
Salmonella enterica Serovar Dublin from Cattle in California from 1993 to 2019: Characterization and Analysis of Antimicrobial Resistance Diversity


Abstract
:1. Introduction
2. Results
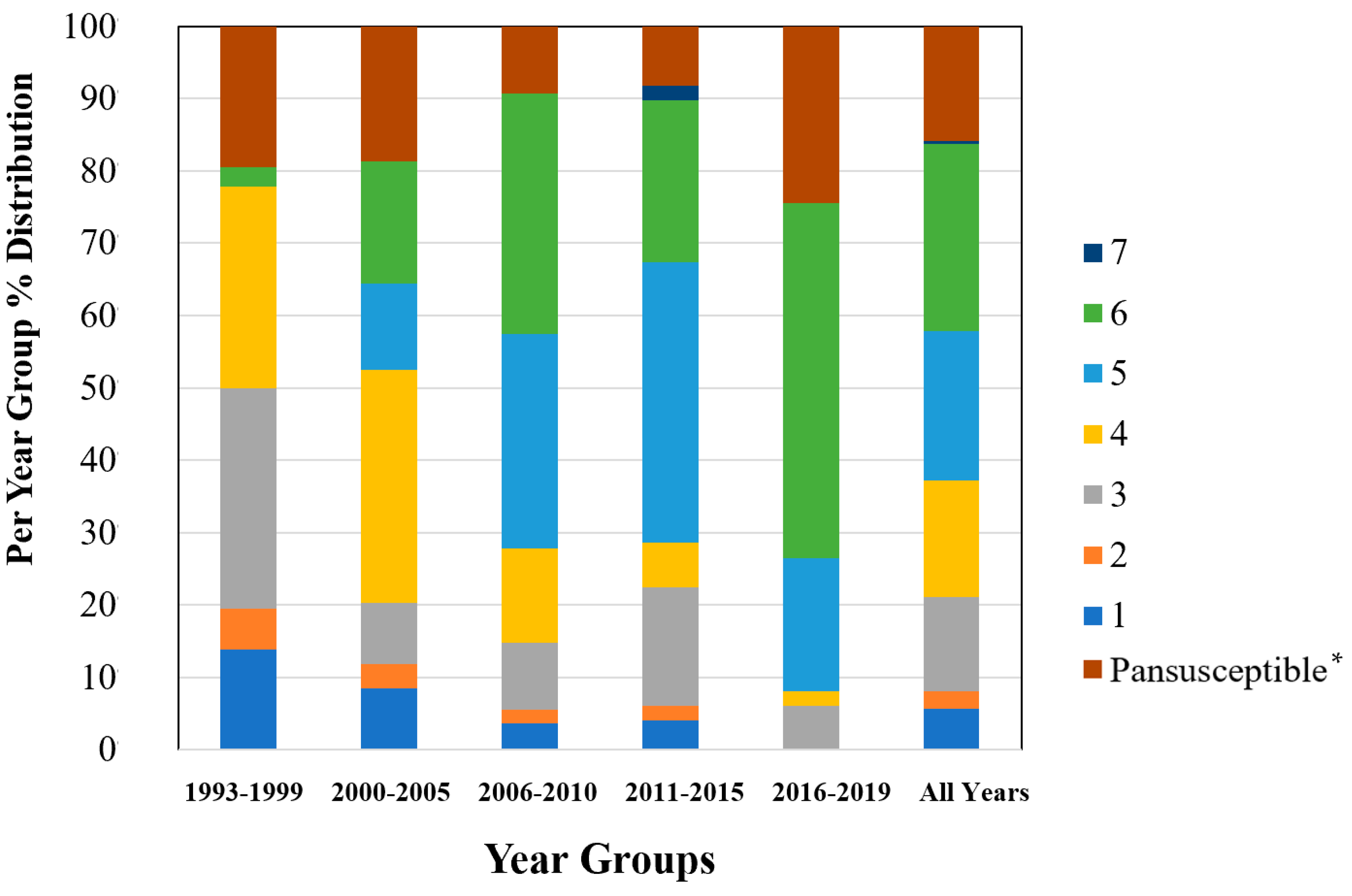
2.1. AMR Profiles Diversity
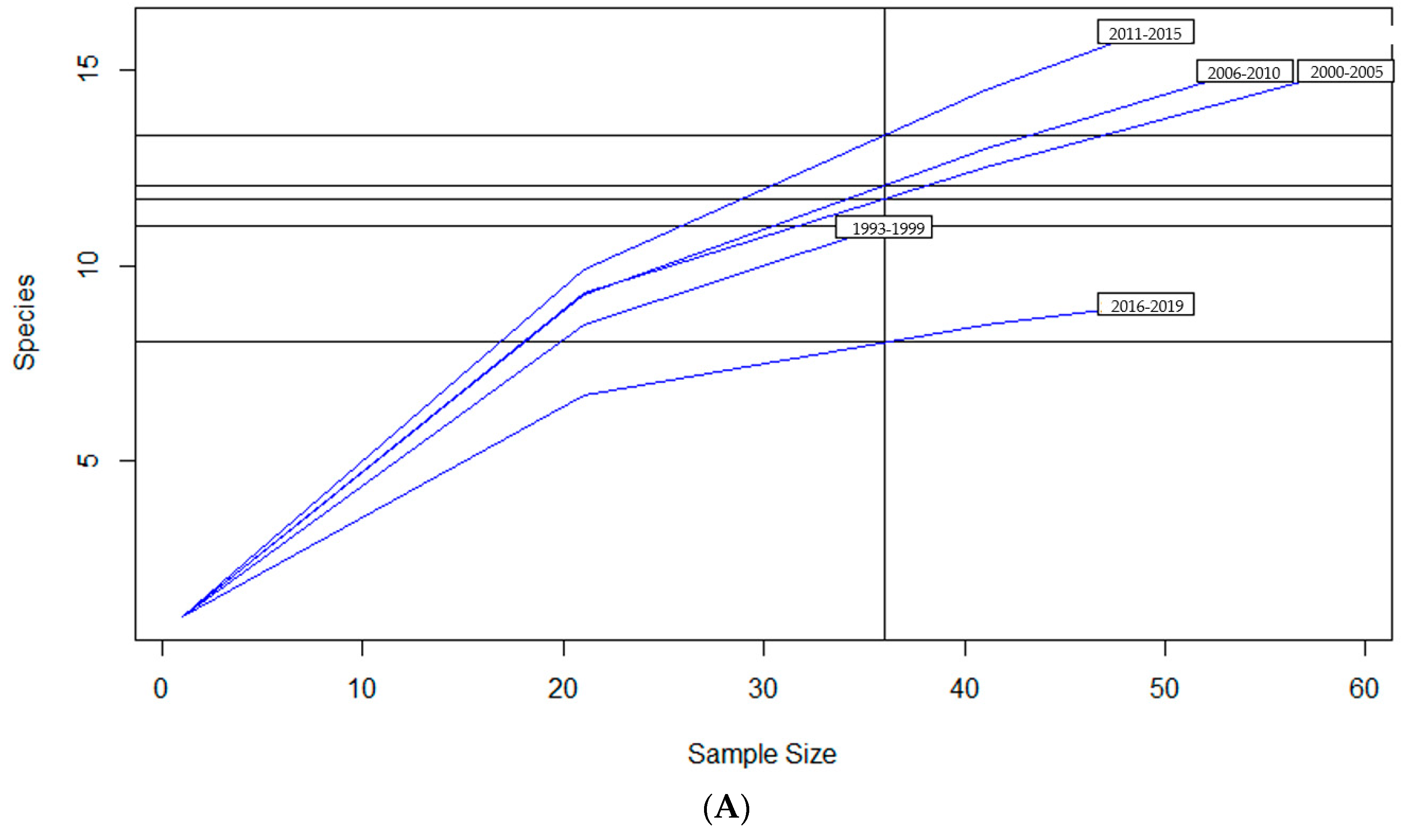
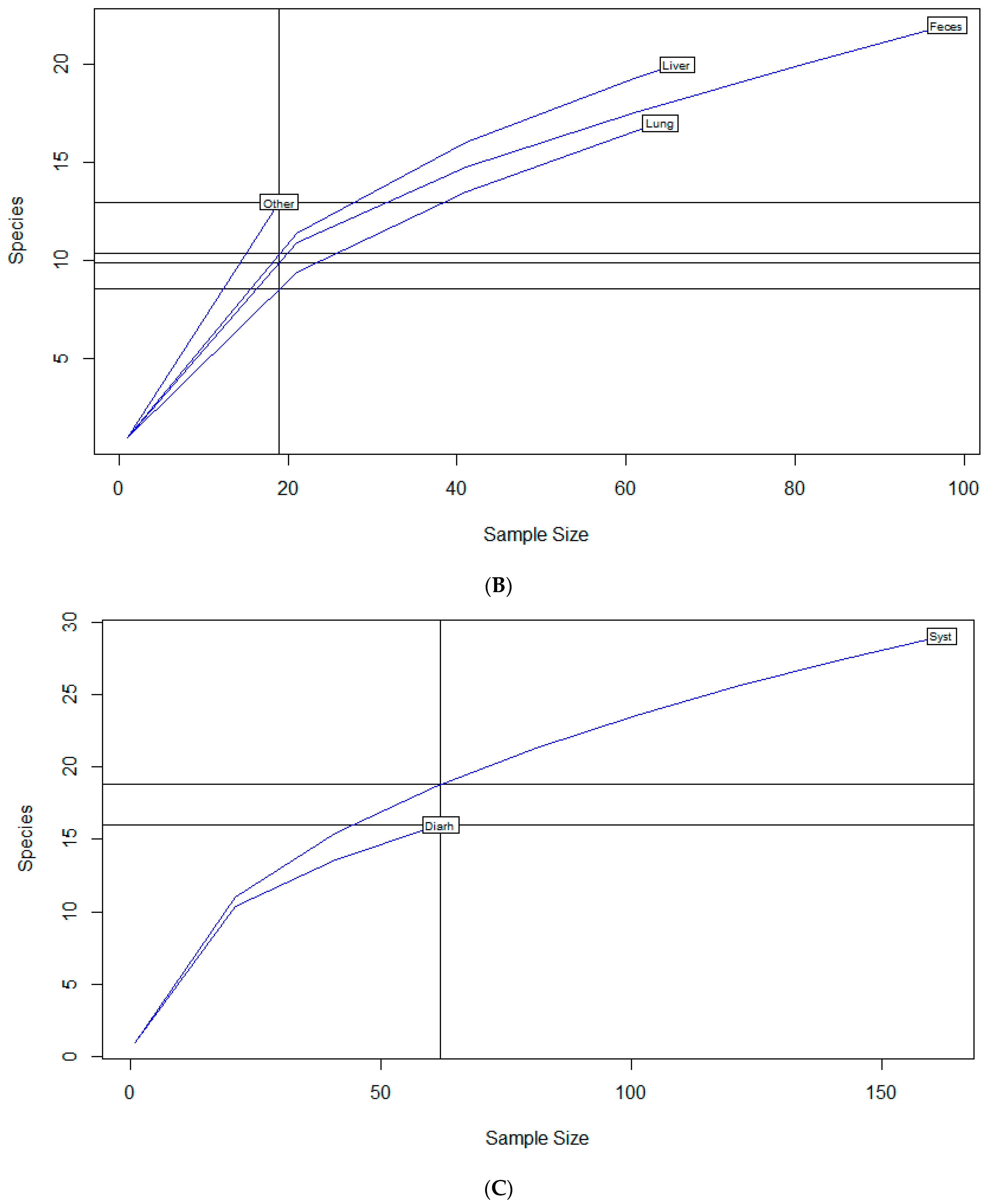
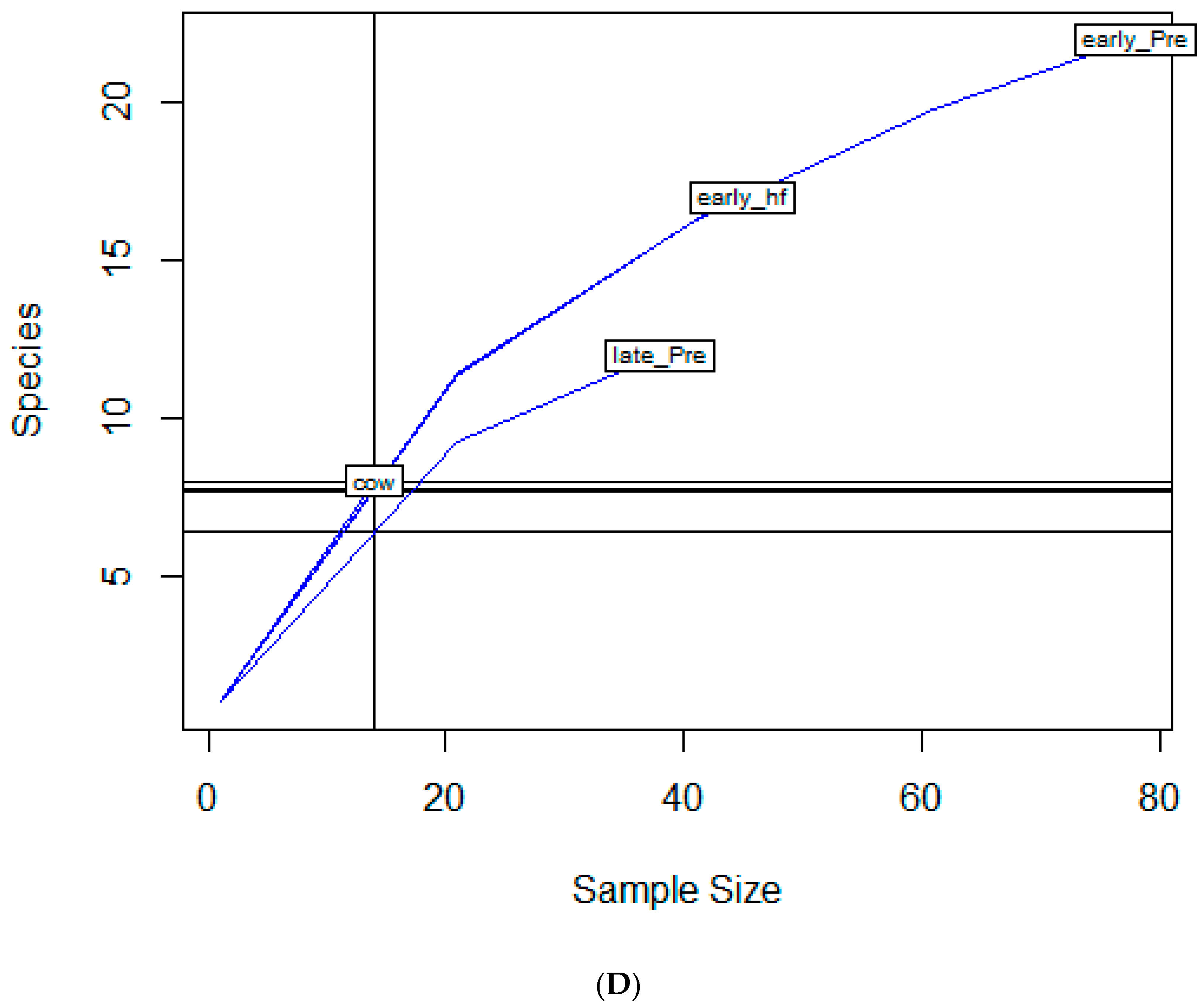
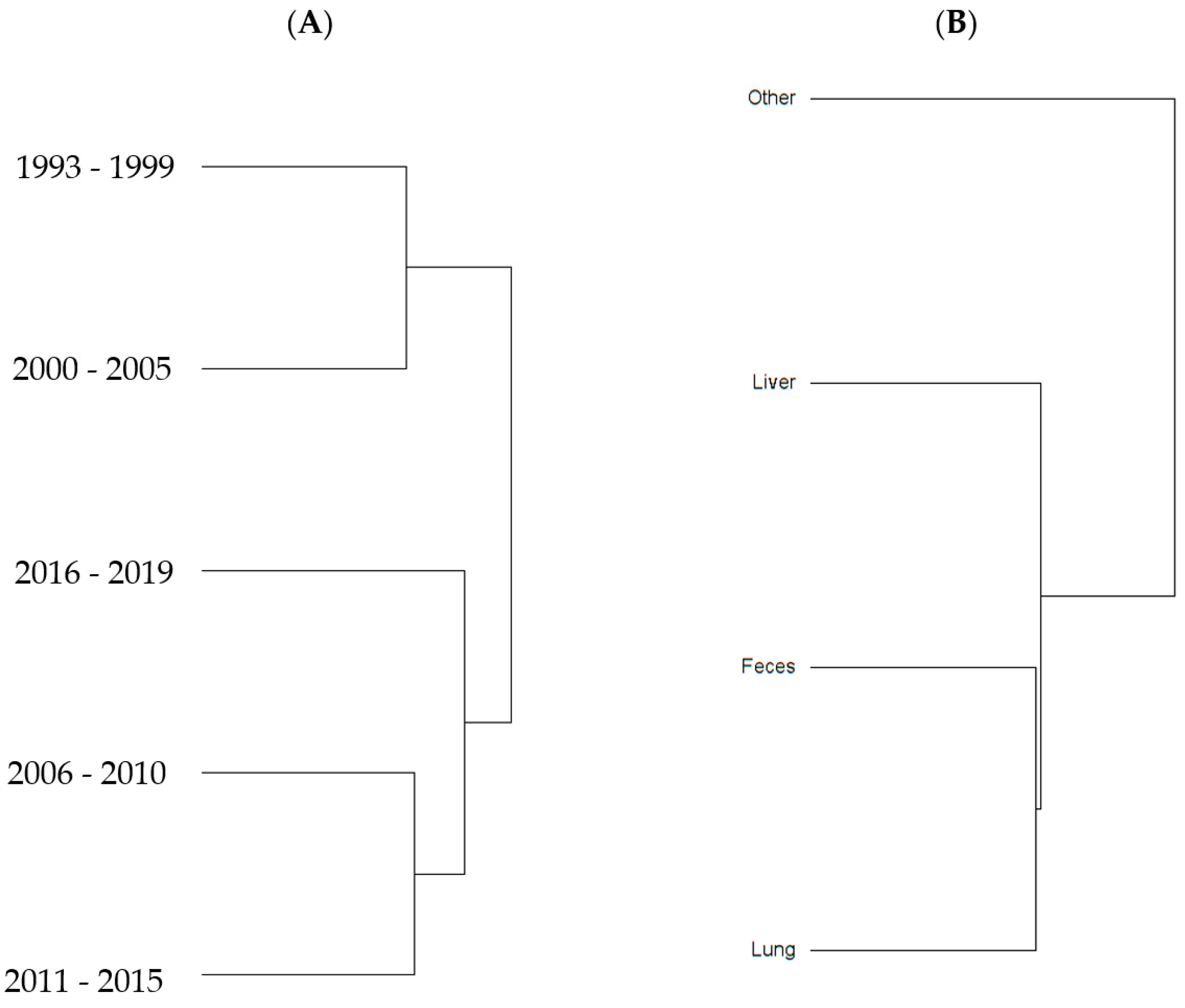
2.2. Diversity Descriptive Evaluation of AMR Profiles
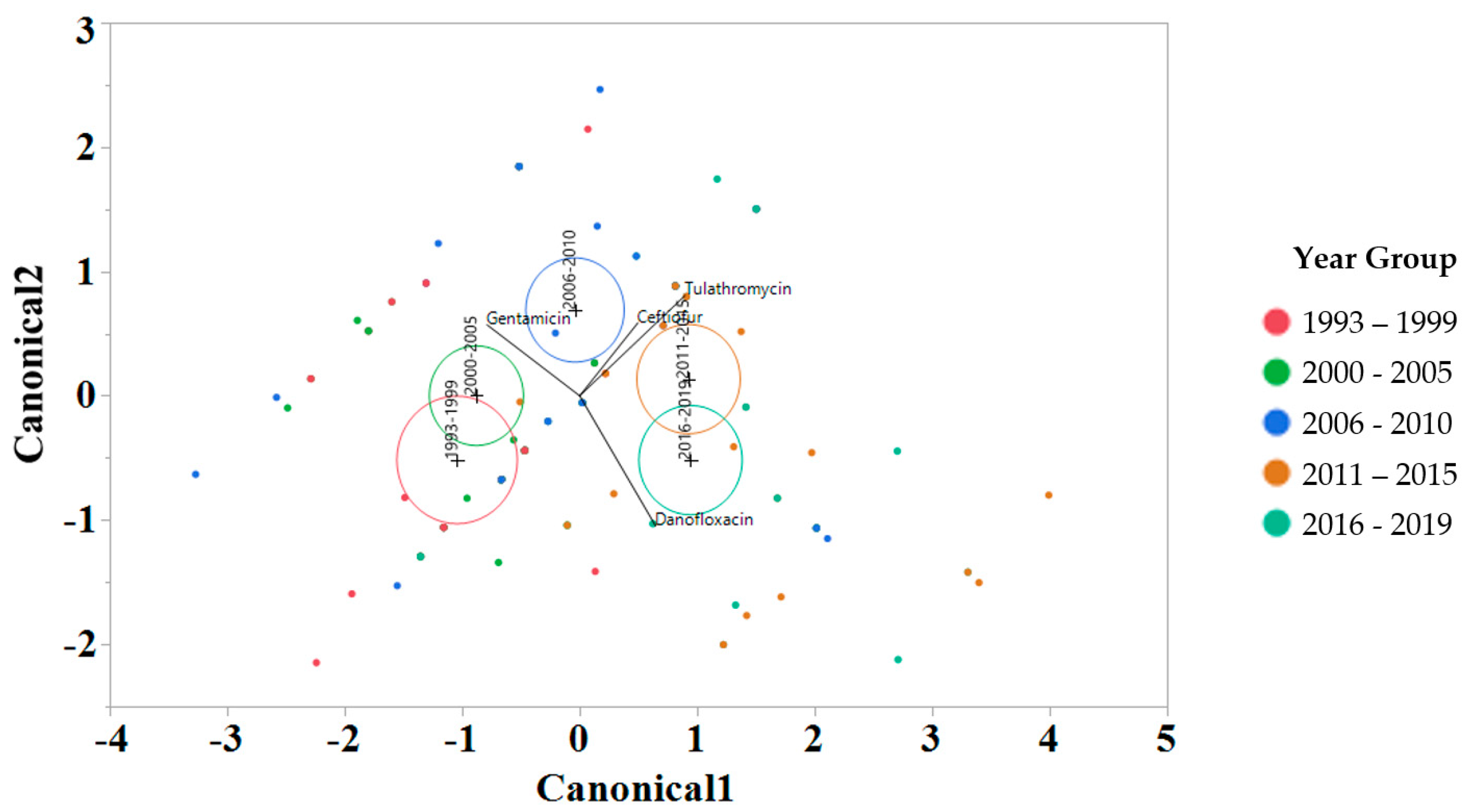
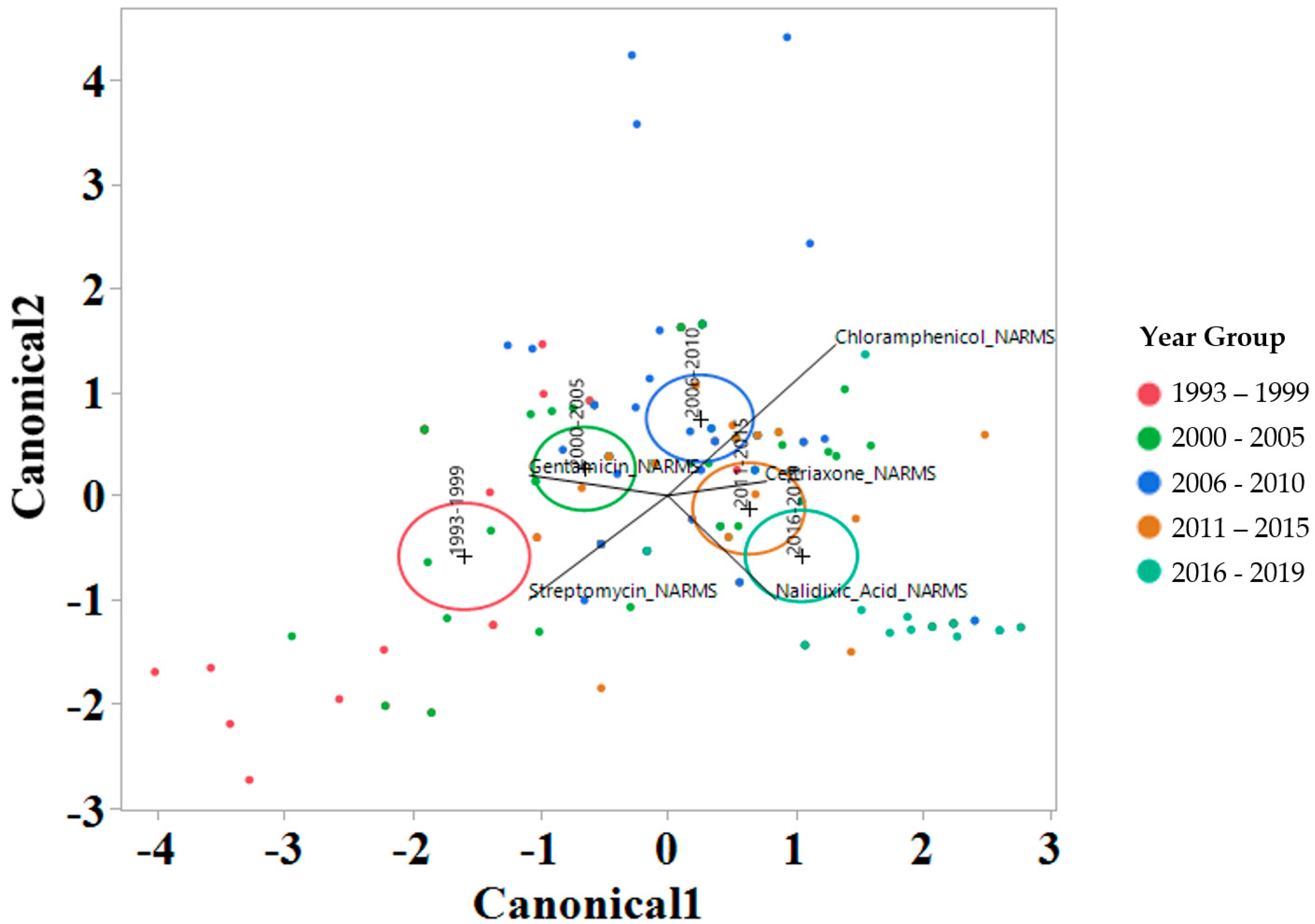
2.3. Linear Discriminant Analysis of MIC for Various Factors
3. Discussion
4. Materials and Methods
4.1. Sample Source, Salmonella Identification, and Serotyping
4.2. Antimicrobial Susceptibility Testing
4.3. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Davidson, K.E.; Byrne, B.A.; Pires AF, A.; Magdesian, K.G.; Pereira, R.V. Antimicrobial resistance trends in fecal Salmonella isolates from northern California dairy cattle admitted to a veterinary teaching hospital, 2002–2016. PLoS ONE 2018, 13, e0199928. [Google Scholar] [CrossRef] [PubMed]
- Holschbach, C.L.; Peek, S.F. Salmonella in Dairy Cattle. Vet. Clin. N. Am. Food Anim. Pract. 2018, 34, 133–154. [Google Scholar] [CrossRef] [PubMed]
- Harvey, R.R.; Friedman, C.R.; Crim, S.M.; Judd, M.; Barrett, K.A.; Tolar, B.; Folster, J.P.; Griffin, P.M.; Brown, A.C. Epidemiology of Salmonella enterica Serotype Dublin Infections among Humans, United States, 1968–2013. Emerg. Infect. Dis. 2017, 23, 1493–1501. [Google Scholar] [CrossRef] [PubMed]
- CDC. Outbreak of Salmonella Infections Linked to Ground Beef; CDC: Atlanta, GA, USA, 2019.
- Turnidge, J.; Paterson, D.L. Setting and revising antibacterial susceptibility breakpoints. Clin. Microbiol. Rev. 2007, 20, 391–408. [Google Scholar] [CrossRef] [PubMed]
- Kowalska-Krochmal, B.; Dudek-Wicher, R. The Minimum Inhibitory Concentration of Antibiotics: Methods, Interpretation, Clinical Relevance. Pathogens 2021, 10, 165. [Google Scholar] [CrossRef] [PubMed]
- Srednik, M.E.; Lantz, K.; Hicks, J.A.; Morningstar-Shaw, B.R.; Mackie, T.A.; Schlater, L.K. Antimicrobial resistance and genomic characterization of Salmonella Dublin isolates in cattle from the United States. PLoS ONE 2021, 16, e0249617. [Google Scholar] [CrossRef] [PubMed]
- NARMS. National Antimicrobial Resistance Monitoring System (NARMS): 2019 Integrated Report Summary; NARMS: Athens, GA, USA, 2019. [Google Scholar]
- CDC. Salmonella—Information for Healthcare Professionals and Laboratories. 2023. Available online: https://www.cdc.gov/salmonella/general/technical.html#:~:text=Healthcare%20providers%20might%20need%20to,azithromycin%2C%20and%20third%20generation%20cephalosporins (accessed on 13 December 2023).
- Constable, P.D. Antimicrobial use in the treatment of calf diarrhea. J. Vet. Intern. Med. Am. Coll. Vet. Intern. Med. 2004, 18, 8–17. [Google Scholar] [CrossRef]
- Fritz, H.M.; Pereira, R.V.; Toohey-Kurth, K.; Marshall, E.; Tucker, J.; Clothier, K.A. Salmonella enterica Serovar Dublin from Cattle in California from 1993–2019: Antimicrobial Resistance Trends of Clinical Relevance. Antibiotics 2022, 11, 1110. [Google Scholar] [CrossRef] [PubMed]
- CLSI. Performance Standards for Antimicrobial Disk and Dilution Susceptibility Tests for Bacteria Isolated from Animals. VET01S; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2015. [Google Scholar]
- CLSI. Performance Standards for Antimicrobial Disk and Dilution Susceptibility Tests for Bacteria Isolated from Animals; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2018. [Google Scholar]
- USDA. Dairy. Cattle Management Practices in the United States, 2014. Fort Collins, CO.; USDA: Washington, DC, USA, 2016.
- Zhang, H.; John, R.; Peng, Z.; Yuan, J.; Chu, C.; Du, G.; Zhou, S. The relationship between species richness and evenness in plant communities along a successional gradient: A study from sub-alpine meadows of the Eastern Qinghai-Tibetan Plateau, China. PLoS ONE 2012, 7, e49024. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Wang, C.; O’Connor, A. A hierarchical Bayesian latent class mixture model with censorship for detection of linear temporal changes in antibiotic resistance. PLoS ONE 2020, 15, e0220427. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| AMR Profile | Nº Drug Classes * | 1993–1999 | 2000–2005 | 2006–2010 | 2011–2015 | 2016–2019 | All Years |
|---|---|---|---|---|---|---|---|
| Count (n) | 36 | 59 | 54 | 49 | 49 | 247 | |
| Percent Distribution (by Column) | |||||||
| PANSUSCEPTIBLE ** | 0 | 19.4 | 18.6 | 9.3 | 8.2 | 24.5 | 15.8 |
| AmpStrFoxAmcChlXnlCroTet | 5 | 0.0 | 6.8 | 24.1 | 34.7 | 16.3 | 17.0 |
| AmpStrFoxAmcChlXnlCroTetGen | 5 | 2.8 | 15.3 | 25.9 | 4.1 | 0.0 | 10.5 |
| AmpStrFoxAmcChlXnlCroTetNal | 6 | 0.0 | 0.0 | 5.6 | 10.2 | 34.7 | 10.1 |
| AmpStrChlTet | 4 | 13.9 | 13.6 | 9.3 | 0.0 | 0.0 | 7.3 |
| AmpStrChlTetGen | 4 | 13.9 | 18.6 | 0.0 | 0.0 | 0.0 | 6.5 |
| StrChlTet | 3 | 0.0 | 1.7 | 7.4 | 14.3 | 6.1 | 6.1 |
| AmpStrTet | 3 | 25.0 | 5.1 | 0.0 | 0.0 | 0.0 | 4.9 |
| Str | 1 | 11.1 | 8.5 | 1.9 | 4.1 | 0.0 | 4.9 |
| StrChlNalTet | 4 | 0.0 | 0.0 | 0.0 | 6.1 | 2.0 | 1.6 |
| AmpStrAmcChlXnlCroNalTet | 6 | 0.0 | 0.0 | 0.0 | 0.0 | 8.2 | 1.6 |
| StrChlTetGen | 3 | 2.8 | 1.7 | 1.9 | 0.0 | 0.0 | 1.2 |
| StrFox | 2 | 0.0 | 1.7 | 1.9 | 2.0 | 0.0 | 1.2 |
| AmpStrFoxSxtAmcChlXnlCroTet | 6 | 0.0 | 0.0 | 0.0 | 2.0 | 4.1 | 1.2 |
| AmpStr | 2 | 2.8 | 1.7 | 0.0 | 0.0 | 0.0 | 0.8 |
| AmpStrFoxAmcXnlCroTet | 3 | 2.8 | 0.0 | 0.0 | 2.0 | 0.0 | 0.8 |
| AmpStrAmcChlXnlCroTet | 5 | 0.0 | 1.7 | 1.9 | 0.0 | 0.0 | 0.8 |
| AmpFoxAmcChlXnlCroTetGen | 5 | 0.0 | 0.0 | 3.7 | 0.0 | 0.0 | 0.8 |
| StrFoxSxtChlNalCipTet | 6 | 0.0 | 0.0 | 0.0 | 2.0 | 2.0 | 0.8 |
| AmpStrChlNalTetGen | 5 | 0.0 | 0.0 | 0.0 | 2.0 | 2.0 | 0.8 |
| StrTet | 2 | 2.8 | 0.0 | 0.0 | 0.0 | 0.0 | 0.4 |
| StrGen | 1 | 2.8 | 0.0 | 0.0 | 0.0 | 0.0 | 0.4 |
| AmpStrSxtAmcChlXnlCroTetGen | 6 | 0.0 | 1.7 | 0.0 | 0.0 | 0.0 | 0.4 |
| AmpStrFoxChlTetGen | 5 | 0.0 | 1.7 | 0.0 | 0.0 | 0.0 | 0.4 |
| AmpChlNalTetGen | 5 | 0.0 | 1.7 | 0.0 | 0.0 | 0.0 | 0.4 |
| AmpStrSxtAmcChlXnlCroTet | 6 | 0.0 | 0.0 | 1.9 | 0.0 | 0.0 | 0.4 |
| AmpFoxAmcXnlCroTetGen | 4 | 0.0 | 0.0 | 1.9 | 0.0 | 0.0 | 0.4 |
| AmpFoxAmcChlXnlCroGen | 4 | 0.0 | 0.0 | 1.9 | 0.0 | 0.0 | 0.4 |
| Amp | 1 | 0.0 | 0.0 | 1.9 | 0.0 | 0.0 | 0.4 |
| AmpStrFoxSxtAmcChlXnlCroNalTet | 7 | 0.0 | 0.0 | 0.0 | 2.0 | 0.0 | 0.4 |
| AmpStrFoxAmcChlXnlCroNalTetGen | 6 | 0.0 | 0.0 | 0.0 | 2.0 | 0.0 | 0.4 |
| AmpStrFoxAmcChlXnlCroNalCipTet | 6 | 0.0 | 0.0 | 0.0 | 2.0 | 0.0 | 0.4 |
| AmpFoxAmcChlXnlCroNalTetGen | 6 | 0.0 | 0.0 | 0.0 | 2.0 | 0.0 | 0.4 |
| Variables | Richness 1 | Diversity 2 | Evenness 3 |
|---|---|---|---|
| Year Group | |||
| 1993–1999 | 11 | 2 | 0.85 |
| 2000–2005 | 15 | 2.3 | 0.84 |
| 2006–2010 | 15 | 2.2 | 0.81 |
| 2011–2015 | 16 | 2.2 | 0.8 |
| 2016–2019 | 9 | 1.7 | 0.79 |
| Age Group | |||
| Early Pre-weaned | 22 | 2.61 | 0.84 |
| Late Pre-weaned | 12 | 2.19 | 0.88 |
| Early Heifer | 17 | 2.51 | 0.88 |
| Cow | 8 | 1.9 | 0.91 |
| Source of Isolate | |||
| Feces | 22 | 2.6 | 0.84 |
| Liver | 20 | 2.6 | 0.87 |
| Lung | 17 | 2.3 | 0.8 |
| Other | 13 | 2.5 | 0.96 |
| Clinical Signs | |||
| Diarrhea | 16 | 2.4 | 0.88 |
| Systemic | 29 | 2.7 | 0.8 |
| California Region | |||
| Central | 23 | 2.7 | 0.85 |
| South or North | 25 | 2.6 | 0.8 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pereira, R.V.; Fritz, H.M.; Toohey-Kurth, K.; Clothier, K.A. Salmonella enterica Serovar Dublin from Cattle in California from 1993 to 2019: Characterization and Analysis of Antimicrobial Resistance Diversity. Antibiotics 2024, 13, 22. https://doi.org/10.3390/antibiotics13010022
Pereira RV, Fritz HM, Toohey-Kurth K, Clothier KA. Salmonella enterica Serovar Dublin from Cattle in California from 1993 to 2019: Characterization and Analysis of Antimicrobial Resistance Diversity. Antibiotics. 2024; 13(1):22. https://doi.org/10.3390/antibiotics13010022
Chicago/Turabian StylePereira, Richard V., Heather M. Fritz, Kathy Toohey-Kurth, and Kristin A. Clothier. 2024. "Salmonella enterica Serovar Dublin from Cattle in California from 1993 to 2019: Characterization and Analysis of Antimicrobial Resistance Diversity" Antibiotics 13, no. 1: 22. https://doi.org/10.3390/antibiotics13010022
APA StylePereira, R. V., Fritz, H. M., Toohey-Kurth, K., & Clothier, K. A. (2024). Salmonella enterica Serovar Dublin from Cattle in California from 1993 to 2019: Characterization and Analysis of Antimicrobial Resistance Diversity. Antibiotics, 13(1), 22. https://doi.org/10.3390/antibiotics13010022