
Efflux, Signaling and Warfare in a Polymicrobial World

Abstract
:1. Introduction
2. Ecological Role of Efflux Pumps
2.1. Efflux Pumps, Homeostasis and Stress Management
2.2. Efflux Pumps and Signalling
2.3. Efflux Pumps, Antibiotic Stress, and Outer Membrane Vesicles
2.4. Efflux Pumps and Host Colonisation
3. Efflux Pump Regulation
3.1. RND Efflux Pumps in Pseudomonas aeruginosa and Their Regulation
3.2. MFS Efflux Pumps in Staphylococcus aureus and Their Regulation
3.3. Cell-Cell Communication, Sensing the Qourum, and Efflux Regulation
4. The Microbial Ecology of Efflux Inhibition and Sources for Exploration
4.1. On the Ecological Basis of Efflux Inhibition
4.2. Plants as a Source for Efflux Inhibitors
4.3. Marine Efflux Inhibitors
4.4. Synthetic Efflux Inhibitors
4.5. Phage Steering
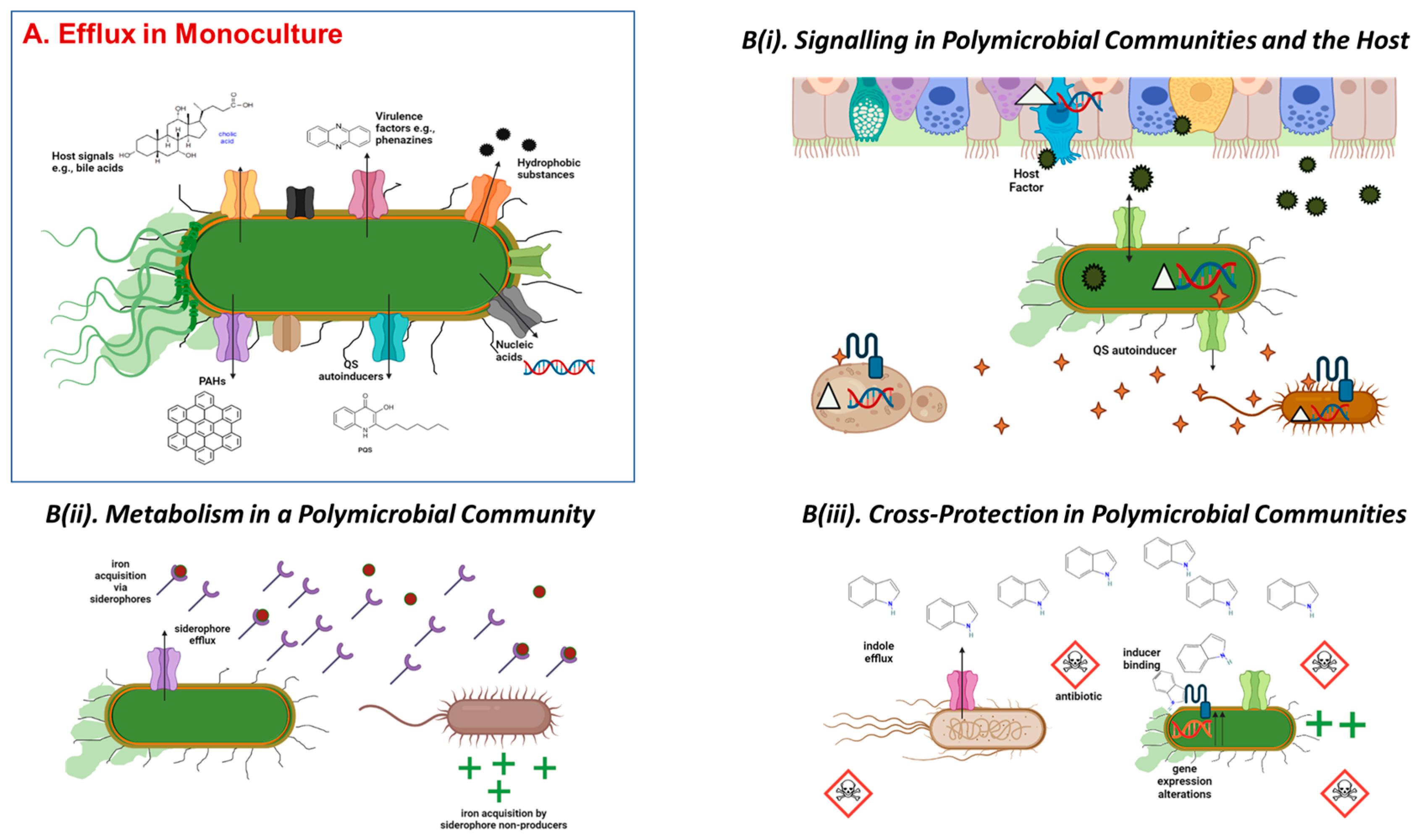
5. Efflux Pumps and Polymicrobial Communities
Efflux Inhibition in the Polymicrobial Quorate
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Henderson, P.J.F.; Maher, C.; Elbourne, L.D.H.; Eijkelkamp, B.A.; Paulsen, I.T.; Hassan, K.A. Physiological Functions of Bacterial “Multidrug” Efflux Pumps. Chem. Rev. 2021, 121, 5417–5478. [Google Scholar] [CrossRef]
- Teelucksingh, T.; Thompson, L.K.; Cox, G. The Evolutionary Conservation of Escherichia coli Drug Efflux Pumps Supports Physiological Functions. J. Bacteriol. 2020, 202, e00367-20. [Google Scholar] [CrossRef] [PubMed]
- Cooper, M.A.; Shlaes, D. Fix the antibiotics pipeline. Nature 2011, 472, 32. [Google Scholar] [CrossRef] [PubMed]
- Available online: https://www.who.int/news/item/09-12-2022-report-signals-increasing-resistance-to-antibiotics-in-bacterial-infections-in-humans-and-need-for-better-data (accessed on 13 March 2023).
- Ruekit, S.; Srijan, A.; Serichantalergs, O.; Margulieux, K.R.; Mc Gann, P.; Mills, E.G.; Stribling, W.C.; Pimsawat, T.; Kormanee, R.; Nakornchai, S.; et al. Molecular characterization of multidrug-resistant ESKAPEE pathogens from clinical samples in Chonburi, Thailand (2017–2018). BMC Infect. Dis. 2022, 22, 695. [Google Scholar] [CrossRef]
- Available online: https://www.who.int/news-room/fact-sheets/detail/antimicrobial-resistance (accessed on 13 March 2023).
- Rahman, T.; Yarnall, B.; Doyle, D.A. Efflux drug transporters at the forefront of antimicrobial resistance. Eur. Biophys. J. 2017, 46, 647–653. [Google Scholar] [CrossRef] [PubMed]
- Blanco, P.; Hernando-Amado, S.; Reales-Calderon, J.A.; Corona, F.; Lira, F.; Alcalde-Rico, M.; Bernardini, A.; Sanchez, M.B.; Martinez, J.L. Bacterial Multidrug Efflux Pumps: Much More Than Antibiotic Resistance Determinants. Microorganisms 2016, 4, 14. [Google Scholar] [CrossRef] [Green Version]
- Alav, I.; Kobylka, J.; Kuth, M.S.; Pos, K.M.; Picard, M.; Blair, J.M.A.; Bavro, V.N. Structure, Assembly, and Function of Tripartite Efflux and Type 1 Secretion Systems in Gram-Negative Bacteria. Chem. Rev. 2021, 121, 5479–5596. [Google Scholar] [CrossRef]
- Nikaido, H.; Pages, J.M. Broad-specificity efflux pumps and their role in multidrug resistance of Gram-negative bacteria. FEMS Microbiol. Rev. 2012, 36, 340–363. [Google Scholar] [CrossRef] [Green Version]
- Greene, N.P.; Kaplan, E.; Crow, A.; Koronakis, V. Antibiotic resistance mediated by the MacB ABC transporter family: A structural and functional perspective. Front. Microbiol. 2018, 9, 950. [Google Scholar] [CrossRef] [Green Version]
- Dong, N.; Zeng, Y.; Wang, Y.; Liu, C.; Lu, J.; Cai, C.; Liu, X.; Chen, Y.; Chen, Y.; Fang, Y. Distribution and spread of the mobilised RND efflux pump gene cluster tmexCD-topRJ in clinical Gram-negative bacteria: A molecular epidemiological study. Lancet Microbe 2022, 3, e846–e856. [Google Scholar] [CrossRef]
- Aguilar-Rodea, P.; Zúñiga, G.; Cerritos, R.; Rodríguez-Espino, B.A.; Gomez-Ramirez, U.; Nolasco-Romero, C.G.; López-Marceliano, B.; Rodea, G.E.; Mendoza-Elizalde, S.; Reyes-López, A. Nucleotide substitutions in the mexR, nalC and nalD regulator genes of the MexAB-OprM efflux pump are maintained in Pseudomonas aeruginosa genetic lineages. PLoS ONE 2022, 17, e0266742. [Google Scholar] [CrossRef]
- Huang, L.; Wu, C.; Gao, H.; Xu, C.; Dai, M.; Huang, L.; Hao, H.; Wang, X.; Cheng, G. Bacterial multidrug efflux pumps at the frontline of antimicrobial resistance: An overview. Antibiotics 2022, 11, 520. [Google Scholar] [CrossRef]
- Pasqua, M.; Grossi, M.; Scinicariello, S.; Aussel, L.; Barras, F.; Colonna, B.; Prosseda, G. The MFS efflux pump EmrKY contributes to the survival of Shigella within macrophages. Sci. Rep. 2019, 9, 2906. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Handzlik, J.; Matys, A.; Kieć-Kononowicz, K. Recent advances in multi-drug resistance (MDR) efflux pump inhibitors of Gram-positive bacteria S. aureus. Antibiotics 2013, 2, 28–45. [Google Scholar] [CrossRef] [Green Version]
- Hassan, K.A.; Liu, Q.; Elbourne, L.D.H.; Ahmad, I.; Sharples, D.; Naidu, V.; Chan, C.L.; Li, L.; Harborne, S.P.D.; Pokhrel, A. Pacing across the membrane: The novel PACE family of efflux pumps is widespread in Gram-negative pathogens. Res. Microbiol. 2018, 169, 450–454. [Google Scholar] [CrossRef] [PubMed]
- Delmar, J.A.; Yu, E.W. The AbgT family: A novel class of antimetabolite transporters. Prot. Sci. 2016, 25, 322–337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pasqua, M.; Grossi, M.; Zennaro, A.; Fanelli, G.; Micheli, G.; Barras, F.; Colonna, B.; Prosseda, G. The Varied Role of Efflux Pumps of the MFS Family in the Interplay of Bacteria with Animal and Plant Cells. Microorganisms 2019, 7, 285. [Google Scholar] [CrossRef] [Green Version]
- Pacheco, J.O.; Alvarez-Ortega, C.; Rico, M.A.; Martínez, J.L. Metabolic Compensation of Fitness Costs Is a General Outcome for Antibiotic-Resistant Pseudomonas aeruginosa Mutants Overexpressing Efflux Pumps. mBio 2017, 8, e00500-17. [Google Scholar]
- Nishino, K.; Yamasaki, S.; Nakashima, R.; Zwama, M.; Hayashi-Nishino, M. Function and Inhibitory Mechanisms of Multidrug Efflux Pumps. Front. Microbiol. 2021, 12, 737288. [Google Scholar] [CrossRef] [PubMed]
- Rojo-Molinero, E.; Macia, M.D.; Oliver, A. Social Behavior of Antibiotic Resistant Mutants Within Pseudomonas aeruginosa Biofilm Communities. Front. Microbiol. 2019, 10, 570. [Google Scholar] [CrossRef]
- Lee, A.; Mao, W.; Warren, M.S.; Mistry, A.; Hoshino, K.; Okumura, R.; Ishida, H.; Lomovskaya, O. Interplay between efflux pumps may provide either additive or multiplicative effects on drug resistance. J. Bacteriol. 2000, 182, 3142–3150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leventhal, G.E.; Ackermann, M.; Schiessl, K.T. Why microbes secrete molecules to modify their environment: The case of iron-chelating siderophores. J. R. Soc. Interface 2019, 16, 20180674. [Google Scholar] [CrossRef]
- Woolridge, D.P.; Vazquez-Laslop, N.; Markham, P.N.; Chevalier, M.S.; Gerner, E.W.; Neyfakh, A.A. Efflux of the natural polyamine spermidine facilitated by the Bacillus subtilis multidrug transporter Blt. J. Biol. Chem. 1997, 272, 8864–8866. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fleeman, R.M.; Debevec, G.; Antonen, K.; Adams, J.L.; Santos, R.G.; Welmaker, G.S.; Houghten, R.A.; Giulianotti, M.A.; Shaw, L.N. Identification of a Novel Polyamine Scaffold With Potent Efflux Pump Inhibition Activity Toward Multi-Drug Resistant Bacterial Pathogens. Front. Microbiol. 2018, 9, 1301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hassan, K.A.; Naidu, V.; Edgerton, J.R.; Mettrick, K.A.; Liu, Q.; Fahmy, L.; Lip, L.; Jackson, S.M.; Ahmad, I.; Sharples, D.; et al. Short-chain diamines are the physiological substrates of PACE family efflux pumps. Proc. Natl. Acad. Sci. USA 2019, 116, 18015–18020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, B.; Pachaiyappan, B.; Gruber, J.D.; Schmidt, M.G.; Zhang, Y.-M.; Woster, P.M. Antibacterial Diamines Targeting Bacterial Membranes. J. Med. Chem. 2016, 59, 3140–3151. [Google Scholar] [CrossRef] [Green Version]
- Mukherjee, S.; Bossier, B.L. Bacterial quorum sensing in complex and dynamically changing environments. Nat. Rev. Microbiol. 2019, 17, 371–382. [Google Scholar] [CrossRef]
- O’Brien, S.; Fothergill, J.L. The role of multispecies social interactions in shaping Pseudomonas aeruginosa pathogenicity in the cystic fibrosis lung. FEMS Microbiol. Lett. 2017, 364, fnx128. [Google Scholar] [CrossRef] [Green Version]
- Reen, F.J.; McGlacken, G.P.; O’Gara, F. The expanding horizon of alkyl quinolone signalling and communication in polycellular interactomes. FEMS Microbiol. Lett. 2018, 365, fny076. [Google Scholar] [CrossRef] [Green Version]
- Whiteley, M.; Diggle, S.P.; Greenberg, E.P. Progress in and promise of bacterial quorum sensing research. Nature 2017, 551, 313–320. [Google Scholar] [CrossRef] [Green Version]
- Kaur, B.; Gupta, J.; Sharma, S.; Sharma, D.; Sharma, S. Focused review on dual inhibition of quorum sensing and efflux pumps: A potential way to combat multi drug resistant Staphylococcus aureus infections. Intern. J. Biol. Macromol. 2021, 190, 33–43. [Google Scholar] [CrossRef] [PubMed]
- Alcalde-Rico, M.; Olivares-Pacheco, J.; Alvarez-Ortega, C.; Cámara, M.; Martínez, J.L. Role of the Multidrug Resistance Efflux Pump MexCD-OprJ in the Pseudomonas aeruginosa Quorum Sensing Response. Front. Microbiol. 2018, 9, 2752. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olivares, J.; Alvarez-Ortega, C.; Linares, J.F.; Rojo, F.; Köhler, T.; Martínez, J.L. Overproduction of the multidrug efflux pump MexEF-OprN does not impair Pseudomonas aeruginosa fitness in competition tests, but produces specific changes in bacterial regulatory networks. Environ. Microbiol. 2012, 14, 1968–1981. [Google Scholar] [CrossRef] [PubMed]
- Alcalde-Rico, M.; Olivares-Pacheco, J.; Halliday, N.; Cámara, M.; Martínez, J.L. The impaired quorum sensing response of Pseudomonas aeruginosa MexAB-OprM efflux pump overexpressing mutants is not due to non-physiological efflux of 3-oxo-C12-HSL. Environ. Microbiol. 2020, 22, 5167–5188. [Google Scholar] [CrossRef]
- Pearson, J.P.; Pesci, E.C.; Iglewski, B.H. Roles of Pseudomonas aeruginosa las and rhl quorum-sensing systems in control of elastase and rhamnolipid biosynthesis genes. J. Bacteriol. 1997, 179, 5756–5767. [Google Scholar] [CrossRef] [Green Version]
- Legendre, C.; Reen, F.J.; Mooij, M.J.; McGlacken, G.P.; Adams, C.; O’Gara, F. Pseudomonas aeruginosa Alkyl Quinolones Repress Hypoxia-Inducible Factor 1 (HIF-1) Signaling through HIF-1 alpha Degradation. Infect. Immun. 2012, 80, 3985–3992. [Google Scholar] [CrossRef] [Green Version]
- Reen, F.J.; Mooij, M.J.; Holcombe, L.J.; McSweeney, C.M.; McGlacken, G.P.; Morrissey, J.P.; O’Gara, F. The Pseudomonas quinolone signal (PQS), and its precursor HHQ, modulate interspecies and interkingdom behaviour. FEMS Microbiol. Ecol. 2011, 77, 413–428. [Google Scholar] [CrossRef] [Green Version]
- Alav, I.; Sutton, J.M.; Rahman, K.M. Role of bacterial efflux pumps in biofilm formation. J. Antimicrob. Chemother. 2018, 73, 2003–2020. [Google Scholar] [CrossRef] [Green Version]
- Penesyan, A.; Paulsen, I.T.; Kjelleberg, S.; Gillings, M.R. Three faces of biofilms: A microbial lifestyle, a nascent multicellular organism, and an incubator for diversity. NPJ Biofilms Microbiomes 2021, 7, 80. [Google Scholar] [CrossRef]
- Cecil, J.D.; Sirisaengtaksin, N.; O’Brien-Simpson, N.M.; Krachler, A.M. Outer Membrane Vesicle-Host Cell Interactions. Microbiol. Spectr. 2019, 7, 7. [Google Scholar] [CrossRef] [Green Version]
- Jan, A.T. Outer Membrane Vesicles (OMVs) of Gram-negative Bacteria: A Perspective Update. Front. Microbiol. 2017, 8, 1053. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.; Zhang, Q.; Li, W.; Yuan, M.; Zhou, J.; Hua, L.; Chen, Y.; Ye, C.; Ma, Y. Development of novel nanoantibiotics using an outer membrane vesicle-based drug efflux mechanism. J. Control. Release 2020, 317, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Furuyama, N.; Sircili, M.P. Outer Membrane Vesicles (OMVs) Produced by Gram-Negative Bacteria: Structure, Functions, Biogenesis, and Vaccine Application. BioMed Res. Intern. 2021, 2021, 1490732. [Google Scholar] [CrossRef] [PubMed]
- Kulp, A.; Kuehn, M.J. Biological Functions and Biogenesis of Secreted Bacterial Outer Membrane Vesicles. Ann. Rev. Microbiol. 2010, 64, 163–184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagakubo, T.; Nomura, N.; Toyofuku, M. Cracking Open Bacterial Membrane Vesicles. Front. Microbiol. 2020, 10, 3026. [Google Scholar] [CrossRef] [Green Version]
- Toyofuku, M.; Morinaga, K.; Hashimoto, Y.; Uhl, J.; Shimamura, H.; Inaba, H.; Schmitt-Kopplin, P.; Eberl, L.; Nomura, N. Membrane vesicle-mediated bacterial communication. ISME J. 2017, 11, 1504–1509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaparakis-Liaskos, M.; Ferrero, R.L. Immune modulation by bacterial outer membrane vesicles. Nat. Rev. Immunol. 2015, 15, 375–387. [Google Scholar] [CrossRef]
- Barber, O.M.; Miramontes, I.M.; Jain, M.; Ozer, E.; Hartmann, E.M. The Future of Bacteriophage Therapy Will Promote Antimicrobial Susceptibility. MSystems 2021, 6, e0021821. [Google Scholar] [CrossRef]
- Schaible, B.; Schaffer, K.; Taylor, C.T. Hypoxia, innate immunity and infection in the lung. Resp. Physiol. Neurobiol. 2010, 174, 235–243. [Google Scholar] [CrossRef] [Green Version]
- Page, L.K.; Staples, K.J.; Spalluto, C.M.; Watson, A.; Wilkinson, T.M.A. Influence of Hypoxia on the Epithelial-Pathogen Interactions in the Lung: Implications for Respiratory Disease. Front. Immunol. 2021, 12, 653969. [Google Scholar] [CrossRef]
- Schaible, B.; Taylor, C.T.; Schaffer, C.T. Hypoxia Increases Antibiotic Resistance in Pseudomonas aeruginosa through Altering the Composition of Multidrug Efflux Pumps. Antimicrob. Agents Chemother. 2012, 56, 2114–2118. [Google Scholar] [CrossRef] [Green Version]
- Toyofuku, M.; Uchiyama, H.; Nomura, N. Social Behaviours under Anaerobic Conditions in Pseudomonas aeruginosa. Int. J. Microbiol. 2012, 2012, 405191. [Google Scholar] [CrossRef] [Green Version]
- Comerford, K.M.; Wallace, T.J.; Karhausen, J.; Louis, N.A.; Montalto, M.C.; Colgan, S.P. Hypoxia-inducible factor-1-dependent regulation of the multidrug resistance (MDR1) gene. Cancer Res. 2002, 62, 3387–3394. [Google Scholar]
- Black, P.A.; Warren, R.M.; Louw, G.E.; van Helden, P.D.; Victor, T.C.; Kana, B.D. Energy metabolism and drug efflux in Mycobacterium tuberculosis. Antimicrob. Agents Chemother. 2014, 58, 2491–2503. [Google Scholar] [CrossRef] [Green Version]
- Mukodani, J.; Ishikawa, Y.; Fukuzaki, H. Effects of Hypoxia on Sterol Synthesis, Acyl-Coa—Cholesterol Acyltransferase Activity, and Efflux of Cholesterol in Cultured Rabbit Skin Fibroblasts. Arteriosclerosis 1990, 10, 106–110. [Google Scholar] [CrossRef] [Green Version]
- Wang, D.; Wang, Q.; Yan, G.; Qiao, Y.; Zhu, B.; Liu, B.; Tang, C. Hypoxia induces lactate secretion and glycolytic efflux by downregulating mitochondrial pyruvate carrier levels in human umbilical vein endothelial cells. Mol. Med. Rep. 2018, 18, 1710–1717. [Google Scholar] [CrossRef]
- Vleugels, A.; Carmeliet, E. Hypoxia Increases Potassium Efflux from Mammalian Myocardium. Experientia 1976, 32, 483–484. [Google Scholar] [CrossRef]
- McGettrick, A.F.; O’Neill, L.A.J. The Role of HIF in Immunity and Inflammation. Cell. Metabol. 2020, 32, 524–536. [Google Scholar] [CrossRef]
- Al-Momani, H.; Perry, A.; Nelson, A.; Stewart, C.J.; Jones, R.; Krishnan, A.; Robertson, A.; Bourke, S.; Doe, S.; Cummings, S. Exposure to bile and gastric juice can impact the aerodigestive microbiome in people with cystic fibrosis. Sci. Rep. 2022, 12, 11114. [Google Scholar] [CrossRef]
- Flynn, S.; Reen, F.J.; Caparrós-Martín, J.A.; Woods, D.F.; Peplies, J.; Ranganathan, S.C.; Stick, S.M.; O’Gara, F. Bile Acid Signal Molecules Associate Temporally with Respiratory Inflammation and Microbiome Signatures in Clinically Stable Cystic Fibrosis Patients. Microorganisms 2020, 8, 1741. [Google Scholar] [CrossRef]
- Legendre, C.; Reen, F.J.; Woods, D.F.; Mooij, M.J.; Adams, C.; O’Gara, F. Bile Acids Repress Hypoxia-Inducible Factor 1 Signaling and Modulate the Airway Immune Response. Infect. Immun. 2014, 82, 3531–3541. [Google Scholar] [CrossRef] [Green Version]
- Phelan, J.P.; Reen, F.J.; Dunphy, N.; O’Connor, R.; O’Gara, F. Bile acids destabilise HIF-1 alpha and promote anti-tumour phenotypes in cancer cell models. BMC Cancer 2016, 16, 476. [Google Scholar] [CrossRef] [Green Version]
- Reen, F.J.; Flynn, S.; Woods, D.F.; Dunphy, N.; Ní Chróinín, M.; Mullane, D.; Stick, S.; Adams, C.; O’Gara, F. Bile signalling promotes chronic respiratory infections and antibiotic tolerance. Sci. Rep. 2016, 6, 29768. [Google Scholar] [CrossRef] [Green Version]
- Lamarche, M.G.; Deziel, E. MexEF-OprN Efflux Pump Exports the Pseudomonas Quinolone Signal (PQS) Precursor HHQ (4-hydroxy-2-heptylquinoline). PLoS ONE 2011, 6, e24310. [Google Scholar] [CrossRef]
- Issa, K.H.B.; Phan, G.; Broutin, I. Functional Mechanism of the Efflux Pumps Transcription Regulators From Pseudomonas aeruginosa Based on 3D Structures. Front. Mol. Biosci. 2018, 5, 57. [Google Scholar] [CrossRef] [Green Version]
- Tegos, G.P.; Haynes, M.; Strouse, J.J.; Khan, M.M.T.; Bologa, C.G.; Oprea, T.I.; Sklar, L.A. Microbial Efflux Pump Inhibition: Tactics and Strategies. Curr. Pharm. Des. 2011, 17, 1291–1302. [Google Scholar] [CrossRef] [Green Version]
- Thakur, V.; Uniyal, A.; Tiwari, V. A comprehensive review on pharmacology of efflux pumps and their inhibitors in antibiotic resistance. Eur. J. Pharmacol. 2021, 903, 174151. [Google Scholar] [CrossRef]
- Blanco, P.; Corona, F.; Martinez, J.L. Biolog Phenotype Microarray Is a Tool for the Identification of Multidrug Resistance Efflux Pump Inducers. Antimicrob. Agents Chemother. 2018, 62, e01263-18. [Google Scholar] [CrossRef] [Green Version]
- Kerr, R.; Jabbari, S.; Blair, J.M.A.; Johnston, I.G. Dynamic Boolean modelling reveals the influence of energy supply on bacterial efflux pump expression. J. R. Soc. Interface 2022, 19, 20210771. [Google Scholar] [CrossRef]
- Aendekerk, S.; Diggle, S.P.; Song, Z.; Høiby, N.; Cornelis, P.; Williams, P.; Cámara, M. The MexGHI-OpmD multidrug efflux pump controls growth, antibiotic susceptibility and virulence in Pseudomonas aeruginosa via 4-quinolone-dependent cell-to-cell communication. Microbiology 2005, 151 Pt 4, 1113–1125. [Google Scholar] [CrossRef] [Green Version]
- Aendekerk, S.; Ghysels, B.; Cornelis, P.; Baysse, C. Characterization of a new efflux pump, MexGHI-OpmD, from Pseudomonas aeruginosa that confers resistance to vanadium. Microbiology 2002, 148 Pt 8, 2371–2381. [Google Scholar] [CrossRef] [Green Version]
- Sakhtah, H.; Koyama, L.; Zhang, Y.; Morales, D.K.; Fields, B.L.; Price-Whelan, A.; Hogan, D.A.; Shepard, K.; Dietrich, L.E.P. The Pseudomonas aeruginosa efflux pump MexGHI-OpmD transports a natural phenazine that controls gene expression and biofilm development. Proc. Natl. Acad. Sci. USA 2016, 113, E3538–E3547. [Google Scholar] [CrossRef] [Green Version]
- Flynn, S.; Reen, F.J.; O’Gara, F. Exposure to Bile Leads to the Emergence of Adaptive Signaling Variants in the Opportunistic Pathogen Pseudomonas aeruginosa. Front. Microbiol. 2019, 10, 2013. [Google Scholar] [CrossRef] [Green Version]
- Meirelles, L.A.; Newman, D.K. Both toxic and beneficial effects of pyocyanin contribute to the lifecycle of Pseudomonas aeruginosa. Mol. Microbiol. 2018, 110, 995–1010. [Google Scholar] [CrossRef] [Green Version]
- Schiessl, K.T.; Hu, F.; Jo, J.; Nazia, S.Z.; Wang, B.; Price-Whelan, A.; Min, W.; Dietrich, L.E.P. Phenazine production promotes antibiotic tolerance and metabolic heterogeneity in Pseudomonas aeruginosa biofilms. Nat. Commun. 2019, 10, 762. [Google Scholar] [CrossRef] [Green Version]
- Reen, F.J.; Barret, M.; Fargier, E.; Muinneacháin, M.Ó.; O’Gara, F. Molecular evolution of LysR-type transcriptional regulation in Pseudomonas aeruginosa. Mol. Phylogenet. Evol. 2013, 66, 1041–1049. [Google Scholar] [CrossRef] [Green Version]
- Fargier, E.; Mac Aogáin, M.; Mooij, M.J.; Woods, D.F.; Morrissey, J.P.; Dobson, A.D.W.; Adams, C.; O’Gara, F. MexT Functions as a Redox-Responsive Regulator Modulating Disulfide Stress Resistance in Pseudomonas aeruginosa. J. Bacteriol. 2012, 194, 3502–3511. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.; Kim, S.H.; Ahn, J.; Jo, I.; Lee, Z.-W.; Choi, S.H.; Ha, N.-C. Crystal Structure of the Regulatory Domain of MexT, a Transcriptional Activator of the MexEF-OprN Efflux Pump in Pseudomonas aeruginosa. Mol. Cells 2019, 42, 850–857. [Google Scholar]
- Cuthbertson, L.; Nodwell, J.R. The TetR Family of Regulators. Microbiol. Mol. Biol. Rev. 2013, 77, 440–475. [Google Scholar] [CrossRef] [Green Version]
- Beggs, G.A.; Brennan, R.G.; Arshad, M. MarR family proteins are important regulators of clinically relevant antibiotic resistance. Prot. Sci. 2020, 29, 647–653. [Google Scholar] [CrossRef]
- Nishino, K.; Senda, Y.; Hayashi-Nishino, M.; Yamaguchi, A. Role of the AraC-XylS family regulator YdeO in multi-drug resistance of Escherichia coli. J. Antibiot. 2009, 62, 251–257. [Google Scholar] [CrossRef] [Green Version]
- Dashtbani-Roozbehani, A.; Brown, M.H. Efflux pump mediated antimicrobial resistance by staphylococci in health-related environments: Challenges and the quest for inhibition. Antibiotics 2021, 10, 1502. [Google Scholar] [CrossRef]
- Sabatini, S.; Piccioni, M.; Felicetti, T.; De Marco, S.; Manfroni, G.; Pagiotti, R.; Nocchetti, M.; Cecchetti, V.; Pietrella, D. Investigation on the effect of known potent S. aureus NorA efflux pump inhibitors on the staphylococcal biofilm formation. RSC Adv. 2017, 7, 37007–37014. [Google Scholar] [CrossRef] [Green Version]
- Andersen, J.L.; He, G.-X.; Kakarla, P.; KC, R.; Kumar, S.; Lakra, W.S.; Mukherjee, M.M.; Ranaweera, I.; Shrestha, U.; Tran, T. Multidrug efflux pumps from Enterobacteriaceae, Vibrio cholerae and Staphylococcus aureus bacterial food pathogens. Int. J. Environ. Res. Public Health 2015, 12, 1487–1547. [Google Scholar] [CrossRef] [Green Version]
- Truong-Bolduc, Q.C.; Zhang, X.; Hooper, D.C. Characterization of NorR protein, a multifunctional regulator of norA expression in Staphylococcus aureus. J. Bacteriol. 2003, 185, 3127–3138. [Google Scholar] [CrossRef] [Green Version]
- Pasqua, M.; di Patti, M.C.B.; Fanelli, G.; Utsumi, R.; Eguchi, Y.; Trirocco, R.; Prosseda, G.; Grossi, M.; Colonna, B. Host-Bacterial pathogen communication: The Wily role of the multidrug efflux pumps of the MFS family. Front. Mol. Biosci. 2021, 8, 723274. [Google Scholar] [CrossRef]
- Truong-Bolduc, Q.C.; Hsing, L.C.; Villet, R.; Bolduc, G.R.; Estabrooks, Z.; Taguezem, G.F.; Hooper, D.C. Reduced aeration affects the expression of the NorB efflux pump of Staphylococcus aureus by posttranslational modification of MgrA. J. Bacteriol. 2012, 194, 1823–1834. [Google Scholar] [CrossRef] [Green Version]
- Rahmati, S.; Yang, S.; Davidson, A.L.; Zechiedrich, E.L. Control of the AcrAB multidrug efflux pump by quorum-sensing regulator SdiA. Mol. Microbiol. 2002, 43, 677–685. [Google Scholar] [CrossRef]
- Yang, S.; Lopez, C.R.; Zechiedrich, E.L. Quorum sensing and multidrug transporters in Escherichia coli. Proc. Natl. Acad. Sci. USA 2006, 103, 2386–2391. [Google Scholar] [CrossRef] [Green Version]
- Wolloscheck, D.; Krishnamoorthy, G.; Nguyen, J.; Zgurskaya, H.I. Kinetic Control of Quorum Sensing in Pseudomonas aeruginosa by Multidrug Efflux Pumps. ACS Infect. Dis. 2018, 4, 185–195. [Google Scholar] [CrossRef] [Green Version]
- Minagawa, S.; Inami, H.; Kato, T.; Sawada, S.; Yasuki, T.; Miyairi, S.; Horikawa, M.; Okuda, J.; Gotoh, N. RND type efflux pump system MexAB-OprM of Pseudomonas aeruginosa selects bacterial languages, 3-oxo-acyl-homoserine lactones, for cell-to-cell communication. BMC Microbiol. 2012, 12, 70. [Google Scholar] [CrossRef] [Green Version]
- Compagne, N.; Da Cruz, A.V.; Müller, R.T.; Hartkoorn, R.C.; Flipo, M.; Pos, K.M. Update on the Discovery of Efflux Pump Inhibitors against Critical Priority Gram-Negative Bacteria. Antibiotics 2023, 12, 180. [Google Scholar] [CrossRef]
- Martínez, O.F.; Cardoso, M.H.; Ribeiro, S.M.; Franco, O.L. Recent Advances in Anti-virulence Therapeutic Strategies With a Focus on Dismantling Bacterial Membrane Microdomains, Toxin Neutralization, Quorum-Sensing Interference and Biofilm Inhibition. Front. Cell. Infect. Microbiol. 2019, 9, 74. [Google Scholar] [CrossRef] [Green Version]
- Rampioni, G.; Leoni, L.; Williams, P. The art of antibacterial warfare: Deception through interference with quorum sensing-mediated communication. Bioorg Chem. 2014, 55, 60–68. [Google Scholar] [CrossRef]
- Garcia-Contreras, R.; Maeda, T.; Wood, T.K. Can resistance against quorum-sensing interference be selected? ISME J. 2016, 10, 4–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, W.; Liu, J.; Zhang, M.; Shi, H.; Zheng, S.; Jin, X.; Gao, Y.; Wang, S.; Ji, A.; Lou, H. Efflux pump-mediated resistance to antifungal compounds can be prevented by conjugation with triphenylphosphonium cation. Nat. Commun. 2018, 9, 5102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holmes, A.R.; Cardno, T.S.; Strouse, J.J.; Ivnitski-Steele, I.; Keniya, M.V.; Lackovic, K.; Monk, B.C.; Sklar, L.A.; Cannon, R.D. Targeting efflux pumps to overcome antifungal drug resistance. Future Med. Chem. 2016, 8, 1485–1501. [Google Scholar] [CrossRef] [Green Version]
- Prasad, R.; Rawal, M.K. Efflux pump proteins in antifungal resistance. Front. Pharmacol. 2014, 5, 202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heacock-Kang, Y.; Sun, Z.; Zarzycki-Siek, J.; Poonsuk, K.; McMillan, I.A.; Chuanchuen, R.; Hoang, T.T. Two Regulators, PA3898 and PA2100, Modulate the Pseudomonas aeruginosa Multidrug Resistance MexAB-OprM and EmrAB Efflux Pumps and Biofilm Formation. Antimicrob. Agents Chemother. 2018, 62, e01459-18. [Google Scholar] [CrossRef] [Green Version]
- Lu, W.-J.; Lin, H.-J.; Hsu, P.-H.; Lai, M.; Chiu, J.-Y.; Lin, H.-T.V. Brown and Red Seaweeds Serve as Potential Efflux Pump Inhibitors for Drug-Resistant Escherichia coli. Evid.-Based Complement. Altern. Med. 2019, 2019, 1836982. [Google Scholar] [CrossRef] [Green Version]
- Durães, F.; Szemerédi, N.; Kumla, D.; Pinto, M.; Kijjoa, A.; Spengler, G.; Sousa, E. Metabolites from Marine-Derived Fungi as Potential Antimicrobial Adjuvants. Mar. Drugs 2021, 19, 475. [Google Scholar] [CrossRef]
- Reen, F.J.; Gutiérrez-Barranquero, J.A.; Dobson, A.D.W.; Adams, C.A.; O’Gara, F. Emerging Concepts Promising New Horizons for Marine Biodiscovery and Synthetic Biology. Mar. Drugs 2015, 13, 2924–2954. [Google Scholar] [CrossRef] [Green Version]
- Gutiérrez-Barranquero, J.A.; Reen, F.J.; Parages, M.L.; McCarthy, R.; Dobson, A.D.W.; O’Gara, F. Disruption of N-acyl-homoserine lactone-specific signalling and virulence in clinical pathogens by marine sponge bacteria. Microb. Biotechnol. 2019, 12, 1049–1063. [Google Scholar] [CrossRef]
- Digirolamo, J.A.; Li, X.-C.; Jacob, M.R.; Clark, A.M.; Ferreira, D. Reversal of Fluconazole Resistance by Sulfated Sterols from the Marine Sponge Topsentia sp. J. Nat. Prod. 2009, 72, 1524–1528. [Google Scholar] [CrossRef] [Green Version]
- Cernicchi, G.; Felicetti, T.; Sabatini, S. Microbial Efflux Pump Inhibitors: A Journey around Quinoline and Indole Derivatives. Molecules 2021, 26, 6996. [Google Scholar] [CrossRef]
- Felicetti, T.; Cannalire, R.; Nizi, M.G.; Tabarrini, O.; Massari, S.; Barreca, M.L.; Manfroni, G.; Schindler, B.D.; Cecchetti, V.; Kaatz, G.W. Studies on 2-phenylquinoline Staphylococcus aureus NorA efflux pump inhibitors: New insights on the C-6 position. Eur. J. Med. Chem. 2018, 155, 428–433. [Google Scholar] [CrossRef]
- Cedraro, N.; Cannalire, R.; Astolfi, A.; Mangiaterra, G.; Felicetti, T.; Vaiasicca, S.; Cernicchi, G.; Massari, S.; Manfroni, G.; Tabarrini, O. From Quinoline to Quinazoline-Based S. aureus NorA Efflux Pump Inhibitors by Coupling a Focused Scaffold Hopping Approach and a Pharmacophore Search. ChemMedChem 2021, 16, 3044–3059. [Google Scholar] [CrossRef]
- Lomovskaya, O.; Warren, M.S.; Lee, A.; Galazzo, J.; Fronko, R.; Lee, M.; Blais, J.; Cho, D.; Chamberland, S.; Renau, T.; et al. Identification and characterization of inhibitors of multidrug resistance efflux pumps in Pseudomonas aeruginosa: Novel agents for combination therapy. Antimicrob. Agents Chemother. 2001, 45, 105–116. [Google Scholar] [CrossRef] [Green Version]
- Bhattacharyya, T.; Sharma, A.; Akhter, J.; Pathania, R. The small molecule IITR08027 restores the antibacterial activity of fluoroquinolones against multidrug-resistant Acinetobacter baumannii by efflux inhibition. Int. J. Antimicrob. Agents 2017, 50, 219–226. [Google Scholar] [CrossRef]
- Vargiu, A.V.; Ruggerone, P.; Opperman, T.J.; Nguyen, S.T.; Nikaido, H. Molecular mechanism of MBX2319 inhibition of Escherichia coli AcrB multidrug efflux pump and comparison with other inhibitors. Antimicrob. Agents Chemother. 2014, 58, 6224–6234. [Google Scholar] [CrossRef] [Green Version]
- Opperman, T.J.; Nguyen, S.T. Recent advances toward a molecular mechanism of efflux pump inhibition. Front. Microbiol. 2015, 6, 421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, A.; Gupta, V.K.; Pathania, R. Efflux pump inhibitors for bacterial pathogens: From bench to bedside. Indian J. Med. Res. 2019, 149, 129–145. [Google Scholar] [PubMed]
- Gurney, J.; Pradier, L.; Griffin, J.S.; Gougat-Barbera, C.; Chan, B.K.; Turner, P.E.; Kaltz, O.; Hochberg, M.E. Phage steering of antibiotic-resistance evolution in the bacterial pathogen, Pseudomonas aeruginosa. Evol. Med. Public Health 2020, 2020, 148–157. [Google Scholar] [CrossRef] [PubMed]
- Chan, B.K.; Sistrom, M.; Wertz, J.E.; Kortright, K.E.; Narayan, D.; Turner, P.E. Phage selection restores antibiotic sensitivity in MDR Pseudomonas aeruginosa. Sci. Rep. 2016, 6, 26717. [Google Scholar] [CrossRef] [Green Version]
- Chan, B.K.; Turner, P.E.; Kim, S.; Mojibian, H.R.; Elefteriades, J.A.; Narayan, D. Phage treatment of an aortic graft infected with Pseudomonas aeruginosa. Evol. Med. Public Health 2018, 2018, 60–66. [Google Scholar] [CrossRef] [Green Version]
- Wen, X.; Langevin, A.M.; Dunlop, M.J. Antibiotic export by efflux pumps affects growth of neighboring bacteria. Sci. Rep. 2018, 8, 15120. [Google Scholar] [CrossRef] [Green Version]
- Molina-Santiago, C.; Udaondo, Z.; Cordero, B.F.; Ramos, J.-L. Interspecies cross-talk between co-cultured Pseudomonas putida and Escherichia coli. Environ. Microbiol. Rep. 2017, 9, 441–448. [Google Scholar] [CrossRef]
- Evans, K.C.; Benomar, S.; Camuy-Vélez, L.A.; Nasseri, E.B.; Wang, X.; Neuenswander, B.; Chandler, J.R. Quorum-sensing control of antibiotic resistance stabilizes cooperation in Chromobacterium violaceum. ISME J. 2018, 12, 1263–1272. [Google Scholar] [CrossRef] [Green Version]
- Molina-Santiago, C.; Daddaoua, A.; Fillet, S.; Duque, E.; Ramos, J.-L. Interspecies signalling: Pseudomonas putida efflux pump TtgGHI is activated by indole to increase antibiotic resistance. Environ. Microbiol. 2014, 16, 1267–1281. [Google Scholar] [CrossRef]
- Duan, K.; Dammel, C.; Stein, J.; Rabin, H.; Surette, M.G. Modulation of Pseudomonas aeruginosa gene expression by host microflora through interspecies communication. Mol. Microbiol. 2003, 50, 1477–1491. [Google Scholar] [CrossRef] [Green Version]
- Tan, C.A.Z.; Lam, L.N.; Biukovic, G.; Soh, E.Y.-C.; Toh, X.W.; Lemos, J.A.; Kline, K.A. Enterococcus faecalis Antagonizes Pseudomonas aeruginosa Growth in Mixed-Species Interactions. J. Bacteriol. 2022, 204, e0061521. [Google Scholar] [CrossRef] [PubMed]
- Morgan, B.G.; Warren, P.; Mewis, R.E.; Rivett, D.W. Bacterial dominance is due to effective utilisation of secondary metabolites produced by competitors. Sci. Rep. 2020, 10, 2316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ioannidis, A.; Magana, M.; Bologa, C.G.; Oprea, T.I.; Paulsen, I.T.; Tegos, G.P. Defining the microbial effluxome in the content of the host-microbiome interaction. Front. Pharmacol. 2015, 6, 31. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
| Efflux Pump (Super) Family | Energy Source for Functionality | Membrane-Spanning Nature | Gram-Positive Example | Gram-Negative Example |
|---|---|---|---|---|
| ABC Superfamily | ATP Hydrolysis [1] | Forms a tri-partite complex in Gram-negative bacteria [1] | Sav1866—Staphylococcus aureus [11] | HlyB and MacB (TolC-dependent)—Escherichia coli [11] |
| RND Family | Proton Motive Force (PMF)—largely via respiration [1] | Forms a tri-partite complex [1] | Unique to Gram-negative bacteria [12] | MexAB-OprM in Pseudomonas aeruginosa [13] |
| MFS | Proton Motive Force (PMF)—largely via respiration [1] | Forms a tri-partite complex in Gram-negative bacteria | MdrM and MdrT in Listeria monocytogenes; NorA—Staphylococcus aureus [14] | EmrKY—Shigella flexneri [15] |
| SMR Family | Proton Motive Force (PMF)—largely via respiration [14] | Not known to form tri-partite complexes [9] | QacA/B—Enterococcus faecalis [14] | AbeS—Acinetobacter baumannii [14] |
| MATE Family | Proton Motive Force (PMF)—largely via respiration [14] | Not known to form tri-partite complexes [9] | MepA—Staphylococcus aureus [16] | NorM—Vibrio cholerae [14] |
| PACE Family | Proton Motive Force (PMF)—largely via respiration [17] | Not known to form tri-partite complexes [9] | Not known to be found in Gram-positive bacteria [17] | Acel—Acinetobacter baumannii [14] |
| AbgT Family | Proton Motive Force (PMF)—largely via respiration [18] | Forms a tri-partite complex in Gram—negative bacteria [18] | Homolog in Streptomyces coelicolor [18] | MtrF—Neisseria gonorrhoeae [18] |
| Environmental Cue(s) | Regulator(s) | Efflux Pump(s) Affected | Outcome(s) |
|---|---|---|---|
| Unclear | RocS2-RocA2 (TCS) | MexAB-OprM downregulated | Promotes biofilm formation |
| Antibiotic exposure (fluoroquinolone) | MexT (LTTR, OCS) Linked with the OprD porin repressor | MexEF-OprN activated | Antibiotic extrusion |
| Antibiotic exposure | MexL (TetR, OCS) | MexJK—downregulated by MexL | Antibiotic extrusion (multiple classes) |
| Antibiotic exposure, phenolic compounds | NalC (TetR, OCS) | Indirect repressor of MexAB-OprM (via ArmR) | |
| Antibiotic exposure | NalD (TetR, OCS) | Represses MexAB-OprM—secondary regulator | Antibiotic extrusion |
| Membrane- and envelope-damaging agents, e.g., antibiotics, biocides, dyes, solvents, etc. | NfxB (TetR, OCS) | Represses MexCD-OprJ | Antibiotic extrusion; antibiotic tolerance in biofilms |
| pH, temperature, oxidative stress, nutrient availability, toxins, etc. | MexR (MarR, OCS) linked with AmrR (controlled by the TetR repressor, NalC) | Primary activator MexAB-OprM | Antibiotic and other toxic substance extrusion; homeostasis |
| Ribosome stress, perhaps due to antibiotic (aminoglycoside) exposure—inducer unknown | MexZ (TetR, OCS) Most frequently mutated in Cystic Fibrosis patients | MexXY-OprM | Antibiotic extrusion |
| Envelope stress, membrane perturbation (colistin, polymyxin B) | ParR-ParS (TCS) | MexXY expressed MexEF-OprN upregulated via MexS (activator) activation | Antibiotic extrusion |
| AmgR-AmgS (TCS) | MexXY expressed | Antibiotic extrusion | |
| Heavy metal exposure | CzcR-CzcS (TCS) | CzcABC | Heavy metal extrusion |
| CopR-CopS (TCS) | CzcABC | Heavy metal extrusion |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moore-Machacek, A.; Gloe, A.; O’Leary, N.; Reen, F.J. Efflux, Signaling and Warfare in a Polymicrobial World. Antibiotics 2023, 12, 731. https://doi.org/10.3390/antibiotics12040731
Moore-Machacek A, Gloe A, O’Leary N, Reen FJ. Efflux, Signaling and Warfare in a Polymicrobial World. Antibiotics. 2023; 12(4):731. https://doi.org/10.3390/antibiotics12040731
Chicago/Turabian StyleMoore-Machacek, Ay’sha, Antje Gloe, Niall O’Leary, and F. Jerry Reen. 2023. "Efflux, Signaling and Warfare in a Polymicrobial World" Antibiotics 12, no. 4: 731. https://doi.org/10.3390/antibiotics12040731
APA StyleMoore-Machacek, A., Gloe, A., O’Leary, N., & Reen, F. J. (2023). Efflux, Signaling and Warfare in a Polymicrobial World. Antibiotics, 12(4), 731. https://doi.org/10.3390/antibiotics12040731