × 10−6 | 0.554 (0.373–0.823) × 10−6 |
| C. albicans UM4 | 0.880 (0.809–0.956) × 10−6 | 0.229 (0.220–0.239) × 10−6 |
| C. glabrata OMNI32 | 1.968 (1.504–2.577) × 10−6 | 1.181 (1.021–1.367) × 10−6 |
| Candida Strains | % Biomass Reduction at 50 μg/mL | |
|---|---|---|
| Ma1 | Ma2 | |
| C. albicans SC5314 | 11.64 ± 0.19 | 57.31 ± 0.79 |
| C. albicans UM4 | 72.19 ± 0.63 | 54.44 ± 2.46 |
| C. glabrata OMNI32 | 49.84 ± 0.10 | 71.92 ± 3.55 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ciociola, T.; Giovati, L.; De Simone, T.; Bergamaschi, G.; Gori, A.; Consalvi, V.; Conti, S.; Vitali, A. Novel Arginine- and Proline-Rich Candidacidal Peptides Obtained through a Bioinformatic Approach. Antibiotics 2023, 12, 472. https://doi.org/10.3390/antibiotics12030472
Ciociola T, Giovati L, De Simone T, Bergamaschi G, Gori A, Consalvi V, Conti S, Vitali A. Novel Arginine- and Proline-Rich Candidacidal Peptides Obtained through a Bioinformatic Approach. Antibiotics. 2023; 12(3):472. https://doi.org/10.3390/antibiotics12030472
Chicago/Turabian StyleCiociola, Tecla, Laura Giovati, Tiziano De Simone, Greta Bergamaschi, Alessandro Gori, Valerio Consalvi, Stefania Conti, and Alberto Vitali. 2023. "Novel Arginine- and Proline-Rich Candidacidal Peptides Obtained through a Bioinformatic Approach" Antibiotics 12, no. 3: 472. https://doi.org/10.3390/antibiotics12030472
APA StyleCiociola, T., Giovati, L., De Simone, T., Bergamaschi, G., Gori, A., Consalvi, V., Conti, S., & Vitali, A. (2023). Novel Arginine- and Proline-Rich Candidacidal Peptides Obtained through a Bioinformatic Approach. Antibiotics, 12(3), 472. https://doi.org/10.3390/antibiotics12030472