Abstract
Antibiotic-resistance gene elements (ARGEs) such as antibiotic-resistance genes (ARGs), integrons, and plasmids are key to the spread of antimicrobial resistance (AMR) in marine environments. Kuwait’s marine area is vulnerable to sewage contaminants introduced by numerous storm outlets and indiscriminate waste disposal near recreational beaches. Therefore, it has become a significant public health issue and warrants immediate investigation. Coliforms, especially Gram-negative Escherichia coli, have been regarded as significant indicators of recent fecal pollution and carriers of ARGEs. In this study, we applied a genome-based approach to identify ARGs’ prevalence in E. coli isolated from mollusks and coastal water samples collected in a previous study. In addition, we investigated the plasmids and intl1 (class 1 integron) genes coupled with the ARGs, mediating their spread within the Kuwait marine area. Whole-genome sequencing (WGS) identified genes resistant to the drug classes of beta-lactams (blaCMY-150, blaCMY-42, blaCTX-M-15, blaDHA-1, blaMIR-1, blaOKP-B-15, blaOXA-1, blaOXA-48, blaTEM-1B, blaTEM-35), trimethoprim (dfrA14, dfrA15, dfrA16, dfrA1, dfrA5, dfrA7), fluroquinolone (oqxA, oqxB, qnrB38, qnrB4, qnrS1), aminoglycoside (aadA2, ant(3’’)-Ia, aph(3’’)-Ib, aph(3’)-Ia, aph(6)-Id), fosfomycin (fosA7, fosA_6, fosA, fosB1), sulfonamide (sul1, sul2, sul3), tetracycline (tet-A, tet-B), and macrolide (mph-A). The MFS-type drug efflux gene mdf-A is also quite common in E. coli isolates (80%). The plasmid ColRNAI was also found to be prevalent in E. coli. The integron gene intI1 and gene cassettes (GC) were reported to be in 36% and 33%, respectively, of total E. coli isolates. A positive and significant (p < 0.001) correlation was observed between phenotypic AMR-intl1 (r = 0.311) and phenotypic AMR-GC (r = 0.188). These findings are useful for the surveillance of horizontal gene transfer of AMR in the marine environments of Kuwait.
1. Introduction
The excessive use of antibiotics to treat infectious diseases has legated the world to the public health hazard of antibiotic resistance [1,2,3]. Aquatic environs have been identified as reservoirs of antibiotic resistancegenes (ARGs); however, little is known about their distribution, spread and migration [4,5]. Marine environments are also sinks for pharmaceuticals, disinfectants, heavy metals, organic compounds, microplastics, and atmospheric dust [6,7,8,9,10,11]. Among these, pollutants, pharmaceuticals, and heavy metals have been evidenced to impose selective pressure on inherent bacterial communities that often develop resistance genes against them [9,11,12,13,14,15,16,17,18]. In addition, genetic elements, for example, class 1 integrons, package ARGs into gene cassettes (GCs) and mediate their transfer to non-resistant bacterial communities via horizontal gene transfer (HGT) [19,20,21,22]. Although more consideration has been given to animal and human health with respect to antimicrobial resistance (AMR) monitoring, the environment often remains ignored. The World Health Organization propelled the One Health Concept, which calls for the evaluation of overall environmental health [23]. Immediate action is also warranted to monitor the threats of AMR in the Gulf Cooperation Council [24,25].
Emergency waste and unauthorized sewage discharge in Kuwait have introduced several antibiotic-resistant bacteria (ARBs) into Kuwait’s marine environments [18,26]. Among these, Escherichia coli could become an infectious animal and human bacterium showing resistance to almost all clinically used antibiotics. A wide profile of AMR E. coli has been previously demonstrated through conventional phenotypic antibiotic susceptibility testing (AST) in Kuwait’s marine environment [27,28,29]. However, data on the genes conferring resistance are lacking in these aquatic settings. In addition, class 1 integrons, GCs, and plasmids involved in their spread are unknown. Currently, microbiological methods of AMR surveillance are considered laborious, time-intensive, and provide limited phenotypic information. With the advent of molecular methods, a rapid and detailed genotypic assessment of all these components can be achieved [30,31,32,33,34]. These methods are now being embraced as a novel approach to map the whole genomes of bacterial isolates, ARGs, plasmids, integrons, and GCs simultaneously to complement the AMR phenotypic assays [32,35,36,37]. The goal of this research was to examine the genomic profiles of E. coli through whole-genome sequencing and to study the prevalence of ARGs, plasmids, integrons (intl1), and GCs.
2. Results
For the present investigation, selected E. coli isolates (n = 23) were subjected to whole-genome sequencing. All sequences ranged between 4,577,350 and 5,103,695 bases, with a Phred quality score above 20.
2.1. MLST and Phylogenetic Analysis
The open reading frames corresponding to seven sequence tags (STs), namely, adk-fumC-gyrB-icd-mdh-purA-recA, were identified in the assembled genomes of 23 E. coli isolates. Alignment of sequences against these STs established the Enterobacteriaceae origin of the strains. Further treatment with the mafft software and UGENE software confirmed 20 isolates as E. coli, and three of these closely matched with Enterobacter cloacae (SE111, SE181, and SE158). Molecular data analyses helped to correctly identify the species of E. cloacae; hence, we found the molecular tests to be technically superior for discriminating between closely related E. coli and E. cloaca strains. Further analysis was performed on the 20 E. coli isolates. The phylogenetic relationships of the selected E. coli were distributed into two clusters (Figure 1). A close relationship was observed between the isolates from spatially distant locations, different seasons, and varied matrices (marine waters or mollusks). For example, SE25 and KHE11 isolated from Al-Salam and Khiran were closely related. Strains SE19 and SC118 were isolated from different matrices, i.e., marine water during winter and mollusks collected during summer; however, they were positioned on the same branch of the phylogenetic tree for their similarity. Similarly, SC59 and 756E0 were also analogous. It was noticed that the three isolates of E. cloacae, namely, SE111, SE181, and SE158, exist separately as a group.

Figure 1.
Phylogeny of the strains isolated from Kuwait. A maximum-likelihood tree was plotted on the distances between the MLST sequences of the selected strains.
2.2. Antibiotic-Resistance Gene Elements
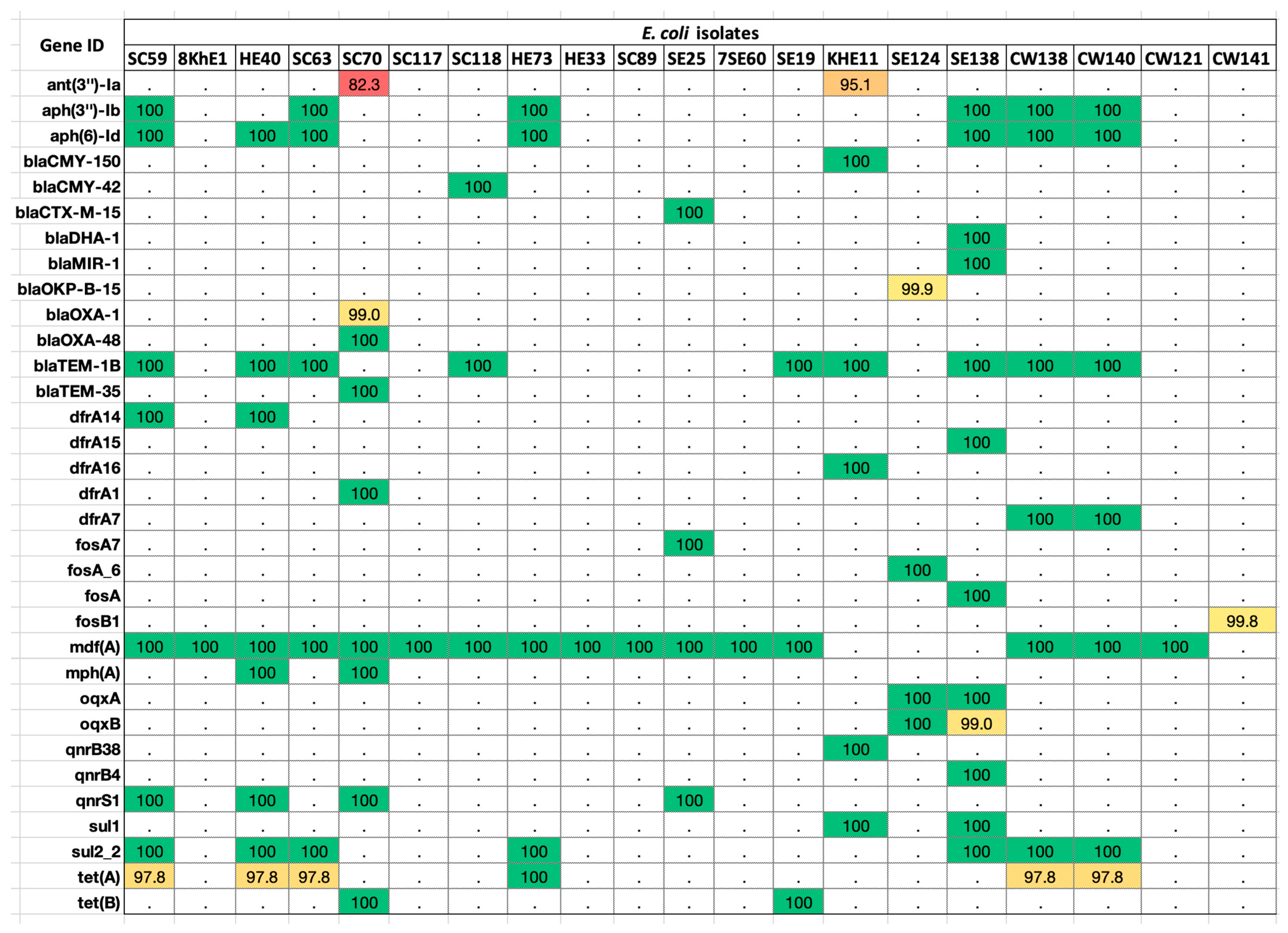
We submitted the assembled E. coli sequences to the ResFinder database and found matches against 33 ARGs (Figure 2; Table S1). The percentage identity of the sequences ranged between 80 and 100%. The highest frequency (80%) was that of the mdf (A) gene found in at least 16 isolates. This was followed by blaTem-1B, present in 45% of E. coli strains.

Figure 2.
ARGs distributed in the E. coli genomes. Colored boxes are positive for the respective ARG listed on the left-hand side panel. The values in each box show the percentage identity in the ResFinder database v4.0.0 (red > 80%; yellow > 90%; green > 100%; white < 0.0%).
The highest number of genes (10/33) originated from beta-lactam family (blaCMY-150, blaCMY-42, blaCTX-M-15, blaDHA-1, blaMIR-1, blaOKP-B-15, blaOXA-1, blaOXA-48, blaTEM-1B, blaTEM-35). This was followed by trimethoprim (6/33),-(dfrA14, dfrA15, dfrA16, dfrA1, dfrA5, dfrA7), fluoroquinolone (5/33) (oqxA, oqxB, QnrB38, QnrB4, QnrS1),aminoglycoside (5/33) (aadA2, ANT (3’’)-Ia, APH (3’’)-Ib, APH (3’)-Ia, APH(6)-Id), fosfomycin (4/33) (FosA7, FosA6, FosA, FosB1), sulfonamide (2/33) (sul1, sul2), and tetracycline (2/33) resistance genes (tetA, tetB). Single genes belonged to the drug class macrolide (mphA) and MFS-type drug efflux (mdfA).
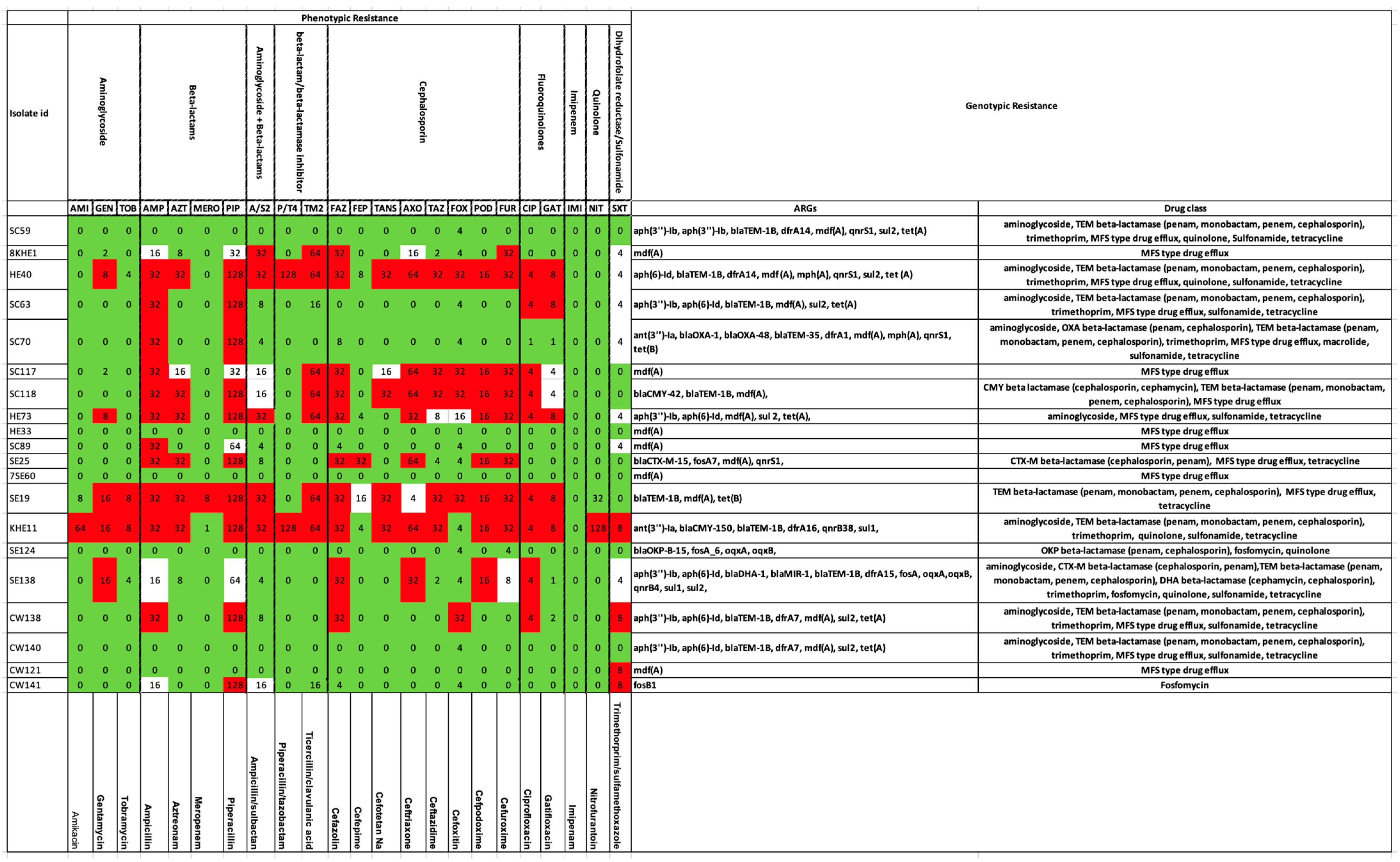
We compared the ARG summaries of these 20 strains with their AMR phenotypes (tested against a panel of 23 antibiotics). According to the genotypic assay, all these strains possessed ARGs; therefore, they were considered as potentially resistant to at least one of the drug classes. Phenotypically, the strains SC59, HE33, 7SE60, SE124, and CW140 were sensitive to all the antibiotics (Figure 3).

Figure 3.
Genotypic versus phenotypic resistance in selected isolates of E. coli. Phenotypes are presented on the left-hand side panel. The color codes represent Red—Resistant, Green—Susceptible, and White—Intermediate. Corresponding genotypes are shown on the right-hand side.
Resistance to beta-lactams was confirmed both genotypically and phenotypically in 40% of the isolates, namely, HE40 (blaTEM-1B), SC63 (blaTEM-1B), SC70 (blaOXA-1, blaOXA-48, blaTEM-35), SC118 (blaCMY-42, blaTEM-1B), SE25 (blaCTX-M-15), SE19 (blaTEM-1B), KHE11 (blaCMY-150, blaTEM-1B), and CW138 (blaTEM-1B). Strain SE138 bearing beta-lactamase genes (blaDHA-1, blaMIR-1, blaTEM-1B) depicted an intermediate phenotype against this drug class. Strain 8KHE1 was phenotypically resistant to beta-lactams + beta-lactamase inhibitors and cephalosporins. None of the genes were resistant to the above drug classes; rather, the mdfA (MFS-type drug efflux) resistant against tetracycline, disinfectants, and antiseptics was found in the genotype. Similar was the case with strains SC117 and SC89. Within the beta-lactamase, genes resistant to CMY-beta lactamase (cephamycin), CTX-M-beta lactamase (penam, cephalosporin), DHA-beta lactamase (cephalosporin, cephamycin), MIR-beta lactamase (monobactam, cephalosporin), OKP-beta lactamase (penam, cephalosporin), OXA-beta lactamase (penam, cephalosporin, carbapenem), and TEM-beta lactamase (penam, monobactam, penem, cephalosporin) were observed in strains SC118; SE25, SE138; SE138; SE124; SC70; and SC59, HE40, SC63, SC70, SC118, SE19, KHE11, SE138, CW138, and CW140, respectively. Cephalosporin was one of the drugs in the antibiotic panel. As evident from the above statements, this first-generation beta-lactamase confers resistance against all the sub-categories of beta-lactams. TEM-beta lactamase was one of the most common genes detected in the tested isolates (45%). Among the carbapenems, the SE19 isolate expressed a meropenem (carbapenem)-resistant phenotype and also possessed blaTEM (TEM-beta lactamase) gene. All the remaining strains were susceptible to meropenem. In addition to this, susceptibility to imipenem (carbapenem) was observed in all the strains (Figure 3). None of the genotypes were positive for the blaIMP group of genes. Strain CW141, phenotypically resistant to piperacillin (beta-lactams), was devoid of relevant genes.
Matching genotypes and phenotypes (20%) for aminoglycoside resistance were recorded in HE40 (aph(6)-Id), HE73 (aph(3’’)-Ib, aph(6)-Id), KHE11 (ant(3’’)-Ia), and SE138 (aph(3’’)-Ib, aph(6)-Id), whereas genes without any phenotypic expression for this drug class were observed in SC59 (aph(3’’)-Ib, aph(3’’)-Ib), SC70 (ant(3’’)-Ia), CW138 (aph(3’’)-Ib, aph(6)-Id), and CW140 (aph(3’’)-Ib, aph(6)-Id). Strains 8KHE1, SC117, and SC89 depicted an aminoglycoside-resistant profile but were devoid of related genes. Contrastingly, its genotype possessed mdfA (MFS-type drug efflux) resistance against tetracycline, disinfectants, and antiseptics.
In the case of fluroquinolones, phenotypic resistance was expressed in 45% of isolates (HE40, SC63, SC117, SC118, HE733, SE19, KHE11, SE138, and CW138). Among these, only 15% (3/20) of the isolates, i.e., HE40, KHE11, and SE138, possessed resistance genes (qnrS1). Phenotypic sensitivity against quinolone (used interchangeably with fluroquinolone) was demonstrated by all the strains except KHE11. ARGs against this drug class were located in strains SC59 (qnrS1), HE40 (qnrS1), SC70 (qnrS1), SE25 (qnrS1), and SE138 (oqxA, oqxB, qnrB4).
Trimethoprim- or sulfonamide-resistant phenotypes were observed in KHE11, CW138, CW121, and CW141, of which only the first two strains (10%) possessed ARGs (KHE11—dfrA15, sul1; CW138—dfrA7, sul2). Four strains with intermediate phenotypes were positive for corresponding ARGs (HE40—dfrA14, sul2; SC63—sul2; HE73—sul2; and SE138—dfrA15, sul1, sul2).
Moreover, SC59, HE40, SC63, SC70, HE73, SE25, SE19, KHE11, SE138, CW138, and CW140 all possessed tetracycline-resistant genotypes (ARG—tetA, tetB), which was not captured by the AST as the chosen panel lacked tetracycline. Similarly, macrolide-resistant genes (mphA) were recorded in SC70 and fosfomycin-resistant genes (fosB1) in CW141 that were not tested phenotypically.
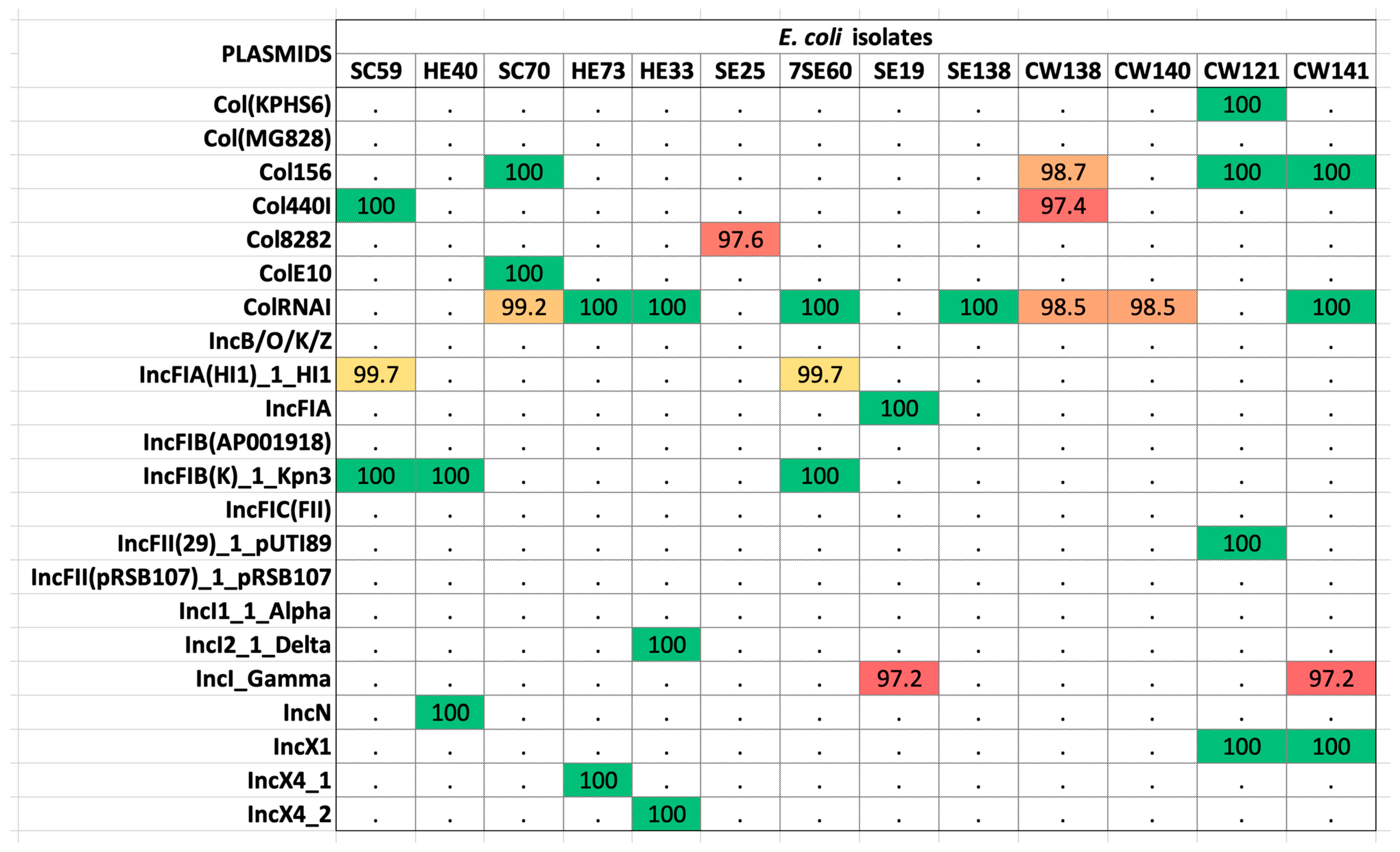
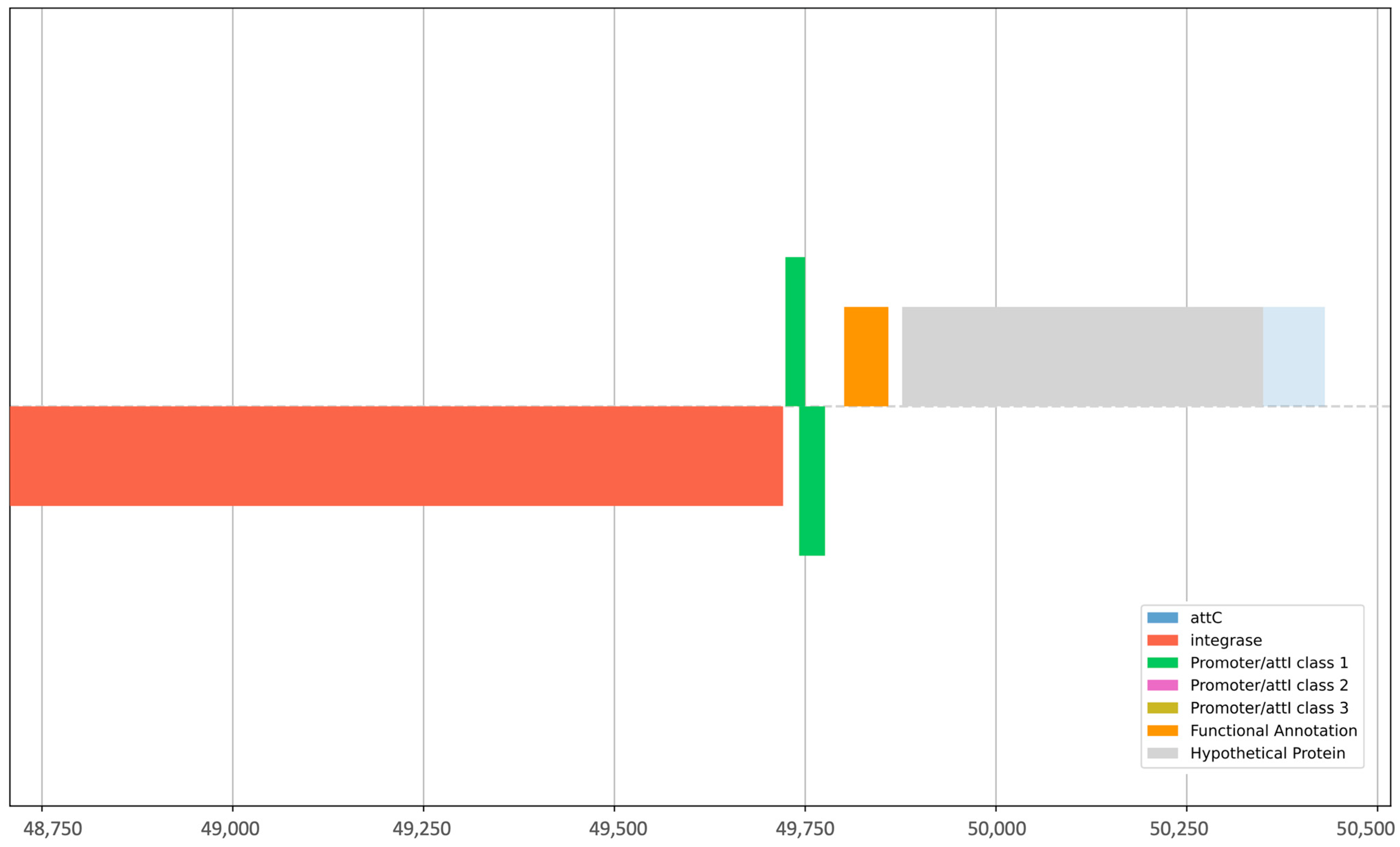
The alignment of sequences against the plasmid finder database revealed 13 isolates as hosts to 22 plasmids (Table S2). The most prevalent was the ColRNAI plasmid found in 11 isolates (42.3%) (Figure 4). Sequences were then concomitantly mined for integrons, recombination sites, and promoters. The genetic elements identified were attC (gene cassette recombination sites), intI (intersection tyr-integrase), Pc_1 (gene cassette promoter class), Pint_1 (integron promoter), and attI_1 (integron recombination site). The elements attC and intI were reported in 20% (4/20) of the strains, whereas Pc_1 and attI_1 were detected in 10% (2/20) of the isolates. Pint_1 was found in 15% (3/20) of the tested samples. Isolates CW138, CW121, and CW141 were positive for integron with the integron integrase (intI) and attC site nearby, whereas HE70 was positive for the latter only, lacking recombination sites (Table 1). These elements aid the mobilization of ARGs in other strains. The intl1 gene encodes the integrase enzyme, the promoter Pc ensures its expression, and the site-specific recombination takes place between the attl and attC sites. The GC is an open reading frame (ORF) without a promoter but with a recombination site. These GCs can integrate novel genes and support the bacterial strains to adapt. On the other hand, strain SC70 possessed only a cluster of attC sites without any integrase in its vicinity (Table S3). Classification of integrons and gene cassettes revealed the presence of class 1 integron (intl1) and GC1. All the genetic elements within strain CW141 are shown in Figure 5. Intriguingly, this strain possessed the fosfomycin-resistant gene (fosB1) and expressed phenotypic AMR against piperacillin (an extended spectrum beta-lactam) and trimethoprim, as well as harbored the incX1 plasmid.

Figure 4.
The plasmids found in selected isolates of E. coli revealed by whole-genome sequencing. Colored boxes mark the presence of the plasmids and their percentage identity in the database (red > 80%; yellow > 90%; green > 100%; white < 0.0%). Plasmids were fetched from the PlasmidFinder database.

Table 1.
Integrons in E. coli isolates.

Figure 5.
Graphical representation of integrons, recombination sites, and promoters present in E. coli strain CW141.
2.3. PCR-Based Identification of Intl1 and GCs
Among all the isolated strains (n = 598 from both mollusks and seawater), 216 were positive for the intl1 (36%) gene, exhibiting a band of 400 bp in size on the agarose gel. These were associated with 198 GCs (36%) (Figure 6A). We looked at the AMR phenotypes of intl1-positive isolates (n = 216); interestingly, 95% of them were phenotypically resistant to at least one antibiotic among the panel (aminoglycoside, beta-lactams, aminoglycoside + beta-lactam inhibitors, beta-lactams/beta-lactamase inhibitors, cephalosporins, fluroquinolones, quinolones, imipenem, and dihydrofolate-reductase/trimethoprim) and the remaining 5% were susceptible (Figure 6C) to all the tested antibiotics. Considering the total AMR phenotypes of E. coli (n = 420), approximately 49% were negative for intl1 (Figure 6B). Spearman’s correlation model established a positive correlation between phenotypic AMR-intl1-GC. Overall, the coefficients were statistically significant (intl1 ß–0.230, p < 0.001; GC ß–0.121, p < 0.001). There was 7.5% variance between intl1-GC resistance (F—24.28, p < 0.001). Positive correlations were observed between phenotypic AMR-intl1 (0.310; p < 0.001) and phenotypic AMR-GC (0.190; p < 0.001). The intl1 and GC were also significantly correlated (0.120; p < 0.003).
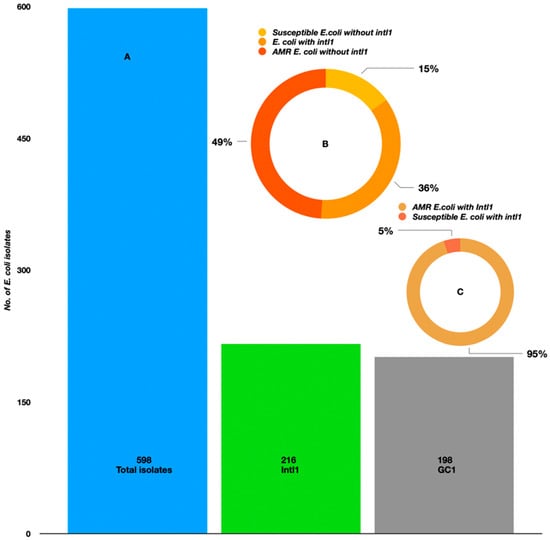
Figure 6.
(A) Prevalence of class 1 integrons and gene cassettes in E. coli isolated from the Kuwait marine waters and molluscans. (B) The occurrence of class 1 integrons in E. coli isolated during summer and winter seasons. (C) AMR-susceptible E. coli with intl1.
3. Discussion
WGS has become the most preferred method to identify ARGs and associated genetic elements [38]. Recently, it identified ARGs in both highly resistant and susceptible isolates in the order of Enterobacterales [38]. This indicates that a population of resistant E.coli is floating in the coastal waters of Kuwait [36,39]. In addition to resistance profiling, WGS was also used presently for the identification of Enterobacteriaceae species. Interestingly, three of the isolates (13.0%) were identified as Enterobacter cloacae. A study from the Norwegian coast demonstrated that MALDI-TOF-MS (Matrix-assisted laser desorption ionization–Time of flight mass spectrometry) identified 90% of isolated strains as E. coli, whereas the remaining 10% were Klebsiella, Citrobacter, and Enterobacter [40]. More robust and precise identification approaches such as molecular methods are preferred over conventional microbiological assays, especially in mollusks and other filter feeders in the marine environment that are prone to pathogenic exposure, in addition to E. coli, for accurate discrimination between closely related species [41]. Further, the MLST-based phylogeny established a close relationship between the 20 isolates. This indicates a common origin of E. coli, most probably one of the point sources of emergency outfalls. The genetic similarity between the E. coli isolates of seawater and mollusks is most probably due to the filter feeding mechanism of the latter [42]. A comprehensive analysis might assist in contact tracing of these AMR strains.
Intriguingly, the E. coli isolates (n = 20) possessed genes resistant against seven drug classes. This is most likely due to the selective pressure imposed by antibiotics such as Azithromycin, Cefalexin, Ciprofloxacin, Clarithromycin, Dimetridazole, Erythromycin, Metronidazole, Metronidazole-OH, Ofloxacin, Ronidazole, Sulfamethoxazole, Tetracycline, and Trimethoprim found in seawater samples [8] and the effluent streams of waste-water treatment plants in Kuwait [43]. Moreover, the presence of conjugative plasmids and integrons in these strains is indicative of ARG transmission through horizontal gene transfer [19,20,22,44,45]. Nevertheless, this study only targeted the fecal coliforms, and the likelihood of other microbial communities with diverse ARG profiles is expected. The advanced method of high-throughput shotgun metagenomic sequencing would be a more valuable tool in the assessment of resistomes at contaminated and non-contaminated sites [26,38].
The presence of beta-lactam genes and their phenotypic expression in strains such as HE40, SC63, SC70, SC118, SE25, SE19, KHE11, SE138, and CW138 draws our immediate attention. Moreover, plasmid vectors were also recorded in HE40, SC70, SE25, SE19, SE138, and CW138. In addition, the strains HE40 and CW138 also possessed integrons and gene cassettes, indicating the possibility of horizontal gene transfer of beta-lactamase genes in the region. Extended spectrum beta-lactams are the last-choice drugs to treat hardcore bacterial infections [46,47]. Worldwide circulation of beta-lactamases have been reported recently with more than 40% in Asian countries [48]. A baseline study also documented the presence of beta-lactams in aerosols collected from a public building situated near the coast of Kuwait [49]. Moreover, trimethoprim resistance was confirmed in strains KHE11 and CW138. The dominance of the MFS-type drug efflux gene (tetracycline, disinfectants, antiseptics) was also noticed in 75% of sequenced isolates. Tetracycline-resistant genes (tetA/tetB) were found in 55% of isolates. The AMR phenotypes for these antibiotics were missing and need to be taken into consideration while performing future phenotypic assays.
We also report that 35.99% of AMR E.coli held the intl1 gene, in parallel with Enterobacteriaceae found in Korea (41.4%) [50]. Contrastingly, only 12.1% of Enterobacteriaceae strains were positive for class 1 integrons in an effluent handling system in Poland [51]. Intl1 positive strains were maximum in locations proximal to the coastal emergency waste outfalls. Numerous storm outlets discharge emergency waste into the country’s marina, leading to deteriorated water quality [24,27,52,53]. These findings are in congruence with our earlier reports on the presence of AMR microbes, ARGs, integrons, plasmids, and insertion sequences near the outfalls [18,26]. We attribute this to the antibiotics, pharmaceuticals, and other contaminants introduced into the seawater [8,43]. A constant selection pressure is thought to elevate the horizontal gene transfer frequency within these aquatic milieus [25,33,36,45,54,55]. Regression analysis predicted a relationship (7.5% variance) between intI1-GC resistance. This suggests that the class 1 integron could potentially play a role in the transmission of AMR in Enterobacteriaceae [45]; however, other contaminants, such as metals, cannot be ignored [5,56,57,58].
In the present investigation, WGS identified ARGs in both resistant and susceptible E. coli phenotypes. The presence of intl1 and GCs indicates the HGT of AMR in the region. WGS is recommended as an accurate and precise monitoring tool for individual AMR strains perpetrating in marine ecosystems.
4. Materials and Methods
4.1. Sampling Locations, Collection and Isolation of Strains
A detailed overview of the sampling task is provided elsewhere [28] as they were previously collected and tested for AMR through standard microbiological methods. However, for ease of readership, a succinct description is provided in Section 4.1.1, Section 4.1.2 and Section 4.1.3. A detailed description of the genomic analysis performed in the current study has been described in Section 4.2, Section 4.3, Section 4.4 and Section 4.5.
4.1.1. Marine Waters
Seawater was collected from Al-Ghazali (29°347784 N, 47°911974 E), Al-Salam (29°357207 N, 47°946784 E), Abu Al-Hasaniya (29°125652 N, 48°633155 E), and Al-Khairan (28°665070 N, 48°387640 E) in an earlier campaign [28]. Six replicates were sampled in 200 ml sterile bottles from each site in both summer (Jul–Aug 2015; 34.0 °C ± 2.0) and winter (Dec–Feb 2016; 17.2 °C ± 2.0) seasons (Table S4). In situ measurements of pH (8.2 ± 0.2), salinity (42 ± 2.0), and temperature were made employing a hand-held, multi-probe water quality meter (Hanna instruments, Smithfield, RI, USA). Samples were transported on ice to the Environment Public Authority (EPA) Kuwait laboratories. Samples were processed at EPA laboratories following an established protocol [36]. Briefly, 10 mL of seawater was pipetted into ¼ strength Ringer’s solution and serially diluted (10 to 105). Serial dilutions were passed through 0.45 µm Merck filters (Merck Life Sciences Limited, Dorset, UK). The filters were placed on Tryptone Bile X-Glucuronide chromogenic media (TBX) selective for E. coli. The Petri dishes were placed inverted at 30 °C for 4 h and 44 °C for 21 h. The plates were examined for the growth of blue–green colonies marking the presence of E. coli. A total of 351 strains were isolated from the collected water samples.
4.1.2. Mollusk Samples
Concurrent to the seawater sampling, mollusk samples were gathered in the summer (Jul–Aug 2015; 34.0 °C ± 2.0) and winter seasons (Dec–Feb 2016; 17.2 °C ± 2.0). Shells (n = 55–60) were hand-picked, sorted in autoclaved bags (4 °C), and transported to the College of Science, Kuwait University laboratories for further processing [28]. The mollusc shells were removed to collect the flesh and fluid in a pre-autoclaved beaker (Borosil, Poole, UK). Approximately 20 g was macerated with 0.1% peptone water (Sigma, Plymouth, UK) and serially diluted [28]. The dilutions were passed through the 0.45 µm nitrocellulose membrane filters and placed on TBX-agar for E. coli, as mentioned previously. In total, 247 mollusc strains were isolated.
4.1.3. Antimicrobial Susceptibility Testing
Antibiotic susceptibility assays for all these isolates were performed, and minimum inhibitory concentration (MIC) was recorded and presented in Al-Sarawi et al. [28]. Briefly, the isolates were tested for susceptibility and resistance against a panel of 23 antibiotics as per the guidelines of Clinical and Laboratory Standards Institutes [59]. The micro-dilution method was employed to determine the MIC by inoculating a loopful of isolate into 10 mL of Cation-Adjusted Muller Hinton (CAMBH) broth (incubated on shaker incubator for 24 h at 37 °C). Turbidity was adjusted with 0.5 M MacFarland solution and loaded to custom, dehydrated, 96-well SensititreTM plates (GN2F, Thermo Scientific, Paisley, UK), which were further incubated at 25 °C for 24 h. Strains were classified as resistant, intermediate, or susceptible based on the MIC breakpoints standardized by CEFAS [27].
4.2. Whole-Genome Sequencing (WGS) and Filtering of ARGEs
From the 598 isolates, selected strains (n = 23, Table S5) with interesting AMR phenotypes, i.e., resistant to 2 or more antibiotics Al- Sarawi et al. [28] or susceptible to all, were subjected to WGS. These isolates were also chosen to represent varied spatiotemporal variability (three sites and two seasons). Whole-genome DNA was isolated from an over-nightly activated broth. Approximately 15 mL of Tryptic Soy Broth was seeded with a loopful of E. coli and incubated at 37 °C. The culture tube was centrifuged at 13,000× g to pellet the organisms and subjected to DNA purification as per the standard protocol of Promega Wizard ®Genomic DNA extraction Kit (Promega, Chilworth Southampton, Hampshire, UK) [60]. Fluorometric quantification of DNA was conducted on a Quantus fluorometer (Promega, Chilworth Southampton, Hampshire, UK). Sequencing libraries were prepared following the Nextera XT kit (Illumina, USA). The libraries were purified using AMPure® XP (Beckman Coulter, Brea, CA, USA) magnetic beads. Libraries were normalized and pooled before loading on a MiSeq V3 sequencing cartridge. Sequencing was performed on an Illumina MiSeq platform available at the Centre for Environment Fisheries and Aquaculture Science (CEFAS), Weymouth laboratories through paired-end (2 × 300 bp) chemistry [36]. Post-sequencing, the Illumina adapters were removed by Trimmomatic v 0.36 [61] and a quality check was performed on the web version of FastQC v 0.11.5 [62]. Raw reads were thereafter aligned in Spades v 3.10.1 for prokaryotic assembly [63] and annotated via Prokka v1.11 [64]. The ABRicate tool [65] was used to screen the ‘assembled fasta’ for antibiotic-resistant genes (ARGs), employing the ResFinder v 4.0.0 [66] database (accessed on 2 April 2023). The sequences were also mined for plasmids through the plasmid finder database [67]. The integron finder database v 2.0.2 was used to annotate the complete integron with integron integrase, attC recombination sites, promoters, and nearby gene cassettes [68].
4.3. MLST (Muti-Locus Sequence Typing) and Phylogenetic Analysis
Seven MLST genes, namely, adk-fumC-gyrB-icd-mdh-purA-recA were retrieved from the Pfam archive. The respective ids of the gene sets were PF00406, PF05683, PF00204, PF00180, PF00056, PF00709, and PF00154, respectively [69]. The hmmscan v 3.1, [70] was then used to align the assembled E. coli genomes with the abovementioned MLSTs. The identified MLSTs were aligned in mafft v 7.305 using the default settings [71]. UGENE v 1.98 was employed to remove and concatenate the unaligned sequences from the 5′ and 3′ ends [72]. A maximum likelihood tree analysis was performed on the aligned MLSTs through FastTree v 2.1.8 [73]. A phylogenetic tree was plotted in FigTree v 1.4.2 [74].
4.4. Screening for Class 1 Integrons and Associated Gene Cassettes
A total of 598 E. coli pure cultures isolated from marine waters (n = 351) and marine organisms/mollusks (n = 247) were used for intl1 gene screening [28,75] and associated gene cassettes (GC). DNA was isolated as per the protocol described elsewhere [76]. Primers int1.F/R (F-5′GGGTCAAGGATCTGGATTTCG 3′; R-5′ACATGCGTGTAAATCATCGTCG3′) and HS286 (F-5′ GGGATCCTCSGCTKGARCGAMTTGTTAGVC) HS287 (R-5′GGGATCCGCSGCTKANCTCVRRCGTTAGSC3′) were used to amplify the intl1 and GCs, respectively [33,77]. A PCR mix (25 μL) was prepared by combining 2 μL (4 µM) each of forward and reverse primers, 6.5 μL of PCR grade water, 12.5 µL of buffer (MangoMixTM, Bioline, London, UK), and 2 μL of template DNA. The reaction was run for initial denaturation (94 °C for 60 s) followed by 35 repetitions (denaturation—94 °C, 60 s; annealing—58 °C for 30 s; extension—72 °C for 150 s) and final extension at 72 °C on a Gene Amp PCR system 9700 (Applied Biosystem, Cheshire, UK). PCR products were passed through a 1.5% agarose gel (Fisher, NJ, USA) and visualized on a GelDoc XR gel imaging system (Bio-Rad, Hertfordshire, UK).
4.5. Statistical Analysis
Statistical analysis was performed in SPSS 23.0 (IBM, SPSS, Hampshire, UK). The normality of data was tested through the Shapiro–Wilk test. A t-test was run to check for the no-normality violation and homogeneity assumptions. Data were considered significant at a confidence interval of 99.95% (p < 0.05). Regression tests applying rectilinear models were employed to establish the correlation between intl1-GC resistance. Homogeneity of variance, data normality, and dimensions of linearity were also explored.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/antibiotics12091366/s1. Table S1: Antibiotic resistance genes filtered through ResFinder; Table S2: Plasmids found in selected isolates of E. coli; Table S3: Integrons and associated elements filtered from the integrall database.; Table S4: GPS coordinates and physicochemical parameters of sampling sites; Table S5: Metadata of E. coli isolates used for sequencing.
Author Contributions
H.A.A.-S.: conceptualization, methodology, validation, resources; N.H.: software, data curation, writing—original draft, writing—review and editing, visualization; S.U.: investigation, writing—review and editing; A.N.J.: investigation, methodology; M.A.A.-S.: investigation, methodology; B.P.L.: conceptualization, supervision, project administration. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not Applicable.
Informed Consent Statement
Not Applicable.
Data Availability Statement
Raw Sequences are hosted on the web portal of the National Centre for Biotechnology Information (NCBI) under accession no. PRJNA955288 (SRR24162651 to SRR24162673) https://dataview.ncbi.nlm.nih.gov/object/PRJNA955288?reviewer=gfeu7sag3l3hgq6o8k49nk974 (accessed on 15 June 2023).
Acknowledgments
We are thankful to Plymouth University for providing the facilities for whole-genome sequencing.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Bungau, S.; Tit, D.M.; Behl, T.; Aleya, L.; Zaha, D.C. Aspects of excessive antibiotic consumption and environmental influences correlated with the occurrence of resistance to antimicrobial agents. Curr. Opin. Environ. Sci. Health 2021, 19, 100224. [Google Scholar]
- Kraemer, S.A.; Ramachandran, A.; Perron, G.G. Antibiotic pollution in the environment: From microbial ecology to public policy. Microorganisms 2019, 7, 180. [Google Scholar]
- Thompson, T. The staggering death toll of drug-resistant bacteria. Nature 2022. Online ahead of print. [Google Scholar]
- Marti, E.; Variatza, E.; Balcazar, J.L. The role of aquatic ecosystems as reservoirs of antibiotic resistance. Trends Microbiol. 2014, 22, 36–41. [Google Scholar] [CrossRef]
- Baquero, F.; Alvarez-Ortega, C.; Martinez, J.L. Ecology and evolution of antibiotic resistance. Environ. Microbiol. Rep. 2009, 1, 469–476. [Google Scholar]
- Al-Ghadban, A.; Uddin, S.; Aba, A.; Ali, L.N.; Al-Sharmoukh, D.; Al-Khabbaz, A.; Al-Mutairi, A. Measurement and Assessment of Radionuclide Concentrations in the Coastal Marine Environment; Kuwait Institute for Scientific Research: Kuwait City, Kuwait, 2011; p. 128. [Google Scholar]
- Gevao, B.; Jaward, F.M.; Uddin, S.; Al-Ghadban, A.N. Occurrence and concentrations of polychlorinated dibenzo-p-dioxins (PCDDs) and polychlorinated dibenzofurans (PCDFs) in coastal marine sediments in Kuwait. Mar. Pollut. Bull. 2009, 58, 452–455. [Google Scholar]
- Gevao, B.; Uddin, S.; Dupont, S. Baseline concentrations of pharmaceuticals in Kuwait’s coastal marine environment. Mar. Pollut. Bull. 2021, 173, 113040. [Google Scholar]
- Uddin, S.; Aba, A.; Fowler, S.; Behbehani, M.; Ismaeel, A.; Al-Shammari, H.; Alboloushi, A.; Mietelski, J.; Al-Ghadban, A.; Al-Ghunaim, A. Radioactivity in the Kuwait marine environment—Baseline measurements and review. Mar. Pollut. Bull. 2015, 100, 651–661. [Google Scholar]
- Uddin, S.; Fowler, S.W.; Saeed, T. Microplastic particles in the Persian/Arabian Gulf–a review on sampling and identification. Mar. Pollut. Bull. 2020, 154, 111100. [Google Scholar]
- Uddin, S.; Fowler, S.W.; Habibi, N.; Behbehani, M. Micro-Nano Plastic in the Aquatic Environment: Methodological Problems and Challenges. Animals 2022, 12, 297. [Google Scholar]
- Hernando-Amado, S.; Coque, T.M.; Baquero, F.; Martínez, J.L. Defining and combating antibiotic resistance from One Health and Global Health perspectives. Nat. Microbiol. 2019, 4, 1432–1442. [Google Scholar]
- Chen, J.; McIlroy, S.E.; Archana, A.; Baker, D.M.; Panagiotou, G. A pollution gradient contributes to the taxonomic, functional, and resistome diversity of microbial communities in marine sediments. Microbiome 2019, 7, 1–12. [Google Scholar]
- Amarasiri, M.; Sano, D.; Suzuki, S. Understanding human health risks caused by antibiotic resistant bacteria (ARB) and antibiotic resistance genes (ARG) in water environments: Current knowledge and questions to be answered. Crit. Rev. Environ. Sci. Technol. 2020, 50, 2016–2059. [Google Scholar]
- Uddin, S.; Fowler, S.W.; Uddin, M.F.; Behbehani, M.; Naji, A. A review of microplastic distribution in sediment profiles. Mar. Pollut. Bull. 2021, 163, 111973. [Google Scholar]
- Habibi, N.; Uddin, S.; Bottein, M.Y.D.; Faizuddin, M. Ciguatera in the Indian Ocean with Special Insights on the Arabian Sea and Adjacent Gulf and Seas: A Review. Toxins 2021, 13, 525. [Google Scholar]
- Uddin, S.; Al-Ghadban, A.; Gevao, B.; Al-Shamroukh, D.; Al-Khabbaz, A. Estimation of suspended particulate matter in Gulf using MODIS data. Aquat. Ecosyst. Health Manag. 2012, 15, 41–44. [Google Scholar] [CrossRef]
- Habibi, N.; Uddin, S.; Al-Sarawi, H.; Aldhameer, A.; Shajan, A.; Zakir, F.; Abdul Razzack, N.; Alam, F. Metagenomes from Coastal Sediments of Kuwait: Insights into the Microbiome, Metabolic Functions and Resistome. Microorganisms 2023, 11, 531. [Google Scholar]
- Wright, G.D. Antibiotic resistance in the environment: A link to the clinic? Curr. Opin. Microbiol. 2010, 13, 589–594. [Google Scholar]
- Perry, J.A.; Wright, G.D. The antibiotic resistance “mobilome”: Searching for the link between environment and clinic. Front. Microbiol. 2013, 4, 138. [Google Scholar]
- Von Wintersdorff, C.J.; Penders, J.; Van Niekerk, J.M.; Mills, N.D.; Majumder, S.; Van Alphen, L.B.; Savelkoul, P.H.; Wolffs, P.F. Dissemination of antimicrobial resistance in microbial ecosystems through horizontal gene transfer. Front. Microbiol. 2016, 7, 173. [Google Scholar]
- Perry, J.A.; Wright, G.D. Forces shaping the antibiotic resistome. BioEssays 2014, 36, 1179–1184. [Google Scholar]
- Anonymous. Antimicrobial Resistance: Global Report on Surveillance; World Health Organization: Geneva, Switzerland, 2014. [Google Scholar]
- Devlin, M.; Le Quesne, W.J.; Lyons, B.P. The marine environment of Kuwait—Emerging issues in a rapidly changing environment. Mar. Pollut. Bull. 2015, 100, 593–596. [Google Scholar]
- Le Quesne, W.J.; Baker-Austin, C.; Verner-Jeffreys, D.W.; Al-Sarawi, H.A.; Balkhy, H.H.; Lyons, B.P. Antimicrobial resistance in the Gulf Cooperation Council region: A proposed framework to assess threats, impacts and mitigation measures associated with AMR in the marine and aquatic environment. Environ. Int. 2018, 121, 1003–1010. [Google Scholar]
- Habibi, N.; Uddin, S.; Lyons, B.; Al-Sarawi, H.A.; Behbehani, M.; Shajan, A.; Razzack, N.A.; Zakir, F.; Alam, F. Antibiotic Resistance Genes Associated with Marine Surface Sediments: A Baseline from the Shores of Kuwait. Sustainability 2022, 14, 8029. [Google Scholar]
- Al-Sarawi, H.A.; Jha, A.N.; Al-Sarawi, M.A.; Lyons, B.P. Historic and contemporary contamination in the marine environment of Kuwait: An overview. Mar. Pollut. Bull. 2015, 100, 621–628. [Google Scholar]
- Al-Sarawi, H.A.; Jha, A.N.; Baker-Austin, C.; Al-Sarawi, M.A.; Lyons, B.P. Baseline screening for the presence of antimicrobial resistance in E. coli isolated from Kuwait’s marine environment. Mar. Pollut. Bull. 2018, 129, 893–898. [Google Scholar]
- Al-Sarawi, H.A.; Najem, A.B.; Lyons, B.P.; Uddin, S.; Al-Sarawi, M.A. Antimicrobial Resistance in Escherichia coli Isolated from Marine Sediment Samples from Kuwait Bay. Sustainability 2022, 14, 11325. [Google Scholar] [CrossRef]
- Zhu, L.-X.; Zhang, Z.-W.; Liang, D.; Jiang, D.; Wang, C.; Du, N.; Zhang, Q.; Mitchelson, K.; Cheng, J. Multiplex asymmetric PCR-based oligonucleotide microarray for detection of drug resistance genes containing single mutations in Enterobacteriaceae. Antimicrob. Agents Chemother. 2007, 51, 3707–3713. [Google Scholar]
- Papan, C.; Meyer-Buehn, M.; Laniado, G.; Nicolai, T.; Griese, M.; Huebner, J. Assessment of the multiplex PCR-based assay Unyvero pneumonia application for detection of bacterial pathogens and antibiotic resistance genes in children and neonates. Infection 2018, 46, 189–196. [Google Scholar]
- Burakoff, A.; Brown, K.; Knutsen, J.; Hopewell, C.; Rowe, S.; Bennett, C.; Cronquist, A. Outbreak of fluoroquinolone-resistant Campylobacter jejuni infections associated with raw milk consumption from a herdshare dairy—Colorado, 2016. Morb. Mortal. Wkly. Rep. 2018, 67, 146. [Google Scholar]
- Nemergut, D.; Martin, A.; Schmidt, S. Integron diversity in heavy-metal-contaminated mine tailings and inferences about integron evolution. Appl. Environ. Microbiol. 2004, 70, 1160–1168. [Google Scholar]
- Pournajaf, A.; Ardebili, A.; Goudarzi, L.; Khodabandeh, M.; Narimani, T.; Abbaszadeh, H. PCR-based identification of methicillin–resistant Staphylococcus aureus strains and their antibiotic resistance profiles. Asian Pac. J. Trop. Biomed. 2014, 4, S293–S297. [Google Scholar]
- Khan, M.W.; Habibi, N.; Shaheed, F.; Mustafa, A.S. Draft genome sequences of five clinical strains of Brucella melitensis isolated from patients residing in Kuwait. Genome Announc. 2016, 4, e01144-16. [Google Scholar] [CrossRef]
- Light, E.; Baker-Austin, C.; Card, R.M.; Ryder, D.; Alves, M.T.; Al-Sarawi, H.A.; Abdulla, K.H.; Stahl, H.; Aliya, A.-G.; Al Ghoribi, M. Establishing a marine monitoring programme to assess antibiotic resistance: A case study from the Gulf Cooperation Council (GCC) region. medRxiv 2022, 9, 100268. [Google Scholar]
- Cabello-Yeves, P.J.; Callieri, C.; Picazo, A.; Mehrshad, M.; Haro-Moreno, J.M.; Roda-Garcia, J.J.; Dzhembekova, N.; Slabakova, V.; Slabakova, N.; Moncheva, S. The microbiome of the Black Sea water column analyzed by shotgun and genome centric metagenomics. Environ. Microbiome 2021, 16, 1–15. [Google Scholar]
- Ruppé, E.; Cherkaoui, A.; Charretier, Y.; Girard, M.; Schicklin, S.; Lazarevic, V.; Schrenzel, J. From genotype to antibiotic susceptibility phenotype in the order Enterobacterales: A clinical perspective. Clin. Microbiol. Infect. 2020, 26, 643.e641–643.e647. [Google Scholar]
- Zhang, X.-X.; Zhang, T.; Zhang, M.; Fang, H.H.; Cheng, S.-P. Characterization and quantification of class 1 integrons and associated gene cassettes in sewage treatment plants. Appl. Microbiol. Biotechnol. 2009, 82, 1169–1177. [Google Scholar]
- Grevskott, D.H.; Svanevik, C.S.; Sunde, M.; Wester, A.L.; Lunestad, B.T. Marine bivalve mollusks as possible indicators of multidrug-resistant Escherichia coli and other species of the Enterobacteriaceae family. Front. Microbiol. 2017, 8, 24. [Google Scholar] [CrossRef]
- Beaz-Hidalgo, R.; Balboa, S.; Romalde, J.L.; Figueras, M.J. Diversity and pathogenecity of Vibrio species in cultured bivalve molluscs. Environ. Microbiol. Rep. 2010, 2, 34–43. [Google Scholar]
- Giacometti, F.; Pezzi, A.; Galletti, G.; Tamba, M.; Merialdi, G.; Piva, S.; Serraino, A.; Rubini, S. Antimicrobial resistance patterns in Salmonella enterica subsp. enterica and Escherichia coli isolated from bivalve molluscs and marine environment. Food Control 2021, 121, 107590. [Google Scholar]
- Gevao, B.; Uddin, S.; Krishnan, D.; Rajagopalan, S.; Habibi, N. Antibiotics in Wastewater: Baseline of the Influent and Effluent Streams in Kuwait. Toxics 2022, 10, 174. [Google Scholar] [CrossRef] [PubMed]
- Wright, G.D. The antibiotic resistome: The nexus of chemical and genetic diversity. Nat. Rev. Microbiol. 2007, 5, 175–186. [Google Scholar] [CrossRef] [PubMed]
- Wright, M.S.; Baker-Austin, C.; Lindell, A.H.; Stepanauskas, R.; Stokes, H.W.; McArthur, J.V. Influence of industrial contamination on mobile genetic elements: Class 1 integron abundance and gene cassette structure in aquatic bacterial communities. ISME J. 2008, 2, 417–428. [Google Scholar] [CrossRef]
- Waśko, I.; Kozińska, A.; Kotlarska, E.; Baraniak, A. Clinically Relevant β-Lactam Resistance Genes in Wastewater Treatment Plants. Int. J. Environ. Res. Public Health 2022, 19, 13829. [Google Scholar] [CrossRef]
- Tooke, C.L.; Hinchliffe, P.; Bragginton, E.C.; Colenso, C.K.; Hirvonen, V.H.; Takebayashi, Y.; Spencer, J. β-Lactamases and β-Lactamase Inhibitors in the 21st Century. J. Mol. Biol. 2019, 431, 3472–3500. [Google Scholar] [CrossRef] [PubMed]
- Pongchaikul, P.; Mongkolsuk, P. Comprehensive Analysis of Imipenemase (IMP)-Type Metallo-β-Lactamase: A Global Distribution Threatening Asia. Antibiotics 2022, 11, 236. [Google Scholar] [CrossRef] [PubMed]
- Habibi, N.; Uddin, S.; Behbehani, M.; Kishk, M.; Abdul Razzack, N.; Zakir, F.; Shajan, A. Antibiotic Resistance Genes in Aerosols: Baseline from Kuwait. Int. J. Mol. Sci. 2023, 24, 6756. [Google Scholar] [CrossRef] [PubMed]
- Ryu, S.-H.; Park, S.-G.; Choi, S.-M.; Hwang, Y.-O.; Ham, H.-J.; Kim, S.-U.; Lee, Y.-K.; Kim, M.-S.; Park, G.-Y.; Kim, K.-S. Antimicrobial resistance and resistance genes in Escherichia coli strains isolated from commercial fish and seafood. Int. J. Food Microbiol. 2012, 152, 14–18. [Google Scholar] [CrossRef]
- Koczura, R.; Mokracka, J.; Jabłońska, L.; Gozdecka, E.; Kubek, M.; Kaznowski, A. Antimicrobial resistance of integron-harboring Escherichia coli isolates from clinical samples, wastewater treatment plant and river water. Sci. Total Environ. 2012, 414, 680–685. [Google Scholar] [CrossRef]
- Lyons, B.; Devlin, M.; Hamid, S.A.; Al-Otiabi, A.; Al-Enezi, M.; Massoud, M.; Al-Zaidan, A.; Smith, A.; Morris, S.; Bersuder, P. Microbial water quality and sedimentary faecal sterols as markers of sewage contamination in Kuwait. Mar. Pollut. Bull. 2015, 100, 689–698. [Google Scholar] [CrossRef]
- Lyons, B.; Barber, J.; Rumney, H.; Bolam, T.; Bersuder, P.; Law, R.; Mason, C.; Smith, A.; Morris, S.; Devlin, M. Baseline survey of marine sediments collected from the State of Kuwait: PAHs, PCBs, brominated flame retardants and metal contamination. Mar. Pollut. Bull. 2015, 100, 629–636. [Google Scholar] [CrossRef]
- Gaze, W.H.; Zhang, L.; Abdouslam, N.A.; Hawkey, P.M.; Calvo-Bado, L.; Royle, J.; Brown, H.; Davis, S.; Kay, P.; Boxall, A. Impacts of anthropogenic activity on the ecology of class 1 integrons and integron-associated genes in the environment. ISME J. 2011, 5, 1253–1261. [Google Scholar] [CrossRef]
- Stalder, T.; Barraud, O.; Casellas, M.; Dagot, C.; Ploy, M.-C. Integron involvement in environmental spread of antibiotic resistance. Front. Microbiol. 2012, 3, 119. [Google Scholar] [CrossRef]
- Zhang, H.; Wang, Y.; Liu, P.; Sun, Y.; Dong, X.; Hu, X. Unveiling the occurrence, hosts and mobility potential of antibiotic resistance genes in the deep ocean. Sci. Total Environ. 2022, 816, 151539. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Liu, R.; Liu, H.; Wang, C.; Yin, X.; Zhang, M.; Fang, J.; Zhang, T.; Ma, L. Evidence for Long-Term Anthropogenic Pollution: The Hadal Trench as a Depository and Indicator for Dissemination of Antibiotic Resistance Genes. Environ. Sci. Technol. 2021, 55, 15136–15148. [Google Scholar] [CrossRef] [PubMed]
- Domingues, S.; Da Silva, G.J.; Nielsen, K.M. Global dissemination patterns of common gene cassette arrays in class 1 integrons. Microbiology 2015, 161, 1313–1337. [Google Scholar] [CrossRef] [PubMed]
- Hastey, C.J.; Boyd, H.; Schuetz, A.N.; Anderson, K.; Citron, D.M.; Dzink-Fox, J.; Hackel, M.; Hecht, D.W.; Jacobus, N.V.; Jenkins, S.G. Changes in the antibiotic susceptibility of anaerobic bacteria from 2007–2009 to 2010–2012 based on the CLSI methodology. Anaerobe 2016, 42, 27–30. [Google Scholar] [CrossRef] [PubMed]
- Al Salameen, F.; Habibi, N.; Uddin, S.; Al Mataqi, K.; Kumar, V.; Al Doaij, B.; Al Amad, S.; Al Ali, E.; Shirshikhar, F. Spatio-temporal variations in bacterial and fungal community associated with dust aerosol in Kuwait. PLoS ONE 2020, 15, e0241283. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef]
- Andrews, S. FastQC: A quality control tool for high throughput sequence data. Retrieved May 2010, 17, 2018. [Google Scholar]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef]
- Seemann, T. Prokka: Rapid prokaryotic genome annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef]
- Seemann, T. ABRicate: Mass Screening of Contigs for Antibiotic Resistance Genes; GitHub: San Francisco, CA, USA, 2016. [Google Scholar]
- Bortolaia, V.; Kaas, R.S.; Ruppe, E.; Roberts, M.C.; Schwarz, S.; Cattoir, V.; Philippon, A.; Allesoe, R.L.; Rebelo, A.R.; Florensa, A.F. ResFinder 4.0 for predictions of phenotypes from genotypes. J. Antimicrob. Chemother. 2020, 75, 3491–3500. [Google Scholar] [CrossRef]
- Carattoli, A.; Hasman, H. PlasmidFinder and In Silico pMLST: Identification and Typing of Plasmid Replicons in Whole-Genome Sequencing (WGS). In Horizontal Gene Transfer. Methods in Molecular Biology; de la Cruz, F., Ed.; Humana: New York, NY, USA, 2020; Volume 2075, pp. 285–294. [Google Scholar] [CrossRef]
- Néron, B.; Littner, E.; Haudiquet, M.; Perrin, A.; Cury, J.; Rocha, E.P. IntegronFinder 2.0: Identification and analysis of integrons across bacteria, with a focus on antibiotic resistance in Klebsiella. Microorganisms 2022, 10, 700. [Google Scholar] [CrossRef] [PubMed]
- Finn, R.D.; Tate, J.; Mistry, J.; Coggill, P.C.; Sammut, S.J.; Hotz, H.-R.; Ceric, G.; Forslund, K.; Eddy, S.R.; Sonnhammer, E.L. The Pfam protein families database. Nucleic Acids Res. 2007, 36, D281–D288. [Google Scholar] [CrossRef] [PubMed]
- Finn, R.D.; Clements, J.; Eddy, S.R. HMMER web server: Interactive sequence similarity searching. Nucleic Acids Res. 2011, 39, W29–W37. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Misawa, K.; Kuma, K.i.; Miyata, T. MAFFT: A novel method for rapid multiple sequence alignment based on fast Fourier transform. Nucleic Acids Res. 2002, 30, 3059–3066. [Google Scholar] [CrossRef]
- Okonechnikov, K.; Golosova, O.; Fursov, M.; Team, U. Unipro UGENE: A unified bioinformatics toolkit. Bioinformatics 2012, 28, 1166–1167. [Google Scholar] [CrossRef]
- Price, M.N.; Dehal, P.S.; Arkin, A.P. FastTree 2–approximately maximum-likelihood trees for large alignments. PLoS ONE 2010, 5, e9490. [Google Scholar] [CrossRef]
- FigTree. Molecular, Evolution, Phylogenetic and Epidemiology v1.4.2. Available online: http://tree.bio.ed.ac.uk/software/figtree/ (accessed on 17 May 2017).
- Alsarawi, H.A. Developing an Integrated Strategy for the Assessment of Hazardous Substances in Kuwait’s Marine Environment; University of Plymouth: Plymouth, UK, 2017. [Google Scholar]
- Mustafa, A.S.; Habibi, N.; Osman, A.; Shaheed, F.; Khan, M.W. Species identification and molecular typing of human Brucella isolates from Kuwait. PLoS ONE 2017, 12, e0182111. [Google Scholar] [CrossRef]
- Mazel, D.; Dychinco, B.; Webb, V.A.; Davies, J. Antibiotic resistance in the ECOR collection: Integrons and identification of a novel aad gene. Antimicrob. Agents Chemother. 2000, 44, 1568–1574. [Google Scholar] [CrossRef] [PubMed]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).